
Abstract
Nonylphenol (NP), a well-known endocrine-disrupter chemical, has several harmful effects on the central nervous system including neuroendocrine disruption, cognitive impairment, and neurotoxicity. Thymoquinone (TQ) is a main bioactive compound in the black seeds of Nigella sativa that has antioxidant, anti-inflammatory, and neuroprotective properties. Here, we investigated the neuroprotective effect of TQ against NP-induced memory deficit and neurotoxicity in rats. To induce memory impairment, NP (25 mg/kg) was used as gavage in male Wistar rats for 21 days. TQ (2.5, 5, and 10 mg/kg) was intraperitoneally administered in NP-treated animals. The morris water maze test was performed to assess spatial learning and memory. The hippocampal tissues were isolated from the brain for histopathological evaluation. Biochemical, molecular, and cellular tests were performed to quantify oxidant (malondialdehyde; MDA)/antioxidant (superoxide dismutase (SOD), total antioxidant capacity (TAC), and reduced glutathione (GSH) parameters) as well as markers for astrocytic activation (glial fibrillary acidic protein; GFAP) and neuronal death (alpha-synuclein; α-syn). Results showed TQ (5 mg/kg) significantly improved NP-induced memory impairment. Histological data revealed a significant increase in the number of necrotic cells in hippocampus, and TQ treatment markedly decreased this effect. The GSH and TAC levels were significantly increased in TQ-treated groups compared to NP group. The molecular analysis indicated that NP increased GFAP and decreased α-syn expression and TQ treatment did the reverse. In vitro study in astrocytes isolated from mice brain showed that TQ significantly increased cell viability in NP-induced cytotoxicity. This study strongly indicates that TQ has neuroprotective effects on NP-induced neurotoxicity through reducing oxidative damages and neuroinflammation.
Graphical abstract
This study investigates the behavioral neurotoxicity induced by Nonylphenol (NP) and the protective effects of Thymoquinone (TQ) as a potent antioxidant compound using molecular, cell culture, histopathological and biochemical techniques.

Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Endocrine-disrupting chemicals (EDCs) are considered a main concern to public health and their potential function on general health has been extensively discussed for many years in science and the public [1]. Adverse effects of EDCs have been shown on memory, cognition, social behavior, and endocrine system [2]. Nonylphenol (NP) is a member of the alkylphenol class and classified as EDCs [3]. NP is found in various products including cosmetics, detergents, pesticides, emulsifiers, and paints[4]. NP is highly toxic to various organs, including liver, pancreas, thyroid, kidneys, eyes, as well as reproductive and respiratory systems [5,6,7,8]. There is evidence that also show deleterious effect of NP on the brain function, with impairment in cognitive function and neurogenesis, and deficits in learning and memory [9]. After ingestion, NP enters the bloodstream and easily passes through the blood–brain barrier (BBB) due to its lipophilic structure [10]. Several mechanisms could be involved in NP neurotoxicity including inflammation [11], oxidative stress [12], gliosis, and neural loss [13]. Previous study indicated that oral administration of NP at dose of 25 mg/kg induced neurotoxicity and learning and memory impairment accompanied by oxidative stress, hippocampal tissue injury, and increased inflammatory and apoptotic-related gene expression [14]. In another study, NP dysregulated the balance of oxidant/antioxidant by reducing the levels of superoxide dismutase (SOD), catalase (CAT), and reduced glutathione (GSH) in the brain [15]. Moreover, exposure to NP was found to stimulate glial cells in the central nervous system and increase inflammatory factors in these cells [16].
Natural compounds with antioxidant and anti-inflammatory properties have attracted considerable attention as potential therapeutic targets for several neurological diseases [17]. Nigella sativa, also known as black cumin, nigella, or kalonji, is an ancient Indian medicine, Ayurveda that is widely used for various ailments treatment including asthma, headache, bronchitis, infection, hypertension, digestive problems, and eczema in Africa and the Middle East [18, 19]. Nigella sativa has also been used for centuries in ancient Indian medicine, Ayurveda, and is still used for various ailments. Thymoquinone (TQ), as a main bioactive constituent of the black seed (Nigella sativa), has anti-inflammatory [20], antioxidant, anti-neurodegenerative, and anti-tumor properties [21]. Under normal conditions, TQ acts as a potent antioxidant and reduces the production of superoxide radicals and lipid peroxidation. It also increases the activity of the antioxidant enzymes, such as glutathione reductase (GR), GSH, CAT, and SOD [22]. Several studies have demonstrated that TQ prevents neuronal apoptosis induced by ethanol cytotoxicity in primary neurons and protects PC12 cells against cytotoxic oxidative injury [23]. Moreover, TQ improved learning and memory function in Alzheimer’s disease animal models via its anti-neurodegenerative properties [24]. A study reported that TQ at doses of 5 and 10 mg/kg can suppress neuroinflammation in BV2 microglia and mixed glial cells [25]. In addition, TQ at doses of 2.5, 5, and 10 mg/kg has been shown to improve lipopolysaccharide (LPS)-induced learning and memory deficit through decreasing inflammatory cytokine levels and oxidative damage [26]. TQ was also found to mitigate dopaminergic neurodegeneration by reducing oxidative stress in a rodent model of Parkinson’s disease [27].
Although recent studies suggest that antioxidant compounds reduce NP toxicity, there is still no safe and definitive solution to prevent and treat NP neurotoxicity. Thus, this study was designed to assess the neuroprotective effects of TQ on memory impairment and neurotoxicity induced by NP in vitro and in vivo. First, isolation and culture of astrocyte from the mouse brain were performed and then characterized with glial fibrillary acidic protein (GFAP), glutamate aspartate transporter (GLAST), and calcium-binding protein B (S100-β) antibodies by immunofluorescence technique. Moreover, the MTS test evaluated the effects of different doses of TQ and NP on astrocyte cell viability. Spatial learning and memory function was assessed by the morris water maze (MWM) task. Furthermore, histological staining (H&E and NISSL) was used to evaluate the hippocampal injuries. The biochemical assessment measured oxidant parameters (malondialdehyde; MDA), antioxidant parameters (enzymatic antioxidants including SOD and total antioxidant capacity (TAC)), and non-enzymatic antioxidant parameters including GSH. Finally, GFAP and alpha-synuclein (α-syn) gene expression analysis was performed by qPCR.
Material and Methods
Chemicals
Thymoquinone (TQ), Nonylphenol (NP), Dulbecco’s Modified Eagle Medium (DMEM), Fetal bovine serum (FBS), Penicillin/Streptomycin, non-essential amino acid, l-glutamate, Hank’s balanced salt solution (HBSS), Poly-l-ornithine (PLO), Diethyl ether, Trypsin, 4% paraformaldehyde, Triton X-100, PBS, MTS kit, were obtained from Sigma-Aldrich Chemical (USA). Ketamine and Xylazine were purchased from Alfasan (Woerden Co., Netherlands). Primary antibodies GFAP (Sigma-Aldrich, G3893, 1/200), GLAST (Abcam, Ab416, 1/200), S100β (Abcam, Ab52642, 1/200) were used in our study.
Cell Culture
Astrocyte Isolation from Wild-Type NMRI Mice
The astrocyte cell was isolated from neonate mice vehicle (wild-type NMRI) aged 5–7 days. The astrocyte cell culture media (DMEM, high glucose + 1% Penicillin/Streptomycin + 15% fetal bovine serum + 1% non-essential amino acid + 1% l-glutamate), as well as cold HBSS (Hank’s balanced salt solution) with 1% Penicillin/Streptomycin for isolation medium, were prepared before the procedure. Poly-l-ornithine (PLO, 0.001%, Sigma-Aldrich, P4707) was used to coat T25 flasks for one day at 37 °C in the CO2 incubator.
Then, mice were euthanized, and their brains were extracted and put on ice in a dish filled with HBSS/Penicillin/ Streptomycin. The cerebellum and olfactory bulbs were removed and placed into another dish filled with HBSS to remove the meninges of the cortex. Now, in the third dish, the tissues were cut into small pieces by sharp blades. The fragments were then moved to a 50 ml Falcon tube containing HBSS/0.5% trypsin, combined, and incubated for about 7–10 min in a water bath at 37 °C. The suspension was then passed through a 100 M cell strainer after incubation. The pieces of cortex tissue were centrifuged at 1500 × g for 5 min. Then, the supernatant was eliminated, and the tissue was dissociated into a single cell suspension by adding 10 ml of astrocyte media and then intensive pipetting 20–30 times. The cell suspension was then seeded in a T25 culture flask and incubated in a CO2 incubator at 37 °C. The primary cell culture medium was changed 4 days after plating and every 2 days after that. After 7–8 days, MG was extracted by shaking the cells at 240 rpm for 3 h. After shaking, the culture medium was changed, and the cells were incubated at 37 °C in the CO2 incubator. On day 14, the cells were shaken at 180 rpm overnight to separate oligodendrocytes and were then dissociated using 0.5% trypsin. FBS was used to inactivate the trypsin, and the cells were centrifuged at 1500 g for 5 min before being seeded on T25 culture flasks [28].
Characterization of Astrocytes
Cell cultures were fixed for 15–20 min with 4% paraformaldehyde (Sigma-Aldrich, P6148) and permeabilized, or with 0.5% Triton X-100 (Sigma-Aldrich, T8532) in PBS for 30 min at room temperature (RT). The cells were incubated with primary antibodies in blocking solution overnight at 4 °C, and then secondary antibodies (1:1000; Invitrogen) were applied for 1 h in RT. Then to visualize the nuclei, cells were stained for 5 min with 49, 69-diamidino-2-phenylindole (DAPI, Sigma-Aldrich, D-8617) and observed under Olympus IX71 microscope (Table 1).
Assessment of the Astrocyte Viability by MTS
MTS assay (Promega, G5421) was used to study the effects of different doses of NP, TQ and combination of NP and TQ on cell viability as previously described [29]. Astrocytes were cultured in a 96-well culture plate at a density of 1 × 104 cells per well. After being cultured for 24 h, the cells were treated for another 24 h with different concentrations of NP (0.25, 0.5, 1, 10, 20, 40, 80, 100 µM), TQ (2.5, 5, 7.5, 10, 12.5, 15, 17.5, 20 µM) and combination of NP (40, 80 μM) and TQ (2.5, 5, 7.5, 10 µM). At the end of treatment, 20 μl MTS solution was added to each well, and the plates were incubated at 37 °C for 3 h. On the microplate reader, the absorbance values of each well were then determined at 490 nm.
In Vivo Experiments
Animals
Male Adult Wistar rats (150–200 g) were obtained from central animal house, Babol University of medical science, Babol, Iran. All animals were cared under standard conditions (temperature (22 ± 2 °C), humidity (60%), and a 12 h light/dark cycle), and food and water were freely accessed. All experimental protocols were according to the international guidelines for care and use of laboratory animals and approved by the ethical committee of Babol university committee (IR.MUBABOL.REC.1398.026).
Experimental Design
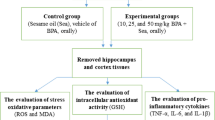
Rats were randomly divided into six groups and the treatment duration for NP and TQ was 21 days: (1) vehicle (vehicle + soy oil): soy oil as an NP and TQ vehicle was administrated orally from the first day of study till day 21. (2) NP: rats received 25 mg/kg NP by gavage, (3) NP + TQ (2.5 mg/kg): rats were intraperitoneally (i.p) injected with 2.5 mg/kg TQ after gavage of NP (25 mg/kg), (4) NP + TQ (5 mg/kg): rats were i.p injected with 5 mg/kg of TQ after gavage of NP (25 mg/kg), (5) NP + TQ (10 mg/kg): rats were i.p injected with 10 mg/kg TQ after gavage of NP (25 mg/kg), (6) TQ (10 mg/kg): rats were the only i.p injected with 10 mg/kg TQ. The doses of NP (25 mg/kg) [14], and TQ (2.5, 5, 10 mg/kg) [30] were selected based on previous studies. NP and TQ were freshly prepared by dissolving NP and TQ in soy oil in volume 0.5 ml. At the end of the 21 days, the MWM behavioral test evaluated learning and memory function. At the end of the study, the rats were fully anesthetized with ketamine/xylazine, and half of the rat population in each group was perfused for histopathological study. Brain tissue of the other half was stored in the freezer − 80 °C for biochemical and molecular study.
Morris Water Maze
The morris water maze (MWM) is commonly used as an accurate behavioral test to assess rodent spatial learning functions [31]. The MWM test was performed to determine the effect of TQ treatment on spatial learning and memory impairment at the end of the 21 days. All experimental trials were done at the same time of the afternoon. Shortly, animals were kept in the MWM test environment for 24 h before the experiments for adaptation and then were trained four trials (times) a day for four consecutive days. There were at least 5-min intervals between two successive trials. The animal was allowed to find a hidden platform (Q4) in 60 s. The swimming escape latency (time spent to find the hidden platform) parameter was analyzed to evaluate the spatial learning. Swimming velocity was used to evaluate the motor activity of the animals in the behavioral experiment. To evaluate the spatial memory on probe trial (day 5), each rat was let to swim for 60 s in the maze without the platform. We used the duration in the target zone (Q4) to assess spatial memory function in all experimental groups. To record and analysis the MWM data, the Noldus Ethovision XT (11) software (Netherlands) was used.
Sample Preparation
To prepare serum, the first 1 ml of blood was collected from each animal without anticoagulants. Then, it was allowed to stay at 25° C for 10 min. Blood samples were centrifuged at 3000 × g for 10 min, and the serum was separated.
To prepare supernatant from brain tissue, 2.7 ml of 150 mM KC1 was added into 300 mg of wet tissue in a 2 ml centrifuge tube. It was then homogenized on ice using a Teflon-Potter–Elvehjem glass or homogenizer and centrifuged at 1600 g of for 10 min at 4 °C. The supernatant was separated and was used in biochemical assays.
Biochemical Assay
To measure biochemical parameters, serum and brain supernatant were used. The levels of MDA, SOD, TAC, and GSH were measured according to the manufacturer’s instructions of an MDA, SOD, and TAC test kits (Teb Pazhouhan Razi, Iran) and GSH reaction kit (Navand Salamat, Urmia, Iran). Briefly, MDA is measured during a colorimetric reaction with thiobarbituric acid in an acidic solution at a temperature of 90–100° C and read at a wavelength of 535 nm. SOD kit catalyzes the free anion radical of superoxide into oxygen and peroxide molecules by producing a chromogen and is assessed at a wavelength of 420 nm by spectrophotometer. TAC assay kit is based on the reduction of Fe3 + to Fe2 + due to antioxidants action and read at a wavelength of 412 nm by spectrophotometer. GSH acts as an antioxidant by protecting cells from free radical damage. GSH was measured based on the GSH recycling system by DTNB and glutathione reductase and samples read at the wavelength 412 nm by spectrophotometer. All biochemical experiments were done in conformity with their kit instructions and were repeated in three independent experiments.
Histological Analysis
Nissl Staining
Animals were anesthetized and then perfused with phosphate buffer solution (PBS 0.1 M) followed by 4% paraformaldehyde in 0.1 M PBS (pH = 7.4). Brains were removed and post-fixed in PFA 4% at 4 °C overnight. Then, tissues were dehydrated in a different alcohol gradient, cleared by incubation in xylene, and placed paraffin and blocked. We obtained serial coronal Sects. (6 µm) of brain tissue by a microtome (Leica RM2135, Germany). For assessing neural cell count and morphology, sections were stained with cresyl fast violet (Nissl). Briefly, after deparaffinization with xylene and rehydration with series of alcohol, tissues were stained with 0.1% cresyl violet (Sigma, Germany) for 5 min at RT. After several washes in distilled water and dehydration in alcohol series, slides were cleaned by xylene and mounted with entellan medium (Merck Chemicals, Germany). Slides were observed under the HUND microscope and the total number of neurons (CFV positive cells = confrontation visual field) counted in DG and CA1-CA4 regions using the ImageJ software in each group and results were shown in Table 3.
Hematoxylin and Eosin Staining
Hematoxylin and eosin (H&E) staining was used to assess neuronal injuries such as necrotic neurons and their structural changes. The brain sections were deparaffinized with xylene and rehydrated in alcohol series. Then, a hematoxylin stain was used for 5 min at RT. Following two washes and clearing in xylene, sections were counterstained with Eosine for 10 min at RT, and the cover slipped after several washes. Slides were observed under HUND Microscope. We captured pictures from the hippocampus in dentate gyrus (DG) and CA1-CA4 regions. Total and necrotic neurons were counted in the CA1-CA4 and DG regions. We considered red and dense neurons necrotic cells in H&E stained section which was counted using the ImageJ software in each experimental groups and results were shown in Table 3. Our images were analyzed from 3 sequential serial Sects. (6 μm) in each slide, and we used three slides (100 μm interval) for each animal. We averaged 9 sections from each animal per group (n = 4). Altogether, 36 sections were analyzed for each experimental group.
Quantitative Real-Time PCR (q-PCR)
In this study, brain tissues of rats were dissected and collected in an RNA-later medium and kept at 4 °C for some hours and then transported to − 20 °C overnight. Then, the RNA-later solution was removed, and the samples were stored at − 80° C until processed for RNA extraction. Total RNA was extracted by an RNA extraction kit (A101231, Pars Tous, Iran). RNA concentration and quality were evaluated by nanodrop spectrophotometer (Thermo Scientific, Wilmington, DE USA), and then, the extracted RNA was immediately transferred to − 80 °C freezer. We synthesized cDNA from 1 µg of total RNA using a reverse transcription reagent kit with random hexamer primers as described by the manufacturer’s protocol (A101161, Pars Tous, Mashhad). The Oligo 7 software (version 7.60) and IDT online software were used to design specific primer sequences (forward and reverse primers) and identify the GFAP and α-syn as target genes and GAPDH as a reference gene. Primer sequences are listed in Table 2. Evaluation of mRNA expression was performed by real-time PCR using Real Q Plus Master Mix Green high rox (A325402, Amplicon, Denmark) and an ABI step-one plus real-time thermocycler (ABI, USA). Briefly, real-time PCR was done by preparing 10 μl of real-time PCR mix, with 0.25 μl of each primer (10 pmol/μl), 1 μl of cDNA, and 2.25 μl of RNase free water. The initial denaturation step was 95 °C for 15 min and then forty cycles at 95 °C for 15 s, 62 °C for 30 s (annealing temperature), and 72 °C for 30 s were exerted in PCR reaction. Finally, the melting curves of amplification were identified via heating the samples from 55 to 95 °C at a linear rate of 0.2 °C/s. The 2-delta-delta CT method was used for the static analysis of the relative expression level of mRNA (Table 2).
Statistical Analysis
Statistical analysis was performed using the GraphPad Prism software (GraphPad Software, Version 7, USA). The results are reported as mean ± SEM. Repeated measure two-way analysis of variance (ANOVA) was used for analyzing the behavioral data in the training period. We used the one-way ANOVA followed by Tukey’s post hoc for analyzing the probe test, histopathology, and qRT-PCR data. The p value < 0.05 was remarked as the minimum level of significance.
Results
Characterization of Astrocyte Cultures Derived from Neonatal Mouse Brains
Primary cultures of astrocytes express important glial markers, such as GFAP, S100 S100-β, and GLAST proteins [32, 33]. Cell-isolated cultures were designated as a relatively pure culture after several passages. Most cells contained astrocytes with a polygonal to fusiform and fat morphology (Fig. 1, a). To confirm the efficacy of the isolation process and purity of astrocytes derived cultures, the immunofluorescence technique was used for GFAP, GLAST, and S100-β antibodies. After 5 passages in vitro, most cells highly expressed GFAP, GLAST, and S100-β as specific astrocyte markers (Fig. 1, b–d). Thus, the isolated cells were purely astrocytes.

Characterization of isolated astrocyte from mice brain. a Phase contrast microscopy reveals that primary astrocytes have a polygonal to fusiform and flat shape under normal condition during 5 passages. b–d Immunofluorescence of GFAP (b), GLAST (c), and S100β (d) as astrocyte markers in the primary culture after passages 5 shows that the culture is purely astrocyte
TQ prevents NP-Induced Cytotoxicity in Astrocyte Culture
To investigate the cytotoxicity of NP and TQ, primary astrocytes were treated with different concentrations of NP and TQ for 24 h, and cell viability was assessed using the MTS assay. NP, at concentrations of 0.25–30 μM, did not noticeably affect cell viability. However, at higher concentrations (40 to 100 µM), it induced a significant cytotoxic effect on cells (all, p < 0.001, Fig. 2, a). The cells treated with TQ showed a reduction in cell viability in a dose-dependent manner compared to the vehicle group. Treatment with 2.5–10 µM TQ did not significantly reduce the cell viability. However, a noticeable decrease was observed at doses of 12.5–20 μM in comparison with the vehicle group (all, p < 0.001, Fig. 2, b). Finally, combination of the most cytotoxic doses of NP and the most effective doses of TQ were examined in vitro. Our results showed that the number of viable cells in the NP 40/TQ 2.5 concentrations was more than that in the vehicle group. Moreover, the number of viable cells in NP 40 + TQ 5, 7.5 and 10 μM was more than the vehicle, but the changes were not statistically significant. However, treatment with different concentrations of TQ in the NP 80 group resulted in more cell death than that in the NP 40 group (Fig. 2, c). Therefore, we can assume that TQ decreases the cytotoxic effect of NP in astrocyte cell culture.

The effect of Nonylphenol (NP) and Thymoquinone (TQ) on cell survival. a Astrocyte treated with graded concentrations of NP and tested after 24 h using the MTS assay. Cell viability of NP exposed cells was noticeably decreased in the concentrations of 40, 80, and 100 µM compared to the vehicle group (all, p < 0.0001). Data were obtained from three independent experiments. Bars indicate mean ± SEM, one-way ANOVA with post hoc Tukey’s test, (***p < 0.001). b Cells were treated with graded concentrations of TQ and tested after 24 h using the MTS assay. TQ significantly reduced primary astrocyte viability at 12.5, 15, 17.5, and 20 µM (all, p < 0.001). Bars indicate mean ± SEM, one-way ANOVA with post hoc Tukey’s test, (*p < 0.05 ** p < 0.01 *** p < 0.001). c Cells were treated with combined concentrations of NP and TQ and tested after 24 h using the MTS assay. MTS assay for astrocyte cells after exposure to the NP plus TQ showed an increase in cell survival at dose of 40/2.5 µM compared to the vehicle and other groups but it was not significant. Data gathered from three independent experiments. Bars indicate mean ± SEM, one-way ANOVA with post hoc Tukey’s test, (*p < 0.05 ** p < 0.01 *** p < 0.001)
TQ Attenuated Learning and Memory Impairment in NP-Treated Animals
Several studies have reported that exposure to NP can lead to the hippocampus injuries, including learning and memory impairment [34]. The MWM task was performed to assess spatial learning and memory in different experimental groups. The averaged escape latency on 4 days of experimental trials was analyzed in different treatment groups as a spatial learning index. Spatial memory was assessed by a probe trial which was based on times that the rat spent in the target quadrant (Q4). Tukey’s post hoc analysis showed that the escape latency in the NP-treated animals was considerably longer than that in the vehicle group in the third (p < 0.0001) and fourth (p < 0.001) days of the training trials (Fig. 3, a). On day 1, TQ supplementation at dose of 10 mg/kg and NP + TQ (10 mg/kg) decreased the escape latency compared to the NP group (p = 0.04). On day 2, TQ supplementation at doses of 2.5 mg/kg (p = 0.0001), 5and 10 mg/kg (p < 0.0001) and TQ alone (p < 0.0001) decreased the escape latency compared to the NP group. On day 3, the escape latency index in NP + TQ (2.5 mg/kg, p = 0.002) and NP + TQ (5 mg/kg, p < 0.0001) groups was noticeably shorter than that in the NP group. On day 4, the escape latency in the NP + TQ (2.5 mg/kg, p = 0.016), NP + TQ (5 mg/kg, p = 0.001), NP + TQ (10 mg/kg, p = 0.007), and TQ (10 mg/kg, p = 0.01) groups was considerably reduced compared to the NP group (Fig. 3, b).

TQ improves NP-induced spatial learning and memory deficit. a–b Comparison of escape latency, c time duration in the target zone, d swimming velocity in all treatment groups. The escape latency in Nonylphenol (NP) group was considerably more than that in vehicle group on days 3 and 4 of training (++++p < 0.0001, +++ p < 0.001). In the vehicle group, the escape latency index significantly decreased on days 3 and 4 compared to the first day of examination (^^^^p < 0.0001, Fig. 3, a). Moreover, the escape latency in TQ treated groups at all doses was significantly decreased at days 2, 3, and 4 of training trails compared to the first day. On day 1, the escape latency in the NP + TQ (10 mg/kg) treated group was significantly decreased compared to the NP group (*p = 0.04). On day 2, TQ decreased the escape latency at all doses compared to the NP group. On day 3, it was observed that the escape latency index in the NP + TQ (2.5 mg/kg, **** p = 0.002) and NP + TQ (5 mg/kg, **** p < 0.0001) groups was noticeably shorter than that compared to the NP group. On day 4, the escape latency in the TQ treated groups at all doses was significantly lesser than that in the NP group (Fig. 3, b). In assessment of time duration spent in the target zone, NP-treated animals spent less time in target quadrant (Q4) than the vehicle group (++++p < 0.0001, Fig. 3, d). Moreover, TQ treatment only at dose of 5 mg/kg significantly enhanced the time spent in the target quadrant compared to the NP group (*p < 0.05, Fig. 3, c). (^) Sign indicates significant difference in the NP group compared to the vehicle group and (*) sign indicates significant difference in the NP group compared to the TQ treatment groups
In the vehicle group, the escape latency index on days 3 and 4 (p < 0.0001) was significantly decreased compared to day 1, but this pattern was not seen in the NP group (Fig. 3, a). Moreover, comparison within NP + TQ (2.5 mg/kg) group showed that the escape latency on days 2 (p = 0.037), 3 (p = 0.003) and 4 (p < 0.001) was reduced compared to day 1. In the NP + TQ (5 mg/kg) group, a significant reduction in escape latency was seen on days 2, 3, and 4 compared to day 1 (p < 0.0001). In addition, in the NP + TQ (10 mg/kg) group, the escape latency was decreased on days 2, 3, and 4 (p < 0.0001) compared to day 1. Moreover, in the NP + TQ (10 mg/kg) group, the escape latency on days 2 (p = 0.011), 3, and 4 (p < 0.0001) was shorter than that on day 1. Also, in the TQ (10 mg/kg) group, there was a significant reduction in the escape latency on day 3 compared to day 1 (p < 0.001, Fig. 3, b).
For evaluation of spatial memory, the platform was removed at the end of the experiment after 4 days training period. The time spent in Q4 was measured and analyzed in all groups. The analysis showed that NP-treated animals spent less time in Q4 than that in the vehicle group (p < 0.0001). TQ treatment at dose of 5 mg/kg significantly enhanced the time spent in Q4 compared to the NP group (p < 0.05, Fig. 4, c). Furthermore, no significant changes were seen in swimming velocity after NP and TQ treatments. These data indicate that TQ improves learning and memory function in NP-treated animals.

Thymoquinone (TQ) treatment enhances antioxidant capacity against NP-induced neurotoxicity. Rat brain was used at the end of the experiment to assess the levels of TAC (a–b), MDA (c–d), GSH (e), and SOD (f). The results showed that NP significantly increased the brain (++++p < 0.0001) and serum (+++p < 0.001) levels of TAC (a–b) compared to the vehicle group. The brain TAC level was significantly increased in TQ treatment at doses of 2.5, 5, and 10 mg/kg, respectively, * p < 0.05, ** p < 0.01, ** p < 0.01. In serum, TQ only at dose of 5 mg/kg significantly increased TAC level (**p < 0.01). Results of MDA assay indicated that both tissue (++++p < 0.0001) and serum (+++p < 0.001) levels of the MDA increase in the NP-treated group compared to the vehicle group. Moreover, TQ in all doses except 2.5 mg/kg modulated brain MDA level (*p < 0.05, c-d). Furthermore, the results indicated that the brain GSH (++++p < 0.0001) and serum SOD (++++p < 0.0001) levels in the NP group were lesser than those in the vehicle group (e–f). Moreover, a significant increase was observed in GSH level in the NP + TQ (5 mg/kg, * p < 0.05) and TQ (10 mg/kg, *** p < 0.001) groups compared to the NP group (e). SOD assay showed that TQ at doses of 5 and 10 mg/kg increased serum SOD level (*p < 0.05, f)

Effect of Thymoquinone (TQ) on histopathological changes induced by Nonylphenol (NP) in rat models. Histopathological changes were evaluated using Nissl staining. Representative images for the vehicle, NP, NP + TQ2.5, NP + TQ5, NP + TQ10, and TQ10 groups are provided (magnification: × 400, × 40)
TQ Reduced Oxidative Stress and Enhanced Total Antioxidant Capacity in NP-Treated Animals
To evaluate the antioxidant effects of TQ on NP-induced oxidative stress, the TAC, MDA, SOD, and GSH parameters were assessed based on their kit instructions. The TAC level in tissue (p < 0.0001) and serum (p < 0.001) was considerably reduced in the NP group compared to the vehicle group (p < 0.0001, Fig. 4, a). Tissue TAC level in the NP + TQ (2.5 mg/kg, p < 0.05), NP + TQ (5 mg/kg, p < 0.01), NP + TQ (10 mg/kg, p < 0.01) groups significantly increased compared to the NP groups (Fig. 4, a). Moreover, serum TAC level in all TQ treated groups was higher than that in the NP group, but a significant change was observed only in the NP + TQ (5 mg/kg, p < 0.01) group (Fig. 4, b). Our results showed that MDA levels in serum (p < 0.001) and tissue (p < 0.0001) were significantly increased in the NP group compared to the vehicle group (Fig. 4, c). Tissue MDA level in the NP + TQ (5 mg/kg), NP + TQ (10 mg/kg), and TQ (10 mg/kg) groups was significantly reduced compared to the NP group (p < 0.05, Fig. 4, c). However, no significant difference was observed in serum MDA level in the TQ treated groups compared to the NP group (Fig. 4, d). In the NP group, serum SOD level was significantly higher than that in the vehicle group (p < 0.0001, Fig. 4, f). However, SOD level was significantly increased in the NP + TQ (5 mg/kg) and NP + TQ (10 mg/kg) groups compared to the NP group (Fig. 4, f). Biochemical analysis for GSH level indicated that NP significantly decreased GSH level compared to the vehicle group (p < 0.0001), while GSH level was significantly increased in the NP + TQ (5 mg/kg, p < 0.05) and NP + TQ (10 mg/kg, p < 0.001) groups compared to the NP group. These data suggest that TQ enhances the antioxidant capacity of the CNS by increasing antioxidant factors and reducing oxidative markers.
The Neuroprotective Effect of TQ Decreases NP-Induced Neural Injury in the Hippocampus
To evaluate the neuroprotective effect of TQ, the hippocampal regions were analyzed by H&E (Fig. 5) and Nissl (Fig. 6) staining. We found that the total number of neurons in the DG (140.63 ± 4.91, p < 0.0001) and CA1-CA4 (120.3 ± 2.59, p < 0.0001) regions of the NP-treated group was less than that in the vehicle group (Table 3). Moreover, no significant difference was observed between the TQ treated and NP groups in the total number of neurons. The population of necrotic neurons was shown as a percentage of the total number of neurons counted in the DG and CA1-CA4 regions of the hippocampus (Table 3). Our results showed that the percentage of necrotic neurons in in the DG (12.53%, p < 0.001) and CA1-CA4 (9.59%, p < 0.0001) regions of the NP-treated group was significantly higher than that in the vehicle group. However, TQ treatment in the NP + TQ (2.5 mg/kg) group decreased the percentage of necrotic neurons in the DG (7.28%, p < 0.01) and CA1-CA4 (5.85%, p < 0.001) regions compared to the NP group. Moreover, the percentage of necrotic neurons was considerably reduced in the DG (6.31%, p < 0.001) and CA1-CA4 (4.88%, p < 0.001) regions of the NP + TQ (5 mg/kg) group, and in the CA1-CA4 regions (4.86%, p < 0.01) of the TQ (10 mg/kg) group compared to the NP group. Therefore, it could be concluded that TQ treatment has neuroprotective effect against NP induced brain injuries such as necrosis and neuronal loss.

Effect of Thymoquinone (TQ) on histopathological changes induced by Nonylphenol (NP) in rat models. Histopathological changes were evaluated using H&E staining. Necrotic cells were indicated on the images by arrows. We considered red and dense neurons as necrotic cells in H&E stained section which was counted using the ImageJ software in each experimental groups and results were shown in Table 3. We counted the total number of neurons (CFV positive cells = confrontation visual field) in DG and CA1-CA4 regions using the ImageJ software in each group and results were shown in Table 3. Representative images for the vehicle, NP, NP + TQ2.5, NP + TQ5, NP + TQ10, and TQ10 groups are provided (magnification: × 400)
TQ Reduces the Expression Level of GFAP and Enhances α-syn in NP- Treated Groups
Upregulation of GFAP expression (as astrocyte marker) indicates gliosis and glial activation [35]. To evaluate the CNS gliosis, we assessed GFAP expression. Results showed that administration of NP at dose of 25 mg/kg significantly increased the GFAP gene expression compared to the vehicle group (p < 0.01, Fig. 7, a). TQ treatment downregulated the level of GFAP in the NP + TQ (2.5 mg/kg, p < 0.05), NP + TQ (5 mg/kg, p < 0.01), and NP + TQ (10 mg/kg, p < 0.01) groups compared to the NP group (Fig. 7, a). α-syn is a synaptic protein that plays a key role in neurotransmitter release and synaptic plasticity [36]. Over-expression of the α-syn gene can lead to α-syn neuronal inclusions that causes neurodegeneration [37]. Our results showed that the α-syn expression was upregulated in the NP group compared to the vehicle group (p = 0.004). Moreover, TQ treatment significantly downregulated the α-syn expression level in all groups, but significant changes were observed only in the NP + TQ (5 mg/kg, p < 0.01) and TQ (10 mg/kg, p < 0.01) groups compared to the NP group (Fig. 7, b). Our results confirmed that TQ significantly reduced the expression of GFAP and α-syn following NP-induced neurotoxicity.

TQ treatment moderated the expression of genes by its neuroprotective properties. The results showed that Nonylphenol (NP)-induced neurotoxicity significantly upregulated the expression level of GFAP compared to the vehicle group (**p < 0.01, a–b). Our molecular results showed that in all Thymoquinone (TQ)-treated groups (2.5, 5, 10 mg/kg), there was a significant reduction in GFAP gene expression (*p < 0.05, ** p < 0.01, and ** p < 0.01, respectively, Fig. 7, a). Moreover, expression of α-syn was increased in the NP group compared to the vehicle group (*p < 0.05, Fig. 7, b). Furthermore, in the TQ treatment groups at doses of 5 and 10 mg/kg, α-syn expression was downregulated compared to the NP group (*p < 0.05, Fig. 7, b). One-way ANOVA (Tukey’s post hoc test) was used to perform statistical analysis
Discussion
We investigated whether TQ could present beneficial impacts on the NP-induced neurotoxicity in an animal model. We observed that treatment with TQ reduces NP-induced cytotoxicity in vitro. Subsequently, TQ prevented neurotoxicity symptoms and improved memory deficit induced by NP. Moreover, TQ increased the levels of antioxidant parameters such as TAC, GSH, and SOD and reduced MDA level as an oxidative stress marker. Histological study indicated that the number of necrotic cells in the TQ treated groups significantly decreased in the hippocampal regions compared to the NP-treated animals. Finally, we found that treatment with TQ reduced GFAP and α-syn expression level. These findings suggest that TQ may be effective in reducing NP-induced neurotoxicity through its antioxidant, neuroprotective and anti-inflammatory effects.
In recent years, there have been growing concerns about the neurotoxic effect of persistent organic pollutants (POPs), such as morphological and physiological changes [38]. Numerous investigations have evidenced that perinatal exposure to POPs can lead to learning and memory deficits in rats [39]. NP, as a potent POP, has been demonstrated to be potentially neurotoxic leading to spatial memory and cognitive impairment in animal models [34]. In this study, we used NP to induce learning and memory impairment as an animal model. Previous studies have indicated that exposure to NP causes memory and learning deficits [40, 41]. Developmental exposure to NP has also been shown to induce axonal damage in the developing neurons in vitro and in vivo [42]. The neurotoxicity of NP may, in part, be owing to an increase in the production of inflammatory mediators. For example, perinatal exposure to NP was found to activate microglia in offspring hippocampus and thereby cause neuroinflammation [16].
In the present study, we observed that NP reduced cell survival and induced cell toxicity. In line with our results, exposure to NP has been reported to induce apoptosis and decrease cell viability in primary cortical neurons [43]. Another study demonstrated that NP induces cytotoxicity through apoptosis by the cell cycle arrest and activation of the caspase cascade [44]. To better understand the interactions between NP toxicity and TQ neuroprotection, we administered different doses of NP and TQ to treat cultured astrocytes and evaluate the cell viability during NP cytotoxicity using MTP test. We observed that TQ protected astrocytes from cytotoxicity induced by NP. It has been reported that TQ (0.01, 0.1, 1, and 10 μM) protected dopaminergic neurons against 1-methyl-4-phenylpyridinium (MPP) by inhibiting apoptotic cell death and maintaining mitochondrial function [45]. Another study demonstrated that TQ protects primary rat cortical neurons against ethanol-induced neuronal apoptosis [23]. Moreover, it was evident that pretreatment with TQ inhibited neurotoxicity induced by beta-amyloid in primary cultured cerebellar granule neurons [46]. These results confirm the neuroprotective properties of TQ against several in vitro neurotoxicity models.
In the present study, we also showed that NP reduced learning and memory ability in rats. Several studies have demonstrated that NP disrupts oxidant/antioxidant balance and causes cognitive and memory impairments in rodents [41, 47]. Our biochemical results showed that NP increases the brain level of MDA is commonly known as a marker of oxidative injury. On the other hand, NP decreased the level of antioxidant parameters of TAC, SOD, and GSH. Consistent with our data, it has been reported that NP increases brain MDA levels and decreases the activity of SOD, CAT in mice [12]. This suggests that oxidative damage may be involved in NP-induced neurotoxicity. Several studies have suggested that NP induces apoptosis and necrosis in neurons [11, 48]. We found that the number of necrotic cells in the NP-treated group was increased compared to the other groups. In line with other studies, we also found that NP causes neurotoxicity through misbalancing oxidant/antioxidant state as well as inducing apoptosis and necrosis in neuronal cells accompanied by learning and memory deficits.
TQ has various therapeutic properties such as antioxidant [49], anti-inflammatory, and anti-cancer effects [50]. In our study, significant improvements in learning level were observed in all TQ treatment groups compared to the NP-treated group. Furthermore, TQ treatment specifically at dose of 5 mg/kg considerably enhanced memory function in the probe task. TQ is considered as a potent neuroprotective compound to protect neurons from degeneration in different models of neurotoxicity [51, 52]. Consistent with our results, Radad et al. (2009) reported that TQ at doses of 15 and 7.5 mg/kg had effective effects in rotenone-induced rat model of Parkinson’s disease. They also showed that TQ significantly improved the neuronal damage caused by rotenone toxin in the animals [53]. It was also reported that TQ at doses of 2, 5, and 10 mg/kg has protective effect on neurotoxicity induced by LPS in rats and improves learning and memory function in the animals [54]. Abulfadl et al. (2018) reported the protective effects of TQ at doses of 10, 20, and 40 mg/kg in an aluminum chloride-induced animal model of Alzheimer’s disease. They found that TQ can reduce beta-amyloid formation and improve animal learning and memory performance in the MWM test [55]. In line with above mentioned studies, we indicated that TQ possesses potent antioxidant and neuroprotective properties and improves learning and memory impairments in our animal model. To realize the therapeutic mechanisms of TQ, we investigated several mechanisms such as cell survival, oxidative stress, the expression of GFAP and α-syn and histopathology. Our results indicated that TQ could reduce glial activation, oxidative damage, and inflammation induced by NP, which might contribute to its effect on learning and memory improvement.
Oxidative stress has a crucial role in neurodegeneration progression [56]. MDA is produced as an end product of lipid peroxidation induced by a variety of oxidative damage [57]. The current study showed that the brain level of MDA was significantly increased in the NP-treated group. Several studies have reported that oxidative stress is one of the main mechanisms of NP toxicity and various neurological diseases [58, 59]. We found that TQ significantly decreased the brain MDA level and increased antioxidant status. The antioxidant defense is predominantly enzymatic and includes glutathione peroxidase (cGPx), SOD, and CAT. GSH is the essential non-enzymatic antioxidant defense that protects cells from reactive oxygen species attacks. GSH measurement of the brain was performed due to its critical role in maintaining balance between oxidative stress and antioxidant system. The brain is very vulnerable to oxidative stress because it is an organ rich in unsaturated fats. Moreover, disturbing the brain antioxidant system causes oxidative stress, which may be involved in the pathogenesis of some neurodevelopmental and neurodegenerative diseases [60, 61]. In the present study, we found that TQ increased brain and serum antioxidant defense by enhancing SOD, GSH, and TAC levels. A large body of evidence confirms that TQ increases antioxidant capacity of the brain tissue and protects neural cells from various injuries due to its antioxidant and anti-inflammatory effects [49, 51, 62]. Kassab and El-Hennamy (2017) reported that TQ reduces arsenic-induced neurotoxicity at dose of 10 mg/kg. They showed that TQ significantly decreased MDA level and increased the level of SOD, GSH, and CAT in the cerebral cortex, cerebellum, and brain stem of arsenic-treated female rats [63]. Another study also indicated that TQ at doses of 5 and 10 mg/kg significantly decreased the levels of MDA and enhanced the GSH levels after administration of acrylamide [64]. Thus, antioxidant property of TQ could be considered as one protective mechanism against learning and memory impairment in our animal model.
Histopathological evaluation of the rat hippocampus indicated some pathological changes including necrotic cells and cell loss in the NP-treated rats compared to the vehicle group. Recently, it has been shown that NP induces apoptosis and necrosis in in cultured cortical neurons by stimulating inflammatory and apoptotic factors [65]. Moreover, TQ was found to improve brain tissue injury through an increase in the hippocampal neurogenesis [66]. Pervious study revealed that TQ and Nigella Sativa improved the hippocampal neuronal pathology induced by toluene exposure in rats [67]. In agreement with previous reports, our data indicated that TQ treatment reduced the necrotic neuron densities in the NP-treated animals. It reduced oxidative stress and gliosis and improved neuronal survival. Altogether, it can be assumed that TQ is a neuroprotective agent and enhances cognitive function in the NP-induced neurotoxicity rat model.
GFAP is a protein expressed by various nerve cells, especially in astrocytes, during development and is known as a sensitive and critical marker of astrocyte activation following stress or injury [68]. Since gliosis, especially microglia and astrocytes reactivation, is one of the symptoms of NP neurotoxicity, we assessed GFAP gene expression in different experimental groups. Nerve damage and infection in the CNS trigger microglia and astrocyte activation. During brain damage and neurodegenerative disorders, astrocytes undergo a series of morphological changes that lead to the activation of the inflammatory responses, an increase in the number of astrocytes, and the expression of the GFAP gene that called astrogliosis [69]. It was reported that NP induces the GFAP overexpression and astrogliosis [70]. In line with these reports, we observed a significant increase in GFAP gene expression in the NP group compared to other groups. In addition to astrocyte, several studies indicated that exposure to different doses of NP affects the number and function of glial cells [16]. Besides, it was demonstrated that TQ (5 mg/kg) significantly reduced the GFAP expression in acrylamide treated animals [64]. We also showed that TQ at doses of 5 and 10 mg/kg significantly decreased the expression level of GFAP in the NP-treated groups. This beneficial effect of TQ in attenuating GFAP gene expression could be attributed to its anti-inflammatory properties.
α-syn is a small peripheral membrane protein particularly found at synaptic terminals, indicating a critical role of this protein in neurotransmitter release [71] and synaptic plasticity [72]. It has been reported that the loss of α-syn has little effect on synaptic transmission, while its over-expression causes cognitive dysfunction in Parkinson’s and Alzheimer’s diseases [73]. TQ was found to effectively protect cultured hippocampal neurons against α-syn–induced synapse damage and interference in regular synaptic activity [74]. Another study discussed that the protective effects of TQ in the MPTP-induced Parkinson’s model was due to α-syn modulation. In this study, TQ prevented accumulations of α-syn protein at the axon terminal of neurons. Furthermore, it has been reported that TQ considerably prevented fibril formation induced by α-syn accumulation and cell death in MPTP toxicity [75].
Consistent with this report, we indicated that α-syn gene expression was upregulated in the rat brain following NP exposure, and TQ significantly downregulated the expression of α-syn in the NP-treated groups. Over-expression of the α-syn gene can lead to the formation of toxic filamentous and α-syn neuronal inclusions, which causes neurodegeneration [37]. Moreover, abnormal accumulation and aggregation of α-syn in the form of Lewy bodies and neurites cause dopaminergic neuronal loss in the substantia nigra as landmark in Parkinson’s disease [76]. Together, it can be concluded that NP interrupts synaptic plasticity by upregulating α-Syn, and treatment with TQ reduces the expression of α-syn and enhances synaptic plasticity and learning and memory function in NP exposed animals.
Conclusion
Overall, the present study indicated that TQ has therapeutic potential in learning and memory impairment associated with NP neurotoxicity through its anti-inflammatory, neuroprotective, and antioxidant properties. It seems that the therapeutic effects of TQ are associated with reductions in gliosis, oxidative damage, and inflammatory mediators. Further study is needed to understand the exact underlying mechanisms of TQ as a hopeful therapeutic target in NP neurotoxicity. Upcoming clinical trials will provide deeper insights into the role of TQ as a useful compound for learning and memory improvement in order to develop strategies to better follow and treat various neurotoxic effects associated with NP.
Data Availability
Data are available from the authors upon request.
References
Diamanti-Kandarakis E, Bourguignon J-P, Giudice LC et al (2009) Endocrine-disrupting chemicals: an endocrine society scientific statement. Endocr Rev 30:293–342. https://doi.org/10.1210/er.2009-0002
Pinson A, Bourguignon JP, Parent AS (2016) Exposure to endocrine disrupting chemicals and neurodevelopmental alterations. Andrology 4:706–722
Chokwe TB, Okonkwo JO, Sibali LL (2017) Distribution, exposure pathways, sources and toxicity of nonylphenol and nonylphenol ethoxylates in the environment. Water SA 43:529–542. https://doi.org/10.4314/wsa.v43i4.01
Kim H, Oh S, Gye MC, Shin I (2018) Comparative toxicological evaluation of nonylphenol and nonylphenol polyethoxylates using human keratinocytes. Drug Chem Toxicol 41:486–491. https://doi.org/10.1080/01480545.2017.1391829
Monteiro-Riviere NA, Van Miller JP, Simon G et al (2000) Comparative in vitro percutaneous absorption of nonylphenol and nonylphenol ethoxylates (NPE-4 and NPE-9) through human, porcine and rat skin. Toxicol Ind Health 16:49–57. https://doi.org/10.1177/074823370001600201
Sciarrillo R, Di Lorenzo M, Valiante S et al (2021) OctylPhenol (OP) alone and in combination with NonylPhenol (NP) Alters the structure and the function of thyroid gland of the lizard Podarcis siculus. Arch Environ Contam Toxicol 80:567–578. https://doi.org/10.1007/s00244-021-00823-5
Geens T, Neels H, Covaci A (2012) Distribution of bisphenol-A, triclosan and n-nonylphenol in human adipose tissue, liver and brain. Chemosphere 87:796–802. https://doi.org/10.1016/j.chemosphere.2012.01.002
Vivacqua A, Recchia AG, Fasanella G et al (2003) The food contaminants bisphenol A and 4-nonylphenol act as agonists for estrogen receptor α in MCF7 breast cancer cells. Endocrine 22:275–284. https://doi.org/10.1385/ENDO:22:3:275
Jie X, JianMei L, Zheng F et al (2013) Neurotoxic effects of nonylphenol: a review. Wien Klin Wochenschr 125:61–70. https://doi.org/10.1007/s00508-012-0221-2
Arukwe A, Thibaut R, Ingebrigtsen K et al (2000) In vivo and in vitro metabolism and organ distribution of nonylphenol in Atlantic salmon (Salmo salar). Aquat Toxicol 49:289–304. https://doi.org/10.1016/S0166-445X(99)00084-3
Gu W, Wang Y, Qiu Z et al (2018) Maternal exposure to nonylphenol during pregnancy and lactation induces microglial cell activation and pro-inflammatory cytokine production in offspring hippocampus. Sci Total Environ 634:525–533. https://doi.org/10.1016/j.scitotenv.2018.03.329
Korkmaz A, Ahbab MA, Kolankaya D, Barlas N (2010) Influence of vitamin C on bisphenol A, nonylphenol and octylphenol induced oxidative damages in liver of male rats. Food Chem Toxicol 48:2865–2871. https://doi.org/10.1016/j.fct.2010.07.019
Eid Z, Mahmoud UM, Mekkawy IAA et al (2021) 4-Nonylphenol induced brain damage in juvenile African catfish (Clarias garepinus). Toxicol Environ Health Sci 13:201–214. https://doi.org/10.1007/s13530-021-00080-y
Tabassum H, Ashafaq M, Parvez S, Raisuddin S (2017) Role of melatonin in mitigating nonylphenol-induced toxicity in frontal cortex and hippocampus of rat brain. Neurochem Int 104:11–26. https://doi.org/10.1016/j.neuint.2016.12.010
Heba SA, Yasser AK, Iman MM (2015) The effect of bisphenol A on some oxidative stress parameters and acetylcholinesterase activity in the heart of male albino rats. Cytotechnology 67(1):145–155. https://doi.org/10.1007/s10616-013-9672-1
Qiu Z, Wang Y, Chen J (2019) Perinatal exposure to nonylphenol induces microglia-mediated nitric oxide and prostaglandin E2 production in offspring hippocampus. Toxicol Lett 301:114–124. https://doi.org/10.1016/j.toxlet.2018.11.013
Rehman MU, Wali AF, Ahmad A et al (2018) Neuroprotective strategies for neurological disorders by natural products: an update. Curr Neuropharmacol 17:247–267. https://doi.org/10.2174/1570159x16666180911124605
Hannan MA, Rahman MA, Sohag AAM et al (2021) Black Cumin (Nigella sativa L.): a comprehensive review on phytochemistry, health benefits, molecular pharmacology, and safety. Nutrients 13:1784. https://doi.org/10.3390/nu13061784
Ahmad MF, Ahmad FA, Ashraf SA et al (2021) An updated knowledge of Black seed (Nigella sativa Linn.): review of phytochemical constituents and pharmacological properties. J Herb Med 25. https://doi.org/10.1016/j.hermed.2020.100404
El Gazzar M, El Mezayen R, Marecki JC et al (2006) Anti-inflammatory effect of thymoquinone in a mouse model of allergic lung inflammation. Int Immunopharmacol 6:1135–1142. https://doi.org/10.1016/j.intimp.2006.02.004
Darakhshan S, Bidmeshki Pour A, Hosseinzadeh Colagar A, Sisakhtnezhad S (2015) Thymoquinone and its therapeutic potentials. Pharmacol Res 95–96:138–158. https://doi.org/10.1016/j.phrs.2015.03.011
Cobourne-Duval MK, Taka E, Mendonca P et al (2016) The antioxidant effects of thymoquinone in activated BV-2 murine microglial cells. Neurochem Res 41:3227–3238. https://doi.org/10.1007/s11064-016-2047-1
Ullah I, Ullah N, Naseer MI, et al (2012) Neuroprotection with metformin and thymoquinone against ethanol-induced apoptotic neurodegeneration in prenatal rat cortical neurons. BMC Neurosci 13. https://doi.org/10.1186/1471-2202-13-11
Poorgholam P, Yaghmaei P, Hajebrahimi Z. Thymoquinone recovers learning function in a rat model of Alzheimer’s disease. Avicenna J phytomedicine 8:188–197. https://doi.org/10.22038/ajp.2018.21828.1820
Alemi M, Sabouni F, Sanjarian F et al (2013) Anti-inflammatory effect of seeds and Callus of Nigella sativa L. extracts on mix glial cells with regard to their thymoquinone content. AAPS PharmSciTech 14:160–167. https://doi.org/10.1208/s12249-012-9899-8
Bargi R, Asgharzadeh F, Beheshti F et al (2017) The effects of thymoquinone on hippocampal cytokine level, brain oxidative stress status and memory deficits induced by lipopolysaccharide in rats. Cytokine 96:173–184. https://doi.org/10.1016/j.cyto.2017.04.015
Dong J, Zhang X, Wang S, et al (2021) Thymoquinone prevents dopaminergic neurodegeneration by attenuating oxidative stress via the Nrf2/ARE pathway. Front Pharmacol 11. https://doi.org/10.3389/fphar.2020.615598
Zarei-Kheirabadi M, Mirsadeghi S, Vaccaro AR et al (2020) Protocol for purification and culture of astrocytes: useful not only in 2 days postnatal but also in adult rat brain. Mol Biol Rep 47:1783–1794. https://doi.org/10.1007/s11033-020-05272-2
Malich G, Markovic B, Winder C (1997) The sensitivity and specificity of the MTS tetrazolium assay for detecting the in vitro cytotoxicity of 20 chemicals using human cell lines. Toxicology 124:179–192. https://doi.org/10.1016/s0300-483x(97)00151-0
Hosseinzadeh H, Parvardeh S, Asl MN et al (2007) Effect of thymoquinone and Nigella sativa seeds oil on lipid peroxidation level during global cerebral ischemia-reperfusion injury in rat hippocampus. Phytomedicine 14:621–627. https://doi.org/10.1016/j.phymed.2006.12.005
D’Hooge R, De Deyn PP (2001) Applications of the Morris water maze in the study of learning and memory. Brain Res Rev 36:60–90. https://doi.org/10.1016/S0165-0173(01)00067-4
Duan S, Anderson CM, Stein BA, Swanson RA (1999) Glutamate induces rapid upregulation of astrocyte glutamate transport and cell-surface expression of GLAST. J Neurosci 19:10193–10200. https://doi.org/10.1523/jneurosci.19-23-10193.1999
Donato R, Sorci G, Riuzzi F et al (2009) S100B’s double life: intracellular regulator and extracellular signal. Biochim Biophys Acta - Mol Cell Res 1793:1008–1022. https://doi.org/10.1016/j.bbamcr.2008.11.009
Kazemi S, Khalili-Fomeshi M, Akbari A et al (2018) The correlation between nonylphenol concentration in brain regions and resulting behavioral impairments. Brain Res Bull 139:190–196. https://doi.org/10.1016/j.brainresbull.2018.03.003
Brahmachari S, Fung YK, Pahan K (2006) Induction of glial fibrillary acidic protein expression in astrocytes by nitric oxide. J Neurosci 26:4930–4939. https://doi.org/10.1523/JNEUROSCI.5480-05.2006
Cheng F, Vivacqua G, Yu S (2011) The role of alpha-synuclein in neurotransmission and synaptic plasticity. J Chem Neuroanat 42:242–248. https://doi.org/10.1016/j.jchemneu.2010.12.001
Desplats P, Spencer B, Crews L et al (2012) α-Synuclein induces alterations in adult neurogenesis in Parkinson disease models via p53-mediated repression of notch. J Biol Chem 287:31691–31702. https://doi.org/10.1074/jbc.M112.354522
Doucet J, Tague B, Arnold DL et al (2009) Persistent organic pollutant residues in human fetal liver and placenta from greater Montreal, Quebec: a longitudinal study from 1998 through 2006. Environ Health Perspect 117:605–610. https://doi.org/10.1289/ehp.0800205
Tiwari SK, Agarwal S, Chauhan LKS et al (2015) Bisphenol-A impairs myelination potential during development in the hippocampus of the rat brain. Mol Neurobiol 51:1395–1416. https://doi.org/10.1007/s12035-014-8817-3
Jie X, Yang W, Jie Y et al (2010) Toxic effect of gestational exposure to nonylphenol on F1 male rats. Birth Defects Res Part B - Dev Reprod Toxicol 89:418–428. https://doi.org/10.1002/bdrb.20268
Li M, You M, Li S et al (2019) Effects of maternal exposure to nonylphenol on learning and memory in offspring involve inhibition of BDNF-PI3K/Akt signaling. Brain Res Bull 146:270–278. https://doi.org/10.1016/j.brainresbull.2019.01.014
Li S, You M, Chai W et al (2019) Developmental exposure to nonylphenol induced rat axonal injury in vivo and in vitro. Arch Toxicol 93:2673–2687. https://doi.org/10.1007/s00204-019-02536-0
Kim SK, Kim BK, Shim JH et al (2006) Nonylphenol and octylphenol-induced apoptosis in human embryonic stem cells is related to Fas-Fas ligand pathway. Toxicol Sci 94:310–321. https://doi.org/10.1093/toxsci/kfl114
Kudo C, Wada K, Masuda T et al (2004) Nonylphenol induces the death of neural stem cells due to activation of the caspase cascade and regulation of the cell cycle. J Neurochem 88:1416–1423. https://doi.org/10.1046/j.1471-4159.2003.02270.x
Radad KS, Al-Shraim MM, Moustafa MF, Rausch WD (2015) Neuroprotective role of thymoquinone against 1-methyl-4-phenylpyridinium-induced dopaminergic cell death in primary mesencephalic cell culture. Neurosciences 20:10–16
Ismail N, Ismail M, Mazlan M et al (2013) Thymoquinone prevents β-amyloid neurotoxicity in primary cultured cerebellar granule neurons. Cell Mol Neurobiol 33:1159–1169. https://doi.org/10.1007/s10571-013-9982-z
Mao Z, Zheng YL, Zhang YQ (2011) Behavioral impairment and oxidative damage induced by chronic application of nonylphenol. Int J Mol Sci 12:114–127. https://doi.org/10.3390/ijms12010114
Mao Z, Zheng Y, Zhang Y et al (2008) Chronic application of nonylphenol-induced apoptosis via suppression of bcl-2 transcription and up-regulation of active caspase-3 in mouse brain. Neurosci Lett 439:147–152. https://doi.org/10.1016/j.neulet.2008.05.006
Staniek K, Gille L (2010) Is thymoquinone an antioxidant? BMC Pharmacol 10. https://doi.org/10.1186/1471-2210-10-s1-a9
Woo CC, Kumar AP, Sethi G, Tan KHB (2012) Thymoquinone: potential cure for inflammatory disorders and cancer. Biochem Pharmacol 83:443–451. https://doi.org/10.1016/j.bcp.2011.09.029
Isaev NK, Chetverikov NS, Stelmashook EV et al (2020) Thymoquinone as a potential neuroprotector in acute and chronic forms of cerebral pathology. Biochem 85:167–176. https://doi.org/10.1134/S0006297920020042
Abulfadl YS, El-Maraghy NN, Ahmed AAE et al (2018) Protective effects of thymoquinone on D-galactose and aluminum chloride induced neurotoxicity in rats: biochemical, histological and behavioral changes. Neurol Res 40:324–333. https://doi.org/10.1080/01616412.2018.1441776
Radad K, Moldzio R, Taha M, Rausch WD (2009) Thymoquinone protects dopaminergic neurons against MPP+ and rotenone. Phyther Res 23:696–700. https://doi.org/10.1002/ptr.2708
Bargi R, Asgharzadeh F, Beheshti F, Hosseini M, Sadeghnia HR, Khazaei M (2017) The effects of thymoquinone on hippocampal cytokine level, brain oxidative stress status and memory deficits induced by lipopolysaccharide in rats. Cytokine 96:173–184. https://doi.org/10.1016/j.cyto.2017.04.015
Abulfadl YS, El-Maraghy NN, Ahmed AAE et al (2018) Thymoquinone alleviates the experimentally induced Alzheimer’s disease inflammation by modulation of TLRs signaling. Hum Exp Toxicol 37:1092–1104. https://doi.org/10.1177/0960327118755256
Federico A, Cardaioli E, Da Pozzo P et al (2012) Mitochondria, oxidative stress and neurodegeneration. J Neurol Sci 322:254–262. https://doi.org/10.1016/j.jns.2012.05.030
Ayala A, Muñoz MF, Argüelles S (2014) Lipid peroxidation: production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid Med Cell Longev 2014. https://doi.org/10.1155/2014/360438
Lotfi M, Hasanpour AH, Moghadamnia AA, Kazemi S (2020) The investigation into neurotoxicity mechanisms of nonylphenol: a narrative review. Curr Neuropharmacol 19:1345–1353. https://doi.org/10.2174/1570159x18666201119160347
Floyd RA (1999) Antioxidants, oxidative stress, and degenerative neurological disorders. Proc Soc Exp Biol Med 222:236–245. https://doi.org/10.1046/j.1525-1373.1999.d01-140.x
Zheng L, Marcusson J, Terman A (2006) Oxidative stress and Alzheimer disease: the autophagy connection? Autophagy 2:143–145. https://doi.org/10.4161/auto.2.2.2444
Zawia NH, Lahiri DK, Cardozo-Pelaez F (2009) Epigenetics, oxidative stress, and Alzheimer disease. Free Radic Biol Med 46:1241–1249. https://doi.org/10.1016/j.freeradbiomed.2009.02.006
Banerjee S, Padhye S, Azmi A et al (2010) Review on molecular and therapeutic potential of thymoquinone in cancer. Nutr Cancer 62:938–946. https://doi.org/10.1080/01635581.2010.509832
Kassab RB, El-Hennamy RE (2017) The role of thymoquinone as a potent antioxidant in ameliorating the neurotoxic effect of sodium arsenate in female rat. Egypt J Basic Appl Sci 4:160–167. https://doi.org/10.1016/j.ejbas.2017.07.002
Tabeshpour J, Mehri S, Abnous K, Hosseinzadeh H (2020) Role of oxidative stress, MAPKinase and apoptosis pathways in the protective effects of thymoquinone against acrylamide-induced central nervous system toxicity in rat. Neurochem Res 45:254–267. https://doi.org/10.1007/s11064-019-02908-z
Li S, Jiang Z, Chai W et al (2019) Autophagy activation alleviates nonylphenol-induced apoptosis in cultured cortical neurons. Neurochem Int 122:73–84. https://doi.org/10.1016/j.neuint.2018.11.009
Beker M, Dallı T, Elibol B (2018) Thymoquinone can improve neuronal survival and promote neurogenesis in rat hippocampal neurons. Mol Nutr Food Res 62. https://doi.org/10.1002/mnfr.201700768
Kanter M (2008) Nigella sativa and derived thymoquinone prevents hippocampal neurodegeneration after chronic toluene exposure in rats. Neurochem Res 33:579–588. https://doi.org/10.1007/s11064-007-9481-z
Sofroniew MV (2020) Astrocyte reactivity: subtypes, states, and functions in CNS innate immunity. Trends Immunol 41:758–770. https://doi.org/10.1016/j.it.2020.07.004
Hausmann R, Rieß R, Fieguth A, Betz P (2000) Immunohistochemical investigations on the course of astroglial GFAP expression following human brain injury. Int J Legal Med 113:70–75. https://doi.org/10.1007/PL00007711
Jie Y, Xuefeng Y, Mengxue Y et al (2016) Mechanism of nonylphenol-induced neurotoxicity in F1 rats during sexual maturity. Wien Klin Wochenschr 128:426–434. https://doi.org/10.1007/s00508-016-0960-6
Maroteaux L, Campanelli JT, Scheller RH (1988) Synuclein: a neuron-specific protein localized to the nucleus and presynaptic nerve terminal. J Neurosci 8:2804–2815. https://doi.org/10.1523/jneurosci.08-08-02804.1988
Davidson WS, Jonas A, Clayton DF, George JM (1998) Stabilization of α-Synuclein secondary structure upon binding to synthetic membranes. J Biol Chem 273:9443–9449. https://doi.org/10.1074/jbc.273.16.9443
Saleh H, Saleh A, Yao H et al (2015) Mini review: linkage between α-Synuclein protein and cognition. Transl Neurodegener 4:1–6. https://doi.org/10.1186/s40035-015-0026-0
Alhebshi AH, Odawara A, Gotoh M, Suzuki I (2014) Thymoquinone protects cultured hippocampal and human induced pluripotent stem cells-derived neurons against α-synuclein-induced synapse damage. Neurosci Lett 570:126–131. https://doi.org/10.1016/j.neulet.2013.09.049
Ardah MT, Merghani MM, Haque ME (2019) Thymoquinone prevents neurodegeneration against MPTP in vivo and modulates α-synuclein aggregation in vitro. Neurochem Int 128:115–126. https://doi.org/10.1016/j.neuint.2019.04.014
Gómez-Benito M, Granado N, García-Sanz P et al (2020) Modeling Parkinson’s disease with the alpha-synuclein protein. Front Pharmacol 11:356. https://doi.org/10.3389/fphar.2020.00356
Acknowledgements
The authors wish to express their sincere gratitude to Ms. Fatemeh Shirafkan for her help in Quantitative Real-Time reaction analysis.
Funding
This work was supported by a grant (9808809) from Babol University of Medical Sciences.
Author information
Authors and Affiliations
Contributions
M. Lotfi, A. Eghbali, and A. Ebrahimpour performed the experiments, collected and analyzed data, and contributed to experimental design. S. Kazemi, F. Pourabdolhossein, and L. Satarian contributed to experimental design and its implementation. A. A. Moghadamnia, M. Lotfi, and F. Pourabdolhossein contributed to the interpretation of the data and wrote preliminary draft of the manuscript. A. A. Moghadamnia and S. Kazemi supervised the experiments. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethical Approval
All experiments and procedures of this study were reviewed and approved by the Research Ethics Committees of Babol University of Medical Sciences (IR.MUBABOL.REC.1398.026).
Consent to Participate
Not applicable.
Consent for Publication
The consent to publish this manuscript has been obtained from all authors.
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Lotfi, M., Kazemi, S., Ebrahimpour, A. et al. Thymoquinone Improved Nonylphenol-Induced Memory Deficit and Neurotoxicity Through Its Antioxidant and Neuroprotective Effects. Mol Neurobiol 59, 3600–3616 (2022). https://doi.org/10.1007/s12035-022-02807-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12035-022-02807-5