
Abstract
Aminoglycoside antibiotics, including gentamicin (GM), induce delayed ototoxic effects such as hearing loss after prolonged use, which results from the death of hair cells. However, the mechanisms underlying the ototoxicity of aminoglycosides warrant further investigation, and there are currently no effective drugs in the clinical setting. Herein, the therapeutic effect of the flavonoid compound rutin against the ototoxic effects of GM in zebrafish hair cells was investigated. Animals incubated with rutin (100–400 µmol/L) were protected against the pernicious effects of GM (200 µmol/L). We found that rutin improves hearing behavior in zebrafish, and rutin was effective in reducing the number of Tunel-positive cells in the neuromasts of the zebrafish lateral line and promoting cell proliferation after exposure to GM. Subsequently, rutin exerted a protective effect against GM-induced cell death in HEI-OC1 cells and could limit the production of cytosolic reactive oxygen species (ROS) and diminish the percentage of apoptotic cells. Additionally, the results of the proteomic analysis revealed that rutin could effectively inhibit the expression of necroptosis and apoptosis related genes. Meanwhile, molecular docking analysis revealed a high linking activity between the molecular docking of rutin and STAT1 proteins. The protection of zebrafish hair cells or HEI-OC1 cells from GM-induced ototoxicity by rutin was attenuated by the introduction of STAT1 activator. Finally, we demonstrated that rutin significantly improves the bacteriostatic effect of GM by in vitro experiments, emphasising its clinical application value. In summary, these results collectively unravel a novel therapeutic role for rutin as an otoprotective drug against the adverse effects of GM.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Aminoglycosides are a class of antibiotics that are extensively used in clinical practice to combat bacterial infections such as Gram-negative bacteria and mycobacteria and can be used in combination with β-lactam antibiotics to enhance efficacy [1]. The toxicity of aminoglycosides is chiefly related to ototoxicity and nephrotoxicity. Ototoxicity refers to the toxic effects of drugs or certain physical factors on the cochlea, vestibular system, and other parts of the cochlea, triggering structural and functional damage to the cochlea, such as ototoxic deafness [2]. Even marginal hearing loss hinders language, cognitive, and social development, leading to reduced psychosocial function, especially in young children. Therefore, the ototoxicity of aminoglycosides severely limits their clinical use [3].
Aminoglycoside antibiotics primarily elicit hearing loss by damaging hair cells on the basilar membrane of the inner ear. Aminoglycoside-induced generation of reactive oxygen species (ROS) is one of the central mechanisms of sensory cell death. Aminoglycosides can penetrate hair cells through vascular striations, mechano-electrical transducer channels, and cytosolization [4]. Following infiltration, they generate large amounts of intracellular ROS through mitochondrial calcium overload and other pathways, which in turn, disrupt normal equilibrium and lead to the oxidation of DNA, lipids, proteins, and other important cellular constituents, ultimately resulting in cell death. The pathway of hair cell death is intricate and encompasses necrosis, as well as caspase-dependent and caspase-independent apoptosis [5].
Gentamicin (GM) is a commonly administered aminoglycoside in clinical practice owing to its favorable efficacy against Gram-negative bacteria and tuberculosis [6]. To date, GM has not been completely replaced by less ototoxic and equally effective drugs in the clinical setting. Hence, there is an urgent need to discover an effective drug for the prevention of gentamicin-induced ototoxicity [7].
Rutin, also termed rutoside or quercetin-3- O -rutinoside, is a naturally occurring bioflavonoid found in vegetables and fruits [8]. Numerous studies have demonstrated its powerful antioxidant capacity, especially its excellent free radical scavenging activity [9]. Prince et al. evaluated the inhibition of lipid peroxidation and the antioxidant status of rutin in diabetes mellitus in a rat model and described that oral administration of rutin for 45 days exerted a significant antioxidant effect in mice with streptozotocin (STZ)-induced experimental diabetes [10]. At the same time, Nafees et al. observed that rutin could alleviate the damage induced by inflammation and oxidative stress in Wistar rats by regulating the MAPK pathway [11].
Importantly, a previous study has reported that rutin mitigated cisplatin-induced hair cell death in neonatal cochlear explants in vitro. The potential mechanism involved the alleviation of mitochondrial damage, the scavenging of reactive oxygen species (ROS), the suppression of the MAPK signaling pathway, and the activation of the PI3K/AKT signaling pathway [12]. However, reports on the ototoxicity of rutin against GM are scarce.
The zebrafish is an ideal animal model for studying human auditory hair cell function. Zebrafish have hair cell-rich neuromast located on the lateral line of the ear sac and body surface, which are very convenient for live labeling staining and observation [13]. This study employed zebrafish as a model organism to examine the potential protective effects of rutin against GM-induced ototoxicity in hair cells and its mechanism of action by conducting in vivo and in vitro experiments.
Materials and Methods
Animals
Wild-type (AB) and Tg (Brn3C: GFP) transgenic zebrafish used in the research were donated by Professor Dong Liu at Nantong University (Nantong, China). All zebrafish were raised and maintained at 28 ± 0.5℃ on a 14 h/10 h light/dark cycle. The breeding program was carried out according to The Zebrafish Book (ZFIN, https://zfin.org/).
Drug Preparation
GM (Solebol, Beijing, China) was dissolved in dimethyl sulfoxide at a concentration of 2 mmol/L. Rutin (Original Bio, Shanghai, China) was dissolved in dimethyl sulfoxide at a concentration of 30 mmol/L. 2-NP, an aselective enhancer of STAT1 transcription, was sourced from Med Chem Express (MCE, Beijing, China) and dissolved in DMSO using an ultrasonic bath. The stock solution was then diluted to working concentrations (Rutin, 25-400 µmol/L; GM, 80 or 200 µmol/L; 2-NP, 200µmol/L) in fish water or culture medium. All the reagents are prepared before each use.
Cells
HEI-OC1 cells used were maintained in high-glucose Dulbecco’s modified Eagle’s medium supplemented with 10% fetal bovine serum under an atmosphere of 10% CO2 at 34 °C. HEI-OC1 cells were cultured in a humidity-controlled incubator without antibiotics.
Drug Treatments
Zebrafish larvae at 5 dpf were treated with the pre-defined drugs in a 24-well plate. Larvae were exposed to rutin for 3 h and then co‐treated with rutin and GM (200 µmol/L) for 12 h.
For cell viability assays, HEI-OC1 cells were incubated with different concentrations of rutin (0-200 µmol/L) for 24 h or a single dose (100 µmol/L) of rutin for 0–48 h, or exposed to rutin for 6 h and then co-treated with rutin and GM (80 µmol/L) for 24 h; For cellular ROS and apoptotic detection, HEI-OC1 cells exposed to rutin (0-100 µmol/L)for 6 h and then co-treated with rutin and GM (80 µmol/L) for 24 h; For proteomic analysis, HEI-OC1 cells were incubated with 100 µmol/L Rutin for 24 h; To explore the role of STAT1, Zebrafish and HEI-OC1 cells were incubated with 200 or 80 µmol/L GM for 24 h or initially treated with 400 or 100 µmol/L rutin or 2-NP (200 µmol/L) for 6 h.
Zebrafish Hearing Behavior Assay
In this study, we used the startle response assay to examine zebrafish hearing. For the behavioral experiments, one zebrafish larvae at 5 dpf was placed in 300µL of fish media per well of a 96-well plate. For each experimental condition, a group of 13 larvae (1 column) was used. The well plate every two seconds. Locomotor activity was subsequently tracked and quantified (as distance travelled) for 60 additional seconds utilizing the Zebrabox system from ViewPoint Behavior Technology.
Whole Mount Immunohistochemistry
The immunohistochemical assay was carried out according to the methodologies outlined in previous studies [14,15,16]. The antibodies used in this study were as follows: anti-Myosin VIIa antibody (1:200, Abcam, UK) with Alexa Fluor 488 antibody (1:200; Sigma‐Aldrich), anti‐BrdU antibody (1:200; Abcam, USA), and Alexa Fluor 594 antibody (1:200; Sigma‐Aldrich). Nuclei were counter-stained using DAPI (1:200; Thermofisher).
FM1-43 Staining
A living cell fluorescent dye FM1-43 (Macklin, Shanghai, China) was used to determine the functionality of existing hair cells. The collected 5dpf zebrafish embryos were anesthetized with 0.03% MS-222 (Sigma-Aldrich) and immersed in 5 µM FM1-43 solution for 45 s in the dark. After discarding the dye, the larvae were washed three times with PBS and images were taken under a fluorescence microscope (Nikon AIR confocal microscope). Fluorescence intensity was quantified using ImageJ, and the background fluorescence intensity was subtracted.
TUNEL Staining
The TUNEL staining is used for measuring apoptotic DNA fragmentation. TUNEL staining (Servicebio, Beijing, China) was performed according to the manufacturers instructions.
CCK-8 Assay
Cell activity was assayed using the Cell Counting Kit kit kit-8 (CCK-8, Servicebio, Jinan, China) according to the instructions of the reagent vendor.
Intracellular ROS Detection
Staining of ROS in HEI-OC1 cells was performed using a fluorescent probe for 2’,7’-dichlorofluorescein diacetate (DCFH-DA, Solepol, Beijing, China).DCFH-DA was diluted in serum-free medium to a final concentration of 5 µmol/L. Then, the cell culture medium was discarded and the appropriate volume of diluted DCFH-DA was added to cover the cells. The cells were incubated at 37 °C for 30 min. The cells were then washed three times with serum-free medium to remove unbound DCFH-DA. finally, the stained cells were analysed using confocal fluorescence microscopy.
Flow Cytometric Analysis
Cell apoptosis was determined by using the Annexin V-PE/7-AAD apoptosis detection kit (BD Biosciences, USA) following the manufacturer’s instructions.
RNA Extraction and qPCR
The steps for real-time fluorescence quantitative PCR were consistent with those reported previously [12]. Real-time quantitative PCR (qRT-PCR) was performed using the following kits: RNA Extraction Kit (Invitrogen, Beijing, China), Super Script II Reverse Transcriptase Kit (Invitrogen, USA) and Power SYBR Green PCR Master Mix (Applied Biosystems, Foster City, USA). The primers used in the experiments are listed in Table 1.
Proteomic Analysis
Proteins were extracted from rutin-treated and solvent-treated HEI-OC1 cells and analyzed proteomically using mass spectrometry (n = 5). KEGG and GO enrichment analyses were subsequently carried out for relevant differential genes.
Binding of Rutin to Predicted Targets
The SDF structure files of the compound rutin were obtained from the PubChem website (https://pubchem.ncbi.nlm.nih.gov/). Subsequently, the SDF file was converted into a PDB file using OpenBabel 2.3.2 software. The receptor proteins STAT1 were sourced from the Protein Data Bank database (www.wwpdb.org). To prepare the receptor proteins for docking studies, water and ligands were removed using PYMOL 2.3.4 software. The docking analysis was conducted using AutoDock Vina (1.1.2), an open-source molecular docking software developed by Scripps. The receptor proteins were further modified using Tools software, and the Grid Box command under the Grid program was utilized to define the binding site. The lattice spacing was set to 1, with the center of the pocket designated as the binding site center. Three-dimensional structures of the binding pockets were visualized using PyMOL software.
Antimicrobial Susceptibility Testing
Antimicrobial susceptibility test disks with 20 µg GM were purchased from Oxoid (Hants, UK). The Escherichia coli-, Klebsiella pneumoniae-, and Salmonella enterica strain was kindly provided by the microbiology laboratory, Shandong Provincial Hospital. GM-disks were treated with different concentrations of rutin (0–200 µM/L) and then placed in bacterial media for drug sensitivity experiments. The Petri dish was incubated at 37 °C for 12 h, and the bacteriostatic zone diameter around the samples was measured using a scale.
Data Analysis and Statistics
Each experiment was repeated at least three times and considered valid with the trials showing similar results. Results were presented as the mean ± standard deviation (SD). Students unpaired t-test was used to compare the results between the GM-treated and control groups or GM with Rutin-treated and GM-treated groups. The significant difference was determined by either Student’s t-test with p < 0.05.
Result
Rutin Improves Hearing Behavior in Zebrafish
First, in order to explore the effect of rutin on hearing damage caused by GM, we conducted an auditory-behavioral experiment in zebrafish (Fig. 1a). The results showed that GM treatment alone caused zebrafish to be less sensitive to sound (vibration) and move less in one corner of the well plate compared to the control. Whereas, after the addition of rutin pretreatment, the range of zebrafish movement was expanded (Fig. 1b).

Rutin improved the hearing behavior of zebrafish. 5 dpf zebrafish were incubated with 200 µmol/L GM for 12 h or pre-treated with 100 µmol/L to 400 µmol/L of rutin for 3 h and then co‐treated with rutin and GM (200 µmol/L) for 12 h. (a) The schematic diagram of experimental process. (b) Representative displacement graph of the hearing behavior in zebrafish. Quantification of the number of startle reflexes per minute (c) and the speed of movement of zebrafish (d), and represented as mean ± SD (n = 13). ****P < 0.0001; ns: no significant difference
Similarly, the results of quantitative analysis showed that rutin significantly reversed the GM-induced reduction in the number of stress reflexes per minute and distance traveled in zebrafish (Fig. 1c, d).
Rutin Protects the Auditory hair Cells of Zebrafish from GM-induced cell Death
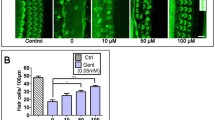
To quantify the number of hair cells following GM exposure, confocal immunofluorescence analysis was utilized. Hair cells were labeled with Myosin VIIa (green), whilst nuclei were labeled with DAPI (Fig. 2aI). The results displayed that GM (200 µmol/L) promoted a significant loss of hair cells in the zebrafish lateral line (Fig. 1a, b). To assess the impact of rutin in protecting zebrafish hair cells against GM-induced hair cell injury, zebrafish larvae (5 pdf) were pre‐treated with 0 µmol/L to 400 µmol/L of rutin for 3 h and then co‐treated with rutin and GM (200 µmol/L) for 12 h (Fig. 2a). The results revealed that pre‐treatment and co‐treatment with rutin effectively increased the number of surviving hair cells following GM treatment (Fig. 2b). Moreover, a Tg (brn3c: GFP) transgenic zebrafish line that expressed a membrane‐bound GFP in hair cells was established (Fig. 2aII). Likewise, co‐treatment with rutin increased the survival rate of GM‐exposed hair cells in a dose‐dependent manner (Fig. 2c).

Rutin alleviates gentamicin (GM)-induced damage to the lateral line neuromast hair cells (hair cells) of zebrafish. 5 dpf zebrafish were incubated with 200 µmol/L GM for 12 h or pre-treated with 100 µmol/L to 400 µmol/L of rutin for 3 h and then co‐treated with rutin and GM (200 µmol/L) for 12 h. Control animals were exposed to the vehicle alone (DMSO). (a) I hair cells were labeled with Myosin VIIa (green), whilst nuclei were labeled with DAPI; II Green fluorescent protein (GFP) expression from the Tg(Brn3C: GFP) transgenic zebrafish depicting the hair cells; III FM1-43 staining displaying functional hair cells in the neuromasts in the different treatment groups. The scale bar represents 5 μm. (b) Quantification of the number of hair cells (MyosinVIIa+) per neuromast after different treatments, presented as mean ± SD (n = 10). ***P < 0.001; ****P < 0.0001; ns: no significant difference. (c) Quantification of the number of surviving cilia per neuromast after the different treatments represented as mean ± SD (n = 10). *P < 0.05; ****P < 0.0001. (d) The FM1‐43 fluorescent intensity per neuromast was calculated for each group and represented as mean ± SD (n = 10). ***P < 0.001; ****P < 0.0001; ns: no significant difference
In addition, to explore the potential of rutin in promoting the functional recovery of damaged hair cell clusters, hair cell mechanotransduction was performed using FM1-43, a fluorescent dye that infiltrates MET channels (Fig. 2aIII) [17]. Interestingly, the fluorescence intensity of FM1-43 in hair cells within each neuromast decreased after GM stimulation and was reversed by rutin in a dose-dependent manner (Fig. 2d).
Rutin Prevents hair cell Death and Promotes cell Proliferation in Neuromasts
To determine the mechanism by which rutin prevents GM-induced hair cell death, a Terminal deoxynucleotidyl transferase (TdT) dUTP Nick‐End labeling (TUNEL) assay was carried out. The results exhibited a large number of TUNEL‐positive cells following 12 h of GM treatment in neuromasts, whereas co‐treatment with rutin markedly reduced the number of TUNEL‐positive cells (Fig. 3a, b).

Rutin promotes cell proliferation and protects against gentamicin (GM)-induced hair cell (hair cells) death in neuromasts. 5 dpf zebrafish were incubated with 200 µmol/L GM for 12 h or pre-treated with 100 µmol/L to 400 µmol/L of rutin for 3 h and then co‐treated with rutin and GM (200 µmol/L) for 12 h. (a) Confocal microscopy images delineating neuromast hair cells. TUNEL (pink) labeling was used to examine hair cell apoptosis, and BrdU (pink) reflects hair cell proliferation. Cell nuclei were labeled with DAPI (blue). The scale bar represents 5 μm. (b) The numbers of TUNEL‐positive cells per neuromast were calculated for each treatment group and presented as mean ± SD (n = 10). ****P < 0.0001. (c) The numbers of proliferated cells (BrdU+) per neuromast were calculated for each treatment group and expressed as mean ± SD (n = 10). ****P < 0.0001
To determine the impact of the oto-protective function of GM in cell proliferation, a 5‐Bromo‐2‐deoxyUridine (BrdU, proliferation marker) immunofluorescence assay was performed. Immunofluorescence analysis determined that BrdU‐positive cells were largely situated at the edge of the neuromast (supporting cells are usually in this area) in the control group, whilst GM treatment significantly reduced the proportion of BrdU-positive cells. In contrast, zebrafish co‐treated with rutin and GM exhibited a significantly increased number of BrdU‐positive cells compared with GM treatment alone (Fig. 3a, c).
Rutin Reverses GM-induced HEI-OC1 cell Damage by Inhibiting ROS Production and Apoptosis
HEI-OC1 cells were initially exposed to progressively increasing concentrations of rutin (0-200 µM/L) for 24 h to determine the ototoxicity of rutin. The results showed that rutin not only exhibited no cytotoxicity towards HEI-OC1 cells but also improved cell viability irrespective of concentration, and its effect was optimal at a concentration of 100 µM/L (Fig. 4a). Specifically, 100 µM/L of rutin significantly increased the cell viability of HEI-OC1 cells between 0 and 48 h compared to the control group (Fig. 4b). Therefore, 100 µM of rutin was selected for the ensuing experiments. Then, the effect of rutin on GM-induced ototoxicity was assessed. The results uncovered that GM (80 µM) significantly reduced the cell viability of HEI-OC1 cells, while rutin (25 to 200 µM) reversed this effect (Fig. 4c).

Effects of rutin on gentamicin (GM)-induced ototoxicity in HEI-OC1 cells. (a) Cell viability after 24 h treatment with rutin (0-200 µmol/L). (b) Cell viability following treatment with or without rutin (100 µmol/L) for different durations (0–48 h). (c) HEI-OC1 cells were pre-treated with 0 µmol/L to 200 µmol/L of rutin for 6 h and then co‐treated with rutin and GM (80 µmol/L) for 24 h. CCK analysis analyzing cell viability. (d) Flow cytometry analysis of cell apoptosis via Annexin V-PE/7-AAD staining after co-incubation with GM and rutin. (e) Reactive oxygen species (ROS) levels in GM and rutin (100 µmol/L) treated for 24 h were detected using DCFH-DA. The scale bar represents 5 μm. (f) DCFH-DA fluorescenceintensity quantification. (g) The expression level of apoptosis-related genes was determined via quantitative real-time PCR. Student’s t-test was used to compare expression levels between the groups’ P < 0.05 (*), P < 0.01 (**), P < 0.001 (***), and P < 0.001 (****); ns: no significant difference. Each experiment was performed in triplicate
Apoptosis was evaluated by fluorescence-activated cell sorting (FACS) of cells stained with AnnexinV antiserum and the DNA intercalator 7-AAD to identify apoptotic cells (AnnexinV- positive and 7-AAD-positive). Compared with the control group, the number of apoptotic cells was high following GM treatment, and this effect was reversed by treatment with rutin (Fig. 4d).
ROS were detected using DCFH-DA, and the fluorescence intensity was calculated. GM exposure significantly increased intracellular ROS levels in HEI-OC1 cells, whereas rutin pretreatment significantly attenuated this effect (Fig. 4e, f).
Of note, GM significantly up-regulated the expression of pro-apoptotic genes (Caspace-1 and bax) and down-regulated that of anti-apoptotic (bcl-2) in HEI-OC1 cells, and this effect was reversed by treatment with rutin (Fig. 4g).
Rutin Inhibits the Necroptosis Signaling Pathway in HEI-OC1 Cells
To further elucidate the mechanism of rutin, proteomics combined with bioinformatics analyses were conducted. As illustrated in Figs. 4a and 389 proteins were differentially expressed after treatment with rutin in HEI-OC1 cells, of which 146 proteins were significantly upregulated and 243 were downregulated (Fig. 5a). Notably, the results of GO analysis showed that various biological processes related to immune response were inhibited compared to controls, such as leukocyte activation involved in immune response (GO:0002366), cell activation involved in immune response (GO:0002263), immune effector process (GO:0002252) and so on (Fig. 5b).

Quantitative proteomics analysis revealing the mechanism by which rutin mediates hair cell death. Proteomics analysis of rutin-treated HEI-OC1 cells compared with control. (a) Volcano plots of differential protein abundances between the rutin-treated groups and the control group. (b) GO enrichment analysis of differentially expressed proteins. (c) KEGG enrichment analysis of differentially expressed proteins. (d) Pathway diagram of cell necroptosis. Green represents downregulated genes, and red denotes upregulated genes
Next, KEGG analysis determined that rutin inhibited several signaling pathways related to cell death, such as necroptosis, apoptosis, and cellular senescence (Fig. 5c). Following this, the necroptosis signaling pathway was investigated to identify potential target proteins of rutin. As anticipated, rutin treatment significantly down-regulated multiple proteins associated with the necroptosis signaling pathway compared to controls, including signal transducer and activator of transcription 1 (STAT1), apoptosis regulator BAX (BAX), ferritin heavy chain (FTH1), apoptosis-associated speck-like protein containing a CARD, and apoptosis-associated speck-like protein containing a CARD, heavy chain (FTH1), apoptosis-associated speck-like protein containing a CARD (ASC), caspase 1, and others (Fig. 5d). Among these, STAT1 was located upstream of the necroptosis signaling pathway.
Rutin Protects hair Cells from GM-induced Ototoxicity by Targeting STAT1
To evaluate the affinity of the rutin for its target, molecular docking analysis was performed. The binding poses and interactions of rutin with STAT1 were determined using Autodock Vina v.1.2.2, and binding energy for each interaction was generated. The results showed that rutin was bound to its protein targets, and the hydrophobic pocket of STAT1 was successfully occupied by rutin. In other words, rutin had a low binding energy of -8.391 kcal/mol, indicating highly stable binding (Fig. 6a).

Rutin protected hair cells by inhibiting the necroptosis pathway. (a) Binding mode of rutin to the target (STAT1) via molecular docking. Cartoon representation and overlay of the crystal structures of small molecule compounds and their targets were illustrated using the Molecule of the Month feature. (b) 5 dpf zebrafish were incubated with 200 µmol/L GM for 12 h or initially treated with 400 µmol/L rutin or a STAT1 agonist, namely 2-NP (200 µmol/L), for 3 h. The expression level of necroptosis pathway-related genes in zebrafish was determined via quantitative real-time PCR. (c) Quantification of the number of surviving cilia per neuromast after different treatments, expressed as mean ± SD (n = 10). ****P < 0.0001. (d) HEI-OC1 cells were incubated with 80 µmol/L GM for 24 h or initially treated with 100 µmol/L rutin or 2-NP (200 µmol/L) for 6 h. The expression level of necroptosis pathway-related genes in mice was determined via quantitative real-time PCR. (e) Flow cytometry analysis of HEI-OC1 cell apoptosis using Annexin V-PE/7-AAD staining after co-incubation with GM, rutin, or 2-NP. (f) Quantification of flow cytometric analyses. The Student’s t-test was used for the comparison; P < 0.01 (**), P < 0.001 (***), and P < 0.001 (****); ns: no significant difference. Each experiment was performed in triplicate
To further validate that STAT1 is a target of rutin, STAT1 activator 2-NP was employed, and the mRNA levels of necroptosis- (stat1b or STAT1, caspa or caspase‐1, and Bax) related genes were quantified via qPCR both in vivo and vitro. The results showed that genes related to programmed necrosis were significantly up-regulated after exposure of zebrafish or HEI-OC1 cells to GM, and this up-regulation was reversed after pretreatment with rutin. Moreover, the down-regulation of necroptosis-related genes by rutin was significantly attenuated by the introduction of 2-NP to embryo culture water or cell culture medium (Fig. 6b, d). Similarly, the protective effect of rutin on the hair cells of zebrafish was significantly suppressed by the addition of 2-NP treatment (Fig. 6c).
Finally, flow cytometry was performed to assess cell apoptosis. The results unveiled that 2-NP attenuated the inhibitory effect of rutin on GM-induced apoptosis in HEI-OC1 cells (Fig. 6e, f).
Rutin improves the bacteriostatic effect of GM
To investigate whether rutin interfered with the efficacy of GM, we performed the disc susceptibility tests of Escherichia coli, Klebsiella pneumoniae, and Salmonella enterica (Fig. 7a). The results showed that rutin at any concentration (30–200 µM/L) significantly increased the bacteriostatic effect of GM against gram-negative bacteria (Fig. 7b-d). Specifically, rutin showed the best antibacterial effect against Escherichia coli and Klebsiella pneumoniae at a concentration of 100 µM/L, and Salmonella at a concentration of 30 µM/L, respectively.

Rutin improves the bacteriostatic effect of GM. (a) Typical images of bacteriostatic drug sensitivity experiments under GM alone or co-incubation with rutin. (b-d) Quantitative analysis of inhibitory ring diameter under different treatments. The Student’s t-test was used for the comparison; P < 0.01 (**), P < 0.001 (***), and P < 0.001 (****); ns: no significant difference. Each experiment was performed in triplicate
Discussion
The high homology between the zebrafish genome and the human genome, as well as the biological characteristics of zebrafish, make it an ideal model animal for modeling various human diseases [18]. The inner ear of the zebrafish possesses vestibular and auditory functions, such as marching through the water to maintain balance, sensing the vibrations of water waves, catching prey, or avoiding danger [19]. The organ that receives these stimuli is called the neuromast, which comprises a ring of supporting cells surrounding a cluster of sensory hair cells. The zebrafish neuromast hair cells are structurally and functionally comparable to mammalian inner ear hair cells. The neuromasts are located in the zebrafish ear sac and the lateral line of the body surface and can be observed under a microscope [20]. In the current study, zebrafish were exposed to GM in order to explore the protective effect of rutin on hair cells.
The Startle Response Assay used to assess zebrafish hearing involves exposing the fish to sudden sound stimuli to observe their startled reaction. This assay takes advantage of zebrafish’s rapid response to abrupt sound stimuli, often manifested as a sudden burst of swimming or a freeze response [21]. We found that rutin significantly improved hearing behavior in zebrafish. In here, we used the startle response assay to examine zebrafish hearing. And, the results collectively signaled that rutin could significantly attenuate GM toxicity in zebrafish hair cells.
To further corroborate this result, in vitro experiments were conducted using hair cell-like HEI-OC1 cells. HEI-OC1 is a widely used progenitor hair cell line derived from mouse auditory organs and has been extensively used in earlier studies to investigate the protective mechanism of hair cells [22]. The results unveiled that rutin could effectively protect HEI-OC1 cells from GM-induced ototoxicity. These findings conjointly suggest that rutin is a promising otoprotective agent for routine use against GM-induced ototoxicity.
In addition, prior studies have documented that rutin exerts antibacterial effects by inhibiting cytoplasmic membrane function, bacterial cell wall synthesis, and nucleic acid synthesis [23]. Therefore, the experiments are necessitated to confirm the influence of rutin on the efficacy of GM while protecting hair cells. Fortunately, we have found that rutin can be very effective in enhancing the bacteriostatic effect of GM against gram-negative bacteria. This suggests that the combination of rutin and GM is well worth trying in clinical applications: Combination therapy with rutin and GM may lead to more effective eradication of bacterial infections compared to GM monotherapy. This can be particularly beneficial in treating multidrug-resistant bacterial strains or chronic infections that are difficult to eradicate with conventional antibiotics alone. Rutin’s ability to enhance the bacteriostatic effect of GM may help reduce the development of antibiotic resistance, thereby preserving the effectiveness of gentamicin for longer durations and reducing the risk of treatment failure due to resistant bacterial strains. Combination therapy with rutin may allow for the use of lower dosages of GM while still achieving therapeutic efficacy. This can help minimize the risk of GM-related side effects and toxicity, which are often dose-dependent.
Next, potential molecular mechanisms underlying the protective effect of rutin were explored. According to a study, rutin enhances fibroblast proliferation at a low concentration and has very low cytotoxicity at high concentrations [24]. In this study, the cytotoxicity of rutin towards either zebrafish hair cells or HEI-OC1 cells was minimal. On the contrary, rutin significantly increased the number of proliferating cells in zebrafish lateral line neuromasts and facilitated the proliferation of HEI-OC1 cells, insinuating that it may assist in stimulating the proliferation of hair cell precursor cells and, in turn, promoting hair cell regeneration. The generation of human hair cell progenitors poses a major challenge in sensory hair cell regeneration research [25]. Therefore, more cases are required in future studies to confirm this generalization.
The permanent hearing loss caused by aminoglycosides has been hypothesized to be predominantly associated with the apoptotic death of outer hair cells [26]. Upon exposure to gentamicin, there is an increase in reactive oxygen species (ROS) within the inner ear cells, leading to oxidative stress [27]. Elevated ROS levels can cause damage to cellular components such as lipids, proteins, and DNA, ultimately resulting in hair cell dysfunction and death. Furthermore, GM can trigger necrotic or apoptotic pathways in inner ear hair cells. This may involve the activation of specific signaling pathways that lead to cell death, such as the JNK (c-Jun N-terminal kinase) pathway or the caspase cascade. Necrotic cell death is characterized by cellular swelling and rupture, while apoptotic cell death involves a series of biochemical events leading to cell shrinkage and fragmentation [28,29,30]. In this study, CCK-8, Tunel staining, flow cytometry, and qPCR assays determined that rutin inhibits GM-induced apoptosis in hair cells. And, herein, rutin alleviated GM-induced ROS production in HEI-OC1 cells.
Inhibition of necroptosis and apoptosis using pharmacological interventions is a viable strategy to ameliorate aminoglycoside-induced damage [31]. Necroptosis is a pro-inflammatory mode of programmed cell death that is hallmarked by the intentional disruption of host membranes and the release of pro-inflammatory cytosolic components into the milieu [14, 23]. According to the results of the KEGG analysis on proteomics, rutin may protect hair cells from GM-induced ototoxicity by modulating the necroptosis pathway, a process mediated by STAT1. Here, drawing from the findings of our proteomic analysis, we posited that STAT1 could be a target of rutin. Subsequently, we conducted molecular docking studies, revealing a strong affinity between STAT1 and rutin. This evidence supports the notion that STAT1 may serve as a direct target of rutin, albeit to a certain extent [16]. Meanwhile, in vivo and in vitro experiments exposed that STAT1 may be a direct target of rutin. It should be noted that in pharmacological experiments, a drug typically does not act on only one target; rather, its efficacy may result from the combined action of multiple molecules and pathways. Therefore, the strengthened evidence presented in this study can only suggest that STAT1 is one of the significant targets of rutin in protecting against ototoxicity. This protein emerges as a promising target for the prevention and management of ototoxicity induced by aminoglycosides.
It should be noted that this study solely demonstrates rutin’s ability to inhibit STAT1 to exert its protective effect. However, further exploration is warranted to elucidate the specific molecular pathways and mechanisms underlying subsequent activation. In the cellular apoptotic signaling pathway, STAT1 plays a crucial regulatory role. Upon cellular stimulation by external factors such as viral infection or cellular damage, signaling molecules like interferons induce the phosphorylation and activation of STAT1. Activated STAT1 forms dimers and translocates to the nucleus, where it binds to specific DNA sequences, regulating the transcription of target genes including apoptotic factors such as Bax and Caspases. Increased expression of these factors triggers the apoptotic signaling pathway within the cell, leading to cell apoptosis. Additionally, activated STAT1 can also cross-talk with other signaling pathways such as NF-κB and JAK-STAT, further modulating cell survival and death decisions [32,33,34]. Therefore, STAT1 plays a critical regulatory role in apoptotic signaling, exerting significant influence over the balance between cell survival and death. Additionally, the binding activity between rutin and STAT1 needs further experimental validation, such as drug-protein co-precipitation, drug-protein immunoprecipitation, surface plasmon resonance, and other [35].
Finally, it is worthwhile emphasizing that mammalian models were not used in this study to evaluate the ototoxic antagonism of rutin. Zebrafish, unlike mice, do not possess a complex cochlear structure and blood-cochlear barrier, resulting in either ototoxic or protective drugs easily impacting their surface hair cells [36,37,38,39,40,41]. Therefore, additional experiments using mammalian models are needed in the future to explore the potential of rutin as a hair cell protector and determine the optimal dose.
Conclusion
To the best of our knowledge, this is the first study to establish the protective effect of rutin against GM-induced ototoxicity. Taken together, this study provides compelling evidence that rutin may enhance hair cell survival by inhibiting STAT1-dependent apoptotic or necroptosis.
Data Availability
No datasets were generated or analysed during the current study.
References
Selimoglu E (2007) Aminoglycoside-induced ototoxicity. Curr Pharm Des 13(1):119–126. https://doi.org/10.2174/138161207779313731
Steyger PS (2021) Mechanisms of Ototoxicity and Otoprotection. Otolaryngol Clin North Am 54(6):1101–1115. https://doi.org/10.1016/j.otc.2021.08.007
Kros CJ, Steyger PS (2019) Aminoglycoside- and Cisplatin-Induced Ototoxicity: mechanisms and otoprotective strategies. Cold Spring Harb Perspect Med 9(11):a033548. https://doi.org/10.1101/cshperspect.a033548
Wargo KA, Edwards JD (2014) Aminoglycoside-induced nephrotoxicity. J Pharm Pract 27(6):573–577. https://doi.org/10.1177/0897190014546836
Lanvers-Kaminsky C, Ciarimboli G (2017) Pharmacogenetics of drug-induced ototoxicity caused by aminoglycosides and cisplatin. Pharmacogenomics 18(18):1683–1695. https://doi.org/10.2217/pgs-2017-0125
Steyger PS (2021) Mechanisms of Aminoglycoside- and Cisplatin-Induced Ototoxicity. Am J Audiol 30(3S):887–900. https://doi.org/10.1044/2021_AJA-21-00006
Zong Y, Chen F, Li S, Zhang H (2021) (-)-Epigallocatechin-3-gallate (EGCG) prevents aminoglycosides-induced ototoxicity via anti-oxidative and anti-apoptotic pathways. Int J Pediatr Otorhinolaryngol 150:110920. https://doi.org/10.1016/j.ijporl.2021.110920
Ghorbani A (2017) Mechanisms of antidiabetic effects of flavonoid rutin. Biomed Pharmacother 96:305–312. https://doi.org/10.1016/j.biopha.2017.10.001
Negahdari R, Bohlouli S, Sharifi S, Maleki Dizaj S, Rahbar Saadat Y, Khezri K, Jafari S, Ahmadian E et al (2021) Therapeutic benefits of rutin and its nanoformulations. Phytother Res 35(4):1719–1738. https://doi.org/10.1002/ptr.6904
Kamalakkannan N, Stanely Mainzen Prince P (2006) Rutin improves the antioxidant status in streptozotocin-induced diabetic rat tissues. Mol Cell Biochem 293(1–2):211–219. https://doi.org/10.1007/s11010-006-9244-1
Nafees S, Rashid S, Ali N, Hasan SK, Sultana S (2015) Rutin ameliorates cyclophosphamide induced oxidative stress and inflammation in Wistar rats: role of NFκB/MAPK pathway. Chem Biol Interact 231:98–107. https://doi.org/10.1016/j.cbi.2015.02.021
Zheng S, Liu C, Tang D, Zheng Z, Yan R, Wu C, Zuo N, Ma J et al (2022) The protective effect of rutin against the cisplatin-induced cochlear damage in vitro. Neurotoxicology 90:102–111. https://doi.org/10.1016/j.neuro.2022.03.005
Chitnis AB, Nogare DD, Matsuda M (2012) Building the posterior lateral line system in zebrafish. Dev Neurobiol 72(3):234–255. https://doi.org/10.1002/dneu.20962
Park MK, Lee BD, Chae SW, Chi J, Kwon SK, Song JJ (2012) Protective effect of NecroX, a novel necroptosis inhibitor, on gentamicin-induced ototoxicity. Int J Pediatr Otorhinolaryngol 76(9):1265–1269. https://doi.org/10.1016/j.ijporl.2012.05.016
Tripathi S, Parmar D, Fathima S, Raval S, Singh G (2023) Coenzyme Q10, Biochanin A and Phloretin Attenuate Cr(VI)-Induced oxidative stress and DNA damage by stimulating Nrf2/HO-1 pathway in the experimental model. Biol Trace Elem Res 201(5):2427–2441. https://doi.org/10.1007/s12011-022-03358-5
Stolzer I, Schickedanz L, Chiriac MT, López-Posadas R, Grassl GA, Mattner J, Wirtz S, Winner B et al (2022) STAT1 coordinates intestinal epithelial cell death during gastrointestinal infection upstream of Caspase-8. Mucosal Immunol 15(1):130–142. https://doi.org/10.1038/s41385-021-00450-2
Gale JE, Marcotti W, Kennedy HJ, Kros CJ, Richardson GP (2001) FM1-43 dye behaves as a permeant blocker of the hair-cell mechanotransducer channel. J Neurosci 21(18):7013–7025. https://doi.org/10.1523/JNEUROSCI.21-18-07013.2001
Baek S, Tran NTT, Diaz DC, Tsai YY, Acedo JN, Lush ME, Piotrowski T (2022) Single-cell transcriptome analysis reveals three sequential phases of gene expression during zebrafish sensory hair cell regeneration. Dev Cell 57(6):799–819e6. https://doi.org/10.1016/j.devcel.2022.03.001
Nicolson T (2017) The genetics of hair-cell function in zebrafish. J Neurogenet 31(3):102–112. https://doi.org/10.1080/01677063.2017.1342246
Kniss JS, Jiang L, Piotrowski T (2016) Insights into sensory hair cell regeneration from the zebrafish lateral line. Curr Opin Genet Dev 40:32–40. https://doi.org/10.1016/j.gde.2016.05.012
Pittman JT, Lott CS (2014) Startle response memory and hippocampal changes in adult zebrafish pharmacologically-induced to exhibit anxiety/depression-like behaviors. Physiol Behav 17:123:174–179. https://doi.org/10.1016/j.physbeh.2013.10.023
Zheng Z, Tang D, Zhao L, Li W, Han J, Hu B, Nie G, He Y (2020) Liproxstatin-1 protects Hair Cell-Like HEI-OC1 cells and cochlear hair cells against Neomycin Ototoxicity. Oxid Med Cell Longev 2020:1782659. https://doi.org/10.1155/2020/1782659
Ivanov M, Novović K, Malešević M, Dinić M, Stojković D, Jovčić B, Soković M (2022) Polyphenols as Inhibitors of Antibiotic Resistant Bacteria-Mechanisms Underlying Rutin Interference with Bacterial Virulence. Pharmaceuticals (Basel) 15(3):385. https://doi.org/10.3390/ph15030385
Tabolacci E, Tringali G, Nobile V, Duca S, Pizzoferrato M, Bottoni P, Clementi ME (2023) Rutin protects fibroblasts from UVA Radiation through Stimulation of Nrf2 pathway. Antioxid (Basel) 12(4):820. https://doi.org/10.3390/antiox12040820
Shu Y, Li W, Huang M, Quan YZ, Scheffer D, Tian C, Tao Y, Liu X et al (2019) Renewed proliferation in adult mouse cochlea and regeneration of hair cells. Nat Commun 10(1):5530. https://doi.org/10.1038/s41467-019-13157-7
He ZH, Li M, Fang QJ, Liao FL, Zou SY, Wu X, Sun HY, Zhao XY et al (2021) FOXG1 promotes aging inner ear hair cell survival through activation of the autophagy pathway. Autophagy 17(12):4341–4362. https://doi.org/10.1080/15548627.2021.1916194
He Z, Guo L, Shu Y, Fang Q, Zhou H, Liu Y, Liu D, Lu L et al (2017) Autophagy protects auditory hair cells against neomycin-induced damage. Autophagy 13(11):1884–1904. https://doi.org/10.1080/15548627.2017.1359449
Ruhl D, Du TT, Wagner EL, Choi JH, Li S, Reed R, Kim K, Freeman M et al (2019) Necroptosis and apoptosis contribute to Cisplatin and Aminoglycoside Ototoxicity. J Neurosc 39(15):2951–2964. https://doi.org/10.1523/JNEUROSCI.1384-18.2019
Gitika B, Sai Ram M, Sharma SK, Ilavazhagan G, Banerjee PK (2006) Quercetin protects C6 glial cells from oxidative stress induced by tertiary-butylhydroperoxide. Free Radic Res 40(1):95–102. https://doi.org/10.1080/10715760500335447
Singh G, Thaker R, Sharma A, Parmar D (2021) Therapeutic effects of biochanin A, phloretin, and epigallocatechin-3-gallate in reducing oxidative stress in arsenic-intoxicated mice. Environ Sci Pollut Res Int 28(16):20517–20536. https://doi.org/10.1007/s11356-020-11740-w
Tripathi S, Fhatima S, Parmar D, Singh DP, Mishra S, Mishra R, Singh G (2022) Therapeutic effects of CoenzymeQ10, biochanin A and Phloretin against arsenic and chromium induced oxidative stress in mouse (Mus musculus) brain. 3 Biotech 12(5):116. https://doi.org/10.1007/s13205-022-03171-w
Kim HS, Lee MS (2007) STAT1 as a key modulator of cell death. Cell Signal 19(3):454–465. https://doi.org/10.1016/j.cellsig.2006.09.003
Wei TT, Zhang MY, Zheng XH, Xie TH, Wang W, Zou J, Li Y, Li HY et al (2022) Interferon-γ induces retinal pigment epithelial cell ferroptosis by a JAK1-2/STAT1/SLC7A11 signaling pathway in Age-related Macular Degeneration. FEBS J 289(7):1968–1983. https://doi.org/10.1111/febs.16272
Murata H, Yasui Y, Oiso K, Ochi T, Tomonobu N, Yamamoto KI, Kinoshita R, Sakaguchi M (2023) STAT1/3 signaling suppresses axon degeneration and neuronal cell death through regulation of NAD+-biosynthetic and consuming enzymes. Cell Signal 108:110717. https://doi.org/10.1016/j.cellsig.2023.110717
Schmitt NC, Rubel EW, Nathanson NM (2009) Cisplatin-induced hair cell death requires STAT1 and is attenuated by epigallocatechin gallate. J Neurosci 29(12):3843–3851. https://doi.org/10.1523/JNEUROSCI.5842-08.2009
Zhang QX, He XJ, Wong HC (2016) Kindt KS (2016) Functional calcium imaging in zebrafish lateral-line hair cells. Methods Cell Biol. 133:229–52. https://doi.org/10.1016/bs.mcb.2015.12.002
Barta CL, Liu H, Chen L, Giffen KP, Li Y, Kramer KL, Beisel KW, He DZ (2018) RNA-seq transcriptomic analysis of adult zebrafish inner ear hair cells. Sci Data 5:180005. https://doi.org/10.1038/sdata.2018.5
Hung GY, Wu CL, Chou YL, Chien CT, Horng JL, Lin LY (2019) Cisplatin exposure impairs ionocytes and hair cells in the skin of zebrafish embryos. Aquat Toxicol 209:168–177. https://doi.org/10.1016/j.aquatox.2019.02.006
Li M, Liu J, Liu D, Duan X, Zhang Q, Wang D, Zheng Q, Bai X et al (2021) Naringin attenuates cisplatin- and aminoglycoside-induced hair cell injury in the zebrafish lateral line via multiple pathways. J Cell Mol Med 25(2):975–989. https://doi.org/10.1111/jcmm.16158
Li M, Zhao J, Tang Q, Zhang Q, Wang Y, Zhang J, Hao Y, Bai X et al (2021) Lamivudine improves cognitive decline in SAMP8 mice: integrating in vivo pharmacological evaluation and network pharmacology. J Cell Mol Med 25(17):8490–8503. https://doi.org/10.1111/jcmm.16811
Li M, Zhang J, Chen W, Liu S, Liu X, Ning Y, Cao Y, Zhao Y (2023) Supraphysiologic doses of 17β-estradiol aggravate depression-like behaviors in ovariectomized mice possibly via regulating microglial responses and brain glycerophospholipid metabolism. J Neuroinflammation 20(1):204. https://doi.org/10.1186/s12974-023-02889-5
Acknowledgements
This research was funded by the Qingdao Medical and Health Research guidance Project (2022-WJZD107) and the National Natural Science Foundation of China (Nos. 82000979).
Funding
This research was funded by the Qingdao Medical and Health Research guidance Project (2022-WJZD107) and the National Natural Science Foundation of China (Nos. 82000979).
Author information
Authors and Affiliations
Contributions
HY and RY designed the experiments; HY performed the animal experiments; TZ, JL, and DW performed the experiments; KG, HY and WZ analyzed the data, reviewed the data, advised the study, and interpreted the analyzed data; HY and TX wrote the manuscript. The manuscript has been revised and approved by all authors.
Corresponding authors
Ethics declarations
Ethics Approval
This study was approved by the Animal Ethics Committee (No.20,230,506) of Shandong Provincial Hospital (Jinan, China).
Consent to Participate
Not applicable.
Consent for publication
All authors have read and approve the manuscript and consent to its publication.
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Yang, H., Zong, T., Liu, J. et al. Rutin Attenuates Gentamycin-induced Hair Cell Injury in the Zebrafish Lateral Line via Suppressing STAT1. Mol Neurobiol (2024). https://doi.org/10.1007/s12035-024-04179-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12035-024-04179-4