
Abstract
Pericytes surrounding endothelial cells in the capillaries are emerging as an attractive cell resource, which can show a large variety of functions in ischemic stroke, including preservation of the blood–brain barrier, regulation of immune function, and support for cerebral vasculature. These functions have been fully elucidated in previous studies. However, in recent years, increasing evidence has shown that pericytes play an important role in neurological recovery after ischemic stroke due to their regenerative function which can be summarized in two aspects according to current discoveries, one is that pericytes are thought to be multipotential themselves, and the other is that pericytes can promote the differentiation of oligodendrocyte progenitor cells (OPCs). Considering the neuroprotective treatment for stroke has not been much progressed in recent years, new therapies targeting pericytes may be a future direction. Here, we will review the beneficial effects of pericytes in ischemic stroke from two directions: the barrier and vascular functions and the regenerative functions of pericytes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Globally, stroke is the second-leading cause of death and the third leading cause of death and disability combined in 2019 (GBD 2019 Stroke Collaborators, 2021). Ischemic stroke is the most common subtype of stroke. In 2019, approximately 73% of stroke cases are ischemic strokes in China (Ma et al., 2021), compared to 62% worldwide (GBD 2019 Stroke Collaborators, 2021). Although multiple innovative approaches to improve the prognosis of ischemic stroke (e.g., reperfusion therapy (Albers et al., 2018; Hacke et al., 2008; Ma et al., 2019; National Institute of Neurological Disorders & Stroke rt-PA Stroke Study Group, 1995; Nogueira et al., 2018; Suzuki et al., 2019; Thomalla et al., 2018; Yang et al., 2020), and neuroprotective therapy (Hill et al., 2020; Xu et al., 2021)) have flourished over the past few decades (Herpich & Rincon, 2020), the treatment of ischemic stroke has now reached a plateau. We have intensely and often been hindered in the translation of preclinical studies into successful clinical studies (Lyden, 2021), particularly in neuroprotective therapy (Paul & Candelario-Jalil, 2021). At the 2017 Stroke Treatment Academic Industry Roundtable X (STAIR X), the concept of Brain Cytoprotection was first proposed because stroke affects not only neurons but also the entire neurovascular unit and white matter (Savitz et al., 2019).
The Neurovascular Unit (NVU) was formalized in 2001 and has drawn attention to the interdependence between brain cells and cerebral blood vessels (Iadecola, 2017). The NVU consists of vascular components (pericytes, smooth muscle cells, endothelial cells), glial cells (astrocytes, oligodendrocytes, microglia), and neurons (Harder et al., 2002; Lo & Rosenberg, 2009; Lo et al., 2003). Interestingly, almost every neuron in the human brain has its own capillaries which account for more than 90% of the total blood vascular volume in the brain (Zlokovic, 2008). Therefore, impaired perfusion of the microvasculature after recanalization therapy can lead to lower-than-expected clinical outcomes (Ames et al., 1968; Goyal et al., 2016). As a member of the vascular part in the NVU, pericytes are present in small cerebral vessels including capillaries, pre-capillary arterioles, and post-capillary venules (Winkler et al., 2011). Multiple studies have highlighted the important role of pericytes in the NVU (Sweeney et al., 2016), such as regulating Blood–Brain Barrier (BBB) permeability (Zlokovic, 2011), neuroinflammation (Rustenhoven et al., 2017), and cerebral blood flow (CBF) (Hall et al., 2014). While these studies focused on the barrier and vascular functions of pericytes, more recent studies have shifted attention to the pluripotent stem cell potential (Sakuma et al., 2016) and the promotion of white matter functions (Shibahara et al., 2020a).
Data show that in previous randomized trials, about 70% of patients treated with reperfusion successfully (modified Thrombolysis in Cerebral Infarction (mTICI) scale score 2b or 3), but only 27% of these patients were disability-free at 90 days (Goyal et al., 2016). This may be partly due to the irreversible injury of brain tissue before reperfusion occurred. Therefore, if pericytes have the ability to transform into other neurovascular unit components or promote white matter differentiation, then treatment of pericytes will play a key role in the recovery of motor, sensory, and emotional impairment after stroke.
In this review, we will start with the morphological structure and the vascular homeostatic functions of pericytes and then focus on the latest research on regenerative characteristics of pericytes which will provide new directions for the treatment of ischemic stroke.
Pericytes in Central Nervous System (CNS)
Since the pericytes were first discovered by Eberth (1871), described by Rouget (1873) as a cluster of contractible cells surrounding endothelial cells, and eventually named “pericytes” by Zimmermann (1923), the studies of pericytes have become increasingly popular in the last 50 years, especially in the brain due to the development of electron microscopy (Caporali et al., 2017). In vertebrates, pericytes are found in almost all tissues and located on the abluminal side of the endothelium in both continuous and fenestrated microvessels (Díaz-Flores et al., 2009). The morphology of pericytes is largely related to location (Joyce et al., 1984) and type of vessels (Armulik et al., 2011). In terms of the location of the pericytes, they can be divided into three types: precapillary, capillary, and postcapillary (Zimmermann, 1923). Usually, pericytes display an elongated, stellate morphology containing a cell body, nuclear region, or perinucleus, and produce a highly branched structure consisting of longitudinal and circumferential branching systems that wrap the endothelium. The primary (longitudinal) processes parallel to the long axis of the vessel, with smaller ones (circumferential) proportionally encircling the vessel wall (Díaz-Flores et al., 2009; Takahashi et al., 1997). Using Cre-recombinase driver mouse lines, Hartmann et al. identified that pericytes can be distinguished as helical pericytes and mesh pericytes in capillaries based on the morphology of the trunk and branches. The helical pericyte is the simplest form, with the primary trunk and branches being thin singular strands, approximately 2 µm in diameter, and the secondary processes branching off from the thin single strands, often present in pairs, forming a helical structure. As for its most complex form, is more commonly found in larger diameter microvessels (6–10 µm). The primary trunk of it shapes a mesh-like structure surrounding the entire vessel, hence comes the name mesh pericyte (Hartmann et al., 2015) (Fig. 1). Pericytes of the CNS and retina have the highest coverage ratio (Daneman et al., 2010). Using Electron Microscopy and 3D Reconstruction, Mathiisen TM et.al demonstrated that pericytes and their protrusions covered 37% of the endothelial tube circumference (Mathiisen et al., 2010), which can partially explain the important role of pericytes in the BBB permeability (Daneman et al., 2010). The ratio of pericytes to endothelial cells and the area of pericytes covering the endothelium are related to the tightness of the junctions between the endothelium, that is, the higher the number of pericytes and their coverage is, the better the microvascular barrier integrity works (Shepro & Morel, 1993).

Two types of pericytes and NVU. According to the morphology of the trunk and branches, pericytes can be distinguished as helical pericytes and mesh pericytes in capillaries. a The helical pericyte: the primary trunk and branches are thin singular strands, and the secondary processes branch off from the thin single strands, forming a helical structure. b The mesh pericyte: the primary trunk shapes a mesh-like structure surrounding the entire vessel, and is more commonly found in larger diameter microvessels (6–10 µm). c NVU: The NVU is composed of vascular components, glial cells, and neurons
The Temporal Dynamics of Pericyte Functions in Ischemic Stroke
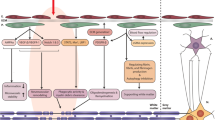
In the acute phase of ischemic stroke, pericytes mainly exhibit the function of constricting blood vessels (Korte et al., 2022), which may further aggravate the injury of cerebral infarction and cause no-reflow phenomenon after reperfusion (Yemisci et al., 2009). Immediately after performing the vasoconstrictive function, pericytes die or separate from blood vessels (Hall et al., 2014), resulting in decreased pericyte coverage and increased BBB permeability in the acute phase. The function of mediating neuroinflammation also manifests after BBB disruption (Rustenhoven et al., 2017). 72 h after ischemic stroke, the coverage of pericytes recovers (Zhou et al., 2018a) and simultaneously exerts the protective effect of the BBB and the function of angiogenesis (Zhang et al., 2022). In the late recovery phase of ischemic stroke, the regenerative function of pericytes gradually appeared (Yang et al., 2017) (Fig. 2).

The temporal dynamics of pericyte functions in ischemic stroke. Pericytes mainly exhibit the function of constricting blood vessels and increasing BBB permeability in the acute phase. Following vasoconstrictive action, pericytes die or separate immediately from blood vessels. The function of mediating neuroinflammation also manifests after BBB disruption and lasts a long time. In the subacute phase, pericytes exert the protective effect of the BBB and the function of angiogenesis. In the late chronic phase of ischemic stroke, the regenerative function of pericytes appeared
Barrier and Vascular Functions of Pericytes
Maintaining BBB Integrity
Pericytes share a basement membrane (BM) with endothelial cells and are also covered by a BM continuous with the BM of endothelial cells (Ayloo et al., 2022). This allows for sophisticated cell-to-cell interactions between two cells. In areas where the direct connection is absent, interdigitations of pericyte and endothelial cell membranes form peg-and-socket contacts (Sims, 1991) which contain tight-, gap-, and adherence junctions (Armulik et al., 2005). Although endothelial cells perform most of the properties of the BBB (Daneman, 2012), their presence alone does not keep the BBB functioning properly (Ayloo et al., 2022). The intricate connection between pericytes and endothelial cells determines the important role of pericytes in BBB. Also, previous studies have demonstrated that pericytes help regulate the BBB from embryogenesis (Armulik et al., 2010; Daneman et al., 2010). Platelet-derived growth factor-BB/platelet-derived growth factor receptor-beta (PDGF-BB/PDGFRβ) signaling and Angiopoietin (Ang)/Tie2 system have a critical role in BBB stabilization throughout growth and development (Sweeney et al., 2016). Lack of endothelial-secreted PDGF-BB or loss of pericyte PDGFRβ can lead to disruption of BBB integrity (Winkler et al., 2010), resulting in embryonic lethality or various neurological disorders. Angiopoietin-1 (Ang-1) has been shown to be constitutively expressed in pericytes and activates the Tie2 receptor in endothelial cells to prevent vascular leakage (Gurnik et al., 2016). While endothelial cell-derived angiopoietin-2 (Ang-2) is reported to increase both the paracellular and the transcellular permeability at the BBB in a mouse stroke model (Gurnik et al., 2016). N-cadherin plays a significant role in endothelial cell-pericyte interactions mediated by brain endothelial cells Smad4 and is thought to be an initial signal for BBB development and increases in angiogenic vessels (Li et al., 2011). Pericyte-endothelial cell interactions may not be unique. Ando et al. found that a pericyte contacts multiple endothelial cells and can extend to more than one capillary (Ando et al., 1999), which means it integrates signals along the length of one vessel and also communicates with other vessels (Bergers & Song, 2005).
In addition to maintaining the BBB through signaling with endothelial cells, the existence of pericytes can also reduce the transmembrane transport of endothelial cells. Mfsd2a−/− mice exhibit a sharp increase in vesicular transport under electron microscopy without variation of endothelial tight junctions, leading to leakage of BBB from embryonic to adult periods (Ben-Zvi et al., 2014). A recent study demonstrates that vitronectin-integrin α5 signaling from pericytes to endothelial cells maintains barrier integrity by actively inhibiting transcytosis in endothelial cells (Ayloo et al., 2022). A single-cell RNA analysis of pericytes-function-loss mice found that hotspot sites were hallmarked by low Mfsd2a and low Ang-2 expression (Mäe et al., 2021) (Fig. 3). Studies related to the interaction between pericytes and endothelial cells in ischemic stroke are also emerging. In oxygen deprivation models, pericytes have a stronger ability to maintain blood–brain barrier integrity than astrocytes during severe and prolonged hypoxic conditions (Abbott, 2002; Al Ahmad et al., 2009). Fernández-Klett F el at. found that early pericyte death (within 24 h) preceded endothelial cell degeneration (after day 5) in the MCAO model (Fernández-Klett et al., 2013).

Cell-to-cell interactions between pericytes and endothelial cells. Pericytes share a BM with endothelial cells and are also covered by a BM. PDGF-BB/PDGFRβ signaling and Ang/Tie2 system are the basis for maintaining the stability of the BBB. Cell-to-cell interactions between two cells keep the BBB functioning properly. In areas where the BM is absent, pericytes and endothelial cells membranes form peg-and-socket contacts. The transmembrane transport in endothelial cells can be reduced by the existence of pericytes through Mfsd2a and vitronectin-integrin α5 signaling
The crosstalk between pericytes and astrocytes also plays an important role in maintaining BBB integrity. Astrocytic end-feet express the water channel protein aquaporin-4 (AQP4), which plays a vital role in the integrity of the BBB (Abbott et al., 2006). Under normal conditions, pericytes regulate the distribution of AQP4 in the end-feet of astrocytes (Armulik et al., 2010; Gundersen et al., 2014). When pericytes are injured in ischemic stroke, the regulator of G-protein signaling 5 (RGS5) is upregulated before they detach from the blood vessels (Özen et al., 2014). And in a subsequent study, it was proved that the loss of RGS5 in pericytes contributes to the retention of AQP4 in the astrocyte end-feet and plays a neurovascular protective role (Özen et al., 2018). Similarly, astrocytes also help pericytes maintain BBB stability by secreting laminin (Yao et al., 2014). In contrast, semaphorin 4D (Sema4D) from astrocytes binds to PlexinB1 in pericytes and disrupts BBB integrity after stroke in rats (Zhou et al., 2018b.
Putting aside the association between pericytes and neighboring cells, only the pericytes themselves were taken out. Pericytes are an important source of extracellular matrix (ECM) in the BM (Xu et al., 2019). There are four major ECM proteins in the BM: collagen IV, laminin, nidogen, and perlecan (Yurchenco, 2011). Pericyte-derived laminin is associated with the maintenance of BBB permeability (Gautam et al., 2016). However, the effect of laminin on BBB is controversial in relation to the type of BBB. Lamininα5 and lamininα4 derived from mural cells may attenuate BBB damage during intracerebral hemorrhage by reducing transcytosis (Gautam et al., 2020). On the contrary, lamininα5 was demonstrated to play a negative role in ischemic stroke, and lamininα5-PKO mice exhibited milder neuronal injury and attenuated vascular damage, suggesting that inhibition of that signaling may have a neuroprotective effect (Nirwane et al., 2019). Also, it was found that endothelial cell-derived perlecan is upregulated in the BM, a process critical to the repair of BBB functions after ischemic stroke (Nakamura et al., 2019). Although current studies on pericyte-associated BM modifications after stroke mainly focus on laminin and perlecan, perhaps BM-targeted therapy is a path forward (Kang & Yao, 2020). HIF-1 once thought to be a protective factor, is produced under hypoxic conditions in the cardiovascular system (Bishop & Ratcliffe, 2015). However, HIF-1 loss-of-function in a model of cerebral ischemia shows less pericyte death, resulting in broader vascular coverage and better integrity in BBB. Therefore, that reduces the degree of infarction and cerebral edema post-stroke (Tsao et al., 2021). From a perspective other than molecular, Tunneling Nanotubes (TNT), open membranous channels for cell-to-cell communication (Rustom et al., 2004), were thought to show a functional role in the crosstalk among BBB. Ischemia-induced astrocyte apoptosis is reduced by TNT-mediated mitochondrial transfer of pericytes (Pisani et al., 2022), which allows for repair of the BBB after ischemic injury.
Recently, increasing studies have found pericytes to be heterogeneous (Dias Moura Park et al., 2016; Prazeres et al., 2017), implying that pericytes may exhibit different features after stroke. Type-1 pericytes are considered to be physiological capillary PCs, while type-2 pericytes are pathological. Type-2 pericytes were first described in skeletal muscle using Nestin-GFP/NG2-DsRed transgenic mice (Birbrair et al., 2013). And then they were claimed to be recruited during tumor angiogenesis (Birbrair et al., 2014). In the brain, PDGFRB + /SMA + /MYH11- (type-2) pericytes may be a cellular biomarker associated with the degree of BBB disruption, not limited to the disease state (Bohannon et al., 2020).
It has been proved that imperfect BBB recovery post-stroke may increase the risk of aftermath events and cognitive impairment (Taheri et al., 2011). Cerebral pericytes are integral components of the neurovascular unit, which governs the BBB (Ding et al., 2021). They are indispensable for BBB, so the opinion that maintaining pericytes function contributes to stroke recovery is increasingly reinforced.
Regulating Cerebral Blood Flow
Pericytes surround around blood vessels and constrict capillaries, leading to a decrease in CBF (Fernández-Klett et al., 2010; Peppiatt et al., 2006). In addition to the constrict ability, pericytes can be relaxed by neuronal activity and the neurotransmitter glutamate to dilate capillaries (Hall et al., 2014). However, the function of regulating CBF is unclear due to the difficulties in distinguishing pericytes from vascular smooth muscle cells (SMCs). What is clear is that SMCs on arterioles and ensheathing pericytes on pre-capillary arterioles (pre-capillary SMCs) control blood flow (Hill et al., 2015). To precisely identify these two types of cells, many markers have been studied in recent years and have been well reviewed (Bohannon et al., 2021; Grant et al., 2019; Zheng et al., 2020). Meanwhile, a technology to label pericytes in live models without marking SMCs was developed in 2017 (Damisah et al., 2017). In addition to difficulties in identification, the intrinsic connectivity of the cerebrovascular system hinders the certainty of pericyte regulation of CBF function of pericytes. Recently, the optical ablation of single capillary pericytes was used to isolate the effect of pericyte loss on local blood flow. The results suggest that capillary pericytes can modulate capillary diameter, influence blood flow in vivo, and establish basal capillary flow resistance (Hartmann et al., 2021). Bohannon et al. demonstrated that pericytes first constrict the capillaries and then end up with death when capillaries are exposed to ischemia (Hall et al., 2014). After stroke, preventing pericytes from shrinking and dying may reduce long-term blood flow to injured neurons. Nonetheless, novel methods are needed to further study the role of pericytes in regulating blood flow.
Mediating Neuroinflammation
Neuroinflammation is involved in the pathophysiology of almost all neurological diseases (Rustenhoven et al., 2017). In ischemic stroke, this process probably includes oxidative stress, increased matrix metalloproteinase (MMP) production, infiltration of peripheral immune cells, and activation of microglia and astrocytes (Candelario-Jalil et al., 2022).
The role of pericytes in neuroinflammation has been studied for decades (Rustenhoven et al., 2017). Pericytes of the central nervous system were thought to have the ability to present antigens to T-lymphocytes (Balabanov et al., 1999). Then in vitro experiments which were stimulated with tumor necrosis factor (TNF) or lipopolysaccharide (LPS) demonstrated pericytes have the ability to detect inflammation because they have pattern recognition receptors (Guijarro-Muñoz et al., 2014; Stark et al., 2013). These studies reveal the active role of pericytes in innate immune responses, concluded to support immune surveillance.
Furthermore, in the experimental autoimmune encephalomyelitis (EAE) models, the infiltration of leukocytes into the CNS was negatively correlated with the coverage of pericytes in the vasculature (Török et al., 2021). However, it is not entirely clear whether CNS pericytes exhibit a pro- or anti-inflammatory profile (Rustenhoven et al., 2016). The polarity of pericytes may be similar to that of microglia in neuroinflammation under an ischemic environment (Ma et al., 2017). Therefore, more studies are needed to demonstrate the role of pericytes in neuroinflammation in ischemic stroke.
Promoting Angiogenesis
New blood vessels are formed through angiogenesis, a multi-factory process requiring synchrony between endothelial cells and pericytes (Mastrullo et al., 2020). Angiogenesis after stroke can mitigate hypoxia-induced damage caused by ischemia (Ergul et al., 2012). And in theory, angiogenic therapy can save the ischemic border zone (Ergul et al., 2012). However, given the notion that treatments that promote angiogenesis may exacerbate stroke outcomes since new angiogenesis-induced vessels are more permeable than usual (Yang & Torbey, 2020). Accordingly, we speculate that the synergy of pericytes especially the function of promoting vascular maturation is important in this process.
In a newly formed blood vessel, pericytes are recruited through communication with endothelial cells, resulting in the formation of a new BM (Stratman et al., 2009). And PDGF-BB/PDGFRβ signal was thought to lead to pericyte recruitment and then stabilize the blood vessel (Gaengel et al., 2009). Also, angiogenesis and vascular integrity in the ischemic brain are partially modulated by pericyte-specific expression of vascular endothelial growth factor receptor 1 (VEGFR1) (Zechariah et al., 2013). When VEGF is present, Ang2/Tie2 signals make the system highly plastic, forming new vessels continuously (Ghori et al., 2022).
Regenerative Functions of Pericytes
Acting as Multipotent Stem Cells
The regenerative potential of pericytes has been discussed in various organs in addition to CNS. Since the discovery of the transformation of pericytes into microglia with the help of astrocytes in the cat cerebral cortex (Barón & Gallego, 1972), the debate on the potential of cerebral pericyte pluripotent stem cells has begun (Table 1).
Using cell markers, nestin/NG2-positive pericytes are considered to be a source of adult stem cells in vitro (Dore-Duffy et al., 2006). Under hypoxic conditions in vitro, human brain-derived pericytes were found to upregulate the expression of activated microglial mRNA, implying that they acquire a microglial cell phenotype (Özen et al., 2014). Furthermore, they showed that pericytes express microglia markers not only in vitro but also in human post-stroke brain tissue. In these observations, ischemia/hypoxia might enhance stem-like activity in brain pericytes, but we do not know which exact mechanisms are essential for their induction (Nakagomi et al., 2011). Reprogramming is reckoned as a pivotal process in transformation following the indications that after ischemia or hypoxia pericyte marker expression was downregulated, while stem cell-like marker was upregulated. Under oxygen/glucose deprivation, pericytes acquire the capacity of multipotential stem cells and can differentiate into major the BBB/neurovascular unit components because of reprogramming (Sakuma et al., 2016). This strategy could be used to induce pluripotency in pericytes to promote regeneration (Karow et al., 2018). Another possible mechanism was recently explained. Oxidative stress after ischemia with the expression of Nrf2 may trigger pericytes to acquire stemness (Sakuma et al., 2022). Similarly, in 2016, Sakuma R et al. revealed that pericytes show multipotent activity in MCAO models indicating that pericytes may be a novel source of microglia after ischemic stroke (Sakuma et al., 2016). Ischemia-induced multipotent stem cells (iSCs) in the human post-stroke brain were first isolated in 2017, and they are likely pericyte derivatives. More importantly, they may help neural repair or regeneration in patients with ischemic stroke (Tatebayashi et al., 2017). The anti-ischemic effect of pericytes due to their multipotency has been used for limb ischemia (Yoshida et al., 2020). Recently, the lineage-tracing technique was used to trace pericyte fates after ischemic stroke. They found that SMAlow/undetectable pericytes differentiated into both microglia and macrophages after the acute period of ischemic stroke (Nirwane & Yao, 2022). In conclusion, these studies suggest that pericytes not only have a strong migratory and proliferative response to ischemic brain injury but also serve as a source of neural lineage cells. While lineage-tracing experiments utilizing an inducible Tbx18-CreERT2 line insist that pericytes as well as vascular smooth muscle cells fail to contribute to other cell lineages, it is noteworthy that this study did not use an ischemia model (Guimarães-Camboa et al., 2017).
Furthermore, apart from transforming into neural lineage cells, pericytes are also revealed to be fibrotic activity. Type A pericytes were first proposed in models of spinal cord injury (Göritz et al., 2011). Blocking the proliferation of such cells will result in the inability to seal the damaged tissue. Interestingly, single-cell analyses later challenged this result suggesting such cells refer to fibroblast-like rather than pericytes (Vanlandewijck et al., 2018). Same to the previous work, Roth M et al. illustrated that the fibrotic ECM is not major coming from pericytes, so targeting pericytes to scar formation after stroke may be useless (Roth et al., 2020). However, type A pericytes were once again shown to be the source of fibrotic ECM, and this result is not limited to ischemic injury but is conserved across diverse central nervous system diseases (Dias et al., 2021). To further prove this idea, a single-cell RNA sequencing analysis was performed revealing that pathways related to fibrosis were enriched in pericytes of cardiac and cerebral ischemic injury (Pham et al., 2021).
The controversy over the presence of pluripotent stem cell potential in pericytes is mainly due to the lack of an appropriate marker for pericytes as well as multipotent pericytes if they existent (Yoshida et al., 2020). Further studies on this area need to be conducted.
Promoting OPCs Differentiation
It has been previously illustrated that more than two of third patients suffered disability after ischemic stroke even if they were successfully treated with reperfusion (Goyal et al., 2016). Cerebral white matter (WM) is particularly vulnerable to vascular occlusion. Numerous studies have demonstrated that WM damage after stroke is associated with long-term sensorimotor deficits and cognitive decline (Arai, 2020; Matute et al., 2013).
The WM is mainly composed of myelinated axons and glial cells. Multiple myelin sheaths produced by oligodendrocytes wrap the axons. Oligodendrocytes as important WM components are originally from OPCs (Bercury & Macklin, 2015). Myelin repair and oligodendrocyte formation in the adult brain are determined by OPCs (Menn et al., 2006) which comprise 5–8% of all the cells in the adult brain and are abundant in both grey and white matter areas (Dawson et al., 2000). Demyelination causes oligodendrocytes to split and differentiate, replacing lost oligodendrocytes with new ones (Levine et al., 2001). Therefore, targeting OPCs to repair WM after ischemic stroke may promote functional recovery. Although there is a vast body of related research on glial cells such as microglia (Shi et al., 2021) and astrocytes (Miyamoto et al., 2015), it mostly falls into pericytes in this section.
It has been reported that in the perivascular region of cerebral WM, pericytes and OPCs may attach and support each other (Maki et al., 2015). Using pericyte-deficient mice, pericyte degeneration was found to disrupt WM microcirculation, which results in an accumulation of toxic blood-derived fibrinogen deposits leading to a loss of myelin, axons, and oligodendrocytes (Montagne et al., 2018). A variety of molecules secreted by pericytes may be required for OPC-to-oligodendrocyte renewal. A-Kinase Anchor Protein 12 (AKAP12) is thought to mainly express on pericytes and is necessary for OPCs function to keep WM homeostasis (Maki et al., 2018). Lama2 has also been identified as a pericyte-derived factor that promotes OPCs differentiation in multiple sclerosis (MS) (De La Fuente et al., 2017). It is shown to support OPCs differentiation into oligodendrocytes without affecting remyelination from OPCs in vivo. Besides functioning in demyelinating diseases, pericytes may promote peri-infarct oligodendrogenesis after ischemic stroke resulting in functional recovery (Shibahara et al., 2020a). The crosstalk between pericytes and macrophages may be critical for this procession in poststroke tissue repair (Shibahara et al., 2020b). However, the origin of pericytes in the infarct area should be further validated.
It is important to note that neurological disorders associated with cognitive dysfunction, cerebrovascular dysfunction, and WM lesions are characterized by a loss of pericyte coverage (Ding et al., 2020). Thus, the functional role of pericytes is not limited to vascular homeostasis but also includes modulating the progenitor cells of adult CNS regeneration.
Conclusions
We summarize the functions of pericytes after stroke as maintaining BBB, regulating CBF, mediating immune responses, promoting angiogenesis, acting as pluripotent stem cells, and promoting OPCs differentiation. Accordingly, pericytes are a promising source of cells for cell therapy and tissue engineering. In the acute phase, inhibiting the contractile function of pericytes and enhancing the function of protecting the BBB may help reduce the occurrence of hypoperfusion, edema, and increased infarct size after ischemic stroke. In subacute and late phases, focusing on the regenerative function of pericytes may contribute to neurological recovery. However, there is still much controversy surrounding the studies of pericytes. Firstly, accurate cellular markers for pericytes need to be identified so that the function of pericytes can be distinguished from vascular smooth muscle cells. Secondly, whether there is the heterogeneity of pericytes in the pathological process after stroke should be further studied. If pericytes, like microglia, are polarized, then drugs that target the polarization of pericytes may be effective in the treatment of stroke. Single-cell studies on pericyte heterogeneity are now available (Vanlandewijck et al., 2018). This may be the way forward as single-cell studies have the ability to differentiate between cell subpopulations (Liu & Zhang, 2022; Qiu et al., 2021). Finally, Research on pericytes after ischemic stroke is more limited to in vitro studies. In vivo, it remains unclear whether they differentiate into neural lineage cells. There is still a long way from clinical trials, so more attention should be paid to pericytes in ischemic stroke. Overall, based on the properties and functions that have been identified so far, further research on pericytes may provide new directions for the treatment of neurological recovery after stroke.
Data availability
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
References
Abbott, N. J. (2002). Astrocyte-endothelial interactions and blood-brain barrier permeability. Journal of Anatomy, 200(6), 629–638. https://doi.org/10.1046/j.1469-7580.2002.00064.x
Abbott, N. J., Rönnbäck, L., & Hansson, E. (2006). Astrocyte-endothelial interactions at the blood-brain barrier. Nature Reviews. Neuroscience, 7(1), 41–53. https://doi.org/10.1038/nrn1824
Al Ahmad, A., Gassmann, M., & Ogunshola, O. O. (2009). Maintaining blood-brain barrier integrity: Pericytes perform better than astrocytes during prolonged oxygen deprivation. Journal of Cellular Physiology, 218(3), 612–622. https://doi.org/10.1002/jcp.21638
Albers, G. W., Marks, M. P., Kemp, S., Christensen, S., Tsai, J. P., Ortega-Gutierrez, S., McTaggart, R. A., Torbey, M. T., Kim-Tenser, M., Leslie-Mazwi, T., Sarraj, A., Kasner, S. E., Ansari, S. A., Yeatts, S. D., Hamilton, S., Mlynash, M., Heit, J. J., Zaharchuk, G., Kim, S., et al. (2018). Thrombectomy for stroke at 6 to 16 hours with selection by perfusion imaging. The New England Journal of Medicine, 378(8), 708–718. https://doi.org/10.1056/NEJMoa1713973
Ames, A., Wright, R. L., Kowada, M., Thurston, J. M., & Majno, G. (1968). Cerebral ischemia: II—The no-reflow phenomenon. The American Journal of Pathology, 52(2), 437–453.
Ando, M., Kakigi, A., & Takeuchi, S. (1999). Elongated pericyte-like cells connect discrete capillaries in the cochlear stria vascularis of gerbils and rats. Cell and Tissue Research, 296(3), 673–676. https://doi.org/10.1007/s004410051327
Arai, K. (2020). Can oligodendrocyte precursor cells be a therapeutic target for mitigating cognitive decline in cerebrovascular disease? Journal of Cerebral Blood Flow and Metabolism, 40(8), 1735–1736. https://doi.org/10.1177/0271678X20929432
Armulik, A., Abramsson, A., & Betsholtz, C. (2005). Endothelial/pericyte interactions. Circulation Research, 97(6), 512–523. https://doi.org/10.1161/01.RES.0000182903.16652.d7
Armulik, A., Genové, G., & Betsholtz, C. (2011). Pericytes: Developmental, physiological, and pathological perspectives, problems, and promises. Developmental Cell, 21(2), 193–215. https://doi.org/10.1016/j.devcel.2011.07.001
Armulik, A., Genové, G., Mäe, M., Nisancioglu, M. H., Wallgard, E., Niaudet, C., He, L., Norlin, J., Lindblom, P., Strittmatter, K., Johansson, B. R., & Betsholtz, C. (2010). Pericytes regulate the blood-brain barrier. Nature, 468(7323), 557–561. https://doi.org/10.1038/nature09522
Ayloo, S., Lazo, C. G., Sun, S., Zhang, W., Cui, B., & Gu, C. (2022). Pericyte-to-endothelial cell signaling via vitronectin-integrin regulates blood-CNS barrier. Neuron, 110(10), 1641-1655.e6. https://doi.org/10.1016/j.neuron.2022.02.017
Balabanov, R., Beaumont, T., & Dore-Duffy, P. (1999). Role of central nervous system microvascular pericytes in activation of antigen-primed splenic T-lymphocytes. Journal of Neuroscience Research, 55(5), 578–587. https://doi.org/10.1002/(SICI)1097-4547(19990301)55:5%3c578::AID-JNR5%3e3.0.CO;2-E
Barón, M., & Gallego, A. (1972). The relation of the microglia with the pericytes in the cat cerebral cortex. Zeitschrift Fur Zellforschung Und Mikroskopische Anatomie, 128(1), 42–57. https://doi.org/10.1007/BF00306887
Ben-Zvi, A., Lacoste, B., Kur, E., Andreone, B. J., Mayshar, Y., Yan, H., & Gu, C. (2014). Mfsd2a is critical for the formation and function of the blood-brain barrier. Nature, 509(7501), 507–511. https://doi.org/10.1038/nature13324
Bercury, K. K., & Macklin, W. B. (2015). Dynamics and mechanisms of CNS myelination. Developmental Cell, 32(4), 447–458. https://doi.org/10.1016/j.devcel.2015.01.016
Bergers, G., & Song, S. (2005). The role of pericytes in blood-vessel formation and maintenance. Neuro-Oncology, 7(4), 452–464. https://doi.org/10.1215/S1152851705000232
Birbrair, A., Zhang, T., Wang, Z.-M., Messi, M. L., Enikolopov, G. N., Mintz, A., & Delbono, O. (2013). Role of pericytes in skeletal muscle regeneration and fat accumulation. Stem Cells and Development, 22(16), 2298–2314. https://doi.org/10.1089/scd.2012.0647
Birbrair, A., Zhang, T., Wang, Z.-M., Messi, M. L., Olson, J. D., Mintz, A., & Delbono, O. (2014). Type-2 pericytes participate in normal and tumoral angiogenesis. American Journal of Physiology. Cell Physiology, 307(1), C25-38. https://doi.org/10.1152/ajpcell.00084.2014
Bishop, T., & Ratcliffe, P. J. (2015). HIF hydroxylase pathways in cardiovascular physiology and medicine. Circulation Research, 117(1), 65–79. https://doi.org/10.1161/CIRCRESAHA.117.305109
Bohannon, D. G., Long, D., & Kim, W.-K. (2021). Understanding the heterogeneity of human pericyte subsets in blood-brain barrier homeostasis and neurological diseases. Cells, 10(4), 890. https://doi.org/10.3390/cells10040890
Bohannon, D. G., Okhravi, H. R., Kim, J., Kuroda, M. J., Didier, E. S., & Kim, W.-K. (2020). A subtype of cerebrovascular pericytes is associated with blood-brain barrier disruption that develops during normal aging and simian immunodeficiency virus infection. Neurobiology of Aging, 96, 128–136. https://doi.org/10.1016/j.neurobiolaging.2020.08.006
Candelario-Jalil, E., Dijkhuizen, R. M., & Magnus, T. (2022). Neuroinflammation, stroke, blood-brain barrier dysfunction, and imaging modalities. Stroke, 53(5), 1473–1486. https://doi.org/10.1161/STROKEAHA.122.036946
Caporali, A., Martello, A., Miscianinov, V., Maselli, D., Vono, R., & Spinetti, G. (2017). Contribution of pericyte paracrine regulation of the endothelium to angiogenesis. Pharmacology & Therapeutics, 171, 56–64. https://doi.org/10.1016/j.pharmthera.2016.10.001
Damisah, E. C., Hill, R. A., Tong, L., Murray, K. N., & Grutzendler, J. (2017). A fluoro-Nissl dye identifies pericytes as distinct vascular mural cells during in vivo brain imaging. Nature Neuroscience, 20(7), 1023–1032. https://doi.org/10.1038/nn.4564
Daneman, R. (2012). The blood-brain barrier in health and disease. Annals of Neurology, 72(5), 648–672. https://doi.org/10.1002/ana.23648
Daneman, R., Zhou, L., Kebede, A. A., & Barres, B. A. (2010). Pericytes are required for blood-brain barrier integrity during embryogenesis. Nature, 468(7323), 562–566. https://doi.org/10.1038/nature09513
Dawson, M. R., Levine, J. M., & Reynolds, R. (2000). NG2-expressing cells in the central nervous system: Are they oligodendroglial progenitors? Journal of Neuroscience Research, 61(5), 471–479. https://doi.org/10.1002/1097-4547(20000901)61:5%3c471::AID-JNR1%3e3.0.CO;2-N
De La Fuente, A. G., Lange, S., Silva, M. E., Gonzalez, G. A., Tempfer, H., van Wijngaarden, P., Zhao, C., Di Canio, L., Trost, A., Bieler, L., Zaunmair, P., Rotheneichner, P., O’Sullivan, A., Couillard-Despres, S., Errea, O., Mäe, M. A., Andrae, J., He, L., Keller, A., et al. (2017). Pericytes stimulate oligodendrocyte progenitor cell differentiation during CNS remyelination. Cell Reports, 20(8), 1755–1764. https://doi.org/10.1016/j.celrep.2017.08.007
Dias, D. O., Kalkitsas, J., Kelahmetoglu, Y., Estrada, C. P., Tatarishvili, J., Holl, D., Jansson, L., Banitalebi, S., Amiry-Moghaddam, M., Ernst, A., Huttner, H. B., Kokaia, Z., Lindvall, O., Brundin, L., Frisén, J., & Göritz, C. (2021). Pericyte-derived fibrotic scarring is conserved across diverse central nervous system lesions. Nature Communications, 12(1), 5501. https://doi.org/10.1038/s41467-021-25585-5
Dias Moura Prazeres, P. H., Sena, I. F. G., da Borges, I. T., de Azevedo, P. O., Andreotti, J. P., de Paiva, A. E., de Almeida, V. M., de Paula Guerra, D. A., Pinheiro Dos Santos, G. S., Mintz, A., Delbono, O., & Birbrair, A. (2017). Pericytes are heterogeneous in their origin within the same tissue. Developmental Biology, 427(1), 6–11. https://doi.org/10.1016/j.ydbio.2017.05.001
Díaz-Flores, L., Gutiérrez, R., Madrid, J. F., Varela, H., Valladares, F., Acosta, E., Martín-Vasallo, P., & Díaz-Flores, L. (2009). Pericytes, Morphofunction, interactions and pathology in a quiescent and activated mesenchymal cell niche. Histology and Histopathology, 24(7), 909–969. https://doi.org/10.14670/HH-24.909
Ding, R., Hase, Y., Ameen-Ali, K. E., Ndung’u, M., Stevenson, W., Barsby, J., Gourlay, R., Akinyemi, T., Akinyemi, R., Uemura, M. T., Polvikoski, T., Mukaetova-Ladinska, E., Ihara, M., & Kalaria, R. N. (2020). Loss of capillary pericytes and the blood-brain barrier in white matter in poststroke and vascular dementias and Alzheimer’s disease. Brain Pathology, 30(6), 1087–1101. https://doi.org/10.1111/bpa.12888
Ding, R., Hase, Y., Burke, M., Foster, V., Stevenson, W., Polvikoski, T., & Kalaria, R. N. (2021). Loss with ageing but preservation of frontal cortical capillary pericytes in post-stroke dementia, vascular dementia and Alzheimer’s disease. Acta Neuropathologica Communications, 9(1), 130. https://doi.org/10.1186/s40478-021-01230-6
Dore-Duffy, P., Katychev, A., Wang, X., & Van Buren, E. (2006). CNS microvascular pericytes exhibit multipotential stem cell activity. Journal of Cerebral Blood Flow and Metabolism, 26(5), 613–624. https://doi.org/10.1038/sj.jcbfm.9600272
Eberth, C. J. (1871). Handbuch der Lehre von der Gewegen des Menschen und der Tiere. W. Engelman.
Ergul, A., Alhusban, A., & Fagan, S. C. (2012). Angiogenesis: A harmonized target for recovery after stroke. Stroke, 43(8), 2270–2274. https://doi.org/10.1161/STROKEAHA.111.642710
Fernández-Klett, F., Offenhauser, N., Dirnagl, U., Priller, J., & Lindauer, U. (2010). Pericytes in capillaries are contractile in vivo, but arterioles mediate functional hyperemia in the mouse brain. Proceedings of the National Academy of Sciences of the United States of America, 107(51), 22290–22295. https://doi.org/10.1073/pnas.1011321108
Fernández-Klett, F., Potas, J. R., Hilpert, D., Blazej, K., Radke, J., Huck, J., Engel, O., Stenzel, W., Genové, G., & Priller, J. (2013). Early loss of pericytes and perivascular stromal cell-induced scar formation after stroke. Journal of Cerebral Blood Flow and Metabolism, 33(3), 428–439. https://doi.org/10.1038/jcbfm.2012.187
Gaengel, K., Genové, G., Armulik, A., & Betsholtz, C. (2009). Endothelial-mural cell signaling in vascular development and angiogenesis. Arteriosclerosis, Thrombosis, and Vascular Biology, 29(5), 630–638. https://doi.org/10.1161/ATVBAHA.107.161521
Gautam, J., Xu, L., Nirwane, A., Nguyen, B., & Yao, Y. (2020). Loss of mural cell-derived laminin aggravates hemorrhagic brain injury. Journal of Neuroinflammation, 17(1), 103. https://doi.org/10.1186/s12974-020-01788-3
Gautam, J., Zhang, X., & Yao, Y. (2016). The role of pericytic laminin in blood brain barrier integrity maintenance. Scientific Reports, 6, 36450. https://doi.org/10.1038/srep36450
GBD 2019 Stroke Collaborators. (2021). Global, regional, and national burden of stroke and its risk factors, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. The Lancet: Neurology, 20(10), 795–820. https://doi.org/10.1016/S1474-4422(21)00252-0
Ghori, A., Prinz, V., Nieminen-Kehlä, M., Bayerl, S. H., Kremenetskaia, I., Riecke, J., Krechel, H., Broggini, T., Scherschinski, L., Licht, T., Keshet, E., & Vajkoczy, P. (2022). Vascular endothelial growth factor augments the tolerance towards cerebral stroke by enhancing neurovascular repair mechanism. Translational Stroke Research, 13(5), 774–791. https://doi.org/10.1007/s12975-022-00991-z
Göritz, C., Dias, D. O., Tomilin, N., Barbacid, M., Shupliakov, O., & Frisén, J. (2011). A pericyte origin of spinal cord scar tissue. Science (NY), 333(6039), 238–242. https://doi.org/10.1126/science.1203165
Goyal, M., Menon, B. K., van Zwam, W. H., Dippel, D. W. J., Mitchell, P. J., Demchuk, A. M., Dávalos, A., Majoie, C. B. L. M., van der Lugt, A., de Miquel, M. A., Donnan, G. A., Roos, Y. B. W. E. M., Bonafe, A., Jahan, R., Diener, H.-C., van den Berg, L. A., Levy, E. I., Berkhemer, O. A., Pereira, V. M., et al. (2016). Endovascular thrombectomy after large-vessel ischaemic stroke: A meta-analysis of individual patient data from five randomised trials. Lancet (London, England), 387(10029), 1723–1731. https://doi.org/10.1016/S0140-6736(16)00163-X
Grant, R. I., Hartmann, D. A., Underly, R. G., Berthiaume, A.-A., Bhat, N. R., & Shih, A. Y. (2019). Organizational hierarchy and structural diversity of microvascular pericytes in adult mouse cortex. Journal of Cerebral Blood Flow and Metabolism, 39(3), 411–425. https://doi.org/10.1177/0271678X17732229
Guijarro-Muñoz, I., Compte, M., Álvarez-Cienfuegos, A., Álvarez-Vallina, L., & Sanz, L. (2014). Lipopolysaccharide activates Toll-like receptor 4 (TLR4)-mediated NF-κB signaling pathway and proinflammatory response in human pericytes. The Journal of Biological Chemistry, 289(4), 2457–2468. https://doi.org/10.1074/jbc.M113.521161
Guimarães-Camboa, N., Cattaneo, P., Sun, Y., Moore-Morris, T., Gu, Y., Dalton, N. D., Rockenstein, E., Masliah, E., Peterson, K. L., Stallcup, W. B., Chen, J., & Evans, S. M. (2017). Pericytes of multiple organs do not behave as mesenchymal stem cells in vivo. Cell Stem Cell, 20(3), 345-359.e5. https://doi.org/10.1016/j.stem.2016.12.006
Gundersen, G. A., Vindedal, G. F., Skare, O., & Nagelhus, E. A. (2014). Evidence that pericytes regulate aquaporin-4 polarization in mouse cortical astrocytes. Brain Structure & Function, 219(6), 2181–2186. https://doi.org/10.1007/s00429-013-0629-0
Gurnik, S., Devraj, K., Macas, J., Yamaji, M., Starke, J., Scholz, A., Sommer, K., Di Tacchio, M., Vutukuri, R., Beck, H., Mittelbronn, M., Foerch, C., Pfeilschifter, W., Liebner, S., Peters, K. G., Plate, K. H., & Reiss, Y. (2016). Angiopoietin-2-induced blood-brain barrier compromise and increased stroke size are rescued by VE-PTP-dependent restoration of Tie2 signaling. Acta Neuropathologica, 131(5), 753–773. https://doi.org/10.1007/s00401-016-1551-3
Hacke, W., Kaste, M., Bluhmki, E., Brozman, M., Dávalos, A., Guidetti, D., Larrue, V., Lees, K. R., Medeghri, Z., Machnig, T., Schneider, D., von Kummer, R., Wahlgren, N., Toni, D., ECASS Investigators. (2008). Thrombolysis with alteplase 3 to 4.5 hours after acute ischemic stroke. The New England Journal of Medicine, 359(13), 1317–1329. https://doi.org/10.1056/NEJMoa0804656
Hall, C. N., Reynell, C., Gesslein, B., Hamilton, N. B., Mishra, A., Sutherland, B. A., O’Farrell, F. M., Buchan, A. M., Lauritzen, M., & Attwell, D. (2014). Capillary pericytes regulate cerebral blood flow in health and disease. Nature, 508(7494), 55–60. https://doi.org/10.1038/nature13165
Harder, D. R., Zhang, C., & Gebremedhin, D. (2002). Astrocytes function in matching blood flow to metabolic activity. News in Physiological Sciences, 17, 27–31. https://doi.org/10.1152/physiologyonline.2002.17.1.27
Hartmann, D. A., Berthiaume, A.-A., Grant, R. I., Harrill, S. A., Koski, T., Tieu, T., McDowell, K. P., Faino, A. V., Kelly, A. L., & Shih, A. Y. (2021). Brain capillary pericytes exert a substantial but slow influence on blood flow. Nature Neuroscience, 24(5), 633–645. https://doi.org/10.1038/s41593-020-00793-2
Hartmann, D. A., Underly, R. G., Grant, R. I., Watson, A. N., Lindner, V., & Shih, A. Y. (2015). Pericyte structure and distribution in the cerebral cortex revealed by high-resolution imaging of transgenic mice. Neurophotonics, 2(4), 041402. https://doi.org/10.1117/1.NPh.2.4.041402
Herpich, F., & Rincon, F. (2020). Management of acute ischemic stroke. Critical Care Medicine, 48(11), 1654–1663. https://doi.org/10.1097/CCM.0000000000004597
Hill, M. D., Goyal, M., Menon, B. K., Nogueira, R. G., McTaggart, R. A., Demchuk, A. M., Poppe, A. Y., Buck, B. H., Field, T. S., Dowlatshahi, D., van Adel, B. A., Swartz, R. H., Shah, R. A., Sauvageau, E., Zerna, C., Ospel, J. M., Joshi, M., Almekhlafi, M. A., Ryckborst, K. J., et al. (2020). Efficacy and safety of nerinetide for the treatment of acute ischaemic stroke (ESCAPE-NA1): A multicentre, double-blind, randomised controlled trial. Lancet (London), 395(10227), 878–887. https://doi.org/10.1016/S0140-6736(20)30258-0
Hill, R. A., Tong, L., Yuan, P., Murikinati, S., Gupta, S., & Grutzendler, J. (2015). Regional blood flow in the normal and ischemic brain is controlled by arteriolar smooth muscle cell contractility and not by capillary pericytes. Neuron, 87(1), 95–110. https://doi.org/10.1016/j.neuron.2015.06.001
Iadecola, C. (2017). The neurovascular unit coming of age: a journey through neurovascular coupling in health and disease. Neuron, 96(1), 17–42. https://doi.org/10.1016/j.neuron.2017.07.030
Joyce, N. C., DeCamilli, P., & Boyles, J. (1984). Pericytes, like vascular smooth muscle cells, are immunocytochemically positive for cyclic GMP-dependent protein kinase. Microvascular Research, 28(2), 206–219. https://doi.org/10.1016/0026-2862(84)90018-9
Jung, K.-H., Chu, K., Lee, S.-T., Bahn, J.-J., Jeon, D., Kim, J.-H., Kim, S., Won, C.-H., Kim, M., Lee, S. K., & Roh, J.-K. (2011). Multipotent PDGFRβ-expressing cells in the circulation of stroke patients. Neurobiology of Disease, 41(2), 489–497. https://doi.org/10.1016/j.nbd.2010.10.020
Kang, M., & Yao, Y. (2020). Basement membrane changes in ischemic stroke. Stroke, 51(4), 1344–1352. https://doi.org/10.1161/STROKEAHA.120.028928
Karow, M., Camp, J. G., Falk, S., Gerber, T., Pataskar, A., Gac-Santel, M., Kageyama, J., Brazovskaja, A., Garding, A., Fan, W., Riedemann, T., Casamassa, A., Smiyakin, A., Schichor, C., Götz, M., Tiwari, V. K., Treutlein, B., & Berninger, B. (2018). Direct pericyte-to-neuron reprogramming via unfolding of a neural stem cell-like program. Nature Neuroscience, 21(7), 932–940. https://doi.org/10.1038/s41593-018-0168-3
Korte, N., Ilkan, Z., Pearson, C. L., Pfeiffer, T., Singhal, P., Rock, J. R., Sethi, H., Gill, D., Attwell, D., & Tammaro, P. (2022). The Ca2+-gated channel TMEM16A amplifies capillary pericyte contraction and reduces cerebral blood flow after ischemia. The Journal of Clinical Investigation, 132(9), e154118. https://doi.org/10.1172/JCI154118
Levine, J. M., Reynolds, R., & Fawcett, J. W. (2001). The oligodendrocyte precursor cell in health and disease. Trends in Neurosciences, 24(1), 39–47. https://doi.org/10.1016/s0166-2236(00)01691-x
Li, F., Lan, Y., Wang, Y., Wang, J., Yang, G., Meng, F., Han, H., Meng, A., Wang, Y., & Yang, X. (2011). Endothelial Smad4 maintains cerebrovascular integrity by activating N-cadherin through cooperation with Notch. Developmental Cell, 20(3), 291–302. https://doi.org/10.1016/j.devcel.2011.01.011
Liu, Z., & Zhang, Z. (2022). Mapping cell types across human tissues. Science (NY), 376(6594), 695–696. https://doi.org/10.1126/science.abq2116
Lo, E. H., Dalkara, T., & Moskowitz, M. A. (2003). Mechanisms, challenges and opportunities in stroke. Nature Reviews. Neuroscience, 4(5), 399–415. https://doi.org/10.1038/nrn1106
Lo, E. H., & Rosenberg, G. A. (2009). The neurovascular unit in health and disease. Stroke, 40, S2–S3. https://doi.org/10.1161/STROKEAHA.108.534404
Lyden, P. D. (2021). Cerebroprotection for acute ischemic stroke: Looking ahead. Stroke, 52(9), 3033–3044. https://doi.org/10.1161/STROKEAHA.121.032241
Ma, H., Campbell, B. C. V., Parsons, M. W., Churilov, L., Levi, C. R., Hsu, C., Kleinig, T. J., Wijeratne, T., Curtze, S., Dewey, H. M., Miteff, F., Tsai, C.-H., Lee, J.-T., Phan, T. G., Mahant, N., Sun, M.-C., Krause, M., Sturm, J., Grimley, R., et al. (2019). Thrombolysis guided by perfusion imaging up to 9 hours after onset of stroke. The New England Journal of Medicine, 380(19), 1795–1803. https://doi.org/10.1056/NEJMoa1813046
Ma, Q., Li, R., Wang, L., Yin, P., Wang, Y., Yan, C., Ren, Y., Qian, Z., Vaughn, M. G., McMillin, S. E., Hay, S. I., Naghavi, M., Cai, M., Wang, C., Zhang, Z., Zhou, M., Lin, H., & Yang, Y. (2021). Temporal trend and attributable risk factors of stroke burden in China, 1990–2019: An analysis for the Global Burden of Disease Study 2019. The Lancet: Public Health, 6(12), e897–e906. https://doi.org/10.1016/S2468-2667(21)00228-0
Ma, Y., Wang, J., Wang, Y., & Yang, G.-Y. (2017). The biphasic function of microglia in ischemic stroke. Progress in Neurobiology, 157, 247–272. https://doi.org/10.1016/j.pneurobio.2016.01.005
Mäe, M. A., He, L., Nordling, S., Vazquez-Liebanas, E., Nahar, K., Jung, B., Li, X., Tan, B. C., Chin Foo, J., Cazenave-Gassiot, A., Wenk, M. R., Zarb, Y., Lavina, B., Quaggin, S. E., Jeansson, M., Gu, C., Silver, D. L., Vanlandewijck, M., Butcher, E. C., et al. (2021). Single-cell analysis of blood-brain barrier response to pericyte loss. Circulation Research, 128(4), e46–e62. https://doi.org/10.1161/CIRCRESAHA.120.317473
Maki, T., Choi, Y. K., Miyamoto, N., Shindo, A., Liang, A. C., Ahn, B. J., Mandeville, E. T., Kaji, S., Itoh, K., Seo, J. H., Gelman, I. H., Lok, J., Takahashi, R., Kim, K.-W., Lo, E. H., & Arai, K. (2018). A-kinase anchor protein 12 is required for oligodendrocyte differentiation in adult white matter. Stem Cells, 36(5), 751–760. https://doi.org/10.1002/stem.2771
Maki, T., Maeda, M., Uemura, M., Lo, E. K., Terasaki, Y., Liang, A. C., Shindo, A., Choi, Y. K., Taguchi, A., Matsuyama, T., Takahashi, R., Ihara, M., & Arai, K. (2015). Potential interactions between pericytes and oligodendrocyte precursor cells in perivascular regions of cerebral white matter. Neuroscience Letters, 597, 164–169. https://doi.org/10.1016/j.neulet.2015.04.047
Mastrullo, V., Cathery, W., Velliou, E., Madeddu, P., & Campagnolo, P. (2020). Angiogenesis in tissue engineering: As nature intended? Frontiers in Bioengineering and Biotechnology, 8, 188. https://doi.org/10.3389/fbioe.2020.00188
Mathiisen, T. M., Lehre, K. P., Danbolt, N. C., & Ottersen, O. P. (2010). The perivascular astroglial sheath provides a complete covering of the brain microvessels: An electron microscopic 3D reconstruction. Glia, 58(9), 1094–1103. https://doi.org/10.1002/glia.20990
Matute, C., Domercq, M., Pérez-Samartín, A., & Ransom, B. R. (2013). Protecting white matter from stroke injury. Stroke, 44(4), 1204–1211. https://doi.org/10.1161/STROKEAHA.112.658328
Menn, B., Garcia-Verdugo, J. M., Yaschine, C., Gonzalez-Perez, O., Rowitch, D., & Alvarez-Buylla, A. (2006). Origin of oligodendrocytes in the subventricular zone of the adult brain. The Journal of Neuroscience, 26(30), 7907–7918. https://doi.org/10.1523/JNEUROSCI.1299-06.2006
Miyamoto, N., Maki, T., Shindo, A., Liang, A. C., Maeda, M., Egawa, N., Itoh, K., Lo, E. K., Lok, J., Ihara, M., & Arai, K. (2015). Astrocytes promote oligodendrogenesis after white matter damage via brain-derived neurotrophic factor. The Journal of Neuroscience, 35(41), 14002–14008. https://doi.org/10.1523/JNEUROSCI.1592-15.2015
Montagne, A., Nikolakopoulou, A. M., Zhao, Z., Sagare, A. P., Si, G., Lazic, D., Barnes, S. R., Daianu, M., Ramanathan, A., Go, A., Lawson, E. J., Wang, Y., Mack, W. J., Thompson, P. M., Schneider, J. A., Varkey, J., Langen, R., Mullins, E., Jacobs, R. E., & Zlokovic, B. V. (2018). Pericyte degeneration causes white matter dysfunction in the mouse central nervous system. Nature Medicine, 24(3), 326–337. https://doi.org/10.1038/nm.4482
Nakagomi, T., Kubo, S., Nakano-Doi, A., Sakuma, R., Lu, S., Narita, A., Kawahara, M., Taguchi, A., & Matsuyama, T. (2015). Brain vascular pericytes following ischemia have multipotential stem cell activity to differentiate into neural and vascular lineage cells. Stem Cells, 33(6), 1962–1974. https://doi.org/10.1002/stem.1977
Nakagomi, T., Molnár, Z., Nakano-Doi, A., Taguchi, A., Saino, O., Kubo, S., Clausen, M., Yoshikawa, H., Nakagomi, N., & Matsuyama, T. (2011). Ischemia-induced neural stem/progenitor cells in the pia mater following cortical infarction. Stem Cells and Development, 20(12), 2037–2051. https://doi.org/10.1089/scd.2011.0279
Nakamura, K., Ikeuchi, T., Nara, K., Rhodes, C. S., Zhang, P., Chiba, Y., Kazuno, S., Miura, Y., Ago, T., Arikawa-Hirasawa, E., Mukouyama, Y.-S., & Yamada, Y. (2019). Perlecan regulates pericyte dynamics in the maintenance and repair of the blood-brain barrier. The Journal of Cell Biology, 218(10), 3506–3525. https://doi.org/10.1083/jcb.201807178
National Institute of Neurological Disorders and Stroke rt-PA Stroke Study Group. (1995). Tissue plasminogen activator for acute ischemic stroke. The New England Journal of Medicine, 333(24), 1581–1587. https://doi.org/10.1056/NEJM199512143332401
Nirwane, A., Johnson, J., Nguyen, B., Miner, J. H., & Yao, Y. (2019). Mural cell-derived laminin-α5 plays a detrimental role in ischemic stroke. Acta Neuropathologica Communications, 7(1), 23. https://doi.org/10.1186/s40478-019-0676-8
Nirwane, A., & Yao, Y. (2022). SMAlow/undetectable pericytes differentiate into microglia- and macrophage-like cells in ischemic brain. Cellular and Molecular Life Sciences, 79(5), 264. https://doi.org/10.1007/s00018-022-04322-1
Nogueira, R. G., Jadhav, A. P., Haussen, D. C., Bonafe, A., Budzik, R. F., Bhuva, P., Yavagal, D. R., Ribo, M., Cognard, C., Hanel, R. A., Sila, C. A., Hassan, A. E., Millan, M., Levy, E. I., Mitchell, P., Chen, M., English, J. D., Shah, Q. A., Silver, F. L., et al. (2018). Thrombectomy 6 to 24 hours after stroke with a mismatch between deficit and infarct. The New England Journal of Medicine, 378(1), 11–21. https://doi.org/10.1056/NEJMoa1706442
Özen, I., Deierborg, T., Miharada, K., Padel, T., Englund, E., Genové, G., & Paul, G. (2014). Brain pericytes acquire a microglial phenotype after stroke. Acta Neuropathologica, 128(3), 381–396. https://doi.org/10.1007/s00401-014-1295-x
Özen, I., Roth, M., Barbariga, M., Gaceb, A., Deierborg, T., Genové, G., & Paul, G. (2018). Loss of regulator of G-protein signaling 5 leads to neurovascular protection in stroke. Stroke, 49(9), 2182–2190. https://doi.org/10.1161/STROKEAHA.118.020124
Park, T.I.-H., Feisst, V., Brooks, A. E. S., Rustenhoven, J., Monzo, H. J., Feng, S. X., Mee, E. W., Bergin, P. S., Oldfield, R., Graham, E. S., Curtis, M. A., Faull, R. L. M., Dunbar, P. R., & Dragunow, M. (2016). Cultured pericytes from human brain show phenotypic and functional differences associated with differential CD90 expression. Scientific Reports, 6, 26587. https://doi.org/10.1038/srep26587
Paul, S., & Candelario-Jalil, E. (2021). Emerging neuroprotective strategies for the treatment of ischemic stroke: An overview of clinical and preclinical studies. Experimental Neurology, 335, 113518. https://doi.org/10.1016/j.expneurol.2020.113518
Peppiatt, C. M., Howarth, C., Mobbs, P., & Attwell, D. (2006). Bidirectional control of CNS capillary diameter by pericytes. Nature, 443(7112), 700–704. https://doi.org/10.1038/nature05193
Pham, T. T. D., Park, S., Kolluri, K., Kawaguchi, R., Wang, L., Tran, D., Zhao, P., Carmichael, S. T., & Ardehali, R. (2021). Heart and brain pericytes exhibit a pro-fibrotic response after vascular injury. Circulation Research, 129(7), e141–e143. https://doi.org/10.1161/CIRCRESAHA.121.319288
Pisani, F., Castagnola, V., Simone, L., Loiacono, F., Svelto, M., & Benfenati, F. (2022). Role of pericytes in blood-brain barrier preservation during ischemia through tunneling nanotubes. Cell Death & Disease, 13(7), 582. https://doi.org/10.1038/s41419-022-05025-y
Qiu, Y.-M., Zhang, C.-L., Chen, A.-Q., Wang, H.-L., Zhou, Y.-F., Li, Y.-N., & Hu, B. (2021). Immune cells in the bbb disruption after acute ischemic stroke: Targets for immune therapy? Frontiers in Immunology, 12, 678744. https://doi.org/10.3389/fimmu.2021.678744
Roth, M., Enström, A., Aghabeick, C., Carlsson, R., Genové, G., & Paul, G. (2020). Parenchymal pericytes are not the major contributor of extracellular matrix in the fibrotic scar after stroke in male mice. Journal of Neuroscience Research, 98(5), 826–842. https://doi.org/10.1002/jnr.24557
Rouget, C. (1873). Memoire sur le developpement, la structure et les proprietes physiologiques des capillaries sanguins et lymphatiques. Arch Physiol Normale Pathol, 5, 603–663.
Rustenhoven, J., Aalderink, M., Scotter, E. L., Oldfield, R. L., Bergin, P. S., Mee, E. W., Graham, E. S., Faull, R. L. M., Curtis, M. A., Park, T.I.-H., & Dragunow, M. (2016). TGF-beta1 regulates human brain pericyte inflammatory processes involved in neurovasculature function. Journal of Neuroinflammation, 13, 37. https://doi.org/10.1186/s12974-016-0503-0
Rustenhoven, J., Jansson, D., Smyth, L. C., & Dragunow, M. (2017). Brain pericytes as mediators of neuroinflammation. Trends in Pharmacological Sciences, 38(3), 291–304. https://doi.org/10.1016/j.tips.2016.12.001
Rustom, A., Saffrich, R., Markovic, I., Walther, P., & Gerdes, H.-H. (2004). Nanotubular highways for intercellular organelle transport. Science (NY), 303(5660), 1007–1010. https://doi.org/10.1126/science.1093133
Sakuma, R., Kawahara, M., Nakano-Doi, A., Takahashi, A., Tanaka, Y., Narita, A., Kuwahara-Otani, S., Hayakawa, T., Yagi, H., Matsuyama, T., & Nakagomi, T. (2016). Brain pericytes serve as microglia-generating multipotent vascular stem cells following ischemic stroke. Journal of Neuroinflammation, 13(1), 57. https://doi.org/10.1186/s12974-016-0523-9
Sakuma, R., Kobayashi, M., Kobashi, R., Onishi, M., Maeda, M., Kataoka, Y., & Imaoka, S. (2022). Brain pericytes acquire stemness via the Nrf2-dependent antioxidant system. Stem Cells (dayton, Ohio), 40(7), 641–654. https://doi.org/10.1093/stmcls/sxac024
Savitz, S. I., Baron, J.-C., Fisher, M., STAIR X Consortium. (2019). Stroke Treatment Academic Industry Roundtable X: Brain Cytoprotection Therapies in the Reperfusion Era. Stroke, 50(4), 1026–1031. https://doi.org/10.1161/STROKEAHA.118.023927
Shepro, D., & Morel, N. M. (1993). Pericyte physiology. FASEB Journal: Official Publication of the Federation of American Societies for Experimental Biology, 7(11), 1031–1038. https://doi.org/10.1096/fasebj.7.11.8370472
Shi, L., Sun, Z., Su, W., Xu, F., Xie, D., Zhang, Q., Dai, X., Iyer, K., Hitchens, T. K., Foley, L. M., Li, S., Stolz, D. B., Chen, K., Ding, Y., Thomson, A. W., Leak, R. K., Chen, J., & Hu, X. (2021). Treg cell-derived osteopontin promotes microglia-mediated white matter repair after ischemic stroke. Immunity, 54(7), 1527-1542.e8. https://doi.org/10.1016/j.immuni.2021.04.022
Shibahara, T., Ago, T., Nakamura, K., Tachibana, M., Yoshikawa, Y., Komori, M., Yamanaka, K., Wakisaka, Y., & Kitazono, T. (2020a). Pericyte-mediated tissue repair through PDGFRβ promotes peri-infarct astrogliosis, oligodendrogenesis, and functional recovery after acute ischemic stroke. ENeuro. https://doi.org/10.1523/ENEURO.0474-19.2020
Shibahara, T., Ago, T., Tachibana, M., Nakamura, K., Yamanaka, K., Kuroda, J., Wakisaka, Y., & Kitazono, T. (2020b). Reciprocal interaction between pericytes and macrophage in poststroke tissue repair and functional recovery. Stroke, 51(10), 3095–3106. https://doi.org/10.1161/STROKEAHA.120.029827
Sims, D. E. (1991). Recent advances in pericyte biology—Implications for health and disease. The Canadian Journal of Cardiology, 7(10), 431–443.
Stark, K., Eckart, A., Haidari, S., Tirniceriu, A., Lorenz, M., von Brühl, M.-L., Gärtner, F., Khandoga, A. G., Legate, K. R., Pless, R., Hepper, I., Lauber, K., Walzog, B., & Massberg, S. (2013). Capillary and arteriolar pericytes attract innate leukocytes exiting through venules and “instruct” them with pattern-recognition and motility programs. Nature Immunology, 14(1), 41–51. https://doi.org/10.1038/ni.2477
Stratman, A. N., Malotte, K. M., Mahan, R. D., Davis, M. J., & Davis, G. E. (2009). Pericyte recruitment during vasculogenic tube assembly stimulates endothelial basement membrane matrix formation. Blood, 114(24), 5091–5101. https://doi.org/10.1182/blood-2009-05-222364
Suzuki, K., Kimura, K., Takeuchi, M., Morimoto, M., Kanazawa, R., Kamiya, Y., Shigeta, K., Ishii, N., Takayama, Y., Koguchi, Y., Takigawa, T., Hayakawa, M., Ota, T., Okubo, S., Naito, H., Akaji, K., Kato, N., Inoue, M., Hirano, T., et al. (2019). The randomized study of endovascular therapy with versus without intravenous tissue plasminogen activator in acute stroke with ICA and M1 occlusion (SKIP study). International Journal of Stroke: Official Journal of the International Stroke Society, 14(7), 752–755. https://doi.org/10.1177/1747493019840932
Sweeney, M. D., Ayyadurai, S., & Zlokovic, B. V. (2016). Pericytes of the neurovascular unit: Key functions and signaling pathways. Nature Neuroscience, 19(6), 771–783. https://doi.org/10.1038/nn.4288
Taheri, S., Gasparovic, C., Huisa, B. N., Adair, J. C., Edmonds, E., Prestopnik, J., Grossetete, M., Shah, N. J., Wills, J., Qualls, C., & Rosenberg, G. A. (2011). Blood-brain barrier permeability abnormalities in vascular cognitive impairment. Stroke, 42(8), 2158–2163. https://doi.org/10.1161/STROKEAHA.110.611731
Takahashi, A., Park, H. K., Melgar, M. A., Alcocer, L., Pinto, J., Lenzi, T., Diaz, F. G., & Rafols, J. A. (1997). Cerebral cortex blood flow and vascular smooth muscle contractility in a rat model of ischemia: A correlative laser Doppler flowmetric and scanning electron microscopic study. Acta Neuropathologica, 93(4), 354–368. https://doi.org/10.1007/s004010050627
Tatebayashi, K., Tanaka, Y., Nakano-Doi, A., Sakuma, R., Kamachi, S., Shirakawa, M., Uchida, K., Kageyama, H., Takagi, T., Yoshimura, S., Matsuyama, T., & Nakagomi, T. (2017). Identification of multipotent stem cells in human brain tissue following stroke. Stem Cells and Development, 26(11), 787–797. https://doi.org/10.1089/scd.2016.0334
Thomalla, G., Simonsen, C. Z., Boutitie, F., Andersen, G., Berthezene, Y., Cheng, B., Cheripelli, B., Cho, T.-H., Fazekas, F., Fiehler, J., Ford, I., Galinovic, I., Gellissen, S., Golsari, A., Gregori, J., Günther, M., Guibernau, J., Häusler, K. G., Hennerici, M., et al. (2018). MRI-guided thrombolysis for stroke with unknown time of onset. The New England Journal of Medicine, 379(7), 611–622. https://doi.org/10.1056/NEJMoa1804355
Török, O., Schreiner, B., Schaffenrath, J., Tsai, H.-C., Maheshwari, U., Stifter, S. A., Welsh, C., Amorim, A., Sridhar, S., Utz, S. G., Mildenberger, W., Nassiri, S., Delorenzi, M., Aguzzi, A., Han, M. H., Greter, M., Becher, B., & Keller, A. (2021). Pericytes regulate vascular immune homeostasis in the CNS. Proceedings of the National Academy of Sciences of the United States of America, 118(10), e2016587118. https://doi.org/10.1073/pnas.2016587118
Tsao, C.-C., Baumann, J., Huang, S.-F., Kindler, D., Schroeter, A., Kachappilly, N., Gassmann, M., Rudin, M., & Ogunshola, O. O. (2021). Pericyte hypoxia-inducible factor-1 (HIF-1) drives blood-brain barrier disruption and impacts acute ischemic stroke outcome. Angiogenesis, 24(4), 823–842. https://doi.org/10.1007/s10456-021-09796-4
Vanlandewijck, M., He, L., Mäe, M. A., Andrae, J., Ando, K., Del Gaudio, F., Nahar, K., Lebouvier, T., Laviña, B., Gouveia, L., Sun, Y., Raschperger, E., Räsänen, M., Zarb, Y., Mochizuki, N., Keller, A., Lendahl, U., & Betsholtz, C. (2018). A molecular atlas of cell types and zonation in the brain vasculature. Nature, 554(7693), 475–480. https://doi.org/10.1038/nature25739
Winkler, E. A., Bell, R. D., & Zlokovic, B. V. (2010). Pericyte-specific expression of PDGF beta receptor in mouse models with normal and deficient PDGF beta receptor signaling. Molecular Neurodegeneration, 5, 32. https://doi.org/10.1186/1750-1326-5-32
Winkler, E. A., Bell, R. D., & Zlokovic, B. V. (2011). Central nervous system pericytes in health and disease. Nature Neuroscience, 14(11), 1398–1405. https://doi.org/10.1038/nn.2946
Xu, J., Wang, A., Meng, X., Yalkun, G., Xu, A., Gao, Z., Chen, H., Ji, Y., Xu, J., Geng, D., Zhu, R., Liu, B., Dong, A., Mu, H., Lu, Z., Li, S., Zheng, H., Chen, X., Wang, Y., et al. (2021). Edaravone dexborneol versus edaravone alone for the treatment of acute ischemic stroke: A phase III, randomized, double-blind. Comparative Trial. Stroke, 52(3), 772–780. https://doi.org/10.1161/STROKEAHA.120.031197
Xu, L., Nirwane, A., & Yao, Y. (2019). Basement membrane and blood-brain barrier. Stroke and Vascular Neurology, 4(2), 78–82. https://doi.org/10.1136/svn-2018-000198
Yang, P., Zhang, Y., Zhang, L., Zhang, Y., Treurniet, K. M., Chen, W., Peng, Y., Han, H., Wang, J., Wang, S., Yin, C., Liu, S., Wang, P., Fang, Q., Shi, H., Yang, J., Wen, C., Li, C., Jiang, C., et al. (2020). Endovascular thrombectomy with or without intravenous alteplase in acute stroke. The New England Journal of Medicine, 382(21), 1981–1993. https://doi.org/10.1056/NEJMoa2001123
Yang, S., Jin, H., Zhu, Y., Wan, Y., Opoku, E. N., Zhu, L., & Hu, B. (2017). Diverse functions and mechanisms of pericytes in ischemic stroke. Current Neuropharmacology, 15(6), 892–905. https://doi.org/10.2174/1570159X15666170112170226
Yang, Y., & Torbey, M. T. (2020). Angiogenesis and blood-brain barrier permeability in vascular remodeling after stroke. Current Neuropharmacology, 18(12), 1250–1265. https://doi.org/10.2174/1570159X18666200720173316
Yao, Y., Chen, Z.-L., Norris, E. H., & Strickland, S. (2014). Astrocytic laminin regulates pericyte differentiation and maintains blood brain barrier integrity. Nature Communications, 5, 3413. https://doi.org/10.1038/ncomms4413
Yemisci, M., Gursoy-Ozdemir, Y., Vural, A., Can, A., Topalkara, K., & Dalkara, T. (2009). Pericyte contraction induced by oxidative-nitrative stress impairs capillary reflow despite successful opening of an occluded cerebral artery. Nature Medicine, 15(9), 1031–1037. https://doi.org/10.1038/nm.2022
Yoshida, Y., Kabara, M., Kano, K., Horiuchi, K., Hayasaka, T., Tomita, Y., Takehara, N., Minoshima, A., Aonuma, T., Maruyama, K., Nakagawa, N., Azuma, N., Hasebe, N., & Kawabe, J.-I. (2020). Capillary-resident EphA7+ pericytes are multipotent cells with anti-ischemic effects through capillary formation. Stem Cells Translational Medicine, 9(1), 120–130. https://doi.org/10.1002/sctm.19-0148
Yurchenco, P. D. (2011). Basement membranes: Cell scaffoldings and signaling platforms. Cold Spring Harbor Perspectives in Biology, 3(2), a004911. https://doi.org/10.1101/cshperspect.a004911
Zechariah, A., ElAli, A., Doeppner, T. R., Jin, F., Hasan, M. R., Helfrich, I., Mies, G., & Hermann, D. M. (2013). Vascular endothelial growth factor promotes pericyte coverage of brain capillaries, improves cerebral blood flow during subsequent focal cerebral ischemia, and preserves the metabolic penumbra. Stroke, 44(6), 1690–1697. https://doi.org/10.1161/STROKEAHA.111.000240
Zhang, Y., Liu, L., Zhao, X., Yan, S., Zeng, F., & Zhou, D. (2022). New insight into ischemic stroke: Circadian rhythm in post-stroke angiogenesis. Frontiers in Pharmacology, 13, 927506. https://doi.org/10.3389/fphar.2022.927506
Zheng, Z., Chopp, M., & Chen, J. (2020). Multifaceted roles of pericytes in central nervous system homeostasis and disease. Journal of Cerebral Blood Flow and Metabolism, 40(7), 1381–1401. https://doi.org/10.1177/0271678X20911331
Zhou, Y.-F., Li, P.-C., Wu, J.-H., Haslam, J. A., Mao, L., Xia, Y.-P., He, Q.-W., Wang, X.-X., Lei, H., Lan, X.-L., Miao, Q. R., Yue, Z.-Y., Li, Y.-N., & Hu, B. (2018a). Sema3E/PlexinD1 inhibition is a therapeutic strategy for improving cerebral perfusion and restoring functional loss after stroke in aged rats. Neurobiology of Aging, 70, 102–116. https://doi.org/10.1016/j.neurobiolaging.2018.06.003
Zhou, Y.-F., Li, Y.-N., Jin, H.-J., Wu, J.-H., He, Q.-W., Wang, X.-X., Lei, H., & Hu, B. (2018b). Sema4D/PlexinB1 inhibition ameliorates blood-brain barrier damage and improves outcome after stroke in rats. FASEB Journal: Official Publication of the Federation of American Societies for Experimental Biology, 32(4), 2181–2196. https://doi.org/10.1096/fj.201700786RR
Zimmermann, K. W. (1923). Der Feinere Bau Der Blutcapillaren., 68(1), 29–109. https://doi.org/10.1007/BF02593544
Zlokovic, B. V. (2008). The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron, 57(2), 178–201. https://doi.org/10.1016/j.neuron.2008.01.003
Zlokovic, B. V. (2011). Neurovascular pathways to neurodegeneration in Alzheimer’s disease and other disorders. Nature Reviews. Neuroscience, 12(12), 723–738. https://doi.org/10.1038/nrn3114
Funding
This study is supported by grants from Zhejiang Provincial Medical and Health Technology Project for Young Backbone Talents (Grant 2019RC234).
Author information
Authors and Affiliations
Contributions
Conceptualization: SH and BY; writing—original draft preparation: SH; writing—review and editing: HZ, BY, SS, XH, HS, XF; supervision: HZ All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interest
The authors have no competing interests to declare that are relevant to the content of this article.
Ethical Approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Hu, S., Yang, B., Shu, S. et al. Targeting Pericytes for Functional Recovery in Ischemic Stroke. Neuromol Med 25, 457–470 (2023). https://doi.org/10.1007/s12017-023-08748-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12017-023-08748-z