
Abstract
Pituitary adenylate cyclase-activating polypeptide (PACAP) is a neuropeptide that exerts general cytoprotective effects, including protection in different kidney disorders. The aim of our study was to investigate the ischemia/reperfusion-induced kidney injury of male and female rats to confirm the protective effects of PACAP in the kidney and to reveal possible gender differences.
Male and female Wistar rats underwent unilateral renal artery clamping followed by 24-h, 48-h, or 14-day reperfusion. PACAP was administered intravenously before arterial clamping in half of the rats. Tubular damage, cytokine expression pattern, oxidative stress marker, antioxidative status and signaling pathways were evaluated using histology, immunohistology, cytokine array, PCR, and Western blot. Tubular damage was significantly less severe in the PACAP-treated male and female rats compared to controls. Results of female animals were significantly better in both treated and untreated groups. Cytokine expression, oxidative stress marker and antioxidative status confirmed the histological results. We also revealed that PACAP counteracted the decreased PKA phosphorylation, influenced the expression of BMP2 and BMP4, and increased the expression of the protein Smad1.
We conclude that PACAP is protective in ischemia/reperfusion-induced kidney injury in both sexes, but females had markedly less pronounced injury after ischemia/reperfusion, possibly also involving further protective factors, the investigation of which could have future therapeutic value in treating ischemic kidney injuries.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Ischemia/reperfusion-induced kidney injury is an important cause or an additional aggravating factor of renal failure. It can lead to chronic kidney disease or to transition from preexisting chronic to end-stage renal disease. It can be evoked by different pathologies, including hypoperfusion, bleeding, and sepsis, and it is an unavoidable consequence following kidney transplantation. In the clinical practice, gender differences have been found in the progression of kidney disorders, with usually better outcome in females (Kher et al. 2005; Neugarten and Golestaneh 2013).
Pituitary adenylate cyclase–activating polypeptide (PACAP) is a neuropeptide with widespread nervous system and peripheral occurrence (Vaudry et al. 2009). PACAP has diverse functions, including regulation of endocrine and exocrine glands (Bardosi et al. 2016; Kanasaki et al. 2016; Sasaki et al. 2017), cardiovascular, urinary and gastrointestinal functions (Vu et al. 2016; Ivic et al. 2017; Heppner et al. 2018; Parsons and May 2018), and development of various organs (Juhasz et al. 2015; Watanabe et al. 2016; Farkas et al. 2017; Fulop et al. 2018). Trophic and protective effects are interconnected and associated with PACAP’s function as a growth factor in the nervous system and peripheral organs. Best known are PACAP’s neuroprotective effects (Reglodi et al. 2011, 2017), but it exerts general cytoprotection in peripheral tissues, such as oxidative stress- and doxorubicin-induced cardiac injury (Gasz et al. 2006; Mori et al. 2010), ischemic and inflammatory intestinal pathologies (Horvath et al. 2016; Heimesaat et al. 2017), endotoxin-induced airway inflammation (Elekes et al. 2011) as well as mouse dermatitis (Kemeny et al. 2010). In the kidney, PACAP’s protective effects are well established under several pathological conditions (rev.: Reglodi et al. 2012a, b; rev.: Khan and Batuman 2016), including gentamicin-, cisplatin-, cyclosporine-, and contrast agent-induced nephrotoxicity, diabetic, and myeloma nephropathy. There is evidence for the protective effects of PACAP in ischemia/reperfusion kidney injury in male rats (rev.: Khan and Batuman 2016) and in PACAP gene-deficient mice proving that endogenous PACAP is also part of the protective machinery (rev.: Reglodi et al. 2012a, b; Laszlo et al. 2015).
Females and males may react to ischemic injuries differently (Kang et al. 2014; Vafapour et al. 2015; Tanaka et al. 2017). PACAP’s protective effect has been shown to differ between sexes in a Parkinson’s disease model (Reglodi et al. 2006) and in tear secretion (Nakamachi et al. 2016), but it is not known whether this is the case in kidney ischemia/reperfusion injury. Therefore, our aim was to investigate the ischemia/reperfusion-evoked kidney injury in female and male rats, evaluating the effects of PACAP on morphology, inflammatory, and oxidative stress status and determining the signaling pathways that play a role in the effect of PACAP’s renoprotection.
Materials and Methods
Animals
Adult male and female Wistar rats (n = 112, 250–300 g) were housed under 12-h light/dark cycle with free access to food and water. Rats were divided into 12 groups: control (untreated) male rats (100 μl saline iv.) with 24-h, 48-h, or 14-day reperfusion; control (untreated) female rats (100 μl saline iv.) with 24-h, 48-h, or 14-day reperfusion; PACAP-treated male rats (100 μg PACAP/300 g dissolved in 100 μl saline iv.) with 24-h, 48-h, or 14-day reperfusion; and PACAP-treated female rats (100 μg PACAP/300 g dissolved in 100 μl saline iv.) with 24-h, 48-h, or 14-day reperfusion. Experimental procedures were carried out under approved protocols BA02/2000-24/2011. Vaginal cytology was performed in female rats to assure that procedures were in the same phases of the reproductive cycle.
Renal Ischemia/Reperfusion
Animals were anesthetized with intraperitoneal ketamine-xylazine. The temperature was kept constant during the operation with a warming blanket. PACAP was injected into the jugular vein before total median laparotomy. The left renal vessels were clamped for 60 min. Following 24 h, 48 h or 14 days, kidneys were removed and processed for further investigations.
Renal Histology and Morphometric Analysis
Rats were perfused with PBS (phosphate buffered saline) and 4% paraformaldehyde under anesthesia 24 h, 48 h, or 14 days after surgery. Removed kidneys were embedded in paraffin, and 5-μm thick sections were stained with periodic acid–Schiff base (PAS) and hematoxylin or with hematoxylin–eosin. Evaluation was performed on digital photographs using Adobe Photoshop version 10.0 and Scion Image 1.47 programs. Minimum ten fields on each slide—where only tubules were seen—were examined. The ratio of tubular lumen area was measured, as its enlargement represents tubular epithelial damage. Tubular lumen and dilated peritubular capillaries were marked manually, then converted to black, while the remaining tubular epithelial area to white. Tubular lumen area was expressed against the total tubular area. To get the total tubular area, the area of the dilated peritubular capillaries was subtracted from the total area (black and white areas together) (Fig. 1).

Morphometric analysis of PAS-H-stained kidney sections—measurement of the ratio of tubular lumen area. A: PAS-H-stained kidney section. B and C: the black area represents the tubular lumen and the peritubular capillaries—tubular lumen and dilated peritubular capillaries were marked manually, then converted to black, while the remaining tubular epithelial area to white in the photo C. D and E: the black area represents the tubular lumen—tubular lumen was marked manually, then converted to black, while the remaining tubular epithelial area and the peritubular capillaries to white in the photo E. Ratio of the tubular lumen area = E/(total area – C + E). Total area: area of the visual field (black + white area). Tubular lumen area was expressed against total tubular area. To get the total tubular area, the area of the dilated peritubular capillaries was subtracted from the total area
Cytokine Expression—Cytokine Array Analysis
Kidneys removed 24 h after ischemia were processed for cytokine array (Panel A, R&D Systems, Minneapolis, USA). The array, in duplicate sets, was performed as described by the manufacturer. Briefly, kidneys were homogenized in PBS containing protease inhibitor. Triton X-100 was added to the final concentration of 1%. After blocking the array membranes for 1 h and adding the reconstituted detection antibody cocktail for another 1 h at room temperature, the membranes were incubated in 1 ml of tissue homogenates at 2–8 °C overnight on a rocking platform. After washing with buffer three times and addition of horseradish peroxidase–conjugated streptavidin to each membrane, we exposed them to a chemiluminescent detection reagent (Amersham Biosciences, Little Chalfont, UK).
Biochemical Assay of Oxidative Stress Marker and Antioxidative Status
Kidneys removed 24 h after ischemia were processed for biochemical assay of oxidative stress marker and antioxidative status to measure superoxide dismutase (SOD), malondialdehyde (MDA), and reduced glutathione (GSH) as described previously. Kidney homogenates were centrifuged for 20 min at 16,000 rpm, and the supernatant was used to measure the concentration of superoxide dismutase (SOD) at absorbance 480 nm. MDA was evaluated with addition of TBA (saturated thiobarbituric acid in 10% perchloric acid)-TCA (20% trichloroacetic acid) reagent. After incubation at 100 °C for 20 min, samples were placed in ice-cold water and were centrifuged for 15 min at 4000 rpm. MDA concentration was determined spectrophotometrically by measuring the absorbance at 532 nm. For GSH measurements, kidney homogenates were centrifuged for 15 min at 4000 rpm after adding 10% TCA. TRIS buffer (tris(hydroxymethyl)methylamine) was added to the supernatant, and samples were measured at 412 nm after adding DTNB (5,5′-dithio-bis (2-nitrobenzoic acid)).
RT-PCR Analysis
Kidneys removed 24 h or 48 h after surgery, in three independent experiments, were processed for RT-PCR analysis. Tissues were mechanically ground in TriReagent (Applied Biosystems, Foster City, CA, USA) and incubated at room temperature for 30 min. After the addition of 20% RNase-free chloroform, samples were centrifuged at 4 °C at 10,000×g for 15 min. Samples were incubated in 500 μl of RNase-free isopropanol at − 20 °C for 1 h then total RNA was harvested in RNase-free water and stored at − 20 °C. The reverse transcriptase reaction was performed according to the suggestions of the manufacturer. For the sequences of primer pairs and further details of PCR, see Table 1. Amplifications were performed in a thermal cycler (Labnet MultiGene™ 96-well Gradient Thermal Cycler; Labnet International, Edison, NJ, USA) in a final volume of 11 μl (containing 1 μl forward and reverse primers [0.4 μM], 0.5 μl dNTP [200 μM], and 5 units of Promega GoTaq® DNA polymerase in 1× reaction buffer) as follows: 95 °C, 2 min, followed by 35 cycles (denaturation, 94 °C, 1 min; annealing at optimized temperatures as given in Table 1 for 1 min; extension, 72 °C, 90 s) and then 72 °C, 10 min. PCR products were analyzed by electrophoresis in 1.2% agarose gel containing ethidium bromide. Actin was used as internal control. Signals were developed with gel documentary system (Fluorchem E, ProteinSimple, CA, USA). Optical signal density was measured using ImageJ 1.40 g freeware and results were normalized to controls.
Western Blot Analysis
Kidneys removed 24 h or 48 h after surgery, in three independent experiments, were washed in physiological saline then collected in 100 μl of homogenization RIPA (Radio Immuno Precipitation Assay)-buffer (150 mM sodium chloride; 1.0% NP40, 0.5% sodium deoxycholate; 50 mM Tris, pH 8.0) containing protease inhibitors. Samples were stored at − 70 °C. Kidneys were sonicated by pulsing burst for 30 s at 40 A (Cole-Parmer, IL, USA). Laemmli electrophoresis sample buffer (Bio-Rad Laboratories, CA, USA) was added to total tissue lysates and boiled for 10 min. Twenty micrograms of protein was separated by 7.5% SDS-PAGE gel for detection of PKA (protein kinase A), P-PKA (phosphorylated protein kinase A), BMPR1 (bone morphogenetic protein type I receptor), BMP2 (bone morphogenetic protein 2), BMP4 (bone morphogenetic protein 4), Smad1, and Col. IV (collagen IV). Proteins were transferred electrophoretically to nitrocellulose membranes. After blocking in 5% non-fat dry milk in PBST (phosphate buffered saline with 0.1% Tween 20; 20 mM Na2HPO4, 115 mM NaCl, pH 7.4), membranes were washed and exposed to primary antibodies (Table 2) overnight at 4 °C. After washing three times for 10 min with PBST, membranes were incubated with the secondary antibody, anti-rabbit IgG (Bio-Rad Laboratories, CA, USA, 1:1500), or with anti-mouse IgG (Bio-Rad Laboratories, CA, USA, 1:1500) in PBST containing 1% non-fat dry milk for 2 h at room temperature. Signals were detected by enhanced chemiluminescence (Advansta Inc., Menlo Park, CA, USA) and developed with gel documentary system (Fluorchem E, ProteinSimple, CA, USA). Signals of male samples were not visible with this enhancing procedure; therefore, SuperSignal West Femto Maximum Sensitivity Substrate (Pierce, Rockford, IL, USA) was used according to the instructions of the manufacturer. Actin was used as internal control. The optical signal density was measured by ImageJ 1.40 g freeware and normalized to controls.
Immunohistochemistry
Perfused kidneys, removed 24 h or 48 h after surgery under anesthesia, were washed in PBS three times and fixed in Saint-Marie’s fixative (99% ethanol, 1% anhydrous acetic acid) for 24 h. After embedding, 5-μm sections were made. Nonspecific binding sites were blocked in PBST supplemented with 1% bovine serum albumin (BSA, Amresco LLC, Solon, OH, USA) at 37 °C for 30 min. For Col. IV, Smad1, BMPR1, and BMP4 immunohistochemistry, sections were incubated in anti-Col. IV (Abcam, Cambridge, UK, 1:500), anti-Smad1 (Cell Signaling, Danvers, MA, USA, 1:600), anti-BMPR1 (Abcam, UK, 1:400), and anti-BMP4 (Cell Signaling, Danvers, MA, USA, 1:500) at 4 °C overnight. Primary antibodies were visualized with anti-rabbit Alexa555 and/or anti-mouse Alexa488 secondary antibodies (Life Technologies Corporation, Carlsbad, CA, USA) at a dilution of 1:1000. Slides were mounted in Vectashield Hard Set mounting medium (Vector Laboratories Ltd., Peterborough, UK) containing DAPI to visualize cell nuclei. Photomicrographs of Col. IV and BMPR1 were taken using an Olympus DP72 camera on a Nikon Eclipse E800 microscope (Nikon Corporation, Tokyo, Japan). Images were acquired using cellSense Entry 1.5 software (Olympus, Shinjuku, Tokyo, Japan) using constant camera settings to allow comparison of fluorescent signal intensities. For investigation of subcellular localization of Smad1 and BMP4, fluorescent images were taken with an Olympus FV1000S confocal microscope (Olympus Co. Tokyo, Japan) using 60× oil immersion objective (NA, 1.3). Laser lines of 543 nm and 488 nm were used. The average pixel time was 4 μs. Z image series of 1-μm optical thickness were recorded in sequential scan mode. Images of Alexa555, Alexa488, and DAPI were overlaid using Adobe Photoshop version 10.0 software.
Statistical Analysis
Morphometric analysis and biochemical assay were analyzed with ANOVA, Western blot, and RT-PCR with two-sample t test. p < 0.05 was considered statistically significant.
Results
Renal Histology and Morphometric Analysis
Dilation of tubular lumen caused by tubular epithelial damage was markedly greater in the control than in the PACAP-treated groups. In control males, the tubular damage already reached maximum at 24 h, and there was no remarkable deterioration at 48-h or 14-day ischemia/reperfusion. Tubular damage became more severe with time in control females, gradually reaching the level observed in males at 24-h ischemia/reperfusion. Significantly less severe histological alterations were found in the PACAP-treated groups in both genders. Results of PACAP-treated females were significantly better compared to males at 48 h and 14 days. The damage was significantly more severe after longer reperfusion times in control females, while the duration of the reperfusion had no influence on the severity of the damage in PACAP-treated females (Figs. 2 and 3).

Representative photomicrographs of PAS-H-stained kidney sections of control (A–C) and PACAP-treated (D–F) male and female rats following 24-h (A, D)/48-h (B, E)/14-day (C, F) ischemia/reperfusion. Scale bar, 20 μm

Result of the morphometric analysis. Ratio of the tubular lumen area in the kidneys of control male/female and PACAP-treated male/female rats following 24-h, 48-h, and 14-day ischemia/reperfusion. Data are given as mean + SEM. **p < 0.01, ***p < 0.001 vs time-matched control male, xxp < 0.01, xxxp < 0.001 vs 24-h control female, &&&p < 0.001 vs time-matched control female, ##p < 0.01, ###p < 0.001 vs 24-h PACAP-treated male, ++p < 0.01 vs time-matched PACAP-treated male (n = 60)
Cytokine Expression
Expression of fractalkine, l-selectin, RANTES, and sICAM-1 was nearly the same in both genders following ischemia/reperfusion and was decreased by PACAP, more in females in the case of fractalkine, l-selectin, and RANTES. Major sex differences could be observed in TIMP-1 and MIP-3α, which were higher in males. Expression of thymus chemokine was higher in females after ischemia/reperfusion and was reduced by PACAP in both genders. Presence of the neurotrophic factor CNTF could be observed only in females after ischemia/reperfusion and could no longer be detected following PACAP treatment (Fig. 4).

Representative cytokine array showing cytokine expression in the kidneys of control male/female and PACAP-treated male/female rats following 24-h ischemia/reperfusion. Table shows the cytokines of the array in the representative double spots. The examined cytokines/chemokines are CINC-1 (cytokine-induced neutrophil chemoattractant 1), CINC-2αβ (cytokine-induced neutrophil chemoattractant 2αβ), CINC-3 (cytokine-induced neutrophil chemoattractant 3), CNTF (ciliary neurotrophic factor), fractalkine, GM-CSF (granulocyte-macrophage colony-stimulating factor), sICAM-1 (soluble intercellular adhesion molecule-1), IFN-μ (interferon μ), IL-1α (interleukin 1α), IL-1β (interleukin 1β), IL-1ra (interleukin-1 receptor antagonist), IL-2 (interleukin 2), IL-3 (interleukin 3), IL-4 (interleukin 4), IL-6 (interleukin 6), IL-10 (interleukin 10), IL-13 (interleukin 13), IL-17 (interleukin 17), IP-10 (interferon gamma–induced protein 10), LIX (lipopolysaccharide-induced CXC chemokine), l-Selectin, MIG (monokine induced by gamma interferon), MIP-1α (macrophage inflammatory protein 1α), MIP-3α (macrophage inflammatory protein 3α), RANTES (regulated on activation, normal T cell expressed and secreted), thymus chemokine, TIMP-1 (tissue inhibitor of metalloproteinases 1), TNF-α (tumor necrosis factor-α), VEGF (vascular endothelial growth factor) (n = 12)
Biochemical Assay of Oxidative Stress Marker and Antioxidative Status
Higher SOD activity was measured after PACAP treatment in both sexes, but a significant difference was only found in females. Levels of MDA were moderately lower in both PACAP-treated male and female rats, though differences were not significant. No significant alterations were found in GSH levels between any groups (Fig. 5)

Renal expression of SOD (superoxide dismutase), MDA (malondialdehyde), and GSH (glutathione) in control male/female and PACAP-treated male/female rats following 24-h ischemia/reperfusion. *p < 0.05 vs female control (n = 12)
BMP Signaling Assessed by RT-PCR, Western Blot, and Immunohistochemistry
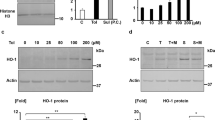
Results of PCR and Western blot are shown in Figs. 6 and 7. Binding of PACAP to its receptors activates PKA, the mRNA and protein expression of which increased after 24-h ischemia/reperfusion in the PACAP-treated females and males. Phosphorylation (activation) of PKA also showed a significant elevation in both genders after PACAP treatment following 24-h ischemia/reperfusion. However, after 48-h ischemia/reperfusion, the expression profiles of PKA showed a significant decrease in the case of PACAP treatment. BMP2 mRNA was significantly elevated in males, while no alteration was detected in females following PACAP treatment at 24 h; BMP2 protein expression did not change in either sex. On the contrary, BMP4 mRNA and protein were significantly elevated in both genders. The immunopositivity of this transcription factor was stronger in the PACAP-treated groups accumulating mainly in the cortical tubules (Fig. 8). PACAP altered the mRNA expression of BMPs in the 48-h ischemia/reperfusion group: BMP2 and BMP4 increased in males, while no significant changes were detected in females. Protein expression of BMP2 and BMP4 was elevated only in females, but no alterations were observed in the PACAP-treated males after 48-h ischemia/reperfusion. BMPR1 receptor was detected in both genders, and PACAP increased its mRNA and protein expression in male and female rats after 24 h. In the cortical tubules, BMPR1 showed stronger signals following PACAP administration (Fig. 8). Interestingly, BMPR1 mRNA expression was reduced after PACAP treatment at 48 h in both sexes, although its protein level remained constant (Figs 6 and 7).

mRNA expression of PKA (protein kinase A), BMP2 (bone morphogenetic protein 2), BMP4 (bone morphogenetic protein 4), BMPR1 (bone morphogenetic protein type I receptor), Smad1, and Col41a in the kidneys of control male/female and PACAP-treated male/female rats following 24-h and 48-h ischemia/reperfusion assessed by RT-PCR. Optical density of signals was measured, and results were normalized to the optical density of controls. Numbers below the signals represent integrated densities of signals determined by ImageJ software. *p < 0.05 vs control (n = 24)

Western blot analysis of PKA (protein kinase A), P-PKA (phosphorylated protein kinase A), BMP2 (bone morphogenetic protein 2), BMP4 (bone morphogenetic protein 4), BMPR1 (bone morphogenetic protein type I receptor), Smad1, and Col. IV (collagen IV) in the kidneys of control male/female and PACAP-treated male/female rats following 24-h and 48-h ischemia/reperfusion. Optical density of signals was measured, and results were normalized to the optical density of controls. Numbers below the signals represent integrated densities of signals determined by ImageJ software. *p < 0.05 vs control (n = 24)

Immunohistochemistry of BMP4 (bone morphogenetic protein 4), BMPR1 (bone morphogenetic protein type I receptor), Smad1, and collagen IV in the kidneys of control male/female and PACAP-treated male/female rats following 24-h and 48-h ischemia/reperfusion. Scale bar, 50 μm (n = 16)
One of the downstream targets of BMPR1 is Smad1, the mRNA expression of which was significantly elevated in the PACAP-treated groups following 24-h ischemia/reperfusion (Fig. 6). The protein level of Smad1 in PACAP-treated males and females was increased 24 h after ischemia/reperfusion (Fig. 7). Moreover, immunopositivity of tubules dramatically increased in the PACAP-treated groups (Fig. 8). In contrast, the mRNA expression of Smad1 was reduced after 48-h ischemia/reperfusion in PACAP-treated animals, but surprisingly, the protein expression still showed an elevated level (Fig. 7). One of the target genes of this signaling pathway is collagen type IV, the mRNA expression of which was also augmented by PACAP treatment after 24-h ischemia/reperfusion in male and female rats equally (Fig. 6). Moreover, a strong elevation in their protein expression was detected in both genders (Fig. 7). The tubules and the renal corpuscles were strongly surrounded by a collagen type IV–immunopositive line in PACAP-treated groups (Fig. 8). On the contrary, PACAP treatment resulted in reduced collagen type IV mRNA after 48-h ischemia/reperfusion, but its protein expression was still higher than in the control group in both females and males (Figs. 6 and 7).
Discussion
In the present study, the role of PACAP and possible gender differences were investigated in ischemia/reperfusion-induced kidney injury. Morphometric analysis revealed that the histological alterations were more severe in control males than in females, with the exception of the 14-day reperfusion group. This corresponds to descriptions showing that kidney diseases are more severe, the damage progrediates more rapidly and leads earlier to end-stage renal failure in males (Kher et al. 2005; Neugarten and Golestaneh 2013). Significantly less severe histological alterations were found in the PACAP-treated rats in both sexes. PACAP has already been proven protective in different kidney injuries, such as diabetic and myeloma nephropathy-, gentamicin-, cyclosporine-, cisplatin-, and contrast-induced nephrotoxicity (rev.: Reglodi et al. 2012a, b; Khan and Batuman 2016). PACAP decreased the damage in male rats after 60-min renal ischemia (rev.: Khan and Batuman 2016), similar to protective effects in other organs against ischemia: retina (Werling et al. 2017), brain (rev.: Vaudry et al. 2009), and intestine (rev.: Horvath et al. 2016). In this study, we found less severe tubular damage in PACAP-treated female rats. Similar gender-dependent protective effect of PACAP was described in Parkinson’s disease (Reglodi et al. 2006) and in corneal keratinization (Nakamachi et al. 2016).
We tested the effect of PACAP on the cytokine profile, as one of the mechanisms of PACAP’s renoprotection is its immunomodulatory action (rev.: Vaudry et al. 2009). Our data revealed that PACAP decreased the expression of several cytokines (fractalkine, l-selectin, RANTES, sICAM-1, thymus chemokine, TIMP-1, and MIP-3α) in rats exposed to renal ischemia/reperfusion in both sexes, showing overlap with our previous data in the same model and with findings in diabetic nephropathy (Banki et al. 2014). In retinal hypoperfusion, PACAP also attenuated increased cytokine expression (CNTF, fractalkine, sICAM-1, TIMP-1, MIP-3α, l-selectin, and RANTES) (Szabo et al. 2012). Differences between males and females were found in the expression of CNTF, which was detectable only in females. CNTF is thought to be part of the protective system against renal injury (Yang et al. 2001), but PACAP could not enhance the CNTF expression suggesting that the renoprotective effect of PACAP does not include influence on CNTF in this model. Gender difference was also observed in TIMP-1 and MIP-3α, with higher levels in males in renal ischemia/reperfusion, similar to mice exposed to hepatic ischemia/reperfusion (Crockett et al. 2006). Decrease in fractalkine, l-selectin, and RANTES was more pronounced in females. Oxidative stress also plays an important role in the pathomechanism of ischemia/reperfusion-induced kidney injury (Menger and Vollmar 2007). Therefore, markers of oxidative stress and of antioxidative status were also investigated in our study. We did not find significant difference between the groups in MDA level, but higher activity of the antioxidant SOD was measured in the PACAP-treated groups, similar to earlier findings in renal and intestinal ischemia (Horvath et al. 2011, 2016). Difference between SOD activity of PACAP-treated and control groups was significant only in females, confirming the gender-dependent protective effect of PACAP. PAC1 receptor activation, via the cAMP/PKA pathways, has crosstalks with, among others, WNT (Kim et al. 2013), β-catenin (Yu et al. 2014), and BMP signaling (Chen et al. 2012; Juhasz et al. 2015), through which it can regulate various cellular functions, such as proliferation, differentiation, and matrix production. Here, we found that PACAP augmented expression and phosphorylation of PKA and prevented the harmful effect of ischemia, in accordance with other studies (rev.: Vaudry et al. 2009). Several transcription factors are influenced by PKA, such as CREB (rev.: Vaudry et al. 2009), Sox9, or Runx2, the activation of which can compensate the harmful effects of ischemia/reperfusion and can induce other pathways, like BMP-related signaling (Nguyen et al. 2017). Earlier, we demonstrated that PACAP increased the expression of BMPs in osteoblasts and chondrifying cells, where BMP signaling can induce extracellular matrix production (Juhasz et al. 2015). BMP4 and BMP7 determine nephron differentiation in the kidney (Katagiri and Watabe 2016). BMP, binding to its receptor (BMPRI), can induce nuclear translocation of Smad1, which may induce production or modification of basal membrane component collagen type IV. Earlier studies demonstrated that PACAP reduced the severity of diabetic nephropathy (Banki et al. 2014) and altered the expression of collagen type IV. In oxalate-induced kidney injury, BMP7 overexpression compensated the harmful effects via the TGFβ signaling (Convento et al. 2017), and nephropathy can have direct connections with an increased BMP6 expression (Falke et al. 2016). Here, we demonstrated that PACAP had gender- and time-dependent effects on BMP expression. BMP2 was unaltered in males, but PACAP increased its expression after 48 h in females. BMP4 protein expression was elevated in both sexes after 24 h of PACAP administration, but an increase was detected after 48 h only in females. There are some BMPs the expression and function of which can be gender dependent (Lochab and Extavour 2017). The present results suggest that BMP2 does not have an important function in PACAP-induced mechanism in renal ischemia/reperfusion. BMP4 elevated in both genders, but the increase or the time dependency was altered in genders. The thickening of the basement membrane in the kidney suggests the formation of severe kidney disease like fibrosis (Banki et al. 2014). In diabetic nephropathy, increased expression of Smad1 and collagen type IV has been detected (Korish et al. 2015), and Smad1 can modify collagen type IV inducing nephropathy (Abe et al. 2004). In our experiments, we detected gender-dependent elevation of BMP4-BMPR1-Smad1, resulting in increased collagen type IV. This result interestingly suggests a signaling cascade very precisely regulated by PACAP, which can be disturbed during kidney injury. Addition of PACAP may have a dual effect as it can prevent the inflammatory effects directly and shift the balance of BMP pathway resulting a thickening of basement membrane.
Conclusively, the present study shows that PACAP is able to decrease the damage in renal ischemia/reperfusion in both males and females, with markedly less pronounced injury in females. Whether this difference between sexes is due to hormonal alterations, as it has already been described in earlier studies, or further factors play a role in this gender-dependent effect of PACAP requires further investigation preceding the potential therapeutic use of PACAP in kidney injuries (Kang et al. 2014; Sandhi et al. 2014; Talebi et al. 2016).
Furthermore, it provides data on the mechanism of PACAP’s renoprotective effect, although its dual actions should also be taken into account, especially regarding the production of collagen type IV.
References
Abe H, Matsubara T, Iehara N, Nagai K, Takahashi T, Arai H, Kita T, Doi T (2004) Type IV collagen is transcriptionally regulated by Smad1 under advanced glycation end product (AGE) stimulation. J Biol Chem 279:14201–14206. https://doi.org/10.1074/jbc.M310427200
Banki E, Kovacs K, Nagy D, Juhasz T, Degrell P, Csanaky K, Kiss P, Jancso G, Toth G, Tamas A, Reglodi D (2014) Molecular mechanisms underlying the nephroprotective effects of PACAP in diabetes. J Mol Neurosci 54:300–309. https://doi.org/10.1007/s12031-014-0249-z
Bardosi S, Bardosi A, Zs N, Reglodi D (2016) Expression of PACAP and PAC1 receptor in normal human thyroid gland and in thyroid papillary carcinoma. J Mol Neurosci 60:171–178. https://doi.org/10.1007/s12031-016-0823-7
Chen G, Deng C, Li YP (2012) TGF-beta and BMP signaling in osteoblast differentiation and bone formation. Int J Biol Sci 8:272–288. https://doi.org/10.7150/ijbs.2929
Convento MB, Pessoa EA, Cruz E, da Glória MA, Schor N, Borges FT (2017) Calcium oxalate crystals and oxalate induce an epithelial-to-mesenchymal transition in the proximal tubular epithelial cells: contribution to oxalate kidney injury. Sci Rep 7:45740. https://doi.org/10.1038/srep45740
Crockett ET, Spielman W, Dowlatshahi S, He J (2006) Sex differences in inflammatory cytokine production in hepatic ischemia-reperfusion. J Inflamm (Lond) 3:16. https://doi.org/10.1186/1476-9255-3-16
Elekes K, Sandor K, Moricz A, Kereskai L, Kemeny A, Szoke E, Perkecz A, Reglodi D, Hashimoto H, Pinter E, Szolcsanyi J, Zs H (2011) Pituitary adenylate cyclase activating polypeptide plays an anti-inflammatory role in endotoxin-induced airway inflammation: in vivo study with gene-deleted mice. Peptides 32:1439–1446. https://doi.org/10.1016/j.peptides.2011.05.008
Falke LL, Kinashi H, Dendooven A, Broekhuizen R, Stoop R, Joles JA, Nguyen TQ, Goldschmeding R (2016) Age-dependent shifts in renal response to injury relate to altered BMP6/CTGF expression and signaling. Am J Physiol Renal Physiol 311:F926–F934. https://doi.org/10.1152/ajprenal.00324.2016
Farkas J, Sandor B, Tamas A, Kiss P, Hashimoto H, Nagy AD, Fulop BD, Juhasz T, Manavalan S, Reglodi D (2017) Early neurobehavioral development of mice lacking endogenous PACAP. J Mol Neurosci 61:468–478. https://doi.org/10.1007/s12031-017-0887-z
Fulop BD, Sandor B, Szentleleky E, Karanyicz E, Reglodi D, Gaszner B, Zakany R, Hashimoto H, Juhasz T, Tamas A (2018) Altered notch signaling in developing molar teeth of pituitary adenylate cyclase-activating polypeptide (PACAP)-deficient mice. J Mol Neurosci. https://doi.org/10.1007/s12031-018-1146-7
Gasz B, Racz B, Roth E, Borsiczky B, Ferencz A, Tamas A, Cserepes B, Lubics A, Gallyas F Jr, Toth G, Lengvari I, Reglodi D (2006) Pituitary adenylate cyclase activating polypeptide protects cardiomyocytes against oxidative stress-induced apoptosis. Peptides 27:87–94. https://doi.org/10.1016/j.peptides.2005.06.022
Heimesaat MM, Reifenberger G, Vicena V, Illes A, Horvath G, Tamas A, Fulop BD, Bereswill S, Reglodi D (2017) Intestinal microbiota changes in mice lacking pituitary adenylate cyclase activating polypeptide (PACAP) - Bifidobacteria make the difference. Eur J Microbiol Immunol 7:187–199. https://doi.org/10.1556/1886.2017.00021
Heppner TJ, Hennig GW, Nelson MT, May V, Vizzard MA (2018) PACAP38-mediated bladder afferent nerve activity hyperexcitability and Ca2+ activity in urothelial cells from mice. J Mol Neurosci. https://doi.org/10.1007/s12031-018-1119-x
Horvath G, Brubel R, Kovacs K, Reglodi D, Opper B, Ferencz A, Szakaly P, Laszlo E, Hau L, Kiss P, Tamas A, Racz B (2011) Effects of PACAP on oxidative stress-induced cell death in rat kidney and human hepatocyte cells. J Mol Neurosci 43:67–75. https://doi.org/10.1007/s12031-010-9428-8
Horvath G, Illes A, Heimesaat MM, Bardosi A, Bardosi S, Tamas A, Fulop BD, Opper B, Nemeth J, Ferencz A, Reglodi D (2016) Protective intestinal effects of pituitary adenylate cyclase activating polypeptide. In: Reglodi D, Tamas A (eds) Pituitary adenylate cyclase activating polypeptide - PACAP. Springer, Switzerland, pp 271–289
Ivic I, Fulop BD, Juhasz T, Reglodi D, Toth G, Hashimoto H, Tamas A, Koller A (2017) Backup mechanism maintains PACAP/VIP-induced arterial relaxations in PACAP-deficient mice. J Vasc Res 54:180–192. https://doi.org/10.1159/000457798
Juhasz T, Helgadottir SL, Tamas A, Reglodi D, Zakany R (2015) PACAP and VIP signaling in chondrogenesis and osteogenesis. Peptides 66:51–57. https://doi.org/10.1016/j.peptides.2015.02.001
Kanasaki H, Oride A, Hara T, Mijiddorj T, Sukhbaatar U, Kyo S (2016) Interactions between two different G protein-coupled receptors in reproductive hormone-producing cells: the role of PACAP and its receptor PAC1R. Int J Mol Sci 17:1635. https://doi.org/10.3390/ijms17101635
Kang KP, Lee JE, Lee AS, Jung YJ, Kim D, Lee S, Hwang HP, Kim W, Park SK (2014) Effect of gender differences on the regulation of renal ischemia-reperfusion-induced inflammation in mice. Mol Med Rep 9:2061–2068. https://doi.org/10.3892/mmr.2014.2089
Katagiri T, Watabe T (2016) Bone morphogenetic proteins. Cold Spring Harb Perspect Biol 8:a021899. https://doi.org/10.1101/cshperspect.a021899
Kemeny A, Reglodi D, Cseharovszky R, Hashimoto H, Baba A, Szolcsanyi J, Pinter E, Helyes Z (2010) Pituitary adenylate cyclase-activating polypeptide deficiency enhances oxazolone-induced allergic contact dermatitis in mice. J Mol Neurosci 42:443–449. https://doi.org/10.1007/s12031-010-9368-3
Khan MA, Batuman V (2016) Renoprotective effects of pituitary adenylate cyclase-activating polypeptide 38 (PACAP38). In: Reglodi D, Tamas A (eds) Pituitary adenylate cyclase activating polypeptide - PACAP. Springer, Switzerland, pp 289–313
Kher A, Meldrum KK, Wang M, Tsai BM, Pitcher JM, Meldrum DR (2005) Cellular and molecular mechanisms of sex differences in renal ischemia-reperfusion injury. Cardiovasc Res 67:594–603. https://doi.org/10.1016/j.cardiores.2005.05.005
Kim JH, Liu X, Wang J, Chen X, Zhang H, Kim SH, Cui J, Li R, Zhang W, Kong Y, Zhang J, Shui W, Lamplot J, Rogers MR, Zhao C, Wang N, Rajan P, Tomal J, Statz J, Wu N, Luu HH, Haydon RC, He TC (2013) Wnt signaling in bone formation and its therapeutic potential for bone diseases. Ther Adv Musculoskelet Dis 5:13–31. https://doi.org/10.1177/1759720X12466608
Korish AA, Abdel Gader AG, Korashy HM, Al-Drees AM, Alhaider AA, Arafah MM (2015) Camel milk attenuates the biochemical and morphological features of diabetic nephropathy: inhibition of Smad1 and collagen type IV synthesis. Chem Biol Interact 229:100–108. https://doi.org/10.1016/j.cbi.2015.01.013
Laszlo E, Varga A, Kovacs K, Jancso G, Kiss P, Tamas A, Szakaly P, Fulop B, Reglodi D (2015) Ischemia/reperfusion-induced kidney injury in heterozygous PACAP deficient mice. Transplant Proc 47:2210–2215. https://doi.org/10.1016/j.transproceed.2015.07.027
Lochab AK, Extavour CG (2017) Bone morphogenetic protein (BMP) signaling in animal reproductive system development and function. Dev Biol 427:258–269. https://doi.org/10.1016/j.ydbio.2017.03.002
Menger MD, Vollmar B (2007) Pathomechanisms of ischemia-reperfusion injury as the basis for novel preventive strategies: is it time for the introduction of pleiotropic compounds? Transplant Proc 39:485–488. https://doi.org/10.1016/j.transproceed.2007.01.022
Mori H, Nakamachi T, Ohtaki H, Yofu S, Sato A, Endo K, Iso Y, Suzuki H, Takeyama Y, Shintani N, Hashimoto H, Baba A, Shioda S (2010) Cardioprotective effect of endogenous pituitary adenylate cyclase-activating polypeptide on doxorubicin-induced cardiomyopathy in mice. Circ J 74:1183–1190
Nakamachi T, Ohtaki H, Seki T, Yofu S, Kagami N, Hashimoto H, Shintani N, Baba A, Mark L, Lanekoff I, Kiss P, Farkas J, Reglodi D, Shioda S (2016) PACAP suppresses dry eye signs by stimulating tear secretion. Nat Commun 7:12034. https://doi.org/10.1038/ncomms12034
Neugarten J, Golestaneh L (2013) Gender and the prevalence and progression of renal disease. Adv Chronic Kidney Dis 20:390–395. https://doi.org/10.1053/j.ackd.2013.05.004
Nguyen V, Meyers CA, Yan N, Agarwal S, Levi B, James AW (2017) BMP-2-induced bone formation and neural inflammation. J Orthop 14:252–256. https://doi.org/10.1016/j.jor.2017.03.003
Parsons RL, May V (2018) PACAP-induced PAC1 receptor internalization and recruitment of endosomal signaling regulate cardiac neuron excitability. J Mol Neurosci. https://doi.org/10.1007/s12031-018-1127-x
Reglodi D, Lubics A, Kiss P, Lengvari I, Gaszner B, Toth G, Hegyi O, Tamas A (2006) Effect of PACAP in 6-OHDA-induced injury of the substantia nigra in intact young and ovariectomized female rats. Neuropeptides 40:265–274. https://doi.org/10.1016/j.npep.2006.06.001
Reglodi D, Kiss P, Lubics A, Tamas A (2011) Review of the protective effects of PACAP in models of neurodegenerative diseases in vitro and in vivo. Curr Pharm Des 17:962–972
Reglodi D, Kiss P, Horvath G, Lubics A, Laszlo E, Tamas A, Racz B, Szakaly P (2012a) Effects of pituitary adenylate cyclase activating polypeptide in the urinary system, with special emphasis on its protective effects in the kidney. Neuropeptides 46:61–70. https://doi.org/10.1016/j.npep.2011.05.001
Reglodi D, Kiss P, Szabadfi K, Atlasz T, Gabriel R, Horvath G, Szakaly P, Sandor B, Lubics A, Laszlo E, Farkas J, Matkovits A, Brubel R, Hashimoto H, Ferencz A, Vincze A, Helyes Z, Welke L, Lakatos A, Tamas A (2012b) PACAP is an endogenous protective factor-insights from PACAP-deficient mice. J Mol Neurosci 48:482–492. https://doi.org/10.1007/s12031-012-9762-0
Reglodi D, Renaud J, Tamas A, Tizabi Y, Socías B, Del-Bel E, Raisman-Vozari R (2017) Novel tactics for neuroprotection in Parkinson’s disease: role of antibiotics, polyphenols and neuropeptides. Prog Neurobiol 155:120–148. https://doi.org/10.1016/j.pneurobio.2015.10.004
Sandhi J, Singh JP, Kaur T, Ghuman SS, Singh AP (2014) Involvement of progesterone receptors in ascorbic acid-mediated protection against ischemia-reperfusion-induced acute kidney injury. J Surg Res 187:278–288. https://doi.org/10.1016/j.jss.2013.10.025
Sasaki S, Watanabe J, Ohtaki H, Matsumoto M, Murai N, Nakamachi T, Hannibal J, Fahrenkrug J, Hashimoto H, Watanabe H, Sueki H, Honda K, Miyazaki A, Shioda S (2017) Pituitary adenylate cyclase-activating polypeptide promotes eccrine gland sweat secretion. Br J Dermatol 176:413–422. https://doi.org/10.1111/bjd.14885
Szabo A, Danyadi B, Bognar E, Szabadfi K, Fabian E, Kiss P, Mester L, Manavalan S, Atlasz T, Gabriel R, Toth G, Tamas A, Reglodi D, Kovacs K (2012) Effect of PACAP on MAP kinases, Akt and cytokine expressions in rat retinal hypoperfusion. Neurosci Lett 523:93–98. https://doi.org/10.1016/j.neulet.2012.06.044
Talebi N, Nematbakhsh M, Monajemi R, Mazaheri S, Talebi A, Vafapour M (2016) The protective effect of γ-aminobutyric acid on kidney injury induced by renal ischemia-reperfusion in ovariectomized estradiol-treated rats. Int J Prev Med 7:6. https://doi.org/10.4103/2008-7802.173796
Tanaka R, Yazawa M, Morikawa Y, Tsutsui H, Ohkita M, Yukimura T, Matsumura Y (2017) Sex differences in ischaemia/reperfusion-induced acute kidney injury depends on the degradation of noradrenaline by monoamine oxidase. Clin Exp Pharmacol Physiol 44:371–377. https://doi.org/10.1111/1440-1681.12713
Vafapour M, Nematbakhsh M, Monajemi R, Mazaheri S, Talebi A, Talebi N, Shirdavani S (2015) Effect of Γ-aminobutyric acid on kidney injury induced by renal ischemia-reperfusion in male and female rats: gender-related difference. Adv Biomed Res 4:158. https://doi.org/10.4103/2277-9175.161585
Vaudry D, Falluel-Morel A, Bourgault S, Basille M, Burel D, Wurtz O, Fournier A, Chow BK, Hashimoto H, Galas L, Vaudry H (2009) Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol Rev 61:283–357. https://doi.org/10.1124/pr.109.001370
Vu JP, Benhammou JN, Goyal D, Luong L, Oh S, Germano P, Pisegna JR (2016) PACAP regulation of gastrointestinal function and obesity. In: Reglodi D, Tamas A (eds) Pituitary adenylate cyclase activating polypeptide – PACAP. Springer, Switzerland, pp 261–270
Watanabe J, Seki T, Shioda S (2016) PACAP and neural development. In: Reglodi D, Tamas A (eds) Pituitary adenylate cyclase activating polypeptide - PACAP. Springer, Switzerland, pp 65–83
Werling D, Banks WA, Salameh TS, Kvarik T, Kovacs LA, Vaczy A, Szabo E, Mayer F, Varga R, Tamas A, Toth G, Biro Z, Atlasz T, Reglodi D (2017) Passage through the ocular barriers and beneficial effects in retinal ischemia of topical application of PACAP1-38 in rodents. Int J Mol Sci 18:675. https://doi.org/10.3390/ijms18030675
Yang CW, Lim SW, Han KW, Ahn HJ, Park JH, Kim YH, Kirsh M, Cha JH, Park JH, Kim YS, Kim J, Bang BK (2001) Upregulation of ciliary neurotrophic factor (CNTF) and CNTF receptor alpha in rat kidney with ischemia-reperfusion injury. J Am Soc Nephrol 12:749–757
Yu R, Cui Z, Li M, Yang Y, Zhong J (2014) Dimer-dependent intrinsic/basal activity of the class B G protein-coupled receptor PAC1 promotes cellular anti-apoptotic activity through Wnt/β-catenin pathways that are associated with dimer endocytosis. PLoS One 9:e113913. https://doi.org/10.1371/journal.pone.0113913
Funding
This study was financially supported by the following: MTA-TKI 14016, EFOP-3.6.2-16-2017-00008 “The role of neuro-inflammation in neurodegeneration: from molecules to clinics”/Center for neuroscience, PTE AOK Research Grant KA-2017-17, NKFIH119759, 2017-1.2.1-NKP-2017-00002, Bolyai Scholarship, GINOP-2.3.2-15-2016-00050 “PEPSYS.” EFOP-3.6.1.-16-2016-00004—Comprehensive Development for Implementing Smart Specialization Strategies at the University of Pécs, Higher Education Institutional Excellence Program of the Ministry of Human Capacities in Hungary, within the framework of the 20765-3/2018/FEKUTSTRAT. EFOP-3.6.3-VEKOP-16-2017-00009.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Experimental procedures were carried out under approved protocols BA02/2000-24/2011.
Rights and permissions
About this article

Cite this article
Laszlo, E., Juhasz, T., Varga, A. et al. Protective Effect of PACAP on Ischemia/Reperfusion-Induced Kidney Injury of Male and Female Rats: Gender Differences. J Mol Neurosci 68, 408–419 (2019). https://doi.org/10.1007/s12031-018-1207-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12031-018-1207-y