
Abstract
In the present study, we characterize the antinociceptive effects produced by the chemokine CCL4 in mice. The intraplantar administration of very low doses of CCL4 (0.1–3 pg) produced bilateral antinociception assessed by the unilateral hot-plate test (UHP) without evoking chemotactic responses at the injection site. Moreover, the subcutaneous administration of CCL4 (3–100 pg/kg) also yielded bilateral antinociception in the UHP and the paw pressure test and reduced the number of spinal neurons that express Fos protein in response to noxious stimulation. The implication of peripheral CCR5 but not CCR1 in CCL4-evoked antinociception was deduced from the inhibition produced by systemic but not intrathecal, administration of the CCR5 antagonist DAPTA, and the inefficacy of the CCR1 antagonist J113863. Besides, the inhibition observed after subcutaneous but not intrathecal administration of naloxone demonstrated the involvement of peripheral opioids and the efficacy of naltrindole but not cyprodime or nor-binaltorphimine supported the participation of δ-opioid receptors. In accordance, plasma levels of met-enkephalin, but not β-endorphin, were augmented in response to CCL4. Likewise, CCL4-evoked antinociception was blocked by the administration of an anti-met-enk antibody. Leukocyte depletion experiments performed with cyclophosphamide, anti-Ly6G, or anti-CD3 antibodies indicated that the antinociceptive effect evoked by CCL4 depends on circulating T lymphocytes. Double immunofluorescence experiments showed a four times more frequent expression of met-enk in CD4+ than in CD8+ T lymphocytes. CCL4-induced antinociception almost disappeared upon CD4+, but not CD8+, lymphocyte depletion with selective antibodies, thus supporting that the release of met-enk from CD4+ lymphocytes underlies the opioid antinociceptive response evoked by CCL4.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Chemokines constitute a group of cytokines that promotes immune cell recruitment to injured tissues by acting through G-protein-coupled receptors. The nearly 50 molecules included so far in this family are classified as CC, CXC, XC, or CX3C according to cysteine residues location at the N-terminus of the protein [1]. Their structure determines the affinity for different G-coupled protein receptors, in such a way that the corresponding receptors have been designed as CCR, CXCR, XCR, or CX3CR. Nevertheless, since no exact match exists between each chemokine and a particular receptor, functional redundancy among the effects produced by different chemokines is frequently observed [1].
Besides chemotaxis, another relevant function exerted by chemokines is the modulation of nociceptive transmission. Most of the available literature attributes a hypernociceptive role to several of these molecules. Thus, some chemokine receptors have been described to be expressed at dorsal root ganglia or CNS structures related to nociceptive transmission [2, 3]. Accordingly, peripheral [4] or spinal [5,6,7] administration of exogenous chemokines to rodents has been reported to induce hyperalgesic responses. Not surprisingly, endogenous chemokines are involved in painful responses present in several settings, such as inflammation [8, 9], neuropathy [10], or cancer [11]. This pronociceptive role has been well established in the case of CC chemokines, as it occurs with CCL1 [6], CCL3 [8], CCL5 [12], and especially, CCL2 [13]. Although less thoroughly studied, CXC chemokines, such as CXCL1 [14], CXCL5 [15], or CXCL12 [16] have also been associated with the amplification of nociceptive transmission. Finally, the unique member of the CX3CL family, fractalkine, has also hyperalgesic properties [17].
Contrasting with these data, analgesic responses induced by chemokines have also been reported [18]. Probably, the most representative examples are two members of the CXC subfamily, CXCL2/3 acting on CXCR2 [19] and CXCL10 through CXCR3 activation [20]. The analgesic action exerted by these chemokines is not due to the direct inhibition of nociceptive neurons but mediated by the release of endogenous opioids from several types of leukocytes [19, 20]. These chemokine-induced analgesic effects have been demonstrated by the inhibition of local hypernociceptive responses in damaged tissues and are due to the participation of immune cells attracted in response to injury. However, a tentative classification of chemokines as pronociceptive or antinociceptive does not seem advisable, taking into account that the aforementioned analgesic chemokines, CXCL2 and CXCL10, can also produce nociceptive responses. In particular, CXCL2 can contribute to neuropathic pain in mice with partial sciatic nerve ligation [21] and CXCL10 can act as algogenic in an experimental model of bone cancer-induced pain [22].
Concerning the dual effect displayed by some chemokines, we have recently communicated that the acute intraplantar administration of low doses of CCL5, a CCR1 and CCR5 agonist, induces thermal hyperalgesia in mice, whereas a slight increase of the dose can prevent it by evoking the local release of the endogenous κ-opioid agonist dynorphin from neutrophils [23]. In the present experiments, we aimed to assess the role on nociception of another CC chemokine that, as CCL5, is a CCR5 agonist. Thus, we have measured the effects induced by the peripheral administration of CCL4, also called Macrophage Inflammatory Protein 1β (MIP-1β) in healthy mice. This chemokine can activate the migration of eosinophils [24], neutrophils [25], lymphocytes [26], and more weakly, monocytes [27]. Regarding pain modulation, a previous report describes that perineural administration of CCL4 to rats evoked hypernociceptive effects 1 day after treatment [28]. Conversely, our results describe that the exogenous administration of very low doses of CCL4 can produce antinociceptive responses that are not restricted to the site of the injection. The possibility that CCL4 could be related to analgesic responses is so far an almost unexplored possibility and only a recent publication proposed that the antihyperalgesic effect evoked by the administration of bone marrow stromal cells in rats is linked to the action of CCL4 at CNS [29]. In this manuscript, we study the mechanisms involved in the peripheral antinociceptive effect evoked by the exogenous administration of CCL4 in mice.
Material and Methods
Animals
Swiss CD-1 male mice 6–8 weeks old, bred (6–8/cage) in the Animalario de la Universidad de Oviedo (Reg. 33044 13A) in a temperature-controlled room, on a 12-h dark–light cycle with free access to food and water were used. Behavioral experiments were performed during the light cycle in a quiet room, and all efforts were made to limit distress and to use minimal number of animals required to produce scientifically reliable data. Protocols were approved by the Comité Ético de Experimentación Animal de la Universidad de Oviedo (Spain) and performed according to the guidelines of European Communities Council Directive (2010/63/EU) for animal experiments.
Drugs
CCL4 (ProSpec) was disolved in 0.1% bovine serum albumin in distilled water. The CCR1 antagonist J113863 (Tocris) was dissolved in 10% dimethyl sufoxide in distilled water. The CCR5 antagonist DAPTA (Tocris), the non-selective opioid receptor antagonist naloxone (Tocris), the selective μ-opioid receptor antagonist cyprodime (Tocris), the selective δ-opioid receptor antagonist naltrindole (Tocris), the selective κ-opioid receptor antagonist nor-binaltorphimine (Tocris), the immunosuppressant agent cyclophosphamide (Sigma-Aldrich), the IL-1 receptor antagonist anakinra (Sigma-Aldrich), the CRF receptor antagonist α-helical CRF (9–41) (Tocris), and the anti-met-enkephalin (met-enk) antibody (ab1975, Millipore) and its corresponding control, IgG from rabbit serum (Sigma-Aldrich), were dissolved in saline.
In depletion experiments, 50 mg/kg of cyclophosphamide were intraperitoneally (i.p.) injected twice 72 and 24 h before testing. The anti-Ly6G antibody (100 μg, BioXCell) was i.p. injected 18 h before testing [30], whereas the anti-CD3 (1 μg, BioLegend), anti-CD4 (30 μg, Stemcell), or anti-CD8α (20 μg, SouthernBiotech) antibodies were i.p. administered 24 h before testing [31]. These antibodies and their corresponding controls (rat IgG2a isotype for anti-Ly6G and anti-CD8α antibodies or rat IgG2b for anti-CD3 and anti-CD4 antibodies) were dissolved in phosphate-buffered saline (PBS).
Subcutaneous (s.c.) administration under the fur of the neck and i.p. injections were performed in a volume of 10 ml/kg. Intraplantar (i.pl.) injections consisted in the administration of 25 μl generally into the right hind paw and in a particular experiment into the forepaw under isoflurane anesthesia (3%, Isoflo®, Esteve). For intrathecal (i.t.) injections, mice were anesthetized with isoflurane (3%, Isoflo®, Esteve) and a small cut in the skin at the dorsal lumbar level was done. Next, the tip of a 30-gauge needle inserted in a Hamilton microsyringe was introduced at the level of L5–L6, and finally, a volume of 5 μl was injected [32]. Intravenous (i.v.) injections were performed into a tail vein in a volume of 100 μl. In all cases, control mice received an injection of the corresponding solvent or the appropriate IgG isotype.
CCL4 was i.pl. or s.c. administered at different times before testing, although the majority of experiments were performed 1 h after its s.c. administration. The s.c. administration of J113863 or DAPTA was performed 30 min before testing. Naloxone was s.c. administered 15 min before testing. The i.t. administration of DAPTA or naloxone was done 30 min before testing. Cyprodime, naltrindole, and nor-binaltorphimine were s.c. administered 30 min before testing. Anakinra was i.p. administered 1 h before testing. α-Helical CRF (9–41) or the anti-met-enk antibody were i.v. administered 60 or 30 min before testing, respectively.
Nociceptive Tests
Unilateral UHP
As previously described [23], mice were gently restrained and the plantar side of the tested paw was placed on a hot plate (IITC, Life Science). The temperature of the plate was 49 °C. Measurements of withdrawal latencies from the heated surface of each hind paw were made separately at 2-min intervals, and the mean of two measures was considered. A cut-off of 30 s was established. In initial experiments, the value obtained in each paw was considered separately, but once observed, the bilateral effect evoked by CCL4, the mean of measures obtained in both paws was used.
von Frey Test
Mechanical allodynia was assessed by applying von Frey filaments (Stoelting) to the plantar side of the paws, as previously reported [33]. Mice were placed on the wire mesh platform during 60 min for habituation. The von Frey filaments 2.44, 2.83, 3.22, 3.61, 4.08, and 4.56 (corresponding to the application of 0.028, 0.068, 0.166, 0.407, 1.202, and 3.63 g bending forces) were used, and starting with the 3.61 filament, six measurements were taken in each animal randomly starting with the left or right paw. Based on the “up and down” method [34], the observation of a positive response (lifting, shaking, or licking of the paw or marked toe spreading) was followed by the application of the immediate thinner filament or the immediate thicker one if the response was negative. The 50% response threshold was calculated using the following formula: 50% g threshold = (10Xf + κδ)/10,000; where Xf is the value of the last von Frey filament applied; κ is a correction factor based on pattern of responses (from the Dixon’s calibration table); and δ is the mean distance in log units between stimuli (here, 0.4).
Paw Pressure Test
Mechanical nociceptive thresholds were measured by using a paw pressure test adapted for mice [35] from the original method described in rats by Randall and Selitto [36]. Mice were gently restrained, and both hind paws were separately exposed to a linearly increased pressure from 0 to 750 g in 15 s (Ugo Basile 7200). When mice struggled to withdraw the corresponding paw, the stimulus was stopped and the pressure value recorded. One measure was taken in each hind paw and their mean was considered.
Actimetry
Mice were exposed for 15 min periods to testing environment during the previous 3 days. On the experimental day, mice received the s.c. administration of CCL4 or saline and were immediately placed in the actimeter (Letica LE 8811). Horizontal locomotor activity was automatically recorded in four periods of 5 min from 50 to 70 min after drug administration.
Blood Extraction Procedure
Mice anesthetized with isoflurane (3%, Isoflo®, Esteve) were manually restrained by grasping the loose skin over the shoulders and the facial vein was punctured slightly behind the mandible in front of the ear canal with a 25-gauge needle by a swift lancing movement. A volume of 200–300 μl was usually harvested.
β-Endorphin and Met-Enk Enzyme-Linked Immunosorbent Assay
Blood was collected in an Eppendorf tube containing 10 μl of EDTA (0.5 M, pH = 8) and centrifuged (1000 g, 15 min, 4 °C) to obtain plasma. To prevent β-endorphin (β-end) degradation aprotinin (0.6 trypsin inhibitor units/ml, Sigma-Aldrich) was added, and in the case of met-enk, a mixture of bestatin (500 μM, Tocris), captopril (100 μM, Tocris), and tiorfan (100 μM, Sigma-Aldrich). Commercial enzyme-linked immunosorbent assay (ELISA) kits were used to determine plasma levels of β-end (Elabscience) and met-enk (MyBioSource). Following manufacturer recommendations, 50 or 100 μl of plasma were used to quantify β-end or met-enk, respectively. In both cases, plates were read at 450 nm (PowerWave™, BioTek). The procedure was repeated in at least three independent assays with samples coming from different mice.
White Cell Count and Flow Cytometry Analysis
White Cell Count
Blood was collected in an Eppendorf tube containing 10 μl of EDTA (0.5 M, pH = 8). A differential hematology analyzer based in the measure of cell impedance when passing through a small aperture with a laser optical technique (Abacus Junior Vet, Diatron) was used to quantify white cell populations. The apparatus gives separated counts of the total number of white blood cells as well as the number of lymphocytes, granulocytes (mainly neutrophils but also basophils and some eosinophils), and mid-size cells including monocytes together with some eosinophils.
Flow Cytometry
Three hundred microliters of blood was collected in EDTA-coated tubes. Blood samples were incubated with 3 ml of red blood cell lysis buffer (NH4Cl 150 mM, NaHCO3 10 mM, and EDTA 1 mM, pH = 7.4) for 10 min on ice. After erythrocyte lysis, blood suspension was centrifuged at 1200 rpm for 5 min at 4 °C. The supernatant was removed and the lysis procedure was repeated for 5 min. After centrifugation, the pellet was resuspended in 100 μl PBS + 10% fetal bovine serum (FBS, Fisher Bioblock) for cell staining. Leukocytes were stained with anti-CD3-FITC (clone 145-2C11, BioLegend), anti-CD4-PE (clone GK 1.5, eBioscience), and anti-CD8-Alexa Fluor 700 (clone 53–6.7, BioLegend) for 30 min on ice. After washing, DAPI was added to label dead cells. Samples were analyzed using a MoFlo XDP flow cytometer (Beckman Coulter) and Summit software 5.2 version. CD4+ and CD8+ T cells were reported as percentages of CD3+ lymphocytes after excluding dead cells and cellular debris based on DAPI staining and forward and side scatter profiles.
Tissue Processing and Immunohistochemical Assays
Hind Paws
Immunohistochemical experiments addressed to identify neutrophils, macrophages, and lymphocytes were performed in plantar skin paws fixed with 4% formol for 4 h, washed under tap water, immersed in 50% ethanol overnight, and finally, embedded in paraffin. Immunohistochemical assays were performed on deparaffinized cross sections (4 μm) with the corresponding antibody. Samples were exposed to the anti-myeloperoxidase antibody (polyclonal rabbit anti-mouse, A0398 DAKO, 1:500) for 30 min at room temperature (RT) after antigen retrieval in buffer solution (pH = 6) for 20 min (PTLink, Dako). The incubation with the anti-F4/80 antibody (polyclonal rat anti-mouse, sc71086 Santa Cruz, 1:200, 30 min, RT + polyclonal rabbit anti-rat immunoglobulins/horseradish peroxidase, DAKO) was performed after antigen retrieval in buffer solution (pH = 6) for 20 min (PTLink, Dako), and the incubation with the anti-CD3 antibody (polyclonal rabbit anti-mouse, ab5690 Abcam, 1:75) lasted 30 min at RT. Sections were next incubated in secondary antibody Envision Rabbit (k4002 Dako) 30 min at RT, and stained with 3–30-diaminobenzidine (Dako). Finally, they were counterstained for 10 min with hematoxylin (Dako). The specificity of the staining was checked with negative controls that included tissues unexposed to primary antibodies. Images were taken through a × 40 objective of an Olympus BX63 microscope and recorded on an Olympus DP73 camera. The number of stained cells were counted in three fields of microphotographs done at × 40 (23,198 μm2/area) from each paw tissue taken from regions containing epidermis, dermis, and subcutaneous tissue of at least four different mice per group.
Blood Smears
Blood collection and red blood cells lysis were performed as explained above. The pellet was suspended in PBS (30 μl, pH = 7.4), and 5 μl of this cell suspension was spread on gelatin-coated slides (Super-Frost Plus, Menzel-Glaser) with a yellow tip sized for 20–200 μl micropipettes. Slides were dried (2 h, RT), fixed (2 min, RT) in acetone:methanol:40% formaldehyde (19:19:2), washed (X2; 10 min) in Milli-Q water, and finally, dried (2 h, RT) and stored at − 20 °C [37].
For met-enk immunofluorescence assays, slides were initially immersed during 30 min in PBS-T (PBS 0.01 M with 0.01% of Triton X-100, Sigma-Aldrich) and incubated (in a humid chamber) with a rabbit anti-met-enk antibody (AB1975, Millipore, 1:100 in 0.01 M PBS) for 1 h at RT. Slides were washed in PBS-T (30 min, RT) and further incubated with secondary antibody Alexa Fluor 488-conjugated goat anti-rabbit IgG (A-11034, Invitrogen, 1:250 in 0.01 M PBS) for 1 h at RT.
For double immunolabeling assays, slides were initially incubated with the rat anti-CD4 (1540-01, SouthernBiotech, 1:100) or anti-CD8α (1550–01, SouthernBiotech, 1:100) antibodies in 0.01 M PBS for 1 h at RT. Next, slides were immersed 30 min in PBS and incubated (1 h, RT) with the corresponding secondary antibody (goat anti-rat IgG-TR, sc-2782, Santa Cruz, 1:200; PBS). Then, after washing 15 min with PBS and 30 min with PBS-T, the staining procedure with the anti-met-enk antibody was performed as explained above.
Finally, slides were washed in PBS (2 × 15 min) and mounted with DAPI Fluoromount-G™ (SouthernBiotech). Immunostained samples were imaged using a BX61 Olympus microscope with a × 20/0.75 NA objective. Images were acquired with a CCD camera Olympus DP-70 using blue-excitation (BP470–490) and green-excitation (BP530-550) filters and the software Olympus DP-controller 1.2.1.108 and Olympus DP-manager 1.2.1.107. In double- staining assays, the green color represents the fluorescence of the anti-met-enk antibody and red corresponds to the fluorescence of anti-CD4 or anti-CD8 antibodies. Merged images were obtained by using the ImageJ software. The percentage of white cells immunoreactive to met-enk, CD4 or CD8 was calculated in each smear. With this aim, five random photographs were taken from each slide and the number of cells clearly positive for the corresponding staining as well as the number of DAPI-stained nuclei were counted independently by two members of the lab. The mean of these independent measures, always showing an error of less than 15%, was considered. Thus, the percentage of met-enk-, CD4-, or CD8-stained cells was calculated in relation to the number of DAPI-stained cells in each slide. Each smear came from blood of a mouse and assays were performed in at least four slides coming from different mice.
Lumbar Spinal Cord
Mice received the s.c. administration of CCL4 or solvent. One hour later, they were anesthetized (3% isoflurane, Isoflo®, Esteve) and were submitted to a noxious stimulus consisting in placing the right paw for 30 s on the hot plate surface set at 49 °C in order to evoke Fos expression in the dorsal horn of the lumbar spinal cord. Mice were killed 60 min after noxious stimulation by cervical dislocation under isoflurane anesthesia and lumbar spinal cords were removed. Next, they were fixed at 4 °C during 24 h in 4% paraformaldehyde and cryoprotected by immersion in 15% sucrose in PBS 0.01 M for 12–24 h and in 30% sucrose for 24 h. A minimal longitudinal cut along the contralateral (left) ventral white matter of the fixed spinal cord was made to serve further as a reference of the ipsi- and contralateral sides in sections. Fifty-micrometer-thick sections of lumbar segments L4–L6 were obtained using a freezing microtome (Microm HM430). Free-floating sections were washed three times in 0.01 M PBS for 10 min, blocked 90 min with 10% FBS containing 0.1% Triton X-100 (Sigma-Aldrich), and further incubated overnight at 4 °C in a humid chamber with rabbit anti-Fos antibody (sc-52, Santa Cruz, 1:500). Next, sections were rinsed three times for 10 min in 0.01 M PBS and incubated for 2 h at room temperature with green Alexa Fluor 488-conjugated goat anti-rabbit IgG (A11034, Life Technologies, 1:500). Slices were collected on gelatin-coated slides (Super-Frost Plus, Menzel-Glaser) and mounted with Fluoromount-G (SouthernBiotech).
Immunostained tissue sections were imaged using a BX61 Olympus microscope with × 4/0.16NA and × 10/0.40NA objectives. Images were acquired with a CCD camera Olympus DP-70 using blue-excitation filter (BP470-490) and processed using the software Olympus DP-controller 1.2.1.108 and Olympus DP-manager 1.2.1.107. The five slices of each spinal cord segment with more Fos-IR neurones were selected. Then, the Fos-IR nuclei of each slice were independently counted by two members of the lab. The mean of these two measures (always showing an error less than 15%) was considered. In each experimental group, the spinal cords of at least five animals were included.
Statistical Analysis
Mean values and their corresponding standard errors were calculated in values of parameters of behavioral assays, ELISA measurements, immunofluorescence experiments, and white blood cell counts. Comparisons of results obtained by the von Frey test were performed by an initial Kruskal-Wallis test followed by the Dunn’s test to achieve specific differences among groups. Data obtained in all other assays were treated as normally distributed and parametric tests applied for their comparisons. When two different variables were considered (for example, withdrawal latencies of right or left paws and different drug treatments in the UHP test after i.pl. administration of drugs) an initial two-way analysis of variance (ANOVA) analysis was followed by Bonferroni’s correction to specifically compare values obtained in the different groups. When only one variable was considered (generally, treatment), one-way ANOVA analysis was initially performed and followed by the Dunnett’s t test to compare effects induced by different doses of the same drug or the Newman-Keuls test to compare the effects induced by different treatments. Comparisons between two groups were done by the unpaired Student’s t test. The criterion for statistical significance was P < 0.05.
Results
Low Doses of CCL4 Evoke Bilateral Antinociception Measured the Unilateral Hot-Plate Test in Mice
The i.pl. administration of very low doses of CCL4 (0.1–3 pg/mouse) into the right hind paw of mice 60 min before testing provoked dose-dependent increases in thermal withdrawal latencies in both hind paws measured by the UHP, being the effect statistically significant upon the administration of both 1 and 3 pg of CCL4 (Fig. 1a). As shown in Fig. 1b, the antinociceptive effect evoked by the i.pl injection of 3 pg of CCL4 in one hind paw was again obtained in both hind paws, 30 and 60 min after the injection of the chemokine. This effect completely disappeared 2 h after CCL4 administration.

Antinociceptive response evoked by the intraplantar (i.pl.) administration by a low dose of CCL4 devoid of chemotactic effect and mechanical allodynia induced by a higher dose. a Antinociceptive effects measured in mice by the unilateral hot plate test after the i.pl. administration of CCL4 (0.1–3 pg) into the right hind paw (N = 7 in solvent and 1-pg-treated groups, N = 5 in 0.3- and 3-pg-treated groups and N = 8 in 0.1-pg-treated group). b Time course of the antinociceptive effect evoked by the i.pl. administration of 3 pg of CCL4 into the right hind paw (N = 7 in solvent-treated group and N = 5 in 3-pg-treated group). In (a) and (b), data are expressed as means ± SEM. A two-way ANOVA (treatment × paws) yielded differences dependent on the dose of CCL4 but not on the paw injected; **p < 0.01, compared with the ipsilateral paw of solvent-treated group, Bonferroni’s correction. c c1, Representative microphotographs of immunohistochemical staining of neutrophils with an anti-myeloperoxidase (MPO) antibody performed in paws of mice-treated i.pl. with solvent 1 h before, an antinociceptive dose of CCL4 (3 pg, 1 h before), or a higher dose of this chemokine (30 ng, 4 h before). In all cases, the scale bar represents 20 μm. c2–c3, Number of lymphocytes (LYMPH), macrophages (MACR), or neutrophils (NEUT) detected by using the selective anti-CD3, anti-F4/80, or anti-myeloperoxidase antibodies, respectively, in a field corresponding to an area of 23,106 μm2 of the paw subcutaneous tissue of mice i.pl. injected 1 h before with solvent or CCL4 (3 pg) or 4 h before with solvent or CCL4 (30 ng). Data are expressed as means ± SEM (N = 4). *p < 0.05, **p < 0.01, compared with the corresponding solvent-treated group, Student’s t test. d Lack of effect in the von Frey test of the i.pl. administration of a dose of CCL4 able to evoke maximal antinociception in the UHP test (3 pg; 1 h before testing) and allodynic effect produced 1 h after the i.pl. administration of 100 ng of CCL4 into the right hind paw. Data are expressed as means ± SEM (N = 8 in solvent-treated group, N = 9 in 3-pg-treated group and N = 6 in 100-ng-treated group). **p < 0.01, compared with the ipsilateral paw of solvent-treated group, Dunn’s test
In order to elucidate whether this antinociceptive effect could be related to the chemotactic properties of CCL4, immunohistochemical assays were performed in paws 1 h after the i.pl. administration of solvent or 3 pg of this chemokine. The staining of lymphocytes, macrophages or neutrophils with anti-CD3, anti-F4/80, or anti-MPO antibodies, respectively, showed that CCL4 does not evoke a chemotactic response when compared with saline (Fig. 1c (c1–c2)). In contrast, the i.pl. administration of a 10,000-fold higher dose of CCL4 (30 ng) produced a remarkable presence of neutrophils (Fig. 1c (c3)) and a slight increase in the number of macrophages in the injected paw, being this response only observed 4 h, and not 1 h (data not shown) after the i.pl. injection (Fig. 1c (c3)).
Since a previous report described the instauration of allodynia following the perineural administration of CCL4 at doses in the range of nanograms [28], we assessed by the von Frey test the effect of the i.pl. administration of an antinociceptive dose (3 pg) or a higher one close to that reported as allodynic (100 ng). As shown in Fig. 1d, the 50% mechanical threshold remained unaltered 1 h after the administration of 3 pg of CCL4, whereas unilateral mechanical allodynia was observed in mice receiving the i.pl. administration of 100 ng of CCL4 (Fig. 1d).
As above mentioned, considering that the antinociceptive effect produced by the i.pl. administration of low doses of CCL4 was observed not only in the injected paw but also in the contralateral one, we have explored whether CCL4 could also evoke this effect when administered in a more distal site, such as the forepaw. As occurred when injected into the hind paw, the antinociceptive effect was detected in both hind paws 30 and 60 min after the administration of 3 pg of CCL4 (Fig. 2a). Accordingly, the s.c. administration of CCL4 under the fur of the neck, the most standard site for s.c. drug administration in rodents, induced dose-dependent increases of withdrawal latencies. Thus, the administration of 10, 30, and 100 pg/kg (corresponding to 0.3, 1 and 3 pg for a 30g mouse) significantly increased withdrawal latencies in both paws 30 and 60 min after drug injection (Fig. 2b). On the basis of this result, CCL4 was always s.c. administered 1 h before testing in subsequent experiments.

Antinociceptive responses produced by CCL4 administered either in a forepaw or under the fur of the neck and lack of effect on locomotor activity. a Antinociceptive effects measured in mice by the unilateral hot plate test after the administration of 3 pg of CCL4 into the right forepaw (N = 5 for solvent-treated and N = 6 for 3pg-treated group). Data are expressed as means ± SEM. A two-way ANOVA (treatment × paws) yielded only statistically significant differences dependent on the treatment received. **p < 0.01, compared with the ipsilateral paw of solvent-treated group, Bonferroni’s correction. b Antinociceptive effects measured in mice by the unilateral hot plate test after the administration of CCL4 (3–100 pg/kg) under the fur of the neck (N = 4 in all groups). c Antinociceptive effects measured in mice by the paw-pressure test 1 h after the s.c. administration of 3–100 pg/kg of CCL4 (N = 12 in solvent group, N = 7 in 3pg/kg- and 10pg/kg-treated groups, N = 11 in 30pg/kg-treated group, and N = 6 in 100pg/kg-treated group). In (b) and (c), data are expressed as means ± SEM. *p < 0.05, **p < 0.01, compared with the solvent-treated group, Dunnett’s t test. d Locomotor horizontal activity measured in an actimeter in four 5-min periods after the s.c. administration of saline or 30 pg/kg of CCL4 50 min before (N = 8 in both groups). Data are expressed as means ± SEM. An initial two-way ANOVA (time × treatment) yielded differences related to time passed in the actimeter but not on the treatment received
In order to assess if CCL4 could also be antinociceptive against mechanical noxious stimuli, the paw pressure test was used. This method is appropriate to measure drug-induced analgesic responses since it is based on the application of overt mechanical noxious stimuli, opposite to the von Frey test, in which light, non-noxious, stimuli are applied in order to detect allodynia. The administration of 3–100 pg/kg of CCL4 1 h before testing provoked dose-dependent antinociception measured by the paw pressure test, being mechanical withdrawal thresholds significantly greater after the administration of 30 and 100 pg/kg (Fig. 2c), as occurred with thermal latencies in the UHP test.
Aiming to evaluate whether behavioral responses in nociceptive tests could be biased by sedative or unspecific effects, we have measured the influence of 30 pg/kg of CCL4 in the locomotor activity measured in an actimeter. Although the activity of mice in the actimeter decreased with time, the treatment applied, solvent or CCL4, did not influence their activity at the different times studies (Fig. 2d).
The ability of CCL4 to modulate the excitability of nociceptive spinal neurons was explored by studying the expression of the protein Fos in laminae I–II of the spinal cord. The nociceptive stimulus applied to mice consisted in a 30s period of contact of the plantar side of the right hind paw with the hot plate (49 °C) under isofluorane anesthesia. As observed in Fig. 3, the number of Fos immunoreactive (IR) cells detected in the contralateral side was scarce either in solvent- or CCL4-treated mice but significantly increased in the side ipsilateral to thermal stimulation of solvent-treated mice. In contrast, the number of Fos-IR cells in the ipsilateral side of mice treated with 30 pg/kg of CCL4 was significantly reduced compared with Fos-IR in the ipsilateral dorsal horn of mice treated with solvent.

Inhibition of the spinal expression of Fos protein obtained in response to noxious thermal stimulation following the administration of an antinociceptive dose of CCL4. Inhibition of the expression of Fos protein in the lumbar spinal cord evoked by CCL4 (30 pg/kg; s.c.; 1 h before) in mice receiving a nociceptive stimulus consisting in the contact of the plantar side of the right hind paw with the hot plate at 49 °C for 30 s. a Data are expressed as means ± SEM of the number of Fos immunoreactive cells observed in laminae I–II of the ipsilateral (right) or contralateral (left) side of the spinal cord of mice treated with solvent (N = 6) or CCL4 (N = 5). A two-way ANOVA (treatment × ipsi/contralateral side of the dorsal horn) showed differences dependent on both variables. ●●p < 0.01, compared with the corresponding contralateral dorsal horn, **p < 0.01, compared with the ipsilateral dorsal horn of solvent-treated group, Bonferroni’s correction. b, c Representative examples of Fos expression obtained in the spinal cord of mice treated either with solvent (b) or CCL4 (c). In both cases, the inset corresponds to the amplification of the external layer of the dorsal horn ipsilateral to the nociceptive stimulus
Involvement of CCR5 and Opioid Receptors in the Antinociceptive Effect Produced by CCL4
Since CCL4 can bind to CCR1 or CCR5 receptors, we have assayed the effect of selective antagonists for these receptors. The antinociceptive response evoked by CCL4 (30 pg/kg; s.c.; 1 h before) remained unmodified when administered together with 30 mg/kg of the CCR1 antagonist J113863 (Fig. 4a), whereas the coadministration of the CCR5 antagonist DAPTA (1–10 mg/kg) inhibited it in a dose-dependent manner (Fig. 4b).

Inhibition of the antinociceptive effect evoked by CCL4 after the systemic administration of CCR5 or δ-opioid receptor antagonists. a–f Modulation of the analgesia evoked in the unilateral hot plate test by CCL4 (30 pg/kg; s.c.; 1 h before) produced by: a the systemic administration of the CCR1 antagonist J113863 (J11, N = 6 for CCL4 + solvent, N = 5 for the other experimental groups), b the CCR5 antagonist (DAPTA, N = 5 for all groups), c the opioid receptor antagonist naloxone (NAL, N = 5 for solvent+solvent, solvent + NAL3 and CCL4 + NAL3, N = 6 for CCL4 + NAL1.7, and N = 7 for resting groups), d the μ-opioid receptor antagonist cyprodime (CYP, N = 8 for CCL4 + solv and N = 5 for the other groups), e the δ-opioid receptor antagonist naltrindole (NTI, N = 8 for solvent + solvent and CCL4 + solvent, N = 5 for CCL4 + NTI 0.3, and N = 9 for the resting groups), f the κ-opioid receptor antagonist nor-binaltorphimine (n-BNI, N = 8 for CCL4 + solvent and N = 5 for the resting groups). g Inhibition produced by s.c. administration of DAPTA or naltrindole (NTI) on the analgesia evoked by 30 pg/kg of CCL4 in the paw pressure test (N = 6 for solvent- and CCL4 + DAPTA-treated groups, N = 7 for CCL4-treated group, and N = 5 for CCL4 + NTI-treated group). Data are expressed as means ± SEM. *p < 0.05, **p < 0.01, compared with solvent-treated group, ●p < 0.05, ●●p < 0.01 compared with CCL4-treated group, Newman-Keuls test
Based on the previous description that the analgesic effect produced by other chemokines can be related to endogenous opioid peptides, we assessed the effect of opioid receptor antagonists on CCL4-evoked antinociception. The administration of the non-selective opioid receptor antagonist naloxone (1–3 mg/kg; s.c.; 15 min before testing) dose-dependently inhibited the response evoked by CCL4 (30 pg/kg; s.c.) in the UHP test (Fig. 4c). In addition, CCL4-evoked antinociception was also prevented by the δ-opioid receptor antagonist naltrindole (0.3–1 mg/kg; s.c.; 30 min before; Fig. 4e) but not the μ- or κ-opioid receptor antagonists cyprodime (10 mg/kg; s.c.; 30 min before testing Fig. 4d) or nor-binaltorphimine (30 mg/kg; s.c.; 30 min before testing Fig. 4f), respectively.
In accordance with a common mechanism underlying thermal and mechanical antinociception, the s.c. administration of 10 mg/kg of DAPTA or 1 mg/kg of naltrindole also inhibited the effect evoked by 30 pg/kg of CCL4 (s.c.; 1 h before) measured in the paw pressure test (Fig. 4g).
Finally, the possible mediation of spinal mechanisms in the effect evoked by systemic CCL4 was evaluated by testing the effect of spinally administered DAPTA or naloxone. The i.t. administration of 5 μg of DAPTA (Fig. 5a) or 10 μg of naloxone (Fig. 5b) did not modify the efficacy of CCL4, suggesting the lack of involvement of spinal mechanisms in its antinociceptive effect.

The intrathecal administration of either a CCR5 or an opioid receptor antagonist does not counteract antinociceptive responses evoked by systemic CCL4. Lack of effect of the intrathecal (i.t.) administration of the CCR5 antagonist DAPTA (a) or the opioid receptor antagonist naloxone (NAL, b) on the analgesia evoked in the UHP test by CCL4 (30 pg/kg; s.c.; 1 h before testing). In (a), N = 5 for all groups; in (b), N = 6 for CCL4-treated group and N = 5 for the resting groups. Data are expressed as means ± SEM. **p < 0.01, compared with solvent-treated group, Newman-Keuls test
Involvement of Met-Enk in CCL4-Induced Antinociception
Considering the participation of opioid receptors in the antinociceptive effects evoked by CCL4, we evaluated if this response could be affected by two drugs able to modulate the release of opioid peptides. Thus, the administration of α-helical-CRF (9–41) (0.5 mg/kg; i.v.; 60 min; Fig. 6a), a drug related to the release of both β-end or met-enk, inhibited CCL4-induced antinociception. In contrast, the effect evoked by CCL4 remained unaltered after the administration of 100 mg/kg of anakinra (i.p.; 60 min; Fig. 6b), an antagonist of IL-1 receptors previously related to the release of β-end and dynorphin, but not enkephalins [38].

Behavioral, ELISA, and immunofluorescence assays suggesting the involvement of met-enk in CCL4-evoked analgesia. a, b Effect of the systemic administration of α-helical-CRF (9–41) (α-He-CRF, a, N = 6 for CCL4 + α-He-CRF and N = 5 for the other groups) or anakinra (ANAK, b N = 5 for anakinra-treated group and N = 6 for the other groups) on the analgesia evoked by CCL4 (30 pg/kg; s.c.; 1 h before testing) in the UHP test. c Plasma concentrations of β-endorphin (β-end) (N = 6 per group) and met-enk (N = 15 per group) measured by ELISA in blood samples of mice treated 1 h before with the s.c. administration of solvent or 30 pg/kg of CCL4. d Dose-dependent inhibition of the analgesia induced by 30 pg/kg of CCL4 (s.c.; 1 h before testing) produced by the i.v. administration 30 min before testing of an anti-met-enk (anti-ME) antibody or a rabbit IgG used as control (N = 5 for anti-ME-treated group, N = 7 for the CCL4 + anti-ME 0.4 group, and N = 6 for the other groups). Data are expressed as means ± SEM. In (a), (b), and (d), *p < 0.05, **p < 0.01, compared with solvent-treated group, ●●p < 0.01 compared with CCL4-treated group, Newman-Keuls test. In (c), **p < 0.01, compared with solvent-treated group, Student’s t test. e–f Microphotographs of an immunofluorescence experiment performed with an anti-met-enk antibody (green) in a murine blood smear. Cell nuclei are stained with DAPI (blue). g Diagram showing the percentage of white blood cells expressing met-enk in mice blood smears (N = 7)
ELISA experiments were performed in blood samples to determine the possible release of β-end or met-enk. As shown in Fig. 6c, β-end blood levels measured in control mice treated with solvent were in the range of 80 pg/ml, a value that was not significantly altered 1 h after the s.c. administration of 30 pg/kg of CCL4. Besides, the levels of met-enk obtained in blood of mice treated with 30 pg/kg of CCL4 1 h before were significantly higher than those obtained in solvent-treated mice (Fig. 6c).
In order to check if the increased levels of met-enk in response to CCL4 administration could be related to the effect evoked by this chemokine, we explored the in vivo effect of an anti-met-enk antibody. As shown in Fig. 6d, the acute i.v. administration of 0.4–1.6 μg/kg of the met-enk antibody 30 min before testing, but not of control IgG from rabbit serum, dose-dependently inhibited CCL4-evoked antinociception, being the effect of the chemokine completely abolished after the administration of the higher dose assayed. The administration of 1.6 μg/kg of the antibody alone did not produce any alteration in paw withdrawal latencies (Fig. 6d).
Since met-enk can be released from immune cells, we next studied if this opioid peptide could be detected in blood cell smears. Blood smears containing white cells coming from seven different mice were used to perform immunofluorescence assays with an anti-met-enk antibody. Percentages of positive cells were calculated separately in each assay yielding a mean value of about 7% from a total number of 3051 DAPI-stained cells (Fig. 6g).
Met-Enk-Related CCL4-Induced Antinociception Is Dependent on CD4+ Lymphocytes
Considering the involvement of met-enk in the effect of CCL4 and its expression in white blood cells, we assessed whether the antinociceptive response evoked by this chemokine could be affected by the reduction in circulating cells following different depleting treatments. Initially, the i.p. administration of 50 mg/kg of the immunosuppressant agent cyclophosphamide 72 and 24 h before testing produced a drastic reduction of all type of cells. A significant reduction of about 70% was measured in either lymphocytes, mid-sized cells, mainly corresponding to monocytes, and granulocytes (Fig. 7a). Interestingly, the effect produced by CCL4 administration was significantly reduced when mice were previously treated with cyclophosphamide (Fig. 7d).

Depletion of lymphocytes prevents CCL4-evoked antinociception. a–c Effect of the treatment with either cyclophosphamide (a; CYCL), anti-Ly6G antibody (b), or anti-CD3 antibody (c) or their corresponding controls (saline, IgG2a, or IgG2b, respectively) on the total number of circulating white blood cells, lymphocytes (LYMPH), mid-size cells (mainly monocytes), and granulocytes (GRAN; mainly neutrophils). In (a), N = 8 per group; in (b), N = 13 for solvent-treated and N = 12 for anti-Ly6G-treated; in (c), N = 7 for solvent-treated and N = 8 for anti-CD3-treated. Data are expressed as means ± SEM. In (a), the two-way ANOVA (cell type x treatment) raised differences depending on treatment but not the particular cell type, whereas in (b) and (c), differences were both related to cell type and treatment administered. **p < 0.01, compared with control group, Bonferroni’s correction. d–f Effects on the CCL4-evoked analgesia (30 pg/kg; s.c.; 1 h before testing) of the depletion produced by cyclophosphamide (d; CYCL), anti-Ly6G antibody (e), or anti-CD3 antibody (f). In (d), N = 8 for each group; in (e), N = 13 for solvent- and CCL4-treated groups and N = 12 for anti-Ly6G + CCL4-treated and anti-Ly6G-treated group; in (f), N = 7 for solvent- and CCL4-treated groups and N = 8 for anti-CD3 + CCL4-treated and anti-CD3-treated group. Data are expressed as means ± SEM. **p < 0.01, compared with solvent-treated group, ●●p < 0.01 compared with CCL4-treated group, Newman-Keuls test
Selective leukocyte depleting experiments were performed with anti-Ly6G and anti-CD3 antibodies to elucidate which particular type(s) of white blood cells could be involved in the antinociceptive effect of CCL4. In particular, mice treated with 100 μg of anti-Ly6G antibody 18 h before testing showed a reduction of granulocyte and monocyte populations that diminished up to approximately 30 and 45% of the respective control values obtained in mice treated with the respective antibody isotype IgG2a, whereas the population of lymphocytes remained unaltered (Fig. 7b). In spite of this marked reduction in granulocyte and monocyte number, the s.c. administration of 30 pg/kg of CCL4 produced a similar antinociceptive effect in depleted and non-depleted mice (Fig. 7e). The administration of 1 μg of anti-CD3 antibody selectively reduced the number of circulating lymphocytes without significantly modifying the levels of granulocytes or monocytes (Fig. 7c). However, the antinociceptive effect evoked by 30 pg/kg of CCL4 was significantly reduced in mice previously treated with the anti-CD3 antibody when compared with mice treated with the corresponding antibody isotype IgG2b.
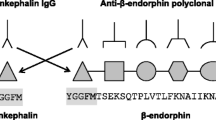
Since the marked reduction of CCL4-induced antinociceptive effect observed in CD3+-depleted mice supported the involvement of T lymphocytes in this response, we next explored which of the main types of CD3+ lymphocytes, CD4+ or CD8+, could participate in this met-enk-related effect. Therefore, we first performed double staining immunofluorescence assays with anti-CD4 or anti-CD8 antibodies together with an anti-met-enk antibody, to analyze the expression of met-enk in CD4+ or CD8+ lymphocytes. In experiments with anti-CD4 and anti-met-enk antibodies (Fig. 8), the expression of the corresponding antigens was analyzed in 2778 cells stained with DAPI from blood smears of white blood cells coming from 4 different mice. From these cells, 11.6 ± 3.03% expressed CD4, 7.68 ± 0.8% expressed met-enk and 1.5 ± 0.42%, showed double staining. Experiments performed with anti-CD8 and anti-met-enk antibodies included 3358 white blood cells stained with DAPI present in blood smears obtained from four different mice. In this case, 6.7 ± 1.3% were CD8+, 7.2 ± 1.5% expressed met-enk, and 0.36 ± 0.07% were positive for both antigens (Fig. 9). When comparing the results obtained in CD4+ and CD8+ cells, we observed that met-enk may be expressed in both types of lymphocytes and that its presence is about four times more frequent in CD4+ than in CD8+ lymphocytes, since 21.7% cells expressing met-enk were CD4+ lymphocytes (Fig. 8c) whereas only 4.9% cells were CD8+ (Fig. 9c).

Expression of met-enk in circulating CD4+ lymphocytes. a Representative microphotographs corresponding to a double-staining immunofluorescence experiment performed in a murine blood smear with an anti-met-enk antibody (green) and an anti-CD4 antibody (red). Cell nuclei were stained with DAPI (blue). Cells stained with the anti-met-enk antibody (green) are shown in the left panel and CD4-positive cells (red) in the middle photograph. Merged images including DAPI staining are shown in the right panel, and the yellow arrow indicates a cell with positive staining for met-enk and CD4. b Group of four cells from another double immunostaining assay performed with anti-met-enk (green, left-hand side) and anti-CD4 (red, middle image). At the right-hand side, the merged image shows that only one of the two CD4+ cells expresses met-enk (yellow arrow). c Diagram depicting the percentage of white blood cells expressing met-enk in this series of experiments as well as the percentage of met-enk-stained cells that also express CD4 in mice blood smears (N = 4)

Expression of met-enk in circulating CD8+ lymphocytes. a Representative microphotographs corresponding to a double-staining immunofluorescence experiment performed in a murine blood smear with an anti-met-enk antibody (green) and an anti-CD8 antibody (red). Cell nuclei were stained with DAPI (blue). Cells stained with the anti-met-enk antibody (green) are shown in the upper panel and CD8-positive cells (red) in the middle photograph. Merged images including DAPI staining are shown in the lower panel, showing that both antigens are not colocalized in this example. b Group of three cells from another double immunostaining assay performed with anti-met-enk (green, top image), anti-CD8 (red, middle image), and at the bottom, the merged image shows that one of the two met-enk-positive cells also expresses CD8 (yellow arrow). c Diagram depicting the percentage of white blood cells expressing met-enk in this series of experiments as well as the percentage of met-enk-stained cells that also express CD8 in mice blood smears (N = 4)
In order to assess if CD4+ or CD8+ T lymphocytes were functionally involved in the antinociceptive effect evoked by CCL4, we performed depletion experiments with anti-CD4 and anti-CD8 antibodies.
Flow cytometry analysis of peripheral blood CD3+ cells showed that CD4+ and CD8+ lymphocytes were significantly depleted 24 h after in vivo treatment with an anti-CD4 antibody (30 μg) or an anti-CD8 antibody (20 μg), respectively. Representative examples of flow cytometry experiments performed in blood of four mice treated with control isotypes, the anti-CD4 antibody, the anti-CD8 antibody, or both antibodies are shown in Fig. 10a. Mean percentages of CD4+ or CD8+ lymphocytes among CD3+ cells were dramatically reduced in mice injected with the anti-CD4 antibody or the anti-CD8 antibody, respectively and accordingly, a drastic depletion of both CD4+ and CD8+ lymphocytes was detected in mice treated with both antibodies (Fig. 10b). Behavioral assays showed that the administration of 30 pg/kg of CCL4 significantly increased withdrawal latencies in mice treated with control IgG, whereas significantly lower withdrawal latencies were obtained in mice treated with anti-CD4 antibody alone or in combination with anti-CD8 antibody after the administration of CCL4 (Fig. 10c). In contrast, administration of this dose of the chemokine produced an increase of withdrawal latencies in mice treated with the anti-CD8 antibody, which was indistinguishable from that obtained in non-depleted mice (Fig. 10c).

Inhibition of CCL4-evoked antinociception by CD4+ lymphocyte depletion. Flow cytometry and effects on CCL4-evoked analgesia after in vivo depletion of CD4+ and CD8+ lymphocytes in mice with anti-CD4 and anti-CD8 antibodies. a Representative flow cytometry profile of peripheral blood CD3+ cells from mice treated with isotype IgG, anti-CD4, anti-CD8, or both anti-CD4 and anti-CD8 antibodies. b Percentage of CD4+ and CD8+ cells among peripheral blood CD3+ lymphocytes detected in mice treated with isotype IgG2a-b (N = 6), anti-CD4 (N = 6), anti-CD8 (N = 6), or both anti-CD4 and anti-CD8 antibodies (N = 7). c Effects of treatment with isotype IgG (N = 6), anti-CD4 (N = 6), anti-CD8 (N = 6), and both anti-CD4 and anti-CD8 antibodies (N = 7) in basal paw withdrawal latencies before and after the administration of CCL4 (30 pg/kg; s.c.; 1 h before testing). Data are expressed as means ± SEM. **p < 0.01, compared with CCL4 + isotype IgG-treated group, Bonferroni’s correction
Discussion
The present results demonstrate that the systemic administration of very low doses of the chemokine CCL4 (MIP-1β) can produce antinociception in mice, being this response related to an increase in plasma levels of met-enk and to the presence of CD4+ lymphocytes.
Although the analgesic effects driven by other chemokines are generally consequence of the local release of antinociceptive molecules from immune cells [19, 20, 23], the effect of CCL4 was not local but systemic and unrelated to any chemotactic reaction at the site of injection. In fact, no modification in the number of neutrophils (stained with MPO), macrophages (F4/80), or lymphocytes (CD3) was found at the injected paw in response to the i.pl. administration of an antinociceptive dose of CCL4 and an increased presence of MPO+ cells was only detected 4 h after the injection of much higher doses of CCL4. Although the induction of mechanical allodynia after the perineural administration of exogenous CCL4 in mice has been described [28], mechanical thresholds measured by the von Frey test were unaffected by the i.pl. administration of an antinociceptive dose of CCL4 and only the injection of a 30,000 times higher dose, similar to that previously reported [28], induced local allodynia in the injected paw. Thus, the bilateral effect evoked by these low doses of CCL4 is not accompanied by local mechanical allodynia.
In order to assess whether, apart from thermal nociception, CCL4 could also inhibit mechanical nociceptive processing, we tested the effect of this chemokine in a paw pressure test. Contrasting with the von Frey test, designed to measure allodynia and based on the application of light, non-noxious, stimulation, the application of mechanical noxious stimulus allows the measurement of drug-induced analgesic responses in uninjured tissues. The administration of low doses of CCL4, that were effective in the UHP test, also induced bilateral mechanical antinociception in the paw pressure test, thus demonstrating the efficacy of CCL4 against noxious mechanical stimulation. Finally, the lack of effect of an antinociceptive dose of CCL4 on motor activity measured by an actimeter and the absence of visible motor alterations or sedation make very unlikely that this response could be biased by any unspecific depressor effect.
CCR5 and CCR1 are the two main receptors for CCL4. Although it has been reported that CCL4 can act on CCR1 either as agonist [39, 40] or antagonist [41], CCR5 is recognized as the main receptor responsible for CCL4 effects [42, 43]. In accordance with this general view, CCL4-induced antinociception was inhibited by the administration of the CCR5 antagonist, DAPTA, but unaffected by the administration of the CCR1 antagonist, J113863, at doses previously shown to block CCR1-mediated responses [44]. Although the stimulation of CCR5 has previously been linked to nociceptive responses [12], a recent report describes that CCL5, another CCR5 agonist, evokes dual effects, leading to hyperalgesia at low doses and triggering hypoalgesic compensatory mechanisms at slightly higher ones [23]. The affinity for CCR1 of CCL5, but not CCL4, could perhaps underlie the different responses evoked by both chemokines, although the possibility that different CCR5 agonists can lead to different consequences on nociception can be also envisaged [28]. Furthermore, experiments performed in the UHP test with naloxone or with selective μ-, δ-, and κ-opioid receptor antagonists demonstrate the participation of opioid mechanisms in CCL4-induced antinociception, a finding further confirmed by using the paw pressure test. Moreover, the lack of effect of i.t. administered DAPTA or naloxone at doses previously reported to inhibit CCR5- [45] or δ-opioid [46] receptor-mediated responses in mice discarded the action of CCL4 at the spinal cord, thus pointing towards the involvement of peripheral mechanisms.
Considering that the effect evoked by CCL4 is not restricted to the site of administration, that endogenous opioids are involved and that no local chemotactic responses occur after the administration of antinociceptive doses of CCL4, we focused our search on the possibility that the source of opioid peptides could be circulating blood cells instead of attracted ones. Aiming to elucidate which of the main endogenous opioids could be involved, we observed that CCL4-induced antinociception was inhibited by blocking CRF receptors, a molecule involved in met-enk or dynorphin release but remained unaffected after the blockade of receptors for IL-1, a mediator related to the release of β-end and dynorphin, but not enkephalins [38]. These results suggested that met-enk could be the endogenous opioid implicated in the effect of CCL4, a possibility that was also compatible with the participation of δ-opioid receptors, a receptor where met-enk binds with high affinity [47]. In keeping with this hypothesis, ELISA assays showed an increased concentration of met-enk, but not β-end, in the plasma of mice treated with an effective dose of CCL4. Moreover, the blockade of the antinociceptive effect evoked by CCL4 in mice treated with an anti-met-enk antibody demonstrates the functional participation of this opioid peptide. The release of met-enk in response to CCL4 administration might be understood in the context of the role played by this endogenous opiate in immune cell intercommunication. Several types of blood leukocytes such as lymphocytes [48], neutrophils [49] or monocytes [50] can be a source of met-enk that can act as a stimulus for the maturation of dendritic cells [51] or the expansion of CD4+ lymphocytes [52]. Accordingly, we observed that about 7% of circulating white blood cells showed met-enk expression detected by immunofluorescence staining.
Thus, considering that white blood cells could be responsible of the release of met-enk evoked by CCL4, immune depletion experiments were performed to depict the putative involvement of different cell lines. Initially, the treatment of mice with the immunosuppressive agent cyclophosphamide produced a great reduction in the population of lymphocytes, monocytes, or neutrophils [23] and strongly inhibited the antinociceptive effect produced by CCL4. In contrast, mice with marked depletion of granulocytes and monocytes due to the treatment with an anti-Ly6G antibody [23] maintained intact their response to CCL4, suggesting that these cell lines are not relevant and that the main role to generate this effect should be played by lymphocytes. The important reduction of the antinociception evoked by CCL4 in mice treated with an anti-CD3 antibody that selectively reduced the presence of lymphocytes but not granulocytes or monocytes, further indicates the participation of T lymphocytes in the antinociceptive effect induced by CCL4. In fact, CCL4 is a key physiological messenger released by B cells to activate T lymphocytes [53] and the induction of peripheral opioid analgesia in inflamed tissues through the release of met-enk from T lymphocytes has been previously demonstrated [54].
To determine the specific involvement of CD4+ or CD8+ subtypes of T lymphocytes, we initially performed double labelling immunofluorescence assays with an anti-met-enk antibody. Our results confirmed the expression of met-enk in CD4+ cells [55] and further demonstrated its presence in CD8+ lymphocytes. Although we have not found any previous quantification of the percentage of T lymphocytes expressing met-enk, our data indicate that, considering total white blood cells expressing met-enk, the staining with the anti-met-enk antibody was about four times more frequent in CD4+ than in CD8+ T lymphocytes (21.7 vs. 4.88%). Aiming to assess the implication of these two particular populations of T lymphocytes in the action of CCL4, flow cytometry studies were combined with behavioral assays in mice treated with selective antibodies against CD4 or CD8. In accordance with the effect obtained when using an anti-CD3 antibody, the simultaneous treatment with anti-CD4 and anti-CD8 antibodies provoked the almost complete disappearance of CCL4-evoked antinociception. Moreover, whereas the selective depletion of CD4+ T lymphocytes with an anti-CD4 antibody completely inhibited the effect induced by CCL4, it remained unaltered in CD8+-depleted mice, indicating that only CD4+, but not CD8+, cells are relevant. The finding that the antinociceptive effect induced by CCL4 through CCR5 activation can be linked to CD4+ T lymphocytes fits well with the previously described ability of this chemokine to activate chemotactic responses in this type of T lymphocytes [53, 56]. The fact that CCR5 expression is characteristic of Th1 CD4+ lymphocytes [57], whilst this receptor is not detected in the surface of circulating native CD8+ T lymphocytes [58], further supports the feasibility of the participation of CD4+ but not CD8+ in the effects induced by CCL4.
Altogether, a logical interpretation of our results is that the activation of CCR5 by low doses of CCL4 triggers the release of met-enk from CD4+ T lymphocytes and that this endogenous opioid evokes systemic antinociception by activating peripheral δ-opioid receptors (Fig. 11). In this sense, it seems interesting to remark that, as occurred in our experiments with CCL4, the aforementioned analgesia due to the release of met-enk from activated CD4+ T lymphocytes in inflamed mice is also mediated by the activation of δ-opioid receptors [55, 59]. However, this previously described analgesia is produced by activated CD4+ cells, appears with a delay of some days and is restricted to the injured paw. Conversely, CCL4-evoked antinociception can be already measured 1 h after CCL4 administration, it is not a localized but a systemic response and occurs in naive, non-immunized, mice.

Hypothetical model to explain CCL4-evoked analgesia. 1, The administration of exogenous CCL4 would provoke the activation of CCR5 but not CCR1, as deduced from the inhibitory effect produced by DAPTA but not J113863. 2, Based on depletion experiments with cyclophosphamide, anti-Ly6G, anti-CD3, anti-CD4, and anti-CD8 antibodies, the main cell line involved in this response should be CD4+ lymphocytes and not granulocytes or monocytes. 3, Immunofluorescence experiments showed that some of these CD4+ lymphocytes express met-enk and ELISA measurements demonstrated that CCL4 evokes an increase in the plasmatic levels of this opioid peptide. 4, This met-enk should be responsible of the stimulation of δ-opioid receptors, as determined by assaying the in vivo effect of an anti-met-enk antibody and selective opioid receptor antagonists, respectively. 5, The lack of effect of intrathecally administered antagonists indicated that neither CCR5 nor opioid receptors expressed at the spinal cord participate in the effect of CCL4. The administration of CCL4 produces a reduction in the number of nociceptive neurons activated in the spinal cord in response to a noxious stimulus, probably reflecting the diminished nociceptor input subsequent to δ-opioid receptor activation
To our knowledge, this is the first description of a systemic antinociceptive effect evoked by a chemokine in non-injured animals. Previous reports related to other chemokines do not refer to the elevation of nociceptive thresholds above basal physiologic levels but, instead, to the suppression of hypernociceptive responses measured in injured tissues [19, 20]. Further experiments seem necessary to draw a more complete picture of the antinociceptive action produced by CCL4. Thus, it should be interesting to assess the efficacy of CCL4 to counteract pain associated to pathological settings, such as inflammation or neuropathic injuries, where the enhanced presence of immune cells could favor the action of CCL4. Besides, the involvement of enkephalins might be an interesting property considering that tolerance in response to endogenous enkephalins could be less marked than that produced by exogenous opiates [60].
References
Turner MD, Nedjai B, Hurst T, Pennington DJ (2014) Cytokines and chemokines: at the crossroads of cell signalling and inflammatory disease. Biochim Biophys Acta 1843:2563–2582
Abbadie C (2005) Chemokines, chemokine receptors and pain. Trends Immunol 26:529–534
White FA, Bhangoo SK, Miller RJ (2005) Chemokines: integrators of pain and inflammation. Nat Rev Drug Discov 4:834–844
Dawes JM, McMahon SB (2013) Chemokines as peripheral pain mediators. Neurosci Lett 557(Pt A):1–8
Baamonde A, Hidalgo A, Menéndez L (2011) Involvement of glutamate NMDA and AMPA receptors, glial cells and IL-1β in the spinal hyperalgesia evoked by the chemokine CCL2 in mice. Neurosci Lett 502:178–181
Akimoto N, Honda K, Uta D, Beppu K, Ushijima Y, Matsuzaki Y, Nakashima S, Kido MA et al (2013) CCL-1 in the spinal cord contributes to neuropathic pain induced by nerve injury. Cell Death Dis 20(4):e679
Yin Q, Fan Q, Zhao Y, Cheng MY, Liu H, Li J, Lu FF, Jia JT et al (2015) Spinal NF-κB and chemokine ligand 5 expression during spinal glial cell activation in a neuropathic pain model. PLoS One 10:e0115120
Llorián-Salvador M, González-Rodríguez S, Lastra A, Fernández-García MT, Hidalgo A, Menéndez L, Baamonde A (2016) Involvement of CC chemokine receptor 1 and CCL3 in acute and chronic inflammatory pain in mice. Basic Clin Pharmacol Toxicol 119:32–40
Llorián-Salvador M, Pevida M, González-Rodríguez S, Lastra A, Fernández-García MT, Hidalgo A, Baamonde A, Menéndez L (2016) Analgesic effects evoked by a CCR2 antagonist or an anti-CCL2 antibody in inflamed mice. Fundam Clin Pharmacol 30:235–238
Zhang ZJ, Jiang BC, Gao YJ (2017) Chemokines in neuron-glial cell interaction and pathogenesis of neuropathic pain. Cell Mol Life Sci 74:3275–3291
Zhou YQ, Gao HY, Guan XH, Yuan X, Fang GG, Chen Y, Ye DW (2015) Chemokines and their receptors: Potential therapeutic targets for bone Cancer pain. Curr Pharm Des 21:5029–5033
Oh SB, Tran PB, Gillard SE, Hurley RW, Hammond DL, Miller RJ (2001) Chemokines and glycoprotein120 produce pain hypersensitivity by directly exciting primary nociceptive neurons. J Neurosci 21:5027–5035
Abbadie C, Lindia JA, Cumiskey AM, Peterson LB, Mudgett JS, Bayne EK, DeMartino JA, MacIntyre DE et al (2003) Impaired neuropathic pain responses in mice lacking the chemokine receptor CCR2. Proc Natl Acad Sci U S A 100:7947–7952
Silva RL, Lopes AH, Guimarães RM, Cunha TM (2017) CXCL1/CXCR2 signaling in pathological pain: Role in peripheral and central sensitization. Neurobiol Dis 105:109–116
Xu W, Zhu M, Yuan S, Yu W (2016) Spinal CXCL5 contributes to nerve injury-induced neuropathic pain via modulating GSK-3β phosphorylation and activity in rats. Neurosci Lett 634:52–59
Yu Y, Huang X, Di Y, Qu L, Fan N (2017) Effect of CXCL12/CXCR4 signaling on neuropathic pain after chronic compression of dorsal root ganglion. Sci Rep 7:5707
Milligan ED, Zapata V, Chacur M, Schoeniger D, Biedenkapp J, O'Connor KA, Verge GM, Chapman G et al (2004) Evidence that exogenous and endogenous fractalkine can induce spinal nociceptive facilitation in rats. Eur J Neurosci 20:2294–2302
Rittner HL, Brack A, Stein C (2008) The other side of the medal: how chemokines promote analgesia. Neurosci Lett 437:203–208
Rittner HL, Labuz D, Schaefer M, Mousa SA, Schulz S, Schäfer M, Stein C, Brack A (2006) Pain control by CXCR2 ligands through Ca2+-regulated release of opioid peptides from polymorphonuclear cells. FASEB J 20:2627–2629
Wang Y, Gehringer R, Mousa SA, Hackel D, Brack A, Rittner HL (2014) CXCL10 controls inflammatory pain via opioid peptide-containing macrophages in electroacupuncture. PLoS One 9:e94696
Kiguchi N, Kobayashi Y, Maeda T, Fukazawa Y, Tohya K, Kimura M, Kishioka S (2012) Epigenetic augmentation of the macrophage inflammatory protein 2/C-X-C chemokine receptor type 2 axis through histone H3 acetylation in injured peripheral nerves elicits neuropathic pain. J Pharmacol Exp Ther 340:577–587
Bu H, Shu B, Gao F, Liu C, Guan X, Ke C, Cao F, Hinton AO Jr et al (2014) Spinal IFN-γ-induced protein-10 (CXCL10) mediates metastatic breast cancer-induced bone pain by activation of microglia in rat models. Breast Cancer Res Treat 143:255–263
González-Rodríguez S, Álvarez MG, García-Domínguez M, Lastra A, Cernuda-Cernuda R, Folgueras AR, Fernández-García MT, Hidalgo A et al (2017) Hyperalgesic and hypoalgesic mechanisms evoked by the acute administration of CCL5 in mice. Brain Behav Immun 62:151–161
Oliveira SH, Lira S, Martinez AC, Wiekowski M, Sullivan L, Lukacs NW (2002) Increased responsiveness of murine eosinophils to MIP-1beta (CCL4) and TCA-3 (CCL1) is mediated by their specific receptors, CCR5 and CCR8. J Leukoc Biol 71:1019–1025
Appelberg R (1992) Macrophage inflammatory proteins MIP-1 and MIP-2 are involved in T cell-mediated neutrophil recruitment. J Leukoc Biol 52:303–306
Schall TJ, Bacon K, Camp RD, Kaspari JW, Goeddel DV (1993) Human macrophage inflammatory protein alpha (MIP-1 alpha) and MIP-1 beta chemokines attract distinct populations of lymphocytes. J Exp Med 177:1821–1826
Uguccioni M, D'Apuzzo M, Loetscher M, Dewald B, Baggiolini M (1995) Actions of the chemotactic cytokines MCP-1, MCP-2, MCP-3, RANTES, MIP-1 alpha and MIP-1 beta on human monocytes. Eur J Immunol 25:64–68
Saika F, Kiguchi N, Kobayashi Y, Fukazawa Y, Kishioka S (2012) CC-chemokine ligand 4/macrophage inflammatory protein-1β participates in the induction of neuropathic pain after peripheral nerve injury. Eur J Pain 16:1271–1280
Guo W, Imai S, Yang JL, Zou S, Watanabe M, Chu YX, Mohammad Z, Xu H et al (2017) In vivo immune interactions of multipotent stromal cells underlie their long-lasting pain-relieving effect. Sci Rep 7:10107
Dejima T, Shibata K, Yamada H, Hara H, Iwakura Y, Naito S, Yoshikai Y (2011) Protective role of naturally occurring interleukin-17A-producing γ δ T cells in the lung at the early stage of systemic candidiasis in mice. Infect Immun 79:4503–4510
Loubaki L, Tremblay T, Bazin R (2013) In vivo depletion of leukocytes and platelets following injection of T cell-specific antibodies into mice. J Immunol Methods 393:38–44
González-Rodríguez S, Llames S, Hidalgo A, Baamonde A, Menéndez L (2012) Potentiation of acute morphine-induced analgesia measured by a thermal test in bone cancer-bearing mice. Fundam Clin Pharmacol 26:363–372
Baamonde A, Curto-Reyes V, Juárez L, Meana A, Hidalgo A, Menéndez L (2007) Antihyperalgesic effects induced by the IL-1 receptor antagonist anakinra and increased IL-1beta levels in inflamed and osteosarcoma-bearing mice. Life Sci 81:673–682
Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL (1994) Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods 53:55–63
Baamonde A, Lastra A, Fresno MF, Llames S, Meana A, Hidalgo A, Menéndez L (2004) Implantation of tumoral XC cells induces chronic, endothelin-dependent, thermal hyperalgesia in mice. Cell Mol Neurobiol 24:269–281
Randall LO, Selitto JJ (1957) A method for measurement of analgesic activity on inflamed tissue. Arch Int Pharmacodyn Ther 111:409–419
Biddolph SC, Gatter KC (1999) Immunohistochemistry of lymphoid organs. In: Rowland-Jones SL, McMichael AJ (eds) Lymphocytes. A practical approach, 2nd edn. Oxford University Press, Oxford, p. 30
Cabot PJ, Carter L, Schäfer M, Stein C (2001) Methionine enkephalin and dynorphin A release from immune cells and control of inflammatory pain. Pain 93:207–212
Lu P, Nakamoto Y, Nemoto-Sasaki Y, Fujii C, Wang H, Hashii M, Ohmoto Y, Kaneko S et al (2003) Potential interaction between CCR1 and its ligand, CCL3, induced by endogenously produced interleukin-1 in human hepatomas. Am J Pathol 162:1249–1258
Repeke CE, Ferreira SB Jr, Claudino M, Silveira EM, de Assis GF, Avila-Campos MJ, Silva JS, Garlet GP (2010) Evidences of the cooperative role of the chemokines CCL3, CCL4 and CCL5 and its receptors CCR1+ and CCR5+ in RANKL+ cell migration throughout experimental periodontitis in mice. Bone 46:1122–1130
Chou CC, Fine JS, Pugliese-Sivo C, Gonsiorek W, Davies L, Deno G, Petro M, Schwarz M et al (2002) Pharmacological characterization of the chemokine receptor, hCCR1 in a stable transfectant and differentiated HL-60 cells: antagonism of hCCR1 activation by MIP-1beta. Br J Pharmacol 137:663–675
Alkhatib G (2009) The biology of CCR5 and CXCR4. Curr Opin HIV AIDS 4:96–103
Chang TT, Chen JW (2016) Emerging role of chemokine CC motif ligand 4 related mechanisms in diabetes mellitus and cardiovascular disease: friends or foes? Cardiovasc Diabetol 15:117
Pevida M, Lastra A, Meana Á, Hidalgo A, Baamonde A, Menéndez L (2014) The chemokine CCL5 induces CCR1-mediated hyperalgesia in mice inoculated with NCTC 2472 tumoral cells. Neuroscience 259:113–125
Hang LH, Li SN, Dan X, Shu WW, Luo H, Shao DH (2017) Involvement of spinal CCR5/PKCγ signaling pathway in the maintenance of cancer-induced bone pain. Neurochem Res 42:563–571
Heyman JS, Koslo RJ, Mosberg HI, Tallarida RJ, Porreca F (1986) Estimation of the affinity of naloxone at supraspinal and spinal opioid receptors in vivo: studies with receptor selective agonists. Life Sci 39:1795–1803
Mansour A, Hoversten MT, Taylor LP, Watson SJ, Akil H (1995) The cloned mu, delta and kappa receptors and their endogenous ligands: evidence for two opioid peptide recognition cores. Brain Res 700:89–98
Padrós MR, Vindrola O, Zunszain P, Fainboin L, Finkielman S, Nahmod VE (1989) Mitogenic activation of the human lymphocytes induce the release of proenkephalin derived peptides. Life Sci 45:1805–1811
Figuerola ML, Vindrola O, Barontini MB, Leston JA (1991) Changes in neutrophil Met-Enk containing peptides in episodic cluster headache. Headache 31:406–408
Hook S, Camberis M, Prout M, Konig M, Zimmer A, Van Heeke G, Le Gros G (1999) Preproenkephalin is a Th2 cytokine but is not required for Th2 differentiation in vitro. Immunol Cell Biol 77:385–390
Shan F, Xia Y, Wang N, Meng J, Lu C, Meng Y, Plotnikoff NP (2011) Functional modulation of the pathway between dendritic cells (DCs) and CD4+T cells by the neuropeptide: methionine enkephalin (MENK). Peptides 32:929–937
Hua H, Lu C, Li W, Meng J, Wang D, Plotnikoff NP, Wang E, Shan F (2012) Comparison of stimulating effect on subpopulations of lymphocytes in human peripheral blood by methionine enkephalin with IL-2 and IFN-γ. Hum Vaccin Immunother 8:1082–1089
Bystry RS, Aluvihare V, Welch KA, Kallikourdis M, Betz AG (2001) B cells and professional APCs recruit regulatory T cells via CCL4. Nat Immunol 2:1126–1132
Boué J, Blanpied C, Brousset P, Vergnolle N, Dietrich G (2011) Endogenous opioid-mediated analgesia is dependent on adaptive T cell response in mice. J Immunol 186:5078–5084
Boué J, Blanpied C, Djata-Cabral M, Pelletier L, Vergnolle N, Dietrich G (2012) Immune conditions associated with CD4+ T effector-induced opioid release and analgesia. Pain 153:485–493
Krzysiek R, Lefèvre EA, Zou W, Foussat A, Bernard J, Portier A, Galanaud P, Richard Y (1999) Antigen receptor engagement selectively induces macrophage inflammatory protein-1 alpha (MIP-1 alpha) and MIP-1 beta chemokine production in human B cell. J Immunol 162:4455–4463
Loetscher P, Uguccioni M, Bordoli L, Baggiolini M, Moser B, Chizzolini C, Dayer JM (1998) CCR5 is characteristic of Th1 lymphocytes. Nature 391:344–345
Askew D, Su CA, Barkauskas DS, Dorand RD, Myers J, Liou R, Nthale J, Huang AY (2016) Transient surface CCR5 expression by naive CD8+ T cells within inflamed lymph nodes is dependent on high endothelial venule interaction and augments Th cell-dependent memory response. J Immunol 196:3653–3664
Basso L, Boué J, Mahiddine K, Blanpied C, Robiou-du-Pont S, Vergnolle N, Deraison C, Dietrich G (2016) Endogenous analgesia mediated by CD4+ T lymphocytes is dependent on Enks in mice. J Neuroinflammation 13:132
Noble F, Turcaud S, Fournié-Zaluski MC, Roques BP (1992) Repeated systemic administration of the mixed inhibitor of enkephalin-degrading enzymes, RB101, does not induce either antinociceptive tolerance or cross-tolerance with morphine. Eur J Pharmacol 223:83–89
Acknowledgements
Grants were provided by the Ministerio de Economía, Industria y Competitividad, and FEDER (European Union) (SAF2017-86799-R). IUOPA is supported by Obra Social Fundación Cajastur-Liberbank (Asturias, Spain). M.G.D. is granted by a FPU predoctoral fellowship (FPU13-04585, MECD, Spain). A.R.F. is recipient of a Ramón y Cajal Fellowship.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
García-Domínguez, M., Lastra, A., Folgueras, A.R. et al. The Chemokine CCL4 (MIP-1β) Evokes Antinociceptive Effects in Mice: a Role for CD4+ Lymphocytes and Met-Enkephalin. Mol Neurobiol 56, 1578–1595 (2019). https://doi.org/10.1007/s12035-018-1176-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12035-018-1176-8