
Abstract
Artemisinins, as a class of bioactive molecules, are mainly derived from the extracts of Artemisia annua L. and are the mainstay for malaria treatment, including severe malaria, uncomplicated malaria and multi-drug resistant malaria. They are well-known for their good tolerability, safety and rapid onset of action. Their efficacy is not only limited to malaria but also extends to a variety of human diseases such as cancer, tuberculosis, viral diseases (e.g. Human cytomegalovirus), immune diseases and parasitic infections like schistosomiasis. Being a cheap and safe drug class, which saves millions of lives at risk from malaria around the globe, can also have significant potential in oncology as they have shown anti-cancer properties in both cell lines and animal models. Active derivatives (e.g. artesunate, artemether and arteether etc.) have also been synthesized which can be used for oral, rectal, intramuscular and intravenous administration. A comprehensive update on the non-malarial use of artemisinins and/or their derivatives and artemisinin-based drug development beyond anti-malarial is discussed in this review. With the collaborative efforts in the clinical pharmacology of artemisinins and novel synthesis of artemisinin analogues, it is likely that artemisinin-based drugs will become an important armamentarium impeding a number of diseases beyond malaria.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Artemisinins are isolated from a Chinese medicinal annual herb Artemisia annua L commonly called as sweet wormwood (Asteraceae). Chemically it is a 1,2,-trioxane bearing an endoperoxide bridge. Professor You-you Tu and her research team isolated and discovered artemisinins and was awarded the Nobel Prize (2015) in Physiology and Medicine for this discovery (Tu 2011). Besides antimalarial properties, it has significant effects against different diseases such as cancer (Cheng et al. 2018), tuberculosis (Miller et al. 2011), autoimmune diseases (e.g. asthma, rheumatoid arthritis (RA), systemic lupus erythematosus (SLE), multiple sclerosis (MS), (Hou and Huang 2016) schistosomiasis (del Villar et al. 2012) and viral diseases (Efferth et al. 2008) and even against some plant tumors (Efferth 2017). Being a major global concern, malaria (a protozoan disease) with 214 million annual cases is the cause of 430,000 annual deaths most of whom are children younger than 5 years (Organization WH 2016). Plasmodium sp. particularly Plasmodium falciparum which is the causative agent of this fatal disease and proliferates in female Anopheles mosquitoes and can kill off the patients, usually in a matter of hours (Cox 2010). Some other species of the genus like P. ovale, P. vivax, P. knowlesi and P. malariae also cause malarial infections in humans (Wilson et al. 2011). Although, continuous efforts have been made since the 1940s to stop the outspread of the disease which was succeeded in North America, Europe, Canada and Russia (Alonso et al. 2011) and parts of Latin America and Asia (Carter and Mendis 2003) but not in Africa, particularly Sub-Saharan parts, where about 90% of malaria deaths take place and more than 80% of the annual patients are found with malaria (Lalloo et al. 2016). Prevalence has fallen up to some extent because of improved control, reasonable increases by funding agencies and enthusiasm for eradication and elimination (Sinka et al. 2012). Biting habits, density, longevity and efficiency of the vector are among the major determinants of the transmission intensity of malaria. With various measures such as insecticide-treated nets and vector control, research and development (R&D) have also initiated for new vaccine and drugs. While the most favoured therapy is the artemisinins combination therapy (ACT) (Yakasai et al. 2015) that is partially or completely based on artemisinins produced by the natural source (Artemisia annua) (Ikram et al. 2017). It is also possible to produce artemisinins in Physcomitrella patens and Nicotiana benthamiana heterologously (Han et al. 2016; Wang et al. 2016). The vaccine for Plasmodium is known as PfSPZ and can be obtained from P. patens and N. benthamiana (Rosales-Mendoza et al. 2017). Quinine as a first antimalarial drug was isolated from Cinchona bark and malaria has been mostly treated with drugs based on quinoline such as quinine, mefloquine and primaquine, chloroquine and antifolates. Unfortunately, many of the P. falciparum strains have developed resistance to chloroquine, halofantrine and mefloquine (Nondo et al. 2017). Interestingly, the bioactivity of artemisinins and/or their derivatives is not only restricted to malaria but have also shown activities against cancer both in vitro and in vivo. A number of case reports, usually, that of human cancer patients reveled that artemisinins have anti-cancer activities (e.g. breast cancer, abdominal ascites and hepatocellular carcinoma, lung cancer, brain cancer, prostate carcinoma and laryngeal squamous cell carcinoma etc.) (Konstat-Korzenny et al. 2018). Likewise malaria, tuberculosis is also lethal infectious and the most widespread disease around the globe (Hurtley et al. 2010). Presently, a huge number of the world population is infected with Mycobacterium tuberculosis which causes more than 2 million deaths around the globe per year (Bloom 1994). Although artemisinins itself are inactive against TB, but synergistically with a mycobacterial-specific siderophore analogue induces selective and significant activity against TB including Mycobacterium tuberculosis which is multi and highly drug-resistance strains (Zheng et al. 2017). Additionally, artemisinins and their semisynthetic derivatives’ (e.g. Artesunate) bioactivity is even more broader and also includes the inhibition of certain viruses, particularly human cytomegalovirus and some members of the family Herpesviridae (Epstein-Barr virus and herpes simplex virus type 1) bovine viral diarrhea virus and hepatitis B and C virus (Hahn et al. 2018). Among the most common parasitic and prevalent diseases, schistosomiasis tends to be continue. In mid-2003 approximately 779 million people were at risk of this disease while the infected people were counted as 207 million (Ross et al. 2017). Trematode worm of the genus Schistosoma is the causative agent of schistosomiasis while 3 main species i.e. S. japonicum, S. haematobium and S. mansoni parasitize humans. Furthermore, some other species such as S. intercalatum and S. malayensis have also been found in Central Africa and Southeast Asia respectively (Pérez et al. 2012). In a large scale, although praziquantel is one of the most valuable and tested control strategy for schistosomiasis, however, the unavailability of therapeutic efficacy, particularly at early stages, is the limiting factor of paraziquantel and could be one of the main reasons for treatment failures. Hence, artemisinins and their derivatives emerged as a family of effective compounds and the best option with schistomicide activity (Crellen et al. 2016). Besides of artemisinins excellent clinical anti-cancer, anti-malarial and anti-tuberculosis effects, various studies also demonstrate that it has the potent anti-inflammatory and immune regulatory functions. Additionally, it has been reported that artesunate reduces the phagocytic index (in vivo) as well as peritoneal macrophages (Li et al. 2013a). Due to their eminent suppressive effects on both adaptive and innate immune cells, a number of experimental autoimmune models have been tested treating with artemisinin family drugs (Hou and Huang 2016). This review will mainly focus on artemisinins, their derivatives (especially synthetic) and artemisinin family drugs as a remedy for various diseases including cancer, malaria, tuberculosis and schistosomiasis.
Brief History and Discovery of Artemisinins
The discovery of artemisinins is somewhat linked with Vietnam War (1967) where Ho Chi Minh (the leader of North Vietnam) was at a war against the United States and South Vietnam. During the war, a majority of the Vietnamese soldiers who were trying to troop down the Ho trail suffered from malaria resistant to chloroquine. The leader asked Chinese Premier En-lai Zhou for assistance to provide some possible cure for his army against malaria (Ali et al. 2017). Meanwhile, malaria also emerged as a cause of hundreds to thousands of deaths in the Southern provinces of China. A meeting was scheduled under the instructions of Premier En-lai Zhou and Chinese Chairman Ze-dong Mao on May 23rd 1967 in China’s capital city Beijing to consider the problem and prevent malaria parasites. In this regard, a program called Project 523 was set up involving more than 500 scientists from 60 different institutes. This project sets 2 goals: 1; short-term goal was to produce some anti-malarial drugs that could instantly be used by the soldiers in the battlefield, 2; the long-term goal of the concerned project was to develop new anti-malarial drugs by searching and screening recipes, synthetic chemicals and practices of traditional Chinese medicines (TCM) (Liu 2017). Initially, You-You Tu (a member of the Project 523) and her team visited Hainan province of China where they studied patients infected with malaria (Miller and Su 2011). In 1969, she had an idea of screening of different Chinese herbs. Over 2000 traditional Chinese recipes were screened out by her team and made a total of 380 herbal extracts, which were first tested on mice infected with malaria (Krungkrai and Krungkrai 2016). A compound from Artemisia annua was more effective against “intermittent fevers” a hallmark of malaria. The preparation techniques were described in a 1600 years old book entitled “The Handbook of Prescriptions for Emergency Treatments” written by Hong Ge in 340. The book also recommends an amount of A. annua mixed thoroughly with water (usually 2 L), extract the juice and drink it all (Tu 2011). She extracted the compound using a low-temperature ether method and it was found completely effective in tests conducted on animals (mice and monkeys). She obtained artemisinins as a pure substance in 1972 and saved millions of lives. She was awarded the Lasker Award in clinical medicine and Nobel Prize in Medicine and Physiology in 2011 and 2015 respectively (Meskill 2017).
Physiochemical Properties of Artemisinins and their Derivatives
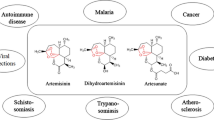
Chemical structure of the compound was determined by different approaches such as mass spectroscopy, spectrophotometry, X-ray crystallography and polyarithmetic analysis (Tu 2016). The chemical structure of artemisinins and their derivatives provided a base for further improvement of the drugs. Some chemical structures of artemisinins and their derivatives are given in Fig. 1. Physically, it is a white and needle-like crystals (without aromatic conjugate system) on 151–153 °C, insoluble in water but soluble in ether, acetone, petroleum ether, ethanol and alkali solution. The molecular formula is confirmed from fundamental analysis and mass spectra as C15H22O5 (Guo 2016). Furthermore, due to their unique chemical structure and activity artemisinins can be kept without any decomposition at room temperature for a long time (Nakase et al. 2008). Qualitative analysis of the compound has shown positive color reactions by the oxidation of NaCI or NaOH while NaOH titration consumes one equivalent. It makes one equivalent of triphenylphosphine oxide by reaction with triphenyphosphine which indicate the presence of the oxidative group in its molecules. Several derivatives such as artesunate and artelinic acid (water soluble), artemether and arteether (lipid soluble) and dihydroartemisinin as an active metabolites are available to treat various diseases including malaria, cancer and tuberculosis (Liu 2017). In addition, a number of other sesquiterpenes are identified from ethereal extraction, including arteannuin A, B and C, and amorphane. Chemically, artemisinin is a 15-carbon sesquiterpene lactone with an endoperoxide (-C-O-C-O-) linkage and a molecular weight of 282 g/ mmol. This intact endoperoxide bridge is of immense importance and has a role in the formation of free radicles (Nakase et al. 2008). As artemisinin derivatives have no endoperoxide bridge and hence are devoid of anti-malarial activity. Furthermore, Fe (III) does not cause the reductive session of the endoperoxide as Fe (II) do. A series of semi-synthetic dihydroartemisinin (fluorinated) derivatives were patented by Centre National de la Recherche Scientifique in 2008 and named as monomeric artemisinin derivatives. These derivatives were exemplified by compounds 7–13 and metabolic stability was also imparted introducing a fluoro-alkyl substituent to C-10 position (Begue et al. 2008). In addition, by incorporating another functional group, mainly an ionizable, to such analogues at either C-16 or C-10 and water-soluble compounds were synthesized. Another group known as diametric artemisinins have been modified from parent artemisinin. In this group, 2 derivatives of monomeric artemisinin are coupled through different linkers connecting with C-10 positions. It is claimed that this group is more stable hydrolytically than other clinically used artemisinin such as artesunate (Blazquez et al. 2013).

Molecular Structures of artemisinin and its derivatives, (Reproduced from Ref. (Li et al. 2016), an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY))
Artemisinin Biosynthesis in planta (A. annua)
Although artemisinin biosynthesis has been investigated since long ago but the detailed regulation and biosynthesis is still not understood. Moreover, in-depth regulatory studies have been facilitated by the fact that biosynthesis as a whole is located in the glandular trichomes of Artemisia annua (Fig. 2) (Olofsson et al. 2011). A total of 3 molecules i.e. one dimethylallyl diphosphate (DMAAP) and two of isopentyl diphosphate (IPP), derived from the general terpenoid biosynthesis, are condensed by farnesyl diphosphate synthase (FPS) into farnesyl diphosphate (FPP, C15 sesquiterpenoid precursor) (Wen and Yu 2011). Interestingly, high artemisinin production has been obtained by the overexpression of FPS in A. annua (Banyai et al. 2010) which not only confirm the role FPS but also the availability of the substrate in artemisinin regulation and biosynthesis similar to that of other sesquiterpene lactones (Simonsen et al. 2013). By cyclization and carbocation formation, FPP is converted to amorpha-4, 11-diene by the enzyme amorpha-4, 11-diene synthase (ADS) (Wen and Yu 2011). In two further oxidization steps, in which, first amorpha-4, 11-diene is hydroxylated into artemisinic alcohol and then by the action of amorphadiene monooxygenase (an enzyme of cytochrome P450) oxidized to atremisinic aldehyde (Wang et al. 2011b). The activity of amorphadiene monooxygenase has confirmed via knock-out of the gene (endogenously) in the plant with no production of downstream products of amorphadiene (Czechowski et al. 2016). Likewise, another enzyme called alcohol dehydrogenase, which has latter been discovered, and has affinity towards artemisinin alcohol and further oxidizes it to aldehyde. The strong expression and specificity of alcohol dehydrogenase in glandular trichomes of A. annua confirms that this enzyme is responsible for artemisinic alcohol oxidation to artemisinic aldehyde (He et al. 2017). By the action of artemisinic aldehyde Δ 111 (13) reductase, artemisinic aldehyde reduced to another compound called dihydroartemisinic aldehyde and eventually oxidized by aldehyde dehydrogenase (an enzyme expressed mainly in the trichomes) to dihyroartemisinic acid (Liu et al. 2016). Aldehyde dehydrogenase not only catalyzing the oxidation of dihydroartemisinic aldehyde to the acid but also catalyzes the oxidation (a reaction catalyzed in yeast by amorphadiene monooxygenase) of artemisinic aldehyde to artemisinic acid (Teoh et al. 2009). In the second last step, an enzyme dihyroartemisinic aldehyde reductase changes dihydroartemisinic aldehyde to a “dead end” substance, dihydroartemisinic alcohol, which mainly affects artemisinin production (Kayser et al. 2011). The last step is a non-enzymatic and spontaneous (light-induced) reaction converting artemisinic acid to arteannuin B and dihydroartemisinic acid to artemisinin (Czechowski et al. 2016).

Artemisinin biosynthesis pathway occurs in the glandular trichomes of Artemisia annua L. The pathway intermediates are defined as FPP, farnesyl diphosphate; AD, amorpha-4,11-diene; AAOH, artemisinic alcohol; AAA, artemisinic aldehyde; AA, artemisinic acid; DHAAA, dihydroartemisinic aldehyde; DHAA, dihydroartemisinic acid. Reproduced from Ref. (Ikram and Simonsen 2017), an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY))
Pharmacological Activities/Effects of Artemisinins and/or their Derivatives
Artemisinin as an Effective Drug to Cure Malaria
Malaria is a parasite infection of red blood cells and caused by a unicellular protozoan parasite plasmodium mainly transmitted by anopheles mosquitoes. Being an endemic disease in several countries (approx 108) of Asia, South America and Africa etc. malaria is the cause of high mortality especially in children less than 5 years and the economic loss is huge in regions endemic with malaria (Stebbins et al. 2018). Although morbidity and mortality have decreased up to some extent but still it kills approximately 2000 people per day. Among strategies, two main factors account for these decreases: 1; insecticide-treated bed nets, 2; artemisinin combination treatments, as they are known for their good tolerability, rapid onset of action and safety. In addition to rapid tests, a valuable addition to microscopy for malaria diagnosis (White et al. 2013), effective treatment are also considered as the mainstay for malaria control (Shayo et al. 2015). An expert panel of the World Health Organization (2001) recommended the use of ACT for malaria treatment (Table 1) (Mandara et al. 2018). In large trials, parenteral artesunate (an artemisinin derivative) reduced severe malaria mortality by 22·5% in Africa and 34·7% in Asia compared with quinine, whereas adjunctive interventions have been uniformly unsuccessful (White et al. 2014). By 2006, ACT had become the most recommended treatment for malaria globally (Maxmen 2016; Wang et al. 2017a). Artemether-lumefantrine (Coartem®; Novartis) as a first ACT was validated by the US Food and Drug Administration (FDA) in April 2009 (Jiao et al. 2017). The most used combinition includes artesunate-sulfadoxine-pyrimethamine, pyronaridine-artesunate, arterolane-piperaquine, artemisinin-napthoquine, artesunate-amodiaquine, artemether-lumefantrine, DHA-piperaquine and artesunate-mefloquine (Premji 2009). To increase the efficacy and further ensure substantial treatment outcomes of ACT, caregiver and patient adherence, measurement methods (if they involved), associated factors and treatment guidelines is of immense importance (McCoy et al. 2017). According to WHO report (2009), ACT as the first-line remedy for malaria had adopted by less than 20 countries (Laxminarayan et al. 2006). Furthermore, with the cooperation of a number of donors including President’s Malaria Initiative (PMI) and Global Fund to Fight AIDS, Tuberculosis and Malaria (GFATM), the number of countries that have established ACT has been further increased (Aregawi et al. 2009). In 2010, ACT was adopted by 84 countries, among them, 60 countries initiate to provide ACT free of cost (for all ages) in the public sector while 8 providing subsidized ACT via the Affordable Medicines Facility–malaria (AMFm) in the private sector (Committee WMPA and Secretariat 2012). A number of research and review articles, clinical trials and case reports have been evaluated the anti-malarial activities of artemisinins and their derivatives (Guo 2016; Wang et al. 2015).
Anti-Cancer Properties of Artemisinin
In the past two decades, besides from artemisinin’s antipaludic activity, studies have evaluated the potential of both artemisinins and their various derivatives having inhibitory effects on the growth and particularly the proliferation of tumour cells (Chen et al. 2017; Jana et al. 2017). This specificity is because of certain characteristics of tumour cells, such that, susceptibility to reactive oxygen species (ROS), elevated the concentration of transferrin and iron and increased metabolism (Leto et al. 2016). High potency and specificity have also been shown by artemisinin tagged to certain transferrin through carbohydrate chain against cancer cells. In this regard, the conjugation is of significant importance, enables artemisinin delivery into cancer cells (Wong et al. 2017). Artemisinins and their derivatives inhibit cancer cell proliferation, yet with much lower efficiencies than their roles in killing malaria parasites (Zhang et al. 2016). In addition, due to its minimal toxicity and adverse effects, various case reports and clinical trials (conducted on humans and animals) have shown that artemisinin drug derivatives have promising in vitro and in vivo activity against certain types of cancer and used as antineoplastic drugs (Li 2012; Wang et al. 2017b). Intriguingly, artemisinin destroys only cancer cells by a mechanism called inducing apoptosis and is totally safe to normal cells (Wei et al. 2017). Dihydroartemisinin (an analogue of artemisinin discovered and developed in 1986) selectively killed cancer cells e.g. Molt-4 lymphoblastoid cells in vitro by co-incubation with holo-transferrin, while the same treatment was essentially non-toxic to normal human lymphocytes (Li et al. 2018). Furthermore, dihydroartemisinin and holotransferrin, as a drug combination, was 100 times (LC50s of Molt-4 and normal lymphocytes were 2.6 μM and 230 μM, respectively) more effective compared with normal lymphocytes on Molt-4 cells (Kumari et al. 2017). Studies have been conducted to assess the in vivo anti-cancer bioactivity of artemisinin e.g. Moore et al. reported that the growth and proliferation of fibrosarcoma tumours in the rat was inhibited by oral administration of dihydroartemisinin along with ferrous sulphate (von Hagens et al. 2017). Artemisinin can induce a number of molecular pathways that may bring about apoptosis and necroptosis (Nakase et al. 2009). For instance, Hooft van Huijsduijnen concluded that artemisinin, dihydroartemisinin and artemisone induced apoptosis via the intrinsic pathway in the presence of caspase-3 and caspase-9 (Van Huijsduijnen et al. 2013). Another researcher, Tilaoui also showed that apoptosis is induced in vitro in murine mastocytoma cells (Tilaoui et al. 2014) while Zhang et al. exhibited that apoptosis is induced in human gastric cell lines and human esophageal cancer cells by artesunate (Liu et al. 2015). In a study conducted by National Cancer Institute with a number of cancer cell lines (to analyze their response in vitro to artesunate) concluded that cancer cells including ovary, kidney, melanoma cells, colon, prostate, central nervous system and breast showed susceptibility to the compound (Bai et al. 2018).
Artemisinin Potential as Anti-Tubercular Agent
Tuberculosis (TB) is among the most infectious diseases caused by a multidrug-resistant strains Mycobacterium tuberculosis while drugs such as rifampicin, pyrazinamide, isoniazid and ethambutol were thought to be the backbone of the current first-line treatment regimen (Organization WH 2011). This pathogen, which was responsible for the most deaths in 2014–2015, can persist for many years without causing any disease symptoms in the host (Zheng et al. 2017). After M tuberculosis emergence as resistant-strains to first and second-line drugs (Skrahina et al. 2011) e.g. viomycin, D-cycloserine and cephalosporin, and further restricted access to second-line drugs for the proper treatment, it’s time for an urgent action (Falzon et al. 2013). Furthermore, long course treatment is among the basic challenges of the present TB therapy. In this regard, drugs with shortening the course of therapy can revolutionize TB control (Leistikow et al. 2010). The spread and development of extensively drug-resistant (XRD) and multi-drug resistant (MDR) strains of M tuberculosis have accelerated research activities around the globe (Rojas Vargas et al. 2016). Consequentially, the pipeline has expanded for potential and new drugs (Makarov et al. 2009), but no fully effective drug against TB has been marketed (Harper 2007). Besides, potent anti-malarial activity, artemisinin (as a conjugate of mycobactin-artemisinin) has potential and profound activity against TB. Although artemisinin itself is inactive against TB, but synergistically with an analogue induces selective and significant activity against TB pathogen (Wencewicz and Miller 2017). Notably, a number of studies have been shown that when artemisinin as a mycobactin-artemisinin conjugate is delivered into M tuberculosis, it causes the burst of reactive oxygen through Fenton reactions and kills M tuberculosis. In addition, whole-cell and physiochemical studies indicated that the iron complex of the conjugation reduces from ferric-to-ferrous which initiates radical chemistry, particularly Fenton-type, with artemisinin component (Liu et al. 2018). The anti-M tuberculosis activity of artesunate was shown in an experiment by Won Hyung Choi via different selective anti-M tuberculosis various assays such as Ogawa slant medium assay, MGIT 960 assay and Resazurin Microtiter (REMA) assay while artemisinin effect was found low as compared to artesunate. Additionally, with a single dose for 21 days, artesunate showed persistent effects and anti-M tuberculosis activity as they inhibit pathogen growth and proliferation in vitro. Artesunate also revealed activity against TB in vivo for four weeks (daily dose of 3.5 mg/kg) with no induce toxicity or side effects (Choi 2017b).
Artemisinin-Based Drugs against Schistosomiasis
In spite of continuing control efforts, schistosomiasis is still the major health problem of tropical and subtropical countries. The causative agent for this infectious disease is Schistosoma, a parasitic trematode worms, retains mainly in the mesenteric portal system of the host. Besides negative effects on pregnancy outcome, child development and agricultural production (Adenowo et al. 2015) this debilitating and chronic disease is a cause of labour loss and emerge as a big threat to the economy (Ross et al. 2017). In 2008, the number of people treated for schistosomiasis was 17.5 million globally while 11.7 million out of total belongs to sub-Saharan Africa. Although, some countries including Tunisia and Japan has been eradicated it successfully, whereas, Morocco and some countries of Caribbean Island have already made a good progress on management and control of schistosomiasis (Utzinger et al. 2009). China, Egypt and Brazil initiates for its elimination whilst a number of countries, particularly the sub-Saharan region, are still facing problems with schistosomiasis (Jones et al. 2018). According to WHO the disease accounts for more than 40% of tropical disease. Several drugs such as praziquantel, metrifonate and oxamniquine etc. have been used for the treatment of the disease, while among them praziquantel becomes the first-line drug. However, in the recent past, the resistance of the pathogen to praziquantel has come into concern, that may further necessitate struggle for some substitutes (Tambo et al. 2014). Although, praziquantel is more active against schistosomes in the adult stage but with high rates of re-infection and lack of therapeutic efficacy (against early stage) are the main limitations of praziquantel (Vale et al. 2017). Thus, being versatile pharmacological tools in nature, artemisinin derivatives like artesunate and artemether have a potential against schistosomiasis which was described for the first time in the1980s in China. In 1996, artemisinin derivatives were approved for the prevention of schistosomiasis by Chinese Ministry of Health (Doenhoff et al. 2008). Furthermore, they are active against a number of schistosome species such as S. mansonai, S. japonicum and S. haematobium by targeting schistosomulum (Liu et al. 2017). Artesunate-sulfamethoxypyrazine plus pyrimethamine, artesunate-sulfadoxine-pyrimethamine and artesunate-amodiaquine have been tested on children infected with S. haematobium (Shuhua et al. 2002).
Anti-Viral Activities of Artemisinin
In addition to its antimalarial, antiangiotensin, anti-inflammatory, antibacterial, and anticancer activity artemisinins and their synthetic derivatives especially artesunate has impressive and relatively broad antiviral activity against hepatitis B (HBV), human cytomegalovirus (HCMV) and Bovine Viral Diarrhea Virus (BVDV) (Morere et al. 2015). The remarkable anti-cancer activity of artemisinin compounds have already been confirmed as they demonstrated the potential antiviral efficacy by applying a concept called chemical hybridization (Fröhlich et al. 2017a, b). It has been reported that artesunate has potential inhibitory effects against herpes viruses like human herpes viruses 6A (HHC-6a), herpes simplex virus 1 (HSV-1) and Epstein-Barr virus. In neonates, patients with transplant recipients and AIDS, human CMV is the main cause of infection (Schreiber et al. 2009). The so far available antiviral drugs such as cidofovir, foscarnet and ganciclovir (mainstream anti-CMV) prevent the elongation of viral DNA by targeting activity of viral DNA polymerase but they have a number of side effects including the emergence of drug resistance and bone marrow suppression (Schreiber et al. 2009). With a profile of high safety and tolerability, the intensive use of artesunate has been demonstrated in malaria patients (Zhang et al. 2018). Fortunately, due to its multi-functionality, the value of the compound is not only restricted to malaria but has also been suggested as a substitute to already available antiviral drugs, particularly, in patients with failing therapy (Kar Han Lau et al. 2011). In addition, it can be used orally, intravenously, intramuscularly and also through the rectal route (Morris et al. 2011). Hence, as the best option, artesunate demonstrate its anti-CMV effects by inhibiting the transactivation of DNA-binding factors Sp1 and NF-κB (Kadambari et al. 2017). Furthermore, artesunate has also inhibitory effects against HBV replication by suppressing surface antigen secretion of HBV, as a result, decrease HBV-DNA levels in vitro (Qi et al. 2013) while in vivo it undergoes a rapid conversion by cellular and plasmatic esterases into dihydroartemisinin (an active metabolite) (Fröhlich et al. 2017a, b). Additionally, pharmacological interest has been increased in the recent past in artemisinin in order to treat viral diseases due to severe limitations of the available antiviral therapy (Sayce et al. 2016). Artesusnate activity is not limited to laboratory strains but it has a prominent effect against clinical isolate mutants with a strong resistance against various conventional antiviral drugs (Durantel et al. 2004). Different sensitivities were shown by herpesviruses analyzed by Kaptein SJ et al. to artemisinin and artesunate; as artemisinin was found with poor and no activity against HCMV and human herpes virus 6A respectively while artesunate was found with strong antiherpsviral potency (Kaptein et al. 2006).
Artemisinin Drugs for the Treatment of Autoimmune Diseases
Autoimmune diseases, a family of about 80 illnesses, mainly share a common pathogenesis in which the body attack (also called immune-mediated attack) on its own cells, tissues and organs. They have highly variable manifestations and can affect any site in the body and are most common in industrialized societies (Schwartz et al. 2016). Diseases of this family often disabling and most of the times leading to complete or partial loss of organ function. Being a threat to public health globally, the prevalence of autoimmune diseases is the common problem of both developing as well as developed countries (Bellone 2010). Since cures for most of the autoimmune diseases are not available and usually patients, particularly women in childbearing years, face debilitating illness with costly treatment (Pagan et al. 2018). Beyond antimalarial agent, artemisinins and their related derivatives have immunoregulatory effects on immune cells such as splenocytes, neutrophils, T-cells, B-cells and macrophages (Yi et al. 2017). Artemisinin-based combination therapies may depress neutrophils as they kill different microbes. Furthermore, artemisinin with semi-synthetic derivatives may also interfere with the most functional competencies of neutrophils (Wang et al. 2008). Additionally, artemisinin can reduce the secretion level of tumor necrosis factor (TNF) in vivo, and have been shown a strong inhibitory effect towards TNF-α in vitro release from macrophage. They have been found to modulate host immune functions mediated by macrophage by interrupting transcriptional signaling pathways in macrophages (Loop and Pahl 2003). The effect of artemisinin is not only restricted to drop-off in cytokines release from macrophage but it also downregulate nitric oxide production, which is important for macrophage-mediated immune reactions (Wang et al. 2009). Artesunate, dihydroartemisinin, artemether and SM905 (a new derivative) have shown protective effects against rheumatoid arthritis (Li et al. 2013b). Among these, artesunate was found resistant to cartilage and bone diseases and development of tissue edema in experimental models, mediated by the suppression of a number of pro-inflammatory cytokines like granulocyte macrophage colony stimulating factor (GM-CSF), TNF- α, interleukin (IL-6), IL-17 α, IL-8 and IL-1β through inhibition of the phosphoinositide 3-kinase (P13K/Akt), mitogen-activated protein kinase and NF- κB signaling pathways (Li et al. 2013b). A series of new artemisinin derivatives (SM735, SM905, SM933 and SM934) with immunosuppressive functions against T cell activation in vitro has been synthesized in the recent past (Ho et al. 2014).
Concluding Remarks and Future Recommendations
Being a class of bioactive molecules, artemisinins and their derivatives have a number of biological activities beyond anti-malarial (severe malaria, uncomplicated malaria and multi-drug resistant malaria), that includes anti-schistosomal, anti-cancer, anti-viral, anti-tuberculosis and anti-immune diseases (Table 1). In addition, they are also becoming established as anti-parasitic, anti-fungal, anti-allergic and anti-protozoal agents. The potential of artemisinin lies in broader anti-disease applications, especially in addressing a tough challenge posed by certain advanced cancers for which more expensive but less effective treatments are available. Furthermore, questions related to safety and long-term use, dosing regimens and particularly interactions, which may be positive or negative, with already available therapies and toxicities (which might be associated to the treatment of different tumours) should be answered by clinical studies on urgent bases to consider drugs like artesunate for oncological indications. Also, the next landmark in research regarding artemisinin will be to further analyze the potential efficacy of novel analogues in various disease models beyond malaria which includes immune disorders and infectious diseases. Although, there is extensive preclinical data (both in vitro and in vivo) in the literatures which supports a number of therapeutic applications for artemisinin, particularly artesunate, in human diseases. However, apart from a well-established safety background for artemisinin, clinical studies about artesunate related non-malaria-diseases are still very limited. With the collaborative efforts in the clinical pharmacology of artemisinin and novel synthesis of artemisinin analogues, it seems that artemisinin based drugs will become an important armamentarium impeding a number of diseases beyond malaria.
References
Adenowo AF, Oyinloye BE, Ogunyinka BI, Kappo AP (2015) Impact of human schistosomiasis in sub-Saharan. Africa Brazilian Journal of Infectious Diseases 19:196–205
Ali Z, Mishra N, Baldi A (2016) Development and characterization of arteether-loaded nanostructured lipid carriers for the treatment of malaria. Artificial cells, nanomedicine, and biotechnology 44:545–549
Ali M, Abbasi BH, Ahmad N, Khan H, Ali GS (2017) Strategies to enhance biologically active-secondary metabolites in cell cultures of Artemisia–current trends. Crit Rev Biotechnol 37:833–851
Alonso PL, Brown G, Arevalo-Herrera M, Binka F, Chitnis C, Collins F, Doumbo OK, Greenwood B, Hall BF, Levine MM, Mendis K, Newman RD, Plowe CV, Rodríguez MH, Sinden R, Slutsker L, Tanner M (2011) A research agenda to underpin malaria eradication. PLoS Med 8:e1000406
Aregawi M, Cibulskis RE, Otten M, Williams R (2009) World malaria report 2009. World Health Organization
Bai Z, Guo X-H, Tang C, Yue S-T, Shi L, Qiang B (2018) Effects of Artesunate on the expressions of insulin-like growth Factor-1, Osteopontin and C-Telopeptides of type II collagen in a rat model of. Osteoarthritis Pharmacology 101:1–8
Banyai W, Kirdmanee C, Mii M, Supaibulwatana K (2010) Overexpression of farnesyl pyrophosphate synthase (FPS) gene affected artemisinin content and growth of Artemisia annua L plant cell. Tissue and Organ Culture (PCTOC) 103:255–265
Barger-Kamate B, Forman M, Sangare CO, Haidara ASA, Maiga H, Vaidya D, Djimde A, Arav-Boger R (2016) Effect of artemether-lumefantrine (Coartem) on cytomegalovirus urine viral load during and following treatment for malaria in children. J Clin Virol 77:40–45
Bassat Q (2011) The use of artemether-lumefantrine for the treatment of uncomplicated Plasmodium vivax malaria. PLoS Negl Trop Dis 5:e1325
Begue J-P, Bonnet-Delpon D, Crousse B, Ourevitch M, Chorki F, Grellepois F, Magueur G (2008) Artemisinine derivatives, and uses thereof for treating malaria. Google Patents
Bellone M (2010) Autoimmune Disease: Pathogenesis eLS
Blazquez AG, Fernandez-Dolon M, Sanchez-Vicente L, Maestre AD, Gomez-San Miguel AB, Alvarez M, Serrano MA, Jansen H, Efferth T, Marin JJG, Romero MR (2013) Novel artemisinin derivatives with potential usefulness against liver/colon cancer and viral hepatitis. Bioorg Med Chem 21:4432–4441
Bloom BR (1994) Tuberculosis: pathogenesis, protection, and control. ASM press
Carter R, Mendis KN (2003) Evolutionary and historical aspects of the burden of Malaria. Clin Microbiol Rev 16:173
Chen J, Chen X, Wang F, Gao H, Hu W (2015) Dihydroartemisinin suppresses glioma proliferation and invasion via inhibition of the ADAM17 pathway. Neurol Sci 36:435–440
Chen X, Wong Y, Lim T, Lim W, Lin Q, Wang J, Hua Z (2017) Artesunate activates the intrinsic apoptosis of HCT116 cells through the suppression of fatty acid synthesis and the NF-κB pathway. Molecules 22:1272
Cheng C et al (2018) Induction of autophagy and autophagy-dependent apoptosis in diffuse large B-cell lymphoma by a new antimalarial artemisinin derivative. SM 1044 Cancer medicine 7:380–396
Choi W (2017a) Novel pharmacological activity of artesunate and artemisinin: their potential as anti-tubercular agents. J Clin Med 6(30)
Choi WH (2017b) Novel pharmacological activity of artesunate and artemisinin: their potential as anti-tubercular agents. J Clin Med 6(30)
Committee WMPA, Secretariat (2012) Malaria policy advisory committee to the WHO: conclusions and recommendations of September 2012 meeting. Springer,
Cox FE (2010) History of the discovery of the malaria parasites and their vectors. Parasit Vectors 3:5
Crellen T, Walker M, Lamberton PH, Kabatereine NB, Tukahebwa EM, Cotton JA, Webster JP (2016) Reduced efficacy of praziquantel against Schistosoma mansoni is associated with multiple rounds of mass drug administration. Clin Infect Dis 63:1151–1159
Cui X, Wang Y, Kokudo N, Fang D, Tang W (2010) Traditional Chinese medicine and related active compounds against hepatitis B virus infection bioscience trends 4
Czechowski T, Larson TR, Catania TM, Harvey D, Brown GD, Graham IA (2016) Artemisia annua mutant impaired in artemisinin synthesis demonstrates importance of nonenzymatic conversion in terpenoid metabolism. Proc Natl Acad Sci 113:15150–15155
Doenhoff MJ, Cioli D, Utzinger J (2008) Praziquantel: mechanisms of action, resistance and new derivatives for schistosomiasis. Curr Opin Infect Dis 21:659–667
Durantel D, Carrouée-Durantel S, Branza-Nichita N, Dwek RA, Zitzmann N (2004) Effects of interferon, ribavirin, and iminosugar derivatives on cells persistently infected with noncytopathic bovine viral diarrhea virus. Antimicrob Agents Chemother 48:497–504
Efferth T (2017) From ancient herb to modern drug: Artemisia annua and artemisinin for cancer therapy. In: Seminars in cancer biology, vol 46. Elsevier, pp 65–83
Efferth T, Romero MR, Wolf DG, Stamminger T, Marin JJ, Marschall M (2008) The antiviral activities of artemisinin and artesunate. Clin Infect Dis 47:804–811
El-Lakkany NM, el-Din SHS (2013) Haemin enhances the in vivo efficacy of artemether against juvenile and adult Schistosoma mansoni in mice parasitology research, vol 112, pp 2005–2015
Falzon D, Jaramillo E, Wares F, Zignol M, Floyd K, Raviglione MC (2013) Universal access to care for multidrug-resistant tuberculosis: an analysis of surveillance data. Lancet Infect Dis 13:690–697
Feng X, Chen W, Xiao L, Gu F, Huang J, Tsao BP, Sun L (2017) Artesunate inhibits type I interferon-induced production of macrophage migration inhibitory factor in patients with systemic lupus erythematosus. Lupus 26:62–72
French MD, Churcher TS, Gambhir M, Fenwick A, Webster JP, Kabatereine NB, Basáñez M-G (2010) Observed reductions in Schistosoma mansoni transmission from large-scale administration of praziquantel in Uganda: a mathematical modelling study. PLoS Negl Trop Dis 4:e897
Fröhlich T et al. (2017a) Synthesis of novel hybrids of quinazoline and artemisinin with high activities against Plasmodium falciparum, human cytomegalovirus, and leukemia cells ACS omega 2:2422
Fröhlich T, Reiter C, Ibrahim MM, Beutel J, Hutterer C, Zeitträger I, Bahsi H, Leidenberger M, Friedrich O, Kappes B, Efferth T, Marschall M, Tsogoeva SB (2017b) Synthesis of novel hybrids of Quinazoline and artemisinin with high activities against Plasmodium falciparum. Human Cytomegalovirus, and Leukemia Cells ACS Omega 2:2422–2431
Greenshields AL, Shepherd TG, Hoskin DW (2017) Contribution of reactive oxygen species to ovarian cancer cell growth arrest and killing by the anti-malarial drug artesunate. Mol Carcinog 56:75–93
Guo Z (2016) Artemisinin anti-malarial drugs in China. Acta Pharm Sin B 6:115–124
von Hagens C, Walter-Sack I, Goeckenjan M, Osburg J, Storch-Hagenlocher B, Sertel S, Elsässer M, Remppis BA, Edler L, Munzinger J, Efferth T, Schneeweiss A, Strowitzki T (2017) Prospective open uncontrolled phase I study to define a well-tolerated dose of oral artesunate as add-on therapy in patients with metastatic breast cancer (ARTIC M33/2). Breast Cancer Res Treat 164:359–369
Hahn F, Fröhlich T, Frank T, Bertzbach LD, Kohrt S, Kaufer BB, Stamminger T, Tsogoeva SB, Marschall M (2018) Artesunate-derived monomeric, dimeric and trimeric experimental drugs–their unique mechanistic basis and pronounced antiherpesviral activity. Antivir Res 152:104–110
Hakacova N, Klingel K, Kandolf R, Engdahl E, Fogdell-Hahn A, Higgins T (2013) First therapeutic use of Artesunate in treatment of human herpesvirus 6B myocarditis in a child. J Clin Virol 57:157–160
Han J, Wang H, Kanagarajan S, Hao M, Lundgren A, Brodelius PE (2016) Promoting artemisinin biosynthesis in Artemisia annua plants by substrate channeling. Mol Plant 9:946–948
Harper C (2007) Tuberculosis, a neglected opportunity. Nat Med 13:309–312
He Q, Fu X, Shi P, Liu M, Shen Q, Tang K (2017) Glandular trichome-specific expression of alcohol dehydrogenase 1 (ADH1) using a promoter-GUS fusion in Artemisia annua L plant cell. Tissue and Organ Culture (PCTOC) 130:61–72
Ho WE, Cheng C, Peh HY, Xu F, Tannenbaum SR, Ong CN, Wong WF (2012) Anti-malarial drug artesunate ameliorates oxidative lung damage in experimental allergic asthma. Free Radic Biol Med 53:498–507
Ho WE, Peh HY, Chan TK, Wong WF (2014) Artemisinins: pharmacological actions beyond anti-malarial. Pharmacol Ther 142:126–139
Hou L, Huang H (2016) Immune suppressive properties of artemisinin family drugs. Pharmacol Ther 166:123–127
Hurtley S, Ash C, Roberts L (2010) Landscapes of Infection. In: Landscapes of infection. American Association for the Advancement of Science
Ikram NK, Simonsen HT (2017) A review of biotechnological artemisinin production in plants. Front Plant Sci 8:1966
Ikram K, Binti NK, Beyraghdar Kashkooli A, Peramuna AV, van der Krol AR, Bouwmeester H, Simonsen HT (2017) Stable production of the antimalarial drug artemisinin in the moss Physcomitrella patens. Frontiers in bioengineering and biotechnology 5:47
Jana S, Iram S, Thomas J, Liekens S, Dehaen W (2017) Synthesis and anticancer activity of novel aza-artemisinin derivatives. Bioorg Med Chem 25:3671–3676
Ji Y, Zhang Y-C, Pei L-B, Shi L-L, Yan J-L, Ma X-H (2011) Anti-tumor effects of dihydroartemisinin on human osteosarcoma. Mol Cell Biochem 351:99–108
Jiao Y, Preston S, Koehler AV, Stroehlein AJ, Chang BCH, Simpson KJ, Cowley KJ, Palmer MJ, Laleu B, Wells TNC, Jabbar A, Gasser RB (2017) Screening of the ‘stasis Box’identifies two kinase inhibitors under pharmaceutical development with activity against Haemonchus contortus. Parasit Vectors 10:323
Jones I, Lund A, Riveau G, Jouanard N, Ndione RA, Sokolow SH, De Leo GA (2018) Ecological control of schistosomiasis in sub-Saharan Africa: restoration of predator-prey dynamics to reduce transmission ecology and evolution of infectious disease: pathogen control and public health management in low-income countries:236
Ju R-J et al (2018) Octreotide-modified liposomes containing daunorubicin and dihydroartemisinin for treatment of invasive breast cancer artificial cells, nanomedicine, and biotechnology, pp 1–13
Kadambari S, Atkinson C, Luck S, Macartney M, Conibear T, Harrison I, Booth C, Sharland M, Griffiths PD (2017) Characterising variation in five genetic loci of cytomegalovirus during treatment for congenital infection. J Med Virol 89:502–507
Kaptein SJ et al (2006) The anti-malaria drug artesunate inhibits replication of cytomegalovirus in vitro and in vivo. Antivir Res 69:60–69
Kar Han Lau P, Woods ML, Kanneyalal Ratanjee S, Tharayil John G (2011) Artesunate is ineffective in controlling valganciclovir-resistant cytomegalovirus infection. Clinical Infectious Diseases 52:279–279
Kayser O, Ryden A, Bouwmeester H, Spira CR, Osada H, Muranaka T (2011) The molecular cloning of dihydroartemisinic aldehyde reductase and its implication in artemisinin biosynthesis in Artemisia annua. Planta Med 77:SL28
Keiser J, N’Guessan NA, Adoubryn KD, Silué KD, Vounatsou P, Hatz C, Utzinger J, N’Goran EK (2010) Efficacy and safety of mefloquine, artesunate, mefloquine-artesunate, and praziquantel against Schistosoma haematobium: randomized, exploratory open-label trial. Clin Infect Dis 50:1205–1213
Kim SH, Kang SH, Kang BS (2016) Therapeutic effects of dihydroartemisinin and transferrin against glioblastoma. Nutr Res Pract 10:393–397
Konstat-Korzenny E, Ascencio-Aragón JA, Niezen-Lugo S, Vázquez-López R (2018) Artemisinin and its synthetic derivatives as a. Possible Therapy for Cancer Medical Sciences 6:19
Krishna S, Ganapathi S, Ster IC, Saeed MEM, Cowan M, Finlayson C, Kovacsevics H, Jansen H, Kremsner PG, Efferth T, Kumar D (2015) A randomised, double blind, placebo-controlled pilot study of oral artesunate therapy for colorectal cancer. EBioMedicine 2:82–90
Krungkrai J, Krungkrai SR (2016) Antimalarial qinghaosu/artemisinin: the therapy worthy of a Nobel prize. Asian Pac J Trop Biomed 6:371–375
Kumari K, Keshari S, Sengupta D, Sabat SC, Mishra SK (2017) Transcriptome analysis of genes associated with breast cancer cell motility in response to artemisinin treatment. BMC Cancer 17:858
del Villar LP, Burguillo FJ, López-Abán J, Muro A (2012) Systematic review and meta-analysis of artemisinin based therapies for the treatment and prevention of schistosomiasis. PLoS One 7:e45867
Lai HC, Singh NP, Sasaki T (2013) Development of artemisinin compounds for cancer treatment. Investig New Drugs 31:230–246
Lalloo DG, Shingadia D, Bell DJ, Beeching NJ, Whitty CJ, Chiodini PL (2016) UK malaria treatment guidelines 2016. J Infect 72:635–649
Laxminarayan R, Over M, Smith DL (2006) Will a global subsidy of new antimalarials delay the emergence of resistance and save lives? Health Aff 25:325–336
Leistikow RL, Morton RA, Bartek IL, Frimpong I, Wagner K, Voskuil MI (2010) The mycobacterium tuberculosis DosR regulon assists in metabolic homeostasis and enables rapid recovery from nonrespiring dormancy. J Bacteriol 192:1662–1670
Leto I, Coronnello M, Righeschi C, Bergonzi MC, Mini E, Bilia AR (2016) Enhanced efficacy of artemisinin loaded in transferrin-conjugated liposomes versus stealth liposomes against HCT-8 Colon Cancer cells. ChemMedChem 11:1745–1751
Li Y (2012) Qinghaosu (artemisinin): chemistry and pharmacology. Acta Pharmacol Sin 33:1141–1146
Li X, Li TT, Zhang XH, Hou LF, Yang XQ, Zhu FH, Tang W, Zuo JP (2013a) Artemisinin analogue SM934 ameliorates murine experimental autoimmune encephalomyelitis through enhancing the expansion and functions of regulatory T cell. PLoS One 8:e74108
Li Y, Wang S, Wang Y, Zhou C, Chen G, Shen W, Li C, Lin W, Lin S, Huang H, Liu P, Shen X (2013b) Inhibitory effect of the antimalarial agent artesunate on collagen-induced arthritis in rats through nuclear factor kappa B and mitogen-activated protein kinase signaling pathway. Transl Res 161:89–98
Li Z, Li Q, Wu J, Wang M, Yu J (2016) Artemisinin and its derivatives as a repurposing anticancer agent: what else do we need to do. Molecules 21:1331
Li X, Gu S, Sun D, Dai H, Chen H, Zhang Z (2018) The selectivity of artemisinin-based drugs on human lung normal and cancer cells. Environ Toxicol Pharmacol 57:86–94
Liang Y-J, Luo J, Yuan Q, Zheng D, Liu YP, Shi L, Zhou Y, Chen AL, Ren YY, Sun KY, Sun Y, Wang Y, Zhang ZS (2011) New insight into the antifibrotic effects of praziquantel on mice in infection with Schistosoma japonicum. PLoS One 6:e20247
Liu C-x (2017) Discovery and development of artemisinin and related compounds. Chinese Herbal Medicines 9:101–114
Liu L, Zuo LF, Zuo J, Wang J (2015) Artesunate induces apoptosis and inhibits growth of Eca109 and Ec9706 human esophageal cancer cell lines in vitro and in vivo. Mol Med Rep 12:1465–1472
Liu M, Shi P, Fu X, Brodelius PE, Shen Q, Jiang W, He Q, Tang K (2016) Characterization of a trichome-specific promoter of the aldehyde dehydrogenase 1 (ALDH1) gene in Artemisia annua. Plant Cell, Tissue and Organ Culture (PCTOC) 126:469–480
Liu L-x, Li-li J, Qiong C, Xiao-lin F (2017) Recent advances in the synthesis of Antischistosomal drugs and. Agents Mini reviews in medicinal chemistry 17:467–484
Liu R, Miller PA, Vakulenko SB, Stewart NK, Boggess WC, Miller MJ (2018) A synthetic dual drug Sideromycin induces gram-negative Bacteria to commit suicide with a gram-positive antibiotic. J Med Chem 61:3845–3854
Loop T, Pahl H (2003) Activators and target genes of Rel/NF-кB transcription factors. In: Nuclear factor кB. Springer, pp 1–48
Lovis L, Mak TK, Phongluxa K, Ayé Soukhathammavong P, Vonghachack Y, Keiser J, Vounatsou P, Tanner M, Hatz C, Utzinger J, Odermatt P, Akkhavong K (2012) Efficacy of praziquantel against Schistosoma mekongi and Opisthorchis viverrini: a randomized, single-blinded dose-comparison trial. PLoS Negl Trop Dis 6:e1726
Luo J, Zhu W, Tang Y, Cao H, Zhou Y, Ji R, Zhou X, Lu Z, Yang H, Zhang S, Cao J (2014) Artemisinin derivative artesunate induces radiosensitivity in cervical cancer cells in vitro and in vivo. Radiat Oncol 9:84
Makarov V et al. (2009) Benzothiazinones kill Mycobacterium tuberculosis by blocking arabinan synthesis science
Mandara CI et al (2018) High efficacy of artemether–lumefantrine and dihydroartemisinin–piperaquine for the treatment of uncomplicated falciparum malaria in Muheza and Kigoma districts. Tanzania Malaria journal 17:261
Maxmen A (2016) Busting the billion-dollar myth: how to slash the cost of drug development. Nature 536:388–390
McCoy A, Lissenden N, Morton A, Worrall E (2017) Towards an economics policy framework to combat malaria, in an era of insecticide resistance
Meskill JMM (2017) A Chinese Pioneer family: the Lins of Wu-feng, Taiwan, 1729–1895 vol 4941. Princeton University Press,
Miller LH, Su X (2011) Artemisinin: discovery from the Chinese herbal garden. Cell 146:855–858
Miller MJ, Walz AJ, Zhu H, Wu C, Moraski G, Möllmann U, Tristani EM, Crumbliss AL, Ferdig MT, Checkley L, Edwards RL, Boshoff HI (2011) Design, synthesis, and study of a mycobactin− artemisinin conjugate that has selective and potent activity against tuberculosis and malaria. J Am Chem Soc 133:2076–2079
Morere L et al (2015) Ex vivo model of congenital cytomegalovirus infection and new combination therapies. Placenta 36:41–47
Morris CA, Duparc S, Borghini-Fuhrer I, Jung D, Shin C-S, Fleckenstein L (2011) Review of the clinical pharmacokinetics of artesunate and its active metabolite dihydroartemisinin following intravenous, intramuscular, oral or rectal administration. Malar J 10:263
Morrissey C, Gallis B, Solazzi JW, Kim BJ, Gulati R, Vakar-Lopez F, Goodlett DR, Vessella RL, Sasaki T (2010) Effect of artemisinin derivatives on apoptosis and cell cycle in prostate cancer cells. Anti-Cancer Drugs 21:423–432
Nakase I, Lai H, Singh NP, Sasaki T (2008) Anticancer properties of artemisinin derivatives and their targeted delivery by transferrin conjugation. Int J Pharm 354:28–33
Nakase I, Gallis B, Takatani-Nakase T, Oh S, Lacoste E, Singh NP, Goodlett DR, Tanaka S, Futaki S, Lai H, Sasaki T (2009) Transferrin receptor-dependent cytotoxicity of artemisinin–transferrin conjugates on prostate cancer cells and induction of apoptosis. Cancer Lett 274:290–298
Nondo RSO, Moshi MJ, Erasto P, Masimba PJ, Machumi F, Kidukuli AW, Heydenreich M, Zofou D (2017) Anti-plasmodial activity of Norcaesalpin D and extracts of four medicinal plants used traditionally for treatment of malaria. BMC Complement Altern Med 17:167
Olofsson L, Engström A, Lundgren A, Brodelius PE (2011) Relative expression of genes of terpene metabolism in different tissues of Artemisia annua L. BMC Plant Biol 11:45
Organization WH (2011) Global tuberculosis control: WHO report 2011
Organization WH (2016) World health statistics 2016: monitoring health for the SDGs sustainable development goals. World Health Organization
Pagan JD, Kitaoka M, Anthony RM (2018) Engineered sialylation of pathogenic antibodies in vivo attenuates autoimmune disease. Cell 172:564–577 e513
Park J, Lai HC, Singh M, Sasaki T, Singh NP (2014) Development of a dihydroartemisinin-resistant Molt-4 leukemia cell line. Anticancer Res 34:2807–2810
Pérez dVL, Burguillo F, Lopez-Aban J, Muro A (2012) Systematic review and meta-analysis of artemisinin based therapies for the treatment and prevention of schistosomiasis
Poespoprodjo JR, Fobia W, Kenangalem E, Lampah DA, Sugiarto P, Tjitra E, Anstey NM, Price RN (2014) Dihydroartemisinin-piperaquine treatment of multidrug resistant falciparum and vivax malaria in pregnancy. PLoS One 9:e84976
Poravuth Y, Socheat D, Rueangweerayut R, Uthaisin C, Pyae Phyo A, Valecha N, Rao BHK, Tjitra E, Purnama A, Borghini-Fuhrer I, Duparc S, Shin CS, Fleckenstein L (2011) Pyronaridine-artesunate versus chloroquine in patients with acute Plasmodium vivax malaria: a randomized, double-blind, non-inferiority trial. PLoS One 6:e14501
Premji ZG (2009) Coartem®: the journey to the clinic Malaria journal, vol 8, p S3
Qi F et al (2013) Traditional Chinese medicine and related active compounds: a review of their role on hepatitis B virus infection. Drug discoveries & therapeutics 7:212–224
Ramacher M, Umansky V, Efferth T (2009) Effect of artesunate on immune cells in ret-transgenic mouse melanoma model. Anti-Cancer Drugs 20:910–917
Rojas Vargas JA, López AG, Froeyen M (2016) Molecular Docking Studies of 1, 2, 4, 5-tetrasubstituted Imidazoles with Different Protein Targets of Mycobacterium tuberculosis Biomirror 7
Rosales-Mendoza S, Nieto-Gómez R, Angulo C (2017) A perspective on the development of plant-made vaccines in the fight against Ebola virus. Front Immunol 8:252
Ross AG, Chau TN, Inobaya MT, Olveda RM, Li Y, Harn DA (2017) A new global strategy for the elimination of schistosomiasis. In: A new global strategy for the elimination of schistosomiasis. Elsevier
Sayce AC, Alonzi DS, Killingbeck SS, Tyrrell BE, Hill ML, Caputo AT, Iwaki R, Kinami K, Ide D, Kiappes JL, Beatty PR, Kato A, Harris E, Dwek RA, Miller JL, Zitzmann N (2016) Iminosugars inhibit dengue virus production via inhibition of ER alpha-glucosidases—not glycolipid processing enzymes. PLoS Negl Trop Dis 10:e0004524
Schnepf N, Corvo J, Sanson-Le Pors M-J, Mazeron M-C (2011) Antiviral activity of ganciclovir and artesunate towards human cytomegalovirus in astrocytoma cells. Antivir Res 89:186–188
Schreiber A, Härter G, Schubert A, Bunjes D, Mertens T, Michel D (2009) Antiviral treatment of cytomegalovirus infection and resistant strains. Expert Opin Pharmacother 10:191–209
Schwartz DM, Bonelli M, Gadina M, O'shea JJ (2016) Type I/II cytokines, JAKs, and new strategies for treating autoimmune diseases. Nat Rev Rheumatol 12:25–36
Shayo A, Buza J, Ishengoma DS (2015) Monitoring of efficacy and safety of artemisinin-based anti-malarials for treatment of uncomplicated malaria: a review of evidence of implementation of anti-malarial therapeutic efficacy trials in Tanzania. Malar J 14:135
Shi JQ et al. (2013) Antimalarial Drug Artemisinin Extenuates Amyloidogenesis and Neuroinflammation in APP swe/PS 1dE9 Transgenic Mice via Inhibition of Nuclear Factor-κ B and NLRP 3 Inflammasome Activation CNS neuroscience & therapeutics 19:262–268
Shuhua X, Tanner M, N’Goran EK, Utzinger J, Chollet J, Bergquist R, Minggang C, Jiang Z (2002) Recent investigations of artemether, a novel agent for the prevention of schistosomiasis japonica, mansoni and haematobia. Acta Trop 82:175–181
Simonsen HT, Weitzel C, Christensen SB (2013) Guaianolide sesquiterpenoids: pharmacology and biosynthesis. In: Natural products. Springer, pp 3069–3098
Sinka ME, Bangs MJ, Manguin S, Rubio-Palis Y, Chareonviriyaphap T, Coetzee M, Mbogo CM, Hemingway J, Patil AP, Temperley WH, Gething PW, Kabaria CW, Burkot TR, Harbach RE, Hay SI (2012) A global map of dominant malaria vectors. Parasit Vectors 5:69
Skrahina A et al (2011) Alarming levels of drug-resistant tuberculosis in Belarus: results of a survey in Minsk. Eur Respir J:erj01454–erj02011
Stebbins RC, Emch M, Meshnick SR (2018) The effectiveness of community bed net use on Malaria Parasitemia among children less than 5 years old in Liberia
Tambo E, Ai L, Zhou X, Chen JH, Hu W, Bergquist R, Guo JG, Utzinger J, Tanner M, Zhou XN (2014) Surveillance-response systems: the key to elimination of tropical diseases. Infectious diseases of poverty 3:17
Teoh KH, Polichuk DR, Reed DW, Covello PS (2009) Molecular cloning of an aldehyde dehydrogenase implicated in artemisinin biosynthesis in Artemisia annua. Botany 87:635–642
Thanaketpaisarn O, Waiwut P, Sakurai H, Saiki I (2011) Artesunate enhances TRAIL-induced apoptosis in human cervical carcinoma cells through inhibition of the NF-κB and PI3K/Akt signaling pathways. Int J Oncol 39:279–285
Tilaoui M, Mouse HA, Jaafari A, Zyad A (2014) Differential effect of artemisinin against cancer cell lines. Natural products and bioprospecting 4:189–196
Tu Y (2011) The discovery of artemisinin (qinghaosu) and gifts from Chinese medicine. Nat Med 17:1217–1220
Tu Y (2016) Artemisinin—a gift from traditional Chinese medicine to the world (nobel lecture). Angew Chem Int Ed 55:10210–10226
Utzinger J et al (2009) Schistosomiasis and neglected tropical diseases: towards integrated and sustainable control and a word of caution. Parasitology 136:1859–1874
Vale N, Gouveia MJ, Rinaldi G, Brindley PJ, Gärtner F, da Costa JMC (2017) Praziquantel for schistosomiasis, single drug revisited metabolism, mode of action and resistance antimicrobial agents and chemotherapy:AAC, pp 02582–02516
Van Huijsduijnen RH et al (2013) Anticancer properties of distinct antimalarial drug classes. PLoS One 8:e82962
Wang JX et al (2008) The new water-soluble artemisinin derivative SM905 ameliorates collagen-induced arthritis by suppression of inflammatory and Th17 responses. Br J Pharmacol 153:1303–1310
Wang J-x, L-f H, YAng Y, TAng W, Li Y, J-p Z (2009) SM905, an artemisinin derivative, inhibited NO and pro-inflammatory cytokine production by suppressing MAPK and NF-κB pathways in RAW 264.7 macrophages. Acta Pharmacol Sin 30:1428
Wang X-Q, Liu HL, Wang GB, Wu PF, Yan T, Xie J, Tang Y, Sun LK, Li C (2011a) Effect of artesunate on endotoxin-induced uveitis in rats. Invest Ophthalmol Vis Sci 52:916–919
Wang Y, Yang K, Jing F, Li M, Deng T, Huang R, Wang B, Wang G, Sun X, Tang KX (2011b) Cloning and characterization of trichome-specific promoter of cpr71av1 gene involved in artemisinin biosynthesis in Artemisia annua L. Mol Biol 45:751–758
Wang J, Zhang CJ, Chia WN, Loh CCY, Li Z, Lee YM, He Y, Yuan LX, Lim TK, Liu M, Liew CX, Lee YQ, Zhang J, Lu N, Lim CT, Hua ZC, Liu B, Shen HM, Tan KSW, Lin Q (2015) Haem-activated promiscuous targeting of artemisinin in Plasmodium falciparum. Nat Commun 6:10111
Wang B, Kashkooli AB, Sallets A, Ting HM, de Ruijter NCA, Olofsson L, Brodelius P, Pottier M, Boutry M, Bouwmeester H, van der Krol AR (2016) Transient production of artemisinin in Nicotiana benthamiana is boosted by a specific lipid transfer protein from a. annua. Metab Eng 38:159–169
Wang J, Xu C, Lun Z-R, Meshnick SR (2017a) Unpacking ‘artemisinin resistance. Trends Pharmacol Sci 38:506–511
Wang J, Zhang J, Shi Y, Xu C, Zhang C, Wong YK, Lee YM, Krishna S, He Y, Lim TK, Sim W, Hua ZC, Shen HM, Lin Q (2017b) Mechanistic investigation of the specific anticancer property of artemisinin and its combination with aminolevulinic acid for enhanced anticolorectal cancer activity. ACS central science 3:743–750
Wei J, Zhang L, Ren L, Zhang J, Liu J, Duan J, Yu Y, Li Y, Peng C, Zhou X, Sun Z (2017) Endosulfan induces cell dysfunction through cycle arrest resulting from DNA damage and DNA damage response signaling pathways. Sci Total Environ 589:97–106
Weifeng T, Feng S, Xiangji L, Changqing S, Zhiquan Q, Huazhong Z, Peining Y, Yong Y, Mengchao W, Xiaoqing J, Wan-yee L (2011) Artemisinin inhibits in vitro and in vivo invasion and metastasis of human hepatocellular carcinoma cells. Phytomedicine 18:158–162
Wen W, Yu R (2011) Artemisinin biosynthesis and its regulatory enzymes: Progress and perspective. Pharmacogn Rev 5:189
Wencewicz TA, Miller MJ (2017) Sideromycins as pathogen-targeted antibiotics
White N, Pukrittayakamee S, Hien T, Faiz M, O a M, Dondorp AM (2013) Malaria Lancet 6736:13
White N, Pukrittayakamee S, Hien T, Faiz M, Mokuolu O, Malaria DA (2014) Malaria. Lancet [Internet] 383(9918):723–735
Wilson ME, Kantele A, Jokiranta TS (2011) Review of cases with the emerging fifth human malaria parasite. Plasmodium knowlesi Clinical infectious diseases 52:1356–1362
Wohlfarth C, Efferth T (2009) Natural products as promising drug candidates for the treatment of hepatitis B and C. Acta Pharmacol Sin 30:25–30
Wong YK, Xu C, Kalesh KA, He Y, Lin Q, Wong WSF, Shen HM, Wang J (2017) Artemisinin as an anticancer drug: recent advances in target profiling and mechanisms of action. Med Res Rev 37:1492–1517
Wu X, Zhang W, Shi X, An P, Sun W, Wang Z (2010) Therapeutic effect of artemisinin on lupus nephritis mice and its mechanisms. Acta Biochim Biophys Sin 42:916–923
Wu H, Hu B, Zhou X, Zhou C, Meng J, Yang Y, Zhao X, Shi Z, Yan S (2018) Artemether attenuates LPS-induced inflammatory bone loss by inhibiting osteoclastogenesis and bone resorption via suppression of MAPK signaling pathway. Cell Death Dis 9
Yakasai AM et al. (2015) Adherence to artemisinin-based combination therapy for the treatment of uncomplicated malaria: a systematic review and meta-analysis Journal of tropical medicine 2015
YANG L, ZHANG D (2017) Summary of dihydroartemisinin and its application for the treatment of lupus erythematosus. Chin Sci Bull 62:2007–2012
Yang Z, Ding J, Yang C, Gao Y, Li X, Chen X, Peng Y, Fang J, Xiao S (2012) Immunomodulatory and anti-inflammatory properties of artesunate in experimental colitis. Curr Med Chem 19:4541–4551
Yi P, Park J-S, Melton DA, Huang KC (2017) NF-κB was discovered 30 years ago as a rapidly inducible transcription factor. Since that time, it has been found to have a broad role in gene induction in diverse cellular responses, particularly throughout the immune system. Here, we summarize elaborate regulatory pathways involving this transcription factor and use recent discoveries in human genetic diseases to place specific proteins within their. Cell 168:1–2
Zhang CJ, Wang J, Zhang J, Lee YM, Feng G, Lim TK, Shen HM, Lin Q, Liu B (2016) Mechanism-guided design and synthesis of a mitochondria-targeting artemisinin analogue with enhanced anticancer activity. Angew Chem Int Ed 55:13770–13774
Zhang S, Shi L, Ma H, Li H, Li Y, Lu Y, Wang Q, Li W (2017) Dihydroartemisinin induces apoptosis in human gastric cancer cell line BGC-823 through activation of JNK1/2 and p38 MAPK signaling pathways. Journal of Receptors and Signal Transduction 37:174–180
Zhang J, Sun X, Wang L, Wong YK, Lee YM, Zhou C, Wu G, Zhao T, Yang L, Lu L, Zhong J, Huang D, Wang J (2018) Artesunate-induced mitophagy alters cellular redox status. Redox Biol 19:263–273
Zheng H, Colvin CJ, Johnson BK, Kirchhoff PD, Wilson M, Jorgensen-Muga K, Larsen SD, Abramovitch RB (2017) Inhibitors of mycobacterium tuberculosis DosRST signaling and persistence. Nat Chem Biol 13:218–225
Zhou H-J, Zhang J-L, Li A, Wang Z, Lou X-E (2010) Dihydroartemisinin improves the efficiency of chemotherapeutics in lung carcinomas in vivo and inhibits murine Lewis lung carcinoma cell line growth in vitro. Cancer Chemother Pharmacol 66:21–29
Acknowledgements
The authors are thankful to the Shanghai Jiao Tong University, Shanghai 200240, China, for providing literature facilities. We are also thankful to the anonymous reviewers for their valuable comments. We would also like to thank Dr. Shujaul Mulk Khan of Quaid-i-Azam University Islamabad, Pakistan for his guidance and checking plagiarism in the first draft of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that there is no conflict of interest to claim.
Additional information
Communicated by: Yuval Cohen
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Saeed-ur-Rahman, Khalid, M., Kayani, SI. et al. Biological Activities of Artemisinins Beyond Anti-Malarial: a Review. Tropical Plant Biol. 12, 231–243 (2019). https://doi.org/10.1007/s12042-019-09228-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12042-019-09228-0