
Abstract
Osteoclasts are multinucleated cells derived from mononuclear phagocyte precursors (monocytes, macrophages); in the canonical pathway of osteoclastogenesis, these cells fuse and differentiate to form specialised bone-resorbing osteoclasts in the presence of receptor activator for nuclear factor kappa B ligand (RANKL). Non-canonical pathways of osteoclastogenesis have been described in which several cytokines and growth factors are able to substitute for RANKL. These humoral factors can generally be divided into those which, like RANKL, are tumour necrosis family (TNF) superfamily members and those which are not; the former include TNFα lymphotoxin exhibiting inducible expression and competing with herpes simplex virus glycoprotein D for herpesvirus entry mediator, a receptor expressed by T lymphocytes (LIGHT), a proliferation inducing ligand (APRIL) and B cell activating factor (BAFF); the latter include transforming growth factor beta (TGF-β), interleukin-6 (IL-6), IL-8, IL-11, nerve growth factor (NGF), insulin-like growth factor-I (IGF-I) and IGF-II. This review summarises the evidence for these RANKL substitutes in inducing osteoclast differentiation from tissue-derived and circulating mononuclear phagocytes. It also assesses the role these factors are likely to play in promoting the pathological bone resorption seen in many inflammatory and neoplastic lesions of bone and joint including rheumatoid arthritis, aseptic implant loosening and primary and secondary tumours of bone.
Similar content being viewed by others
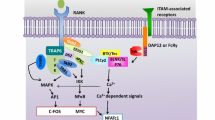
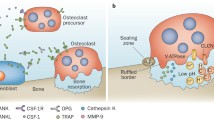

Avoid common mistakes on your manuscript.
Introduction
Bone resorption is required for skeletal modelling during bone growth and for bone remodelling throughout life. Bone remodelling involves co-ordinated bone resorption and bone formation which is carried out by osteoclasts and osteoblasts, respectively. Osteoclasts initiate the remodelling process, removing bone which is subsequently replaced by osteoblasts. Osteoclasts are multinucleated cells that are uniquely specialised to carry out bone resorption. Pathological bone resorption occurs when the resorption component of the remodelling process does not accord with physiological demands and is an invariable accompaniment of a wide range of neoplastic and non-neoplastic musculoskeletal conditions [1, 2]. The extent of pathological bone resorption (osteolysis) is a function of the number of osteoclasts and hence the humoral factors that play a role in osteoclast differentiation.
Osteoclasts are specialised multinucleated cells that form by fusion of mononuclear phagocyte osteoclast precursors [3, 4]. Mononuclear osteoclast precursors are derived from the pluripotential haematopoietic stem cell [5]. These cells circulate in the (CD14+) monocyte fraction and differentiate into osteoclasts at or near the bone surface [3, 4]. It has been shown that osteoclast precursors comprise 1–4 % of circulating monocytes and that osteoclasts can be formed from monocyte/macrophage precursors [4, 5]. Osteoclast formation involves a multistep process during which there is loss and gain of monocyte/macrophage and osteoclast markers, respectively [6].
Canonical (RANKL-Dependent) Osteoclast Formation
Canonical osteoclast formation involves an interaction between receptor activator for nuclear factor kappa B ligand (RANKL)-expressing bone stromal cells and RANK-expressing mononuclear phagocyte osteoclast precursors [7, 8]. RANKL is a Tumour Necrosis Factor (TNF) receptor ligand family member and is expressed as a membrane-bound protein by osteoblasts, stromal cells and lymphocytes [9]. At the tissue level mRNA has been detected in developing and adult bone, bone marrow and lymphoid tissue (lymph nodes, spleen and thymus) [10]. In bone, RANKL is cleaved into a soluble molecule (sRANKL) by metalloproteinases and secreted mainly (but not exclusively) by osteoblastic stromal cells [7, 8]. The expression of RANKL by osteoblasts/bone stromal cells is upregulated by osteotropic hormones, such as 1,25-dihydroxy vitamin D3 (1,25(OH)2D3), parathyroid hormone (PTH), and by locally produced cytokines such as TNFα, Interleukin (IL)-1, and IL-6 [11–13]. RANKL not only promotes the differentiation and fusion of osteoclast precursors cells but also activates the bone resorbing activity of mature osteoclasts [14, 15]. The osteoclastogenic effects of RANKL are mediated through its receptor, RANK (also known as TRANCE-R, and TNFRSF11A), which is a type I membrane protein that associates at the cell surface as a trimer [16]. RANK is expressed by dendritic cells, T cells and osteoclasts. RANK mRNA has been detected in the heart, lung, brain, skeletal muscle, kidney, liver and skin [17].
The importance of the RANKL/RANK system in osteoclast formation is evidenced in RANKL knockout mice which exhibit unopposed bone formation; this results in a form of osteopetrosis that is characterised by the absence of osteoclasts on the bone surface with total bone marrow occlusion and failure of tooth eruption [10, 18, 19]. These mice have domed skulls, shortened limbs and abnormal growth plates and suffer spontaneous fractures due to abnormal mineralisation [19]. The osteoclast precursors in these mice are normal as they are able to differentiate into osteoclasts in vitro with the addition of exogenous RANKL. The osteoblasts, however, are defective as they are unable to support osteoclastogenesis, highlighting the osteoblast as the primary cell source of RANKL [18]. In addition to bone abnormalities, these mice exhibit defective development of the thymus and lymphoid tissue, an indication of the wide-ranging effects of RANKL. RANK-deficient mice also exhibit severe osteopetrosis [16, 20].
Osteoprotegerin (OPG), a member of the TNF receptor superfamily (TNFRSF11B), is a soluble decoy receptor for RANKL which competes with RANK for binding to RANKL and in this way blocks RANKL-induced osteoclastogenesis [14, 21]. OPG is expressed in a variety of tissues including brain, skin, liver, heart and bone as well as haematopoietic and immune cells. The main OPG-producing cells are osteoblasts and bone stromal cells [22]. The OPG knockout mouse develops a severe osteoporosis in which there is loss of trabecular and cortical bone associated with a marked increase in osteoclast formation and resorption [23]. The number of osteoblasts is also increased in response to the increase in osteoclastogenesis. Interestingly, these mice also develop vascular calcification indicating that OPG also plays a role in the prevention of arterial calcification [24].
RANKL/RANK binding initiates activation of a range of signal transduction pathways via interaction of the cytoplasmic tail of RANK with the adaptor proteins TNF-related Factor-1 (TRAF1), TRAF2, TRAF3, TRAF5 and TRAF6 [25]. TRAF6 in particular is important for osteoclast differentiation and function. TRAF6 deficient mice develop severe osteopetrosis with defects in bone resorption and tooth eruption [26]. In contrast, progenitor cells derived from TRAF2 or TRAF5-knockout mice show only mildly reduced osteoclastogenesis [27, 28]. TRAF6 mediates activation of the NFκB and c-src /PI 3-kinase/Akt pathways. PI 3-kinase inhibition in vitro results in impaired osteoclastic bone resorption [29]. Mice deficient in both the p50 and p52 subunits of NFκB [30], in c-src [31] or the Src homology 2-containing inositol-5-phosphatase (SHIP) [32] develop osteopetrosis, suggesting that RANKL-induced activation of NFκB and c-src in osteoclast progenitors is crucial for osteoclast differentiation. Many members of the mitogen-activated protein kinase (MAPK) family are also activated downstream of RANK including the p38-MAPKs, c-Jun N-terminal kinases (JNK1, 2 and 3) and extracellular signal-regulated kinases (ERK1 and ERK2). Bone marrow monocytes derived from JNK1-, but not JNK2-, deficient mice show reduced osteoclastogenesis associated with lack of phosphorylation of the JNK substrate c-Jun [33]. Fos (c-Fos, Fos B, FRA-1, FRA-2) and Jun (c-Jun, JunB, JunD) proteins comprise the homodimeric AP-1 transcription factor. Mice lacking JunB [34] or c-Fos develop osteopetrosis because of an early block of differentiation in the osteoclast lineage [35]. FRA-1 is able to rescue c-Fos functions in bone development [36], suggesting a central role for AP-1 in osteoclastogenesis.
Macrophage colony stimulating factor (M-CSF) is a growth factor which plays a crucial role in the proliferation, differentiation, activation and survival of cells of the mononuclear phagocyte system [37]. M-CSF is synthesised by mesenchymal cells, including fibroblasts and osteoblasts, some epithelial cells, and activated macrophages [38, 39]. M-CSF belongs to the tyrosine kinase receptor family and is encoded by the protooncogene c-fms [40]. M-CSF binds to its receptor c-Fms on osteoclast precursors and mature osteoclasts. The essential role of M-CSF in osteoclast formation has been shown in vivo and in vitro. Osteoblasts/bone stromal cells in osteopetrotic (op/op) mice do not produce functionally active M-CSF because of an insertion of an extra thymidine in the coding region of the M-CSF gene [41, 42]. These animals are severely deficient in mature macrophages and osteoclasts. The administration of recombinant human M-CSF corrects the impaired bone resorption in these animals and induces the appearance of resorbing osteoclasts and bone marrow macrophages [41].
Non-Canonical (RANKL-Independent) Osteoclast Formation
Although osteoclast formation induced by RANKL represents the major pathway whereby osteoclastogenesis occurs, a number of other cytokines and growth factors have been shown to be capable of substituting for RANKL to induce osteoclast formation from marrow-derived and circulating osteoclast precursors. These can be divided into humoral factors which, like RANKL, are TNF superfamily members, and those which are not (Table 1). The former include TNFα, APRIL (a proliferation inducing ligand), BAFF (B cell activating factor), and LIGHT (lymphotoxin exhibiting inducible expression and competing with herpes simplex virus glycoprotein D for herpesvirus entry mediator, a receptor expressed by T lymphocytes); the latter include transforming growth factor β (TGFβ), the interleukins IL-6, IL-11, IL-8, nerve growth factor (NGF) and IGF-I and II. The existence of these non-canonical pathways implies that there is a degree of redundancy in growth factor signalling for osteoclast formation; in general non-canonical osteoclastogenesis from marrow and monocyte precursors results in the formation of osteoclasts that are smaller and have fewer nuclei than those formed in RANKL-treated cultures; these smaller osteoclasts produce smaller lacunar resorption pits; in LIGHT-treated cultures, the number of resorption pits formed is similar to that seen in RANKL-treated cultures but the pits are not as deep [1, 43, 47] (Table 1; Fig. 1). The significance of non-canonical osteoclastogenesis in physiological bone resorption is uncertain, but it is likely to play a role in pathological bone resorption where high levels of the cytokines and growth factors known to be capable of substituting for RANKL are generally found.

a Small TRAP+ multinucleated cells and b correspondingly small resorption pits formed in LIGHT-treated cultures (both at ×400 magnification)
Osteoclast Formation by TNF Superfamily Members
TNFα
TNFα is produced by macrophages, monocytes and T cells. The TNFα transgenic mouse has helped to define the mechanism by which elevated levels of TNFα influence canonical and non-canonical osteoclast formation. In these mice there is a four- to seven-fold increase in the number of osteoclast precursors in the peripheral blood mononuclear cells (PBMC) population and the spleen [64]. In this model, the osteoclastogenic effect was not seen following administration of PTHrP, 1,25(OH)2D3 or IL-1β. The increased number of pre-osteoclasts correlated with detectable levels of TNFα in the serum and the onset of inflammatory arthritis. The addition of a TNFα antagonist inhibited this increase in the pre-osteoclast population. This suggested that TNFα, at elevated concentrations, is able to directly impact osteoclastogenesis by increasing the proliferation of osteoclast precursors. TNFα stimulated the formation of TRAP+ and cathepsin K+ osteoclasts on the bone surface. Osteoclasts were only observed near the site of administration and there were no radiological signs of resorption. Although some groups have not observed formation of multinucleated cells capable of resorption when TNFα is added to osteoclast precursors without other factors [44], both Azuma et al. and Kudo et al. found that TNFα is able to induce mouse and human osteoclast differentiation and activation independently of RANKL without the addition of other cytokines [45, 65]. The combination of TNFα and IL-1α in cultures was found to synergistically increase osteoclast formation and lacunar resorption in mouse marrow and human monocyte cell cultures [45, 65]. IL-1α, however, did not induce osteoclast formation alone; there is evidence suggesting that IL-1α directly targets osteoclast precursors to promote differentiation but that it requires permissive levels of RANKL to do so; a similar mechanism may operate with regard to TNFα [66, 67].
TNF-α can induce biological reactions via two cell-surface receptors: TNF receptor type 1 (TNFR1) and TNF receptor type 2 (TNFR2). Each receptor mediates different intracellular signals. Analysis of TNFR1- and TNFR2-deficient mice revealed that TNFR1 induces osteoclast differentiation, whilst TNFR2 inhibits osteoclast differentiation [68]. More recently, two independent regulators, A20 [69] and TSG-6 [70], have also been suggested to play a role in the negative loop effects of TNFα, which in part may explain the various temporal effects of TNFα that have been recorded in in vitro/vivo models of osteoclast formation [71–73].
LIGHT and Other TNF Superfamily Members
LIGHT (lymphotoxin exhibiting inducible expression and competing with herpes simplex virus glycoprotein D for herpesvirus entry mediator, a receptor expressed by T lymphocytes) is a type II transmembrane protein that is a member of the tumour necrosis factor superfamily (TNFSF14) [74, 75]. LIGHT is primarily expressed by cells that play an immunological role including activated T cells, natural killer cells, spleen cells, immature dendritic cells and macrophages [76–80]. The key biological function of LIGHT is modifying innate and adaptive immune responses via T cell differentiation, activation and homeostasis [79, 80]. LIGHT has been implicated in graft versus host diseases, rheumatoid arthritis and tumour cell apoptosis [48, 55, 81–83]. LIGHT binds three receptors: herpesvirus entry mediator (HVEM), lymphotoxin β receptor (LTβR) and decoy receptor 3 (DcR3), the cytoplasmic tails of which show structural similarity. The functions of HVEM and LTβR are cell-context specific, but in general HVEM has survival and growth-inducing properties whilst LTβR mediates cell death [84, 85]. DcR3 is a soluble non-signalling receptor that modulates the functions of LIGHT. DcR3 has been shown to induce osteoclastogenesis in one study [86], although the mechanism whereby this occurs is unclear given that DcR3 does not have signalling capabilities.
LIGHT is capable of inducing osteoclast formation from both murine RAW264.7 macrophage precursors and human peripheral blood mononuclear cells via a process which is not inhibited by OPG or RANK-Fc [47] (Fig. 1). The effect of LIGHT is not mediated through any of its known receptors (HVEM, lymphotoxin β receptor and DcR3). It was noted that the soluble decoy receptor DcR3 dose-dependently inhibited LIGHT-dependent osteoclast formation [47]. Yang et al. reported that DcR3 is able to stimulate osteoclastogenesis independently of both M-CSF and RANKL in human monocytes, RAW264.7 murine macrophages and rat marrow cells via a mechanism involving secretion of TNFα following reverse signalling to activate ERK and p38 MAPK [86]. LIGHT-mediated osteoclastogenesis was originally shown to be inhibited by a neutralising antibody to TNFRII [47], a receptor LIGHT has not previously been shown to bind [46] and, in common with HVEM and LTβR, is not known to associate with TRAF6.
An analysis of the downstream signalling pathways of LIGHT showed that it activates the Akt, NFκB and JNK pathways in mouse and human monocytes [87]. Activation of these pathways is mediated by TRAFs including TRAF6 which plays an important role in osteoclast differentiation [25]. TRAF6 does not bind to TNFR-II [88] so the precise role of TRAF6 in TNFα and LIGHT-mediated osteoclastogenesis is unclear; TRAF6 also does not associate with HVEM or LTβR. Other TRAFs, however, have been shown to bind to the cytoplasmic tail of TNFR-II and RANK [88]. It has been shown that TRAF2 and TRAF5 are involved in TNFα-mediated osteoclastogenesis [27, 28]. Since the cytoplasmic tail of HVEM and LTβR associates with TRAF2 and/or TRAF5 [88], it is possible that HVEM and LTβR mediate the osteoclastogenic actions of LIGHT via these adapter proteins.
Other members of the TNF superfamily, which have been shown to be capable of inducing osteoclast formation, include APRIL (a proliferation inducing ligand), also known as TRDL1 and TNFSF13, and BAFF (B cell activating factor) [60]. APRIL is not present in normal tissues but is highly expressed by numerous tumour cells [89, 90]. BAFF is expressed by dendritic cells and macrophages and stimulates the proliferation of B lymphocytes and their secretion of immunoglobulin; it is also expressed by tumour cells including B leukaemia cells [91]. Numerous TRAP+/VNR+ multinucleated cells that express F-actin rings are generated in APRIL and BAFF-treated monocyte cultures, but these osteoclasts produce significantly less resorption than those formed in RANKL-treated cultures [60]. This finding suggests that not all the osteoclast-like cells formed in these cultures are functional.
Compared with LIGHT, APRIL and BAFF are much less efficient in terms of producing mature resorbing osteoclasts from monocytes [60]. The addition of LIGHT to monocyte cultures results in the formation of numerous osteoclasts and extensive lacunar resorption, approximately one-half that seen in RANKL-treated cultures. HVEM, a receptor for LIGHT, shows increased mRNA expression during osteoclastogenesis. The formation of TRAP+ osteoclasts and resorption pits in APRIL/BAFF-treated cultures is similar to those in TNFα-treated cultures. It has recently been shown that TNFα-induced osteoclast formation is markedly increased in mice lacking NF-κB2 (p100) [61]. The precursor protein NF-κB2 acts as a negative regulator of osteoclast formation and TNFα was found to induce a sustained accumulation of NF-κB2 in osteoclast precursors. A similar mechanism of inhibition may control osteoclast resorption induced by APRIL and BAFF.
Osteoclast Formation by Non-TNF Superfamily Members
Transforming Growth Factor β
Transforming growth factor β (TGFβ) is a multifunctional growth factor that is abundant in bone; it is produced by many cells in bone, including osteoblasts, fibroblasts and osteoclasts [92]. TGFβ1 is the most common isoform of TGFβ [62]. There are contradictory reports regarding the specific actions of TGFβ1 in bone [63, 93, 94]. TGFβ1 is a potent chemoattractant for osteoblasts and can stimulate osteoblast proliferation and early stage differentiation [95]. TGFβ1 knockout mice have no osteoblasts and as a result have fragile bones with a low mineral:matrix ratio. Although there are many reports of the inhibitory effect of TGFβ1 on osteoclast differentiation, there are also several which have noted that TGFβ1 is able to induce osteoclastogenesis and resorption [96]. It is probable that these reported differences in TGFβ effects are due to differences in cell populations, the culture methods and TGF β1 concentration employed in various studies. TGFβ1 inhibits osteoclast formation in long-term co-cultures of bone cells at high concentrations due to its action on osteoblasts whereby it suppresses RANKL and increases OPG levels [97]. However, TGFβ1 has been shown to stimulate osteoclast formation from monocytes in the absence of RANKL and other factors known to induce osteoclast formation [96]. Osteoclasts formed in these cultures, like those in TNFα-treated cultures, are small and form small lacunar resorption pits.
Interleukins
IL-6 and IL-11 share many biological properties and are members of the glycoprotein 130 (gp130) cytokine family; these cytokines are produced by osteoblasts, stromal cells, monocytes and macrophages and other cells in bone [98]. They exert their effects through a common signal transducer, i.e. gp130. IL-6 can stimulate osteoclast precursor proliferation, differentiation and activation in a dose-dependent manner, independently of RANKL [99, 100]. Roodman et al. proposed a model to explain the actions of IL-6 [101]. At low concentrations (below 10 ng/ml), IL-6 stimulates osteoclast formation from precursors, but at high concentrations (above 10 ng/ml) IL-6 predominantly stimulates the activation of mature osteoclasts. It has been shown that IL-6 and IL-11 can induce osteoclast formation from human monocytes [100]. This osteoclast formation is inhibited by the addition of a human antibody to the gp130 receptor. Another interleukin that has been reported to be capable of substituting for RANKL is IL-8 [102, 103]; the possible role of IL-23 in inducing osteoclastogenesis is discussed in Section 4.
Nerve Growth Factor
Nerve growth factor (NGF), a member of the neurotrophin family, is expressed in a number of skeletal cell populations and plays a role in bone fracture repair through stimulation of osteoblasts which enhance osteoclast resorption [104–107]. The addition of NGF to monocyte cultures induces the formation of TRAP+ multinucleated cells which produce approximately five-fold more resorption than APRIL or BAFF [60]. This amount of resorption is comparable to that seen in cultures treated with IL-6 and TGF-β. Sensory and sympathetic neural innervation of the bone and periosteum is well documented [56, 104–108]. Local administration of NGF is known to enhance bone remodelling and is thought to be particularly important in promoting bone formation in fracture healing [57, 104]. Direct neurite-osteoclast cell communication through adrenergic receptors has been shown and it is therefore possible that NGF could act as a neurogenic coupling factor between osteoblasts and osteoclasts [58].
Insulin-Like Growth Factor
Insulin-like growth factor-I/II (IGF-I/II) are non-TNF superfamily growth factors that are known to play a role in bone remodelling. Bone is a major reservoir of IGF-I and IGF-II. IGF-I predominates in rodent bone and IGF-II in human bone [109]. It has been suggested that IGFs may act as coupling factors between osteoblasts and osteoclasts [59, 110, 111]. The effect of IGFs on osteoclasts is not well-defined but promotion of osteoclast formation and activity in vitro has been described [112–114]. Both, IGF I and IGF II, stimulate osteoblast proliferation and matrix synthesis but the effect on osteoclast differentiation is less well characterised [115]. The IGF I knockout mouse clearly exhibits defective osteoblasts and deficient numbers of osteoclasts [116]. In vitro studies have shown that IGF I and IGF II, in the presence of additional growth factors, are able to promote osteoclast differentiation and activation, and it has been shown that IGF I may influence osteoclast formation through regulation of RANKL and RANK expression [117–119]. It has also been shown that both IGF I and IGF II directly support the formation of mature resorbing osteoclasts from human monocyte precursors [60]. Although lacunar resorption in IGF-treated cultures was significantly less than that in RANKL or LIGHT-treated cultures, given the abundance of both IGF I and IGF II in the bone matrix, this mechanism may be significant in physiological bone remodelling. IGFs are stored in the bone matrix and are released upon resorption and can potentially act on nearby osteoblasts and osteoclasts to increase bone remodelling [116, 118]. It has been shown that resorbing osteoclasts express IGF I, IGF II and IGF I receptor mRNA, indicating that osteoclasts are likely to be directly responsive to IGF [119]. An increase in the expression of IGF I mRNA in mature osteoclasts as compared to monocytes has been noted, giving support to the possibility of an autocrine loop.
RANKL-Independent Pathological Bone Resorption
There is now considerable evidence to show that RANKL-independent pathways of osteoclasts formation are likely to play a role in pathological bone resorption associated with several neoplastic and non-neoplastic diseases of bone and joint.
Giant cell tumour of bone (GCTB) is a locally aggressive highly osteolytic bone tumour that contains numerous large, multinucleated osteoclasts. In addition to the osteoclastic giant cell component, there is a significant mononuclear cell population which includes macrophage-like osteoclast precursors and mononuclear stromal cells that are known to express RANKL. Expression of a number of non-canonical osteoclastogenic factors has been noted in GCTB cells including TGFβ, IL-1, IL-6, TNFα, IGF I and II [49, 120–122]. These factors are likely to play a role in the recruitment and formation of the numerous large osteoclasts characteristically seen in this tumour. The expression of mRNA for IGF I and II and the IGF I receptor has been noted in both giant cells and mononuclear stromal cells in GCTB [122]. Mononuclear stromal cells in GCTB are known to exhibit an osteoblast phenotype and it is possible that IGF I and II could represent a therapeutic target which would inhibit replication of these cells and decrease formation of the giant cells in these tumours. Expression of APRIL and BAFF has also been noted in the mononuclear stromal cells and giant cells of GCTB [60, 123].
RANKL-independent osteoclast formation has also been shown to play a role in the osteolysis associated with Ewing sarcoma, metastatic breast carcinoma and melanoma [124–126]. Cultured breast cancer cells produce soluble factors that inhibit RANKL-induced but stimulate non RANKL-induced osteoclast formation from human monocytes. Ewing sarcoma cells express RANKL but, like melanoma cells, also produce a soluble factor that supports osteoclast formation; this is inhibited by an antibody to TNFα. Several RANKL-independent humoral factors are also likely to play a role in the osteolysis associated with myeloma [127]. LIGHT has been shown to increase osteoclast formation in multiple myeloma [128], a condition in which IL-6 and IL-11 have been strongly implicated with regard to osteoclast formation [127]. IL-6 and IL-11 are also thought to play a role in oestrogen deficiency-associated bone loss [129], Paget’s disease [129] and Gorham-Stout disease [130]. It has been shown that anti-BAFF treatment can inhibit osteoclastogenesis in a SCID human multiple myeloma model [131]. APRIL is expressed in a number of tumour cell lines and over-expression of APRIL has been linked with tumour progression, in particular in multiple myeloma [89, 132, 133].
Inflammatory conditions of bone and joint are often characterised by the presence of a heavy macrophage infiltrate. This is seen particularly in aseptic loosening of implant components which is associated with the presence in periprosthetic tissues of numerous implant-derived wear particle-containing foreign body macrophages and periprosthetic osteolysis. It has been shown that two distinct cellular mechanisms of osteoclast formation and bone resorption are likely to operate in aseptic loosening [134]. CD14+ macrophages isolated from the pseudomembrane of loose hip arthroplasties can be induced to form osteoclasts by both RANKL-dependent and TNFα-dependent mechanisms [50, 134]. The addition of an antibody against the p55 receptor subunit of TNF resulted in significant inhibition of osteoclast formation and lacunar resorption. TNFα mediated lipopolysaccharide-stimulated osteoclastogenesis has also been shown to play a role in periodontal disease [51].
Elevated levels of TNFα have been detected in patients with rheumatoid arthritis (RA), and this cytokine is believed to play a central role in the pathogenesis of this disease [52]. High levels of TNFα are known to be present in synovial fluid in many inflammatory joint conditions including RA where there are numerous macrophages as well as lymphoid cells in the synovial membrane. Synovial fluid macrophages in RA are capable of osteoclast formation when incubated with TNFα [53]. It is likely that TNFα (± IL-1)-induced osteoclast formation contributes to the formation of marginal erosions in this condition [54]. Glucocorticoids, which inhibit TNFα-induced osteolysis are of benefit with regard to controlling pathological bone resorption in RA; in contrast, glucocorticoids promote RANKL-dependent osteoclast formation [65]. This finding would argue for two distinct pathways of osteoclast formation, one RANKL-dependent, one RANKL-independent, being operative in conditions of pathological bone resorption.
LIGHT, a potent RANKL-independent osteoclastogenic factor, has been shown in several studies to play a role in RA. An increase in LIGHT levels has been noted in the serum of RA patients [47]. LIGHT is upregulated on B lymphocytes and monocytes in RA and blocking the action of LIGHT reduces the severity of murine collagen-induced arthritis [135, 136]. The formation of osteoclasts from monocytes is significantly decreased by adding a soluble decoy receptor for LIGHT, DcR3, and by blocking antibodies to the p75 component of the TNF receptor [47]. It has been shown that osteoclast formation from monocytes involves interaction with synovial cells [137]. LIGHT promotes RA synovial fibroblast proliferation, survival and activity [138, 139], and cytokine and metalloproteinase expression by synovial macrophages [54]. Polymorphic variants of LIGHT alter binding to HVEM and DcR3 and may influence inflammation and resorption in RA [140]. In recent experiments, we found that LIGHT is highly expressed in the synovial tissue of RA patients and that LIGHT induces RANKL-dependent osteoclast formation from synovial fluid macrophages. BAFF is also associated with autoimmune diseases such as RA, Sjögren’s syndrome and systemic lupus erythematosus and may play a role in osteolysis associated with these inflammatory arthropathies [141, 142]. Adamopoulos and co-workers have noted that in RA and crystal arthritis, synovial fibroblasts produce soluble factor(s) that induce osteoclast formation by a RANKL-independent mechanism [143]; osteoclastogenesis was not altered by inhibition of TNF or IL-6. They also showed that mice overexpressing IL-23 show increased osteoclast formation with increased trabecular and cortical bone loss as well as inflammatory joint destruction [144, 145]; this was associated with an increase in marrow myeloid precursors in contrast to IL-23 deficient mice which showed impaired osteoclast differentiation and function. More recently, they have shown that IL-23-induced osteoclastogenesis may be RANKL-independent and that it involves activation of a unique 12 kDa myeloid protein [146]. IL-23 may thus represent another non-TNF superfamily member capable of inducing RANKL-independent osteoclast formation from mononuclear phagocyte precursors.
Conclusion
Although the RANKL/RANK axis undoubtedly accounts for osteoclastogenesis that occurs in physiological bone remodelling, non-canonical (RANKL-independent) pathways of osteoclast formation are likely to play a role in pathological bone resorption associated with neoplastic and non-neoplastic diseases of bone and joint. Although osteoclastogenic humoral factors are not as potent as RANKL in inducing osteoclast formation, these factors are present at relatively high concentration in osteolytic bone and joint disorders which also commonly have a significant CD14+ (pre-osteoclast) macrophage infiltrate. The existence of multiple non-canonical pathways of osteoclast formation has particular implications for the use of therapies targeting RANKL, such as the anti-RANKL antibody, Denosumab™, which has been employed to treat osteoporosis and GCTB. RANKL-independent osteoclastogenesis could represent a mechanism of escape from this therapy. Further investigation into the mechanisms of action of humoral factors that induce RANKL-independent osteoclast formation, particularly with regard to specific osteolytic diseases of bone and joint, will advance our understanding of pathological bone resorption and may lead to the development of new anti-resorptive therapies.
References
Knowles HJ, Athanasou NA (2009) Canonical & non-canonical pathways of osteoclast formation. Histol Histopathol 24:337–346
Athanasou NA (2011) The osteoclast—what’s new? Skelet Radiol 40:1137–1140
Massey HM, Flanagan AM (1999) Human osteoclasts derive from CD14-positive monocytes. Br J Haematol 106:167–170
Fujikawa Y, Quinn JM, Sabokbar A, McGee JO’D, Athanasou NA (1996) The human osteoclast precursor circulates in the monocyte fraction. Endocrinology 137:4058–4060
Walker DG (1993) Bone resorption restored in osteopetrotic mice by transplants of normal bone marrow and spleen cells. Clin Orthop Relat Res 294:4–6
Faust J, Lacey DL, Hunt P et al (1999) Osteoclast markers accumulate on cells developing from human peripheral blood mononuclear precursors. J Cell Biochem 72:67–80
Lacey DL, Timms E, Tan HL et al (1998) Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 93:165–176
Yasuda H, Shima N, Nakagawa N et al (1998) Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL. Proc Natl Acad Sci U S A 95:3597–3602
Lum L, Wong BR, Josien R et al (1999) Evidence for a role of a tumour necrosis factor-alpha (TNF-alpha)-converting enzyme-like protease in shedding of TRANCE, a TNF family member involved in osteoclastogenesis and dendritic cell survival. J Biol Chem 274:13613–13618
Kong YY, Yoshida H, Sarosi I et al (1999) OPGL is a key regulator of osteoclastogenesis, lymphocyte development and lymph-node organogenesis. Nature 397:315–323
Kitazawa R, Kitazawa S (2002) Vitamin D(3) augments osteoclastogenesis via vitamin D-responsive element of mouse RANKL gene promoter. Biochem Biophys Res Commun 290:650–655
Horwood NJ, Elliott J, Martin TJ, Gillespie MT (1998) Osteotropic agents regulate the expression of osteoclast differentiation factor and osteoprotegerin in osteoblastic stromal cells. Endocrinology 139:4743–4746
Hofbauer LC, Lacey DL, Dunstan CR, Spelsberg TC, Riggs BL, Khosla S (1999) Interleukin-1beta and tumour necrosis factor-alpha, but not interleukin-6, stimulate osteoprotegerin ligand gene expression in human osteoblastic cells. Bone 25:255–259
Atkins GJ, Bouralexis S, Haynes DR et al (2001) Osteoprotegerin inhibits osteoclast formation and bone resorbing activity in giant cell tumors of bone. Bone 28:370–377
Fuller K, Wong B, Fox S, Choi Y, Chambers TJ (1998) TRANCE is necessary and sufficient for osteoblast-mediated activation of bone resorption in osteoclasts. J Exp Med 188:997–1001
Li J, Sarosi I, Yan XQ et al (2000) RANK is the intrinsic hematopoietic cell surface receptor that controls osteoclastogenesis and regulation of bone mass and calcium metabolism. Proc Natl Acad Sci U S A 97:1566–1571
Khosla S (2001) Minireview: the OPG/RANKL/RANK system. Endocrinology 142:5050–5055
Kim N, Odgren PR, Kim DK, Marks SC Jr, Choi Y (2000) Diverse roles of the tumour necrosis factor family member TRANCE in skeletal physiology revealed by TRANCE deficiency and partial rescue by a lymphocyte-expressed TRANCE transgene. Proc Natl Acad Sci U S A 97:10905–10910
Odgren PR, Kim N, MacKay CA, Mason-Savas A, Choi Y, Marks SC Jr (2003) The role of RANKL (TRANCE/TNFSF11), a tumour necrosis factor family member, in skeletal development: effects of gene knockout and transgenic rescue. Connect Tissue Res 44(Suppl 1):264–271
Dougall WC, Glaccum M, Charrier K et al (1999) RANK is essential for osteoclast and lymph node development. Genes Dev 13:2412–2424
Udagawa N, Takahashi N, Yasuda H et al (2000) Osteoprotegerin produced by osteoblasts is an important regulator in osteoclast development and function. Endocrinology 141:3478–3484
Hofbauer LC (1999) Osteoprotegerin ligand and osteoprotegerin: novel implications for osteoclast biology and bone metabolism. Eur J Endocrinol 141:195–210
Mizuno A, Amizuka N, Irie K et al (1998) Severe osteoporosis in mice lacking osteoclastogenesis inhibitory factor/osteoprotegerin. Biochem Biophys Res Commun 247:610–615
Bucay N, Sarosi I, Dunstan CR et al (1998) Osteoprotegerin-deficient mice develop early onset osteoporosis and arterial calcification. Genes Dev 12:1260–1268
Wada T, Nakashima T, Hiroshi N, Penninger JM (2006) RANKL-RANK signaling in osteoclastogenesis and bone disease. Trends Mol Med 2006(12):17–25
Lomaga MA, Yeh WC, Sarosi I et al (1999) TRAF6 deficiency results in osteopetrosis and defective interleukin-1, CD40, and LPS signalling. Genes Dev 13:1015–1024
Kanazawa K, Kudo A (2000) TRAF2 is essential for TNF-alpha-induced osteoclastogenesis. J Bone Miner Res 20:840–847
Kanazawa K, Azuma Y, Nakano H, Kudo A (2003) TRAF5 functions in both RANKL- and TNF alpha-induced osteoclastogenesis. J Bone Miner Res 18:443–450
Nakamura I, Takahashi N, Sasaki T et al (1995) A specific inhibitor of phosphatidylinositol-3 kinase, blocks osteoclastic bone resorption. FEBS Lett 361:79–84
Iotsova V, Caamaño J, Loy J, Yang Y, Lewin A, Bravo R (1997) Osteopetrosis in mice lacking NF-kappaB1 and NF-kappaB2. Nat Med 3:1285–1289
Soriano P, Montgomery C, Geske R, Bradley A (1991) Targeted disruption of the c-src proto-oncogene leads to osteopetrosis in mice. Cell 64:693–702
Takeshita S, Namba N, Zhao J et al (2002) SHIP-deficient mice are severely osteoporotic due to increased numbers of hyper-resorptive osteoclasts. Nat Med 8:943–949
David JP, Sabapathy K, Hoffmann O, Idarraga MH, Wagner EF (2002) JNK1 modulates osteoclastogenesis through both c-Jun phosphorylation-dependent and -independent mechanisms. J Cell Sci 115:4317–4325
Kenner L, Hoebertz A, Beil FT et al (2004) Mice lacking JunB are osteopenic due to cell-autonomous osteoblast and osteoclast defects. J Cell Biol 164:613–623
Wagner EF (2002) Functions of AP1 (Fos/Jun) in bone development. Ann Rheum Dis 61(Suppl 2):40–42
Fleischmann A, Hafezi F, Elliott C, Remé CE, Rüther U, Wagner EF (2000) Fra-1 replaces c-Fos-dependent functions in mice. Genes Dev 14:2695–2700
Felix R, Hofstetter W, Wetterwald A, Cecchini MG, Fleisch H (1994) Role of colony-stimulating factor-1 in bone metabolism. J Cell Biochem 55:340–349
Flanagan AM, Lader CS (1998) Update on the biologic effects of macrophage colony-stimulating factor. Curr Opin Hematol 5:181–185
Motoyoshi K (1998) Biological activities and clinical application of M-CSF. Int J Hematol 67:109–122
Sherr CJ, Rettenmier CW, Sacca R, Roussel MF, Look AT, Stanley ER (1985) The c-fms proto-oncogene product is related to the receptor for the mononuclear phagocyte growth factor, CSF-1. Cell 41:665–676
Felix R, Cecchini MG, Fleisch H (1990) Macrophage colony stimulating factor restores in vivo bone resorption in the op/op osteopetrotic mouse. Endocrinology 127:2592–2594
Yoshida H, Hayashi S, Kunisada T et al (1990) The murine mutation osteopetrosis is in the coding region of the macrophage colony stimulating factor gene. Nature 345:442–444
Mabilleau G, Pascaretti-Grizon F, Basle MF, Chappard D (2012) Depth and volume of resorption induced by osteoclasts generated in the presence of RANKL, TNF-alpha/IL-1 or LIGHT. Cytokine 57(2):294–299
Lam J, Takeshita S, Barker JE, Kanagawa O, Ross FP, Teitelbaum SL (2000) TNF-alpha induces osteoclastogenesis by direct stimulation of macrophages exposed to permissive levels of RANK ligand. J Clin Invest 106:1481–1488
Azuma Y, Kaji K, Katogi R, Takeshita S, Kudo A (2000) Tumour necrosis factor-alpha induces differentiation of and bone resorption by osteoclasts. J Biol Chem 275:4858–4864
Bossen C, Ingold K, Tardivel A et al (2006) Interactions of tumour necrosis factor (TNF) and TNF receptor family members in the mouse and human. J Biol Chem 281:13964–13971
Edwards JR, Sun SG, Locklin R et al (2006) LIGHT (TNFSF14), a novel mediator of bone resorption, is elevated in rheumatoid arthritis. Arthritis Rheum 54:1451–1462
Pasero C, Barbarat B, Just-Landi S et al (2009) A role for HVEM, but not lymphotoxin-beta receptor, in LIGHT-induced tumour cell death and chemokine production. Eur J Immunol 39:2502–2514
Atkins GJ, Haynes DR, Graves SE et al (2000) Expression of osteoclast differentiation signals by stromal elements of giant cell tumors. J Bone Miner Res 15:640–649
Itonaga I, Sabokbar A, Murray DW, Athanasou NA (2000) Effect of osteoprotegerin and osteoprotegerin ligand on osteoclast formation by arthroplasty membrane derived macrophages. Ann Rheum Dis 59:26–31
Tervahartiala T, Koski H, Xu JW, Häyrinen-Immonen R, Hietanen J, Sorsa T, Konttinen YT (2001) Tumor necrosis factor-alpha and its receptors, p55 and p75, in gingiva of adult periodontitis. J Dent Res 80:1535–1539
Brennan F, McInnes IB (2008) Evidence that cytokines play a role in rheumatoid arthritis. J Clin Invest 118:3537–3545
Adamopoulos IE, Sabokbar A, Wordsworth BP, Carr A, Ferguson DJ, Athanasou NA (2006) Synovial fluid macrophages are capable for osteoclast formation and resorption. J Pathol 208:35–43
Chu CQ, Field M, Feldmann M, Maini RN (1991) Localisation of tumour necrosis factor alpha in synovial tissues and at the cartilage-pannus junction in patients with rheumatoid arthritis. Arthritis Rheum 34:1125–1132
Shaikh RB, Santee S, Granger SW et al (2001) Constitutive expression of LIGHT on T cells leads to lymphocyte activation, inflammation, and tissue destruction. J Immunol 167:6330–6337
Hohmann EL, Elde RP, Rysavy JA, Einzig S, Gebhard RL (1986) Innervation of periosteum and bone by sympathetic vasoactive intestinal peptide-containing nerve fibers. Science 232:868–871
Wang L, Zhou S, Liu B et al (2006) Locally applied nerve growth factor enhances bone consolidation in a rabbit model of mandibular distraction osteogenesis. J Orthop Res 24:2238–2245
Togari A (2002) Adrenergic regulation of bone metabolism: possible involvement of sympathetic innervation of osteoblastic and osteoclastic cells. Microsc Res Tech 58:77–84
Mohan S, Jennings JC, Linkhart TA, Baylink DJ (1988) Primary structure of human skeletal growth factor: homology with human insulin-like growth factor-II. Biochim Biophys Acta 966:44–55
Hemingway F, Taylor R, Knowles HJ, Athanasou NA (2011) RANKL-independent human osteoclast formation with APRIL, BAFF, NGF, IGF I and IGF II. Bone 48:938–944
Yao Z, Xing L, Boyce BF (2009) NF-kappaB p100 limits TNF-induced bone resorption in mice by a TRAF3-dependent mechanism. J Clin Invest 119:3024–3034
Centrella M, Horowitz MC, Wozney JM, McCarthy TL (1994) Transforming growth factor-gene family members and bone. Endocr Rev 15:27–39
Pfeilschifter J, Seyedin SM, Mundy GR (1998) Transforming growth factor beta inhibits bone resorption in fetal rat long bone culture. J Clin Invest 82:680–685
Inoue M, Ross FP, Erdmann JM, Abu-Amer Y, Wei S, Teitelbaum SL (2006) Tumour necrosis factor alpha regulates alpha(v)beta5 integrin expression by osteoclast precursors in vitro and in vivo. Endocrinology 141:284–290
Kudo O, Fujikawa Y, Itonaga I, Sabokbar A, Torisu T, Athanasou NA (2002) Proinflammatory cytokine (TNFalpha/IL-1alpha) induction of human osteoclast formation. J Pathol 198:220–227
Ma T, Miyanishi K, Suen A et al (2004) Human interleukin-1-induced murine osteoclastogenesis is dependent on RANKL, but independent of TNF-α. Cytokine 26:138–144
Wei S, Kitaura H, Zhou P, Ross FP, Teitelbaum SL (2005) IL-1 mediates TNF-induced osteoclastogenesis. J Clin Invest 115:282–290
Teitelbaum SL (2007) Osteoclasts: what do they do and how do they do it? Am J Pathol 170(2):427–435
Mabilleau G, Chappard D, Sabokbar A (2011) Role of the A20-TRAF6 axis in lipopolysaccharide-mediated osteoclastogenesis. J Biol Chem 286(5):3242–3249
Mahoney DJ, Swales C, Athanasou NA et al (2011) TSG-6 inhibits osteoclast activity via an autocrine mechanism and is functionally synergistic with osteoprotegerin. Arthritis Rheum 63(4):1034–1043
Fuller K, Murphy C, Kirstein B, Fox SW, Chambers TJ (2002) TNFα potently activates osteoclasts, through a direct action independent of and strongly synergistic with RANKL. Endocrinology 143:1108–1118
Fuller K, Kirstein B, Chambers TJ (2006) Murine osteoclast formation and function: differential regulation by humoral agents. Endocrinology 147:1979–1985
Iqbal J (2006) Does TNF have anti osteoclastogenic actions? Ann N Y Acad Sci 1068:234–239
Mauri DN, Ebner R, Montgomery RI et al (1998) LIGHT, a new member of the TNF superfamily, and lymphotoxin are ligands for herpesvirus entry mediator. Immunity 8:21–30
Tamada K, Shimozaki K, Chapoval AI et al (2000) LIGHT, a TNF-like molecule, costimulates T cell proliferation and is required for dendritic cell-mediated allogeneic T cell response. J Immunol 164:4105–4110
Montgomery RI, Warner MS, Lum BJ, Spear PG (1996) Herpes simplex virus-1 entry into cells mediated by a novel member of TNF/NGF receptor family. Cell 87:427–436
Zhai Y, Guo R, Hsu TL et al (1998) LIGHT, a novel ligand for lymphotoxin receptor and TR2/HVEM, induces apoptosis and suppresses in vivo tumor formation via gene transfer. J Clin Invest 102:1142–1151
Heo SK, Yun HJ, Park WH, Park SD (2008) NADPH oxidase activation is required for migration by LIGHT in human monocytes. Biochem Biophys Res Commun 371:834–840
Schneider K, Potter KG, Ware CF (2004) Lymphotoxin and LIGHT signaling pathways and target genes. Immunol Rev 202:49–66
Chang TH, Hsieh SL, Chao Y, Chou YC, Lin WW (2005) Proinflammatory effects of LIGHT through HVEM and LTbetaR interactions in cultured human umbilical vein endothelial cells. J Biomed Sci 12:363–375
Kang YM, Kim SY, Kang JH et al (2007) LIGHT up-regulated on B lymphocytes and monocytes in rheumatoid arthritis mediates cellular adhesion and metalloproteinase production by synoviocytes. Arthritis Rheum 56:1106–1117
Ye Q, Fraser CC, Gao W et al (2002) Modulation of LIGHT-HVEM costimulation prolongs cardiac allograft survival. J Exp Med 195:795–800
Rooney IA, Butrovich KD, Glass AA et al (2000) The lymphotoxin-beta receptor is necessary and sufficient for LIGHT-mediated apoptosis of tumor cells. J Biol Chem 275:14307–14315
Granger SW, Rickert S (2003) LIGHT-HVEM signaling and the regulation of T cell-mediated immunity. Cytokine Growth Factor Rev 14:289–296
Marsters SA, Ayres TM, Skubatch M, Gray CL, Rothe M, Ashkenazi A (1997) Herpesvirus entry mediator, a member of the tumor necrosis factor receptor (TNFR) family, interacts with members of the TNFR-associated factor family and activates the transcription factors NF-kappaB and AP-1. J Biol Chem 272:14029–14032
Yang CR, Wang JH, Hsieh SL, Wang SM, Hsu TL, Lin WW (2004) Decoy receptor 3 (DcR3) induces osteoclast formation from monocyte/macrophage lineage precursor cells. Cell Death Differ. Suppl 1:S97-107
Hemingway F, Kashima TG, Knowles HJ, Athanasou NA (2013) Investigation of osteoclastogenic signalling of the RANKL substitute LIGHT. Exp Mol Pathol 94:380–385
Arch RH, Gedrich RW, Thompson CB (1998) Tumour necrosis factor receptor-associated factors (TRAFs) - a family of adapter proteins that regulates life and death. Genes Dev 12:2821–2830
Hahne M, Kataoka T, Schroter M et al (1998) APRIL, a new ligand of the tumor necrosis factor family, stimulates tumor cell growth. J Exp Med 188:1185–1190
Kelly K, Manos E, Jensen G, Nadauld L, Jones DA (2000) APRIL/TRDL-1, a tumor necrosis factor-like ligand, stimulates cell death. Cancer Res 60:1021–1027
Schneider P, MacKay F, Steiner V et al (1999) BAFF, a novel ligand of the tumor necrosis factor family, stimulates B cell growth. J Exp Med 189:1747–1756
Bonewald LF (1995) Regulation and regulatory activities of transforming growth factor beta. Crit Rev Eukaryot Gene Exp 9:33–44
Massey HM, Scopes J, Horton MA, Flanagan AM (2002) Transforming growth factor-beta stimulates the osteoclast-forming potential of peripheral blood haematopoietic precursors in a lymphocyte-rich microenvironment. Bone 28:577–582
Beaudreuil J, Mbalaviele G, Cohen-Solal M, Morieux C, de Vernejoul MC, Orcel P (1995) Short-term local injections of transforming growth factor-beta 1 decrease ovariectomy-stimulated osteoclastic resorption in vivo in rat. J Bone Miner Res 10:971–977
Ota K, Quint P, Weivoda MM et al (2013) Transforming growth factor beta 1 induces CXCL16 and leukemia inhibitory factor expression in osteoclasts to modulate migration of osteoblast progenitors. Bone 57:68–75
Itonaga I, Sabokbar A, Sun SG et al (2004) Transforming growth factor-beta induces osteoclast formation in the absence of RANKL. Bone 34:57–64
Takai H, Kanematsu M, Yano K et al (1998) Transforming growth factor-beta stimulates the production of osteoprotegerin/osteoclastogenesis inhibitory factor by bone marrow stromal cells. J Biol Chem 273:27091–27096
Ishimi Y, Miyaura C, Jin CH et al (1990) IL-6 is produced by osteoblasts and induces bone resorption. J Immunol 145:3297–3303
Tamura T, Udagawa N, Takahashi N et al (1993) Soluble interleukin-6 receptor triggers osteoclast formation by interleukin 6. Proc Natl Acad Sci U S A 90:11924–11928
Kudo O, Sabokbar A, Pocock A, Itonaga I, Fujikawa Y, Athanasou NA (2003) Interleukin-6 and interleukin-11 support human osteoclast formation by a RANKL-independent mechanism. Bone 32:1–7
Roodman GD, Kurihara N, Ohsaki Y et al (1992) Interleukin 6. A potential autocrine/paracrine factor in Paget’s disease of bone. J Clin Invest 89:46–52
Bendre MS, Montague DC, Peery T, Akel NS, Gaddy D, Suva LJ (2003) Interleukin-8 stimulation of osteoclastogenesis and bone resorption is a mechanism for the increased osteolysis of metastatic bone disease. Bone 33:28–37
Bendre MS, Margulies AG, Walser B et al (2005) Tumour-derived interleukin-8 stimulates osteolysis independent of the receptor activator of nuclear factor-kappaB ligand pathway. Cancer Res 65:11001–11009
Asaumi K, Nakanishi T, Asahara H, Inoue H, Takigawa M (2000) Expression of neurotrophins and their receptors (TRK) during fracture healing. Bone 26:625–633
Frenkel SR, Guerra LA, Mitchell OG, Singh IJ (1990) Nerve growth factor in skeletal tissues of the embryonic chick. Cell Tissue Res 260:507–511
Grills B, Schuijers J (1998) Immunohistochemical localization of nerve growth factor in fractured and unfractured rat bone. Acta Orthop Scand 69:415–419
Hukkanen M, Konttinen Y, Santavirta S et al (1993) Rapid proliferation of calcitonin gene-related peptide-immunoreactive nerves during healing of rat tibial fracture suggests neural involvement in bone growth and remodelling. Neuroscience 54:969–979
Serre CM, Farlay D, Delmas PD, Chenu C (1990) Evidence for a dense and intimate innervation of the bone tissue, including glutamate-containing fibers. Bone 25:623–629
Bautista CM, Mohan S, Baylink DJ (1990) Insulin-like growth factors I and II are present in the skeletal tissues of ten vertebrates. Metabolism 39:96–100
Hayden JM, Mohan S, Baylink DJ (1995) The insulin-like growth factor system and the coupling of formation to resorption. Bone 17:93S–98S
Hill PA, Reynolds JJ, Meikle MC (1995) Osteoblasts mediate insulin-like growth factor-I and -II stimulation of osteoclast formation and function. Endocrinology 136:124–131
Fukuoka H, Aoyama M, Miyazawa K, Asai K, Goto S (2005) Hypoxic stress enhances osteoclast differentiation via increasing IGF2 production by non-osteoclastic cells. Biochem Biophys Res Commun 328:885–894
Mochizuki H, Hakeda Y, Wakatsuki N et al (1992) Insulin-like growth factor-I supports formation and activation of osteoclasts. Endocrinology 131:1075–1080
Nakao K, Aoyama M, Fukuoka H, Fujita M, Miyazawa K, Asai K, Goto S (2009) IGF II modulates the microenvironment for osteoclastogenesis. Biochem Biophys Res Commun 378:462–466
Zhang M, Xuan S, Bouxsein ML et al (2002) Osteoblast-specific knockout of the insulin-like growth factor (IGF) receptor gene reveals an essential role of IGF signalling in bone matrix mineralization. J Biol Chem 277:44005–44012
Wang Y, Nishida S, Elalieh HZ, Long RK, Halloran BP, Bikle DD (2006) Role of IGF-I signaling in regulating osteoclastogenesis. J Bone Miner Res 21:1350–1358
Guicheux J, Heymann D, Rousselle AV et al (1998) Growth hormone stimulatory effects on osteoclastic resorption are partly mediated by insulin-like growth factor I: an in vitro study. Bone 22:25–31
Bautista CM, Baylink DJ, Mohan S (1991) Isolation of a novel insulin-like growth factor (IGF) binding protein from human bone: a potential candidate for fixing IGF-II in human bone. Biochem Biophys Res Commun 176:756–763
Middleton J, Arnott N, Walsh S, Beresford J (1995) Osteoblasts and osteoclasts in adult human osteophyte tissue express the mRNAs for insulin-like growth factors I and II and the type 1 IGF receptor. Bone 16:287–293
Zheng MH, Fan Y, Smith A, Wysocki S, Papadimitriou JM, Wood DJ (1998) Gene expression of monocyte chemoattractant protein-1 in giant cell tumors of bone osteoclastoma: possible involvement in CD68+ macrophage-like cell migration. J Cell Biochem 70:121–129
Franchi A, Benvenuti S, Masi L et al (2001) TGF-beta isoform and receptor expression in giant cell tumor and giant cell lesions of bone. Appl Immunohistochem Mol Morphol 9:170–175
Middleton J, Arnott N, Walsh S, Beresford J (1996) The expression of mRNA for insulin-like growth factors and their receptor in giant cell tumors of human bone. Clin Orthop 322:224–231
Mhawech-Fauceglia P, Kaya G, Sauter G et al (2006) The source of APRIL up-regulation in human solid tumor lesions. J Leukoc Biol 80:697–704
Lau YS, Adamopoulos IE, Sabokbar A, Giele H, Gibbons CL, Athanasou NA (2007) Cellular and humoral mechanisms of osteoclast formation in Ewing’s sarcoma. Br J Cancer 96:1716–1722
Lau YS, Danks L, Sun SG et al (2007) RANKL-dependent and RANKL-independent mechanisms of macrophage-osteoclast differentiation in breast cancer. Breast Cancer Res Treat 105:7–16
Lau YS, Sabokbar A, Giele H, Cerundolo V, Hofstetter W, Athanasou NA (2006) Malignant melanoma and bone resorption. Br J Cancer 94(10):1496–1503
Oranger A, Carbone C, Izzo M, Grano M. (2013) Cellular mechanisms of multiple myeloma bone disease. Clin Dev Immunol. 289458
Brunetti G, Rizzi R, Oranger A et al (2014) LIGHT/TNFSF14 increases osteoclastogenesis and decreases osteoblastogenesis in multiple myeloma-bone disease. Oncotarget 5:12950–12967
Galson DL, Roodman GD (2014) Pathobiology of Paget’s Disease of Bone. J Bone Metab 21:85–98
Hirayama T, Sabokbar A, Itonaga I, Watt-Smith S, Athanasou NA (2014) Cellular and humoral mechanisms of osteoclast formation and bone resorption in Gorham-Stout disease. J Pathol 195:624–630
Neri P, Kumar S, Fulciniti MT et al (2007) Neutralizing B-cell activating factor antibody improves survival and inhibits osteoclastogenesis in a severe combined immunodeficient human multiple myeloma model. Clin Cancer Res 13:5903–5909
Moreaux J, Cremer FW, Reme T et al (2005) The level of TACI gene expression in myeloma cells is associated with a signature of microenvironment dependence versus a plasmablastic signature. Blood 106:1021–1030
Abe M, Kido S, Hiasa M et al (2006) BAFF and APRIL as osteoclast-derived survival factors for myeloma cells: a rationale for TACI-Fc treatment in patients with multiple myeloma. Leukemia 20:1313–1315
Sabokbar A, Kudo O, Athanasou NA (2003) Two distinct cellular mechanisms of osteoclast formation and bone resorption in periprosthetic osteolysis. J Orthop Res 21:73–80
Yang YM, Kim SY, Kang JH et al (2007) LIGHT up regulated on B lymphocytes and monocytes in rheumatoid arthritis mediates cellular adhesion and metalloproteinase production by synoviocytes. Arthritis Rheum 56:1106–1107
Fava RA, Notidis E, Hunt J et al (2003) A role for the lymphotoxin/LIGHT axis in the pathogenesis of murine collagen-induced arthritis. J Immunol 171:115–116
Ishida S, Yamane S, Nakano S et al (2009) The interaction of monocytes with rheumatoid synovial cells is a key step in LIGHT-mediated inflammatory bone destruction. Immunology 128:315–324
Ishida S, Yamane S, Ochi T et al (2003) LIGHT induces cell proliferation and inflammatory responses of rheumatoid arthritis synovial fibroblasts via lymphotoxin beta receptor. J Rheumatol 35:960–968
Pierer M, Brentano F, Rethage J. et al. (2009) The TNF superfamily member LIGHT contributes to survival and activation of synovial fibroblasts in rheumatoid arthritis
Cheung TC, Coppieters K, Sanjo H et al (2010) Polymorphic variants of LIGHT (TNF superfamily −14) alter receptor avidity and bioavailability. J Immunol 185:1949–1958
Mariette X, Roux S, Zhang J et al (2003) The level of BLyS (BAFF) correlates with the titre of autoantibodies in human Sjogren’s syndrome. Ann Rheum Dis 62:168–171
Pers JO, Daridon C, Devauchelle V et al (2005) BAFF overexpression is associated with autoantibody production in autoimmune diseases. Ann N Y Acad Sci 1050:34–39
Dickerson TJ, Suzuki E, Stanecki C, Shin HS, Qui H, Adamopoulos IE (2012) Rheumatoid and pyrophosphate arthritis synovial fibroblasts induce osteoclastogenesis independently of RANKL, TNF and IL-6. J Autoimmun 39:369–376
Adamopoulos IE, Tessmer M, Chao CC et al (2011) IL-23 is critical for induction of arthritis, osteoclast formation, and maintenance of bone mass. J Immunol 187:951–959
Adamopoulos IE, Mellins ED (2015) Alternative pathways of osteoclastogenesis in inflammatory arthritis. Nat Rev Rheumatol 11:189–194
Shin HS, Sarin R, Dixit N, Wu J, Gershwin E, Bowman EP, Adamopoulos IE (2015) Crosstalk among IL-23 and DNAX activating protein of 12 kDa-dependent pathways promotes osteoclastogenesis. J Immunol 194:316–324
Acknowledgments
We would like to thank Sarah Turton for typing the manuscript. NAA and AS are supported by the Oxford NIHR BRU, The Rosetrees Trust, Sarcoma (UK) and the Bone Cancer Research Trust.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
Afsie Sabokbar, David Mahoney, Francesca Hemingway and Nicholas Athanasou declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Sabokbar, A., Mahoney, D.J., Hemingway, F. et al. Non-Canonical (RANKL-Independent) Pathways of Osteoclast Differentiation and Their Role in Musculoskeletal Diseases. Clinic Rev Allerg Immunol 51, 16–26 (2016). https://doi.org/10.1007/s12016-015-8523-6
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12016-015-8523-6