
Abstract
Inflammatory bowel disease (IBD) is a chronic inflammatory condition that may emerge at a young age and often lasts for life. It often goes through phases of recurrence and remission and has a devastating effect on quality of life. The exact etiology of the disease is still unclear, but it appears that an inappropriate immune response to intestinal flora bacteria in people with a genetic predisposition may cause the disease. Managing inflammatory bowel disease is still a serious challenge. Oxidative stress and free radicals appear to be involved in the pathogenesis of this disease, and a number of studies have suggested the use of antioxidants as a therapeutic approach. The antioxidant and anti-inflammatory properties of some trace elements have led some of the research to focus on studying these trace elements in inflammatory bowel disease. Zinc and selenium are among the most important trace elements that have significant anti-inflammatory and antioxidant properties. Some studies have shown the importance of these trace elements in inflammatory bowel disease. In this review, we have attempted to provide a comprehensive overview of the findings of these studies and to gather current knowledge about the association of these trace elements with the inflammatory process and inflammatory bowel disease.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Inflammatory bowel disease (IBD) is a complex disease, and several factors are involved in its pathogenesis. Genetic susceptibility, intestinal microflora, immune function, and environmental factors appear to be involved in the pathogenesis of IBD [1, 2]. The prevalence of the IBD has increased significantly in both developing and developed countries over the past three decades [3]. IBD is a chronic disease that is often lifelong in its duration. Diagnosis and monitoring of this disease is a serious challenge, and although recent studies have shown advances in introducing less invasive methods for diagnosis and monitoring of IBD, however, colonoscopy and histopathological examinations are still recognized as the gold standard and used routinely [4, 5]. Treatment of the disease also has many challenges, and despite the advances made, it is still difficult to treat IBD effectively. The peak of IBD incidence is at a young age; it reduces the quality of life and ability of people to work, so it will also be detrimental to society in addition to the suffering and costs it imposes on the patient [6, 7]. IBD is an inflammatory disease, and oxidative stress due to its close association with the inflammatory process possibly plays an important role in the pathogenesis IBD [8,9,10]. This issue has led to a significant amount of research into the study of oxidative stress, antioxidants, and nutritional factors in IBD [11]. Trace elements such as zinc and selenium have also been considered, due to their antioxidant and anti-inflammatory properties. Trace elements are inorganic nutrients that are present in body fluids and tissues in small quantities and are essential for the growth and function of the body [12]. Deficiency of essential trace elements is associated with some clinical disorders, and their excessive amounts can also have toxic effects [13, 14]. Zinc and selenium are among the most important trace elements. It has been reported that about 30% of the world population have zinc deficiency and this deficiency is mostly mild to moderate [15]. About 3000 proteins in the human body require zinc as a cofactor, so severe zinc deficiency may be fatal [16, 17]. Zinc deficiency can cause disorders such as dermatitis, loss of appetite, impaired wound healing, increased blood ammonia, and hypogonadism [17]. Zinc is also closely linked to the immune system and the inflammatory process. Zinc regulates oxidative stress, immune response, and inflammation process by targeting Nf-ҡB signaling pathway which has a crucial role in inflammation [18].
It seems that zinc deficiency is common among patients with IBD and has prevalence between 15 and 45%, and given that IBD is a chronic inflammatory disease and associated with immune system function, monitoring and maintaining normal levels of zinc in IBD patients seem to be necessary [19]. The role of selenium is also very important. It is an essential trace element and ranges between 90.1 and 98.7 μg/L that has been suggested as the optimal serum concentration [20]. Selenium deficiency can impair important biological processes such as DNA repairing, immune response, and cell cycle regulation [21,22,23]. Selenium deficiency appears to be associated with an increased risk of diseases such as cardiovascular disease, cancers, and hypothyroidism [24,25,26]. Although evidence has been obtained that sufficient levels of selenium can prevent the development of some cancers, it is still not possible to say that selenium has a preventive effect against cancers [27, 28]. In addition, it appears that values above 120 μg/L of selenium are associated with an increased risk of type 2 diabetes and some cancers [20]. Selenium is present as a cofactor in about 25 human proteins called selenoproteins [21]. Glutathione peroxidase, a very important antioxidant enzyme, is a selenoprotein [20]. Some studies have been reported selenium deficiency among IBD patients, and our previous findings also indicate the occurrence of oxidative stress in IBD, and the key role of glutathione peroxidase in dealing with oxidative stress has been suggested [9, 29]. In the next sections, we will review the importance of oxidative stress in the pathogenesis of IBD. There have been many studies to date that each have somehow demonstrated the antioxidant and anti-inflammatory role of zinc and selenium; in addition, as mentioned above, some studies have been done regarding the trace elements status in IBD. Therefore, in this review article, we have attempted to give an overview of the findings of these studies and tried to review the current knowledge about the importance of these trace elements in the inflammatory process (Tables 1 and 2).
IBD: a Chronic Inflammatory Disease
IBD is an inflammatory disease with a variable course of onset and remission; it is principally due to Crohn’s disease and ulcerative colitis [30]. The main clinical symptoms of this disease are abdominal pain, chronic diarrhea, and rectal bleeding. It has many extra-intestinal complications, such as anemia, which can affect the patients’ quality of life significantly [31, 32]. Crohn’s disease can cause intestinal fistulas that cause frequent urinary tract complications [33]. Generally, IBD have many consequences, but it seems that the most important consequence of this disease is a significant increase in the risk of colorectal cancer [34]. Routine treatment approaches are helpful for maintaining the clinical remission, but sometimes the severity of IBD symptoms is so severe that the patient needs to be hospitalized and sometimes there is no option other than a colectomy to treat the disease. IBD has the greatest in prevalence in Europe and North America. In Norway and the USA, the incidence of ulcerative colitis is 505per 100,000 And 286 per 100,000 respectively [3]. The significant rise in the prevalence of IBD has also been reported in some African, Asian, and South American countries over the past three decades [3, 35]. The exact etiology IBD is not yet well understood, but evidence suggests that the disease is caused by an inappropriate immune response to intestinal bacteria in genetically susceptible individuals [36]. In physiological conditions, the intestinal epithelial cells are a physical barrier against the bacterial infiltration from the intestinal lumen to the lamina propria. A layer of mucous membrane covers the surface of the intestinal epithelium, known as the mucosal barrier. A healthy mucosal barrier depends on the tight junctions between epithelial cells. These tight junctions limit the para-cellular space between adjacent epithelial cells. This limitation is crucial in preventing bacteria from penetrating into the lower layers. In IBD these tight junctions are disrupted, thereby weakening the mucous membrane, and increasing intestinal permeability can lead to increased bacterial penetration into the lamina propria. The infiltration of bacteria into the lamina propria stimulates the immune system and increases the recruitment of innate immune cells such as neutrophils and macrophages into the lamina propria [2, 36]. In these cells, the NF-ĸB signaling pathway is active, and high levels of inflammatory cytokines including IL6, IL1, IL12, TNFα, and Il23 are produced and secreted by these cells [30, 37]. The free radicals and oxidative stress produced by the function of these immune cells also stimulate the secretion of these cytokines [37].
These cytokines induce the recruitment of effector CD4+ T cells (TCD4+) to the lamina propria. These T cells also secrete numerous cytokines and chemokines and reinforce the recruitment of the innate immune cells to lamina propria. Because of the disruption of the intestinal mucosal barrier, bacteria infiltrate the lamina propria continuously, and these successive cycles of inflammation continue, resulting in chronic inflammation [2, 30]. All of these events are summarized in Fig. 1. Currently, therapeutic approaches for IBD treatment are focused on attenuating NF-ĸB signaling; inhibiting production of cytokines; as well as attenuating T cell proliferation, differentiation, and function [2, 38]. As mentioned earlier, it seems that environmental factors are also involved in IBD pathogenesis. For example, studies have shown that smoking increases the risk of Crohn’s disease [39]. Nutrition also appears to be one of the most important environmental factors, as it seems that fast foods and high carbohydrate and fat diets are increasing the risk of IBD, while consuming fiber foods and fruits are associated with low risk of IBD [1]. It seems that zinc and vitamin D deficiency, oral contraceptives, and some antibiotics are other environmental risk factors of IBD [1, 40]. Although there are good advances in identifying environmental risk factors of IBD, further epidemiological studies required to identify other environmental risk factors IBD. In addition, it is not yet well understood how the risk factors mentioned above increase the risk of IBD; therefore, further studies are also required in this regard.

Summary of events that resulted to IBD
Oxidative Stress: a Key Player in the Pathogenesis of IBD
Free radicals are highly reactive compounds that have an unpaired electron. These compounds are produced by the metabolism of oxygen and nitrogen [41,42,43,44,45,46,47]. Free radicals are implicated in defense against infectious agents and are involved in some signaling pathways, but overproduction of free radicals can cause destructive events such as changes in inflammatory response, changes in protein and lipid structure, and e DNA damage [8]. The body’s cells are equipped with an antioxidant defense system to counter the destructive effects of free radicals. Generally, in cells, there is a balance between the production of oxidants and their clearance by the antioxidant system, and when this balance is impaired, oxidative stress will occur [8, 48]. Superoxide anion is the most abundant free radical in human tissues. Macrophages and neutrophils produce high amounts of superoxide anion, so it is obvious that the concentration of this free radical increases during inflammation. The main source of superoxide anion production is mitochondrial electron transport chain, but xanthine oxidase and NADPH oxidase can also produce superoxide anion [8]. Xanthine oxidase function is a major source of superoxide anion in the gastrointestinal tract [8]. Superoxide anion is the source of some other types of free radicals such as hydroxyl radical and per hydroxyl radical [49]. High concentrations of hydroxyl radicals are involved in lipid peroxidation. The peroxidation of lipids has very detrimental consequences. For example, lipid peroxidation can impair the structure and function of cells membranes and can induce the conversion of lipoproteins into pro-inflammatory molecules [8, 50]. Interestingly, in IBD, lipid peroxidation increases, indicating oxidative stress [51]. Superoxide dismutase, catalase, and glutathione peroxidase are three antioxidant enzymes involved in the cleansing of superoxide anion. In the first step, superoxide dismutase converts superoxide anion to H2O2 which has less harmful effects, and in the next step, catalase and glutathione peroxidase convert H2O2 to oxygen and water. Neutrophils and mylo-peroxidase convert H2O2 to hypochlorite ions. In IBD, the concentration and activity of these antioxidant enzymes increase, possibly due to the cell’s response to oxidative stress and increased lipid peroxidation [9, 51]. During the inflammation process, large quantities of free radicals are produced by immune cells, especially neutrophils and macrophages.
Free radicals along with pro-inflammatory cytokines are involved in the pathogenesis of IBD. For example, nitric oxide that results from activity iNOS stimulates the production of TNFα in the colon cells and TNFα by weakening the mucosal barrier and increasing the surface adhesion molecules and increases the penetration of bacteria and neutrophil recruitment which can lead to exacerbation of IBD [52, 53]. Interestingly, TNFα increases production of superoxide anion in colon cells and enhances oxidative stress, and oxidative stress weakens intestinal mucosal barrier by enhancing tyrosine phosphorylation and breakdown of tight junctions proteins [54, 55]. As we mentioned earlier, weakening the intestinal mucosal barrier is a key event in the pathogenesis of IBD. This should be noted that the elevation of iNOS expression in the leukocytes of active IBD patients have been demonstrated in the previous study [56]. Some studies have also shown that the administration of some antioxidant compounds can attenuate the production of TNFα and IL1 in IBD patients [57]. In addition, it seems that oxidative stress can also intensify the adaptive immune response [58]. Therefore, it seems that oxidative stress plays a very important role in the pathogenesis of IBD and there was a close interaction between oxidative stress and pro-inflammatory cytokines. The relationship between NF-ĸB and oxidative stress is also interesting. IL1 and TNFα are involved in NF-ĸB activation. It seems that H2O2 can also reinforce NF-ĸB signaling in T cells [59]. One study has also shown that lack of NRF2 which is a factor involved in enhancing antioxidant defense enhances the NF-ĸB activity that confirms the role of oxidative stress in NF-ĸB activation and, more interestingly, NF-ĸB can enhance the expression of NADPH oxidase and by these ways exacerbate oxidative stress [60, 61]. However, it seems that NF-ĸB has a dual role and can increase the expression of some antioxidant enzymes such as glutathione peroxidase and glutathione S transferase in some situations [62]. In numerous clinical studies on IBD patients, increased levels of some oxidative markers including lipid peroxidation indicator and significant decrease in the level of some antioxidant compounds such as vitamin E and glutathione have been reported [9, 63,64,65]. As noted earlier, the increase in the concentration and activity of some antioxidant enzymes in IBD patients has been reported that this increase is probably due to a compensatory mechanism to counteract oxidative stress. Given these clinical findings and all of the above, it is expected that oxidative stress can intensify IBD severity. However, there is still no strong evidence for the association of oxidative markers as well as antioxidants with disease severity [9].
It appears that some environmental IBD risk factors that we mentioned earlier exert their effects by exacerbating oxidative stress; for example, smoking, which is known as a risk factor for Crohn’s disease, has a synergistic effect with oxidative stress [66]. Nutritional factors are also associated with oxidative stress; for example, glutathione peroxidase is a selenoprotein, and superoxide dismutase also requires zinc for effective function, so lack of micronutrient can attenuate the effectiveness of antioxidant defense. However, the exact mechanism of the effects of oxidative stress on pathogenesis of IBD is still unclear and has not been fully elucidated, and it seems that many aspects of this complex issue remain unclear. In the following sections, we will discuss more about the role of zinc and selenium in oxidative stress and inflammation and pathogenesis of IBD.
Selenium, Oxidative Stress, Inflammation, and IBD
Selenium (Se) is a vital trace element, several compounds of which are known to have antioxidant properties (e.g., selenomethionine and selenocysteine). Selenium acts as the active site of the enzymes which are involved in protection of cells against oxidative stress [67, 68]. As noted above, selenium is integrated into protein structure as the amino acid selenocysteine (Sec) and is encoded by the UGA codon [68]. The identification of Se as a necessary trace element for human health was published by Schwarz Foltz in the 1950s [69]. Many studies have shown an association between serum Se levels and efficient immune systems in human and animal models [21]. Selenium deficiency is common among IBD patients, and some studies have reported selenium deficiency in 30.9% of IBD patients [70]. Selenium is mainly essential because of its combination in selenoproteins with a capacity to reduce inflammatory response. Some selenoproteins protect cells from harmful oxidative radicals containing the glutathione peroxidases (GPx), thioredoxin reductase (TXNRD 1-3), and methionine sulfoxide reductase (Msr) systems which contain the selenocysteine and the selenomethionine at their active sites. Selenoprotein P (SELENOP) which is required for Se transport also contains selenocysteine [71]. Between selenoproteins, the most intensively studied for their role in reactive oxygen species (ROS) reduction were the four isoforms of glutathione peroxidase (GPx), which are expressed in the intestine. GPx1 is expressed by all of intestinal cell types, while GPx2 is commonly expressed in the intestinal epithelial cells. GPx3 secreted by intestinal epithelial cells. GPx4 is expressed in both intestinal epithelial cells and found in the lamina propria [72, 73]. Two important selenoproteins, GPx3 and the selenoprotein P, are circulating in plasma, and their plasma amounts can be used to evaluate the Se status [74]. Glutathione peroxidase (GPx) is an enzyme that is able to reduce hydrogen peroxide (H2O2), phospholipid hydroperoxides, and organic hydroperoxides; also, TrxR isoenzymes reduce oxidized thioredoxins, H2O2, and organic hydroperoxides [72]. Additionally, several knock-out mouse models have indicated the protective roles of GPx isoenzymes against intestinal inflammation and inflammation associated cancer [75,76,77]. As mentioned earlier, this enzyme is involved in the pathogenesis of IBD, and several clinical evidences of increased concentration and activity of this enzyme in IBD patients are available which indicate a compensatory mechanism to reduce the high levels of free radicals produced by immune cells during the inflammatory process.
Various selenoproteins are highly expressed in the intestine and may be involved in the regulation of redox homeostasis. Redox balance in cells is regulated by the Nrf2/Keap1 signaling pathway. Nrf2 is a transcription factor that plays a role in enhancing antioxidant defense. Nrf2 appears to weaken the inflammatory process in IBD, and some studies have suggested that enhancing its activity with compounds such as polyphenols is effective in protecting against intestinal inflammation [78]. A study of mouse models of IBD has shown that the lack of Nrf2 enhances the expression of inflammatory cytokines such as TNFα and IL6 and in addition increases COX2 expression [79]. Under normal conditions, Keap1 binds to Nrf2 and holds it in the cytosol. Under oxidative stress, free radicals modify the structure of Keap1 and weaken its binding to Nrf2. Nrf2 can enter the nucleus and bind to antioxidant/electrophile responsive element (ARE/EpRE) regions to increase the expression of antioxidant genes including GPx and TrxR [80]. Nrf2 can only increase TrxR and GPx expression when sufficient amounts of selenium are available [81]. In addition, there was a direct and significant relationship between serum selenium levels and the expression of some other Nrf2-related genes, including GSTP1 and SOD2 [82]. All of these findings support the hypothesis that selenium is involved in the regulation of redox balance by Nrf2. Furthermore, selenium deficiency appears to increase nitric oxide levels in macrophages flowing LPS stimulation [83]. Studies showed that selenium could possibly reduce inducible NO synthase (INOS) expression. This effect of selenium is probably associated with attenuation of p38 MAPK and NF-ĸB signaling pathways [84]. Selenium can attenuate the LPS-induced oxidative stress and NO elevation. As noted earlier, NO plays an important role in the pathogenesis of IBD and can exert a stimulatory effect on TNFα production. Some therapeutic approaches for the treatment of IBD also focused on inhibition of TNFα function. Therefore, the role of selenium in reducing NO levels should be considered more seriously. Evidence is also available of the beneficial effects of selenium nanoparticles in the inhibition of NF-ĸB and the treatment of colitis, so the efficacy of selenium supplements for the treatment of IBD should be further investigated [85]. Furthermore, some studies revealed the capacity of Se in changing the M1-type macrophages to M2-type macrophages [86].
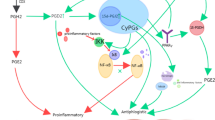
These findings may be important because M1 macrophages have pro-inflammatory properties and play a very destructive role in the pathogenesis of IBD and can exacerbate inflammation by producing high levels of inflammatory cytokines such as IL6 and IL1B. However, M2 macrophages can attenuate inflammation in IBD by producing anti-inflammatory cytokines including IL10. Shifting macrophage polarization to M2 macrophages has been suggested as a therapeutic approach for the treatment of IBD, and many studies have been or are being conducted in this area [87]. Selenoproteins can influence macrophage activity by inhibition of the arachidonic acid pathway and attenuate the production of inflammatory mediator including prostaglandin E2 (PGE2) and decrease the pro-inflammatory genes expressions in macrophages (e.g., NF-휅B) [88, 89]. Selenium is involved in activating PPAR훾 which is involved in production of some anti-inflammatory mediator such as PGD2 [90]. It seems that PGD2 is increased in IBD patients who are in clinical remission [91]. Therefore, PGD2 is likely to play an important role in relieving inflammation. In addition to, PPARγ decreases IL2 secretion by exerting an inhibitory effect on the nuclear factor of activated T cells (NFAT) and thereby attenuates T cell proliferation [92]. It seems that PPARγ expression is reduced in the colonic mucosa of IBD patients [93]. On the other hand, it has been shown that administration of PPARγ agonists to mouse models of IBD has also had some interesting effects to attenuate the severity of chemically induced colitis [94]. All of these findings indicate the protective and important role of PPARγ in IBD, and since selenium plays a role in PPARγ activation, it is necessary to have adequate selenium levels in IBD patients. It seems that selenium may be involved in oxidation of prostaglandin E2 (PGE2) into 15-keto-PGE2 which has a reduced pro-inflammatory ability [89]. PGE2 plays a pro-inflammatory role by enhancing IL23 production and enhancing the IL23/Th17 pathway and is involved in the pathogenesis of IBD [95]. Therefore, it is likely that selenium can reduce the severity of inflammation in IBD by attenuating PGE2 pro inflammatory function. Furthermore, selenium is involved in the regulation of COX activity and prostaglandin biosynthesis, as one of recent studies has shown that selenium can attenuate inflammation in mouse models of colitis by regulating the COX enzyme activity [96]. Selenium seems to be able to decrease COX2 expression in LPS-stimulated macrophages [97]. Because COX2-specific inhibition is considered a therapeutic approach for the treatment of IBD [98], it is likely that sufficient levels of selenium may have beneficial effects in reducing the severity of the disease in IBD patients.
This hypothesis is more supported by some studies findings. For example, Se inadequacy was reported in IBD patients even when they are in remission, and constantly low Se serum status was related to an increased severity of the UC and CD. In addition to low nutritional intake, treatment of IBD with 5-ASA, corticosteroids, and immunosuppressive agents may also decrease selenium levels [29]. Based on the result of some studies, receiving enough selenium supplementation was associated with beneficial health results with respect to oxidative stress-related disorders in the gastrointestinal tract. Similarly, based on studies on mice models, inadequate selenium level in the colitis was revealed, and the severity of the inflammation was reduced with selenium intakes. Some of these studies on mouse models of IBD have shown that selenium enhances the function of IL10, an anti-inflammatory cytokine, and reduces IL17 levels and Th1 and Th17 cell populations [99, 100]. All of these effects indicate the protective role of selenium against chemical induced colitis in mice. We have previously noted the increased risk of colorectal cancer among IBD patients and the role of oxidative stress as a stimulating factor in tumorigenesis. Our previous studies and also other studies have made it clear that lipid peroxidation in IBD increases. Lipid peroxides can damage DNA, which can cause colorectal cancer [101]. Selenium has been shown to have beneficial effects against lipid peroxidation, and it has even been shown that selenium supplements can be useful in this regard [102, 103]. TrxR1 and selenoprotein P can reduce the level of lipid hydroperoxide, and some evidence regarding the protecting roles of selenoprotein P and GPX3 against colitis-associated cancer are available [77, 104,105,106]. Given all of the above, it seems that the assessment of selenium levels in patients with IBD should be taken seriously as low levels of selenium may enhance the inflammatory process and exacerbate the disease. It is not yet clear whether selenium supplements can be effective in maintaining the remission phase in IBD patients, but given the important roles of selenium and selenoproteins in counteracting oxidative stress, attenuating inflammatory signaling pathways, and increasing the population of anti-inflammatory M2 macrophages, this is possible that enough selenium can help maintain remission. Therefore, a clinical trial with large sample size is needed to address this issue.
Zinc, Oxidative Stress, Inflammation, and IBD
Zinc is a nutritionally important mineral for human health. It is absorbed from the gastrointestinal (GI) lumen in the length intestine and basically in the small intestine at the distal duodenum and proximal jejunum [107]. This trace element is essential for stability of structures and function, and over 300 enzymes and 2800 macromolecules containing zinc are involved in cellular signaling pathways and cellular processes [108]. Zinc deficiency is common in IBD patients, during both active and remission phases, with a prevalence rate of 15 to 40% [109,110,111]. Zinc deficiency in IBD disease can occur due to dietary limitation and/or poor zinc absorbency in the small intestine [112]. Preclinical models and human translational studies and animal model studies have demonstrated that decreased serum zinc concentrations may exacerbate inflammation; this Zn effect may be mediated by various pathophysiological mechanisms such as increased generation of pro-inflammatory cells, modulating inflammatory cytokine response, aggravating mucosa leakage, and disruption of epithelial barrier function [113,114,115]. Zinc is a cofactor for several enzymes and plays a crucial role in vital processes including defense against free radicals and regulation cells mediating innate immunity [116]. Zinc is the cofactor of carbonic anhydrase isoenzymes, which maintain acid-base homeostasis, regulate pH, and fluid balance. These isoenzymes are linked to some diseases including mountain sickness, gastric and duodenal ulcers, idiopathic intracranial hypertension, neurological disorders, and osteoporosis [117,118,119,120]. Different cells and tissues have been shown to exhibit oxidative stress under conditions of zinc deficiency, and zinc supplementation can be helpful to prevent oxidative damage [121]. Zinc is incorporated in cellular antioxidant defense system and protects cells from oxidative stress through increasing biosynthesis of glutathione peroxidase (GPx), induces synthesis of metallothioneins (MTs), and inhibits NADPH oxidase as a cofactor of the antioxidant enzyme superoxide dismutase 1 (SOD1) [122,123,124,125,126]. MTs as direct oxidant scavengers are a family of zinc-cytoprotective enzymes that up-regulated in response to an inflammatory stimulus [112, 127]. These proteins are a family of cysteine-rich small proteins that are involved in the suppression of ROS production under oxidative stress condition [127, 128].
Zinc can reinforce the synthesis this zinc-binding proteins by activation of zinc-sensing metal regulatory transcription factor 1 (MTF-1); thus, metallothioneins function seems to be dependent upon the presence of Zn [129, 130]. Studies on both animal models and human have shown that the use of zinc either supplements enhancement or does not alter the level of MTs [131,132,133,134]. A study in a mouse model of dextran sulfate sodium (DSS)-induced colitis showed that MTs play an important role against colonic mucosal inflammation and protection of the intestinal mucosa through their anti-inflammatory function [128]. SOD is a marker of oxidative stress. Cu/Zn-SOD expression, as the main isoform active in IBD, is altered in IBD patients [135,136,137]. Decreased cytoplasmic level and activity of Cu/Zn-SOD may be due to zinc deficiency and chronic inflammation [116, 138]. However, one study has shown that Zn supplementation with Zn gluconate has no effect on the SOD enzyme activity in patients with ulcerative colitis [139].Given the crucial role that this enzyme plays in neutralizing free radicals undoubtedly reduces its activity leading to exacerbated oxidative stress and enhanced inflammation in IBD. As mentioned in the previous sections, NADPH oxidase is one of the major sources of free radical production. Studies have demonstrated that increased activity of the epithelial NADPH oxidase homologs (Nox1, Nox3, Nox4, Nox5, DUOX1, and DUOX2) contributes in intestinal inflammation [140, 141]. A study was performed to investigate the role of NADPH oxidase in intestinal epithelial cells at the active phase of UC. The results of this study showed that in colon epithelial cells, the use of NADPH oxidase inhibitors reduced the levels of oxidants and TNF-α in LPS-treated human colonic epithelium cells isolated from UC patients [142]. As stated, zinc can play an important role in reducing the activity of this enzyme, so it is possible that zinc deficiency can be associated with increased production of free radicals and exacerbated inflammation in IBD. Zinc is also involved in regulation of antioxidant protective responses of nuclear factor erythroid 2-related factor 2 (Nrf2). As noted earlier, Nrf2 plays an essential role in regulation of the cell redox balance and has a significant importance in inflammation [143, 144]. Inflammatory injuries can be alleviated with neutralizing ROS of cells by activating the Nrf2 [145]. In a study on animal model of IBD, effect of Zn in the form of zinc oxide nanoparticles (ZnOnp) has been surveyed, and it has been shown of ZnOnp treatment could attenuate DSS-induced colitis in mice through activation the Nrf2 pathway in the antioxidant defense system. The results of this study suggest that these nanoparticles can be considered a therapeutic approach in IBD patients [146].
In general, it can be said that due to the role of zinc in relation to oxidative stress, zinc supplementation can be considered in IBD patients [147]. In addition to the redox balance, zinc is also closely linked to immune cells and the immunological interactions involved in the development of IBD. As noted earlier, Th17 cells play a critical role in the pathogenesis of IBD. Some studies have shown that Zn can inhibit the development of these cells through its effects on STAT3 signaling which is an essential signaling pathway for Th17 development [115]. In addition, Zn enhances the phosphorylation of IL-1 receptor-associated kinase 4 (IRAK4) and thereby attenuates the IL1β signaling pathway which leads to the attenuation of Th17 cell-dependent inflammatory response [148]. As mentioned in the previous section, impairment of the intestinal mucosal barrier is one of the main causes of IBD. In addition to TNFα, IFN-γ also plays a role in weakening the intestinal mucosal barrier [149]. It seems that Zn can attenuate activator protein-1 (AP-1) signaling, thereby reducing IFN-γ expression [150]. Certainly, reduced expression of this pro-inflammatory cytokine can have positive effects on the intestinal mucosal barrier. In addition, zinc deficiency causes demethylation of the IL6 promoter, thereby increasing the expression of this pro-inflammatory cytokine and enhancing inflammation [151]. Zn also can increase iTreg cells [152]. These cells are involved in the regulation of effector T cell activity and immune tolerance and are important in the pathogenesis of IBD [153]. Zinc deficiency, like selenium deficiency, appears to reduce the population of M2 macrophages as well [154]. Thus, it seems that Zn may also help to weaken the inflammation process in IBD via increasing iTreg cells and M2 macrophage population. Furthermore, zinc can attenuate NF-κB signaling, one of the main inflammatory pathways, leading to a decreased expression of pro-inflammatory cytokines (namely, IL-6 and TNF-α) [155]. Zinc acts as a negative regulator of the NF-κB pathway through the zinc-finger protein A20. It is worth noting that mutations in the gene encoding zinc-finger protein A20 (TNFΑIP3), a susceptibility gene for inflammatory conditions, are positively associated with the risk of developing multiple autoimmune and inflammatory diseases including IBD [151, 156]. The recent studies confirmed that zinc supplementation influences the NF-κB pathway via the alteration of zinc-finger protein A20 activity [157, 158]. It seems that the zinc transporter SLC39A8 or ZIP8 is a negative regulator of the NF-κB pathway and pro-inflammatory response, further supporting the possible role of the trace element in the pathogenesis of IBD [159].
In addition, zinc appears to be associated with the intestinal mucosal barrier. One study showed that zinc depletion enhances occludin proteolysis and down-regulates claudin-3. Because these proteins play a significant role in tight junctions between the intestinal epithelial cells, it seems that low levels of zinc can attenuate the intestinal mucosal barrier [160]. As noted in the preceding sections, attenuation of the intestinal mucosal barrier is one of the key events in the pathogenesis of IBD, so the association of zinc with occludin and claudin-3, discussed above, may be a sign of the preventive role of zinc against IBD development. There is also some clinical evidence on this, which we will refer to in the following sections. Given the above, it seems that further studies on the role of zinc in the pathogenesis of IBD and its mechanisms of action should be undertaken. Given the previously mentioned anti-inflammatory and antioxidant functions of zinc and its protective role in the pathogenesis of IBD as well as the significant prevalence of zinc deficiency among IBD patients, it is likely that providing sufficient levels of zinc can be helpful for more effective treatment of this chronic disease. Monitoring the serum zinc level in these patients will certainly be helpful in this regard.
Clinical Evidence Regarding Zinc and Selenium Importance in IBD
The number of clinical studies examining the selenium and zinc status in IBD is not substantial, and the majority of the research is on animal models. However, lower levels of these trace elements in IBD patients have been reported by the majority of conducted clinical studies. In addition, some of these studies have reported a significant association between deficiency of these trace elements and severity of clinical symptoms and some inflammatory markers in IBD patients. In the preceding sections of this review article, we discuss the role of these trace elements in the pathogenesis of IBD. In this section, we will review the clinical and epidemiological findings to clarify the importance of the subject. One of the most interesting studies in this regard is a study of a population of Korean IBD patients. In this study, decreased selenium levels in 30.9% and decreased zinc levels in 39.0% of patients were reported; in addition, a significant relationship between age under 40 and zinc deficiency was reported. This study also reported a significant relationship between female gender and selenium deficiency as well as between low levels of serum albumin (< 3.3 g/dL) and selenium deficiency [70]. Another study in Iraq reported an inverse correlation between serum selenium levels and serum levels of inflammatory markers (CRP and serum calprotectin) in patients with IBD [161]. These observations are justified by an anti-inflammatory role of zinc and selenium in the pathogenesis of IBD. Another study found that serum selenium levels were lower in IBD patients than in healthy controls and this decrease was more in patients with Crohn’s disease than in ulcerative colitis patients. The findings of this study suggest that factors such as body mass index (BMI), iron status, and duration of disease affect serum selenium levels in patients with IBD [29]. Furthermore, one study, conducted in New Zealand on patients with Crohn’s disease, reported an interesting association between serum selenium levels, three single nucleotide polymorphisms (rs7901303 and rs17529609 in the gene SEPHS1 and rs1553153 in the gene SEPSECS), and Crohn's disease. In addition, lower serum levels of selenium in CD patients compared to healthy subjects were reported. Researchers in the study concluded that there may be an association between low levels of selenium and Crohn’s disease, or low levels of selenium may be due to Crohn’s disease [162]. Further studies seem to be needed to clarify this interesting issue. Evidence has also been provided of the association of selenium with clinical activity in IBD patients, which is in line with the discussions presented in the preceding sections.
In one study, patients with ulcerative colitis were divided into three categories of patients with mild, moderate, and severe disease based on the Truelove and Witts’ severity index, which is a scoring system for evaluation of disease clinical activity. In this study, it was shown that the serum selenium level in patients with severe disease was significantly lower than patients with mild and moderate disease [163]. The benefits of selenium supplements in the treatment of IBD have not been extensively studied yet, although some clinical studies to date have reported promising results. One study has shown that selenium supplementation can reduce some of the clinical symptoms, including stool frequency, in patients with ulcerative colitis [164]. Other study investigating the effect of selenium supplementation on the adverse drug reaction (ADR) of infliximab has shown that selenium supplements can decrease total infliximab ADRs in IBD patients [165]. However, the efficacy of selenium supplementation in the treatment of IBD is still unclear, and given the protective role of selenium in the pathogenesis of IBD that we discussed earlier, as well as the interesting findings from animal models studies, further clinical studies are needed in this regard. Numerous clinical studies have shown a decrease in serum zinc levels in IBD patients compared to healthy control [163, 166]. This reduction in zinc levels may have a nutritional origin, which is likely to be further reduced following IBD, because calprotectin, a protein present in neutrophil granules, is a zinc chelator, and since this protein is secreted in large quantities during inflammation, this is possible that zinc chelating by calprotectin causes a reduction in zinc levels. Evaluation of this hypothesis can be considered in future studies. An epidemiologic study in which 170,776 women were followed for 26 years showed that zinc intake, especially through diet, reduced the risk of IBD [40]. A recent study in Japan also reported a similar result [167]. These interesting clinical findings confirm the protective role of zinc against IBD development discussed in previous sections. Zinc deficiency was associated with an increased risk of adverse disease-specific outcomes, subsequent hospitalizations, and surgeries in IBD patients. In a recent study, 996 IBD patients were analyzed to survey the association of zinc deficiency with the increased risk of subsequent hospitalizations, surgeries, and disease-related complications. The results of this study demonstrated that serum zinc deficiency in these patients was associated with the development of disease-specific complications and also showed the role for monitoring and substitution of zinc in IBD patients associated with improved outcomes [109].
However, based on the results of clinical investigations which were conducted so far, it seems that there is no significant relationship between serum zinc level and clinical activity of the disease in IBD patients [163, 168]. Unfortunately, a clinical trial investigating the efficacy of zinc supplementation in the treatment of IBD is not available. However, as noted earlier, studies on animal and cell models reported promising results. A clinical study that examines the effect of zinc supplements on clinical and endoscopic activity and levels of inflammatory markers can be very useful.
Conclusion and Future Direction
Zinc and selenium are two important trace elements that appear to play a protective role in the pathogenesis of IBD. These trace elements can enhance the body’s antioxidant defense, reduce inflammation by affecting inflammatory signaling pathways and various immune cells, and help strengthen the intestinal mucosal barrier by strengthening tight junctions between epithelial cells. Numerous clinical evidences are available regarding reduced levels of these trace elements in IBD patients, indicating the importance of the subject. On the other hand, the decrease in selenium level seems to be related to the increased clinical activity of the disease, which is a very interesting finding. Although very few clinical studies have investigated the efficacy of the supplementation of these trace elements in the treatment of IBD, studies on animal models have reported very promising findings on the efficacy of nanoparticles containing these trace elements in the treatment of IBD. An in-depth examination of the findings demonstrating the protective role of these trace elements against inflammation and combining these findings with clinical evidence from IBD patients suggests that these trace elements can have preventive effects against IBD and can also help to relieve inflammation and reduce the severity of the disease in IBD patients. It is likely that some events that occur during the pathogenesis of IBD may also result in a decrease in the levels of these trace elements. It seems that the serum levels of these trace elements in patients with IBD should be monitored and the food sources containing these trace elements should be sufficiently consumed. The relationship between serum levels of these trace elements with endoscopic and histopathologic activity of IBD has not been studied yet, but given the anti-inflammatory effects of these elements and the role of zinc in wound healing, it is possible that a relationship exists between the serum levels of these elements and the endoscopic and histopathological activities of IBD. Future studies need to study this topic as it can be very helpful in managing IBD. Another issue that could be addressed in future studies is usefulness of zinc and selenium supplements in the treatment of IBD patients. Given the antioxidant and anti-inflammatory roles of these elements, which are reviewed in depth in this review article, it is possible that these supplements may help to treat IBD more effectively and prolong clinical remission in IBD patients. In overall, the trace elements status in IBD appears to be a neglected subject, and there is a need for more clinical studies to examine this issue more broadly and deeply.
Data Availability
None.
References
Molnar T, Annaházi A (2014) Pathogenesis of ulcerative colitis and Crohn’s disease: similarities, differences and a lot of things we do not know yet. J Clin Cell Immunol 5:2
Abraham C, Cho JH (2009) Inflammatory bowel disease. N Engl J Med 361:2066–2078. https://doi.org/10.1056/NEJMra0804647
Ng SC, Shi HY, Hamidi N, Underwood FE, Tang W, Benchimol EI et al (2018) Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: a systematic review of population-based studies. Lancet. 390:2769–2778. https://doi.org/10.1016/s0140-6736(17)32448-0.
Naganuma M, Hosoe N, Kanai T, Ogata H (2015) Recent trends in diagnostic techniques for inflammatory bowel disease. Korean J Intern Med 30:271–278. https://doi.org/10.3904/kjim.2015.30.3.271
Moein S, Qujeq D, Vaghari Tabari M, Kashifard M, Hajian-Tilaki K (2017) Diagnostic accuracy of fecal calprotectin in assessing the severity of inflammatory bowel disease: from laboratory to clinic. Caspian J Intern Med 8:178–182. https://doi.org/10.22088/cjim.8.3.178
Johnston RD, Logan RF (2008) What is the peak age for onset of IBD? Inflamm Bowel Dis 14(Suppl 2):S4–S5. https://doi.org/10.1002/ibd.20545
Devlen J, Beusterien K, Yen L, Ahmed A, Cheifetz AS, Moss AC (2014) The burden of inflammatory bowel disease: a patient-reported qualitative analysis and development of a conceptual model. Inflamm Bowel Dis 20:545–552. https://doi.org/10.1097/01.mib.0000440983.86659.81
Piechota-Polanczyk A, Fichna J (2014) Review article: the role of oxidative stress in pathogenesis and treatment of inflammatory bowel diseases. Naunyn Schmiedebergs Arch Pharmacol 387:605–620. https://doi.org/10.1007/s00210-014-0985-1
Vaghari-Tabari M, Moein S, Qujeq D, Kashifard M, Hajian-Tilaki K (2018) Positive correlation of fecal calprotectin with serum antioxidant enzymes in patients with inflammatory bowel disease: accidental numerical correlation or a new finding? Am J Med Sci 355:449–455. https://doi.org/10.1016/j.amjms.2017.12.009
Vaghari Tabari M, Moein S, Qujeq D, Kashifard M, Shokri Shirvani J, Hajian Tilaki K et al (2017) Evaluation of the potential antioxidant role of high-density lipoprotein-cholesterol (HDL-C) in patients with ulcerative colitis. Ann Colorectal Res 5:e13699. https://doi.org/10.5812/acr.13699
Moura FA, de Andrade KQ, Dos Santos JCF, Araujo ORP, Goulart MOF (2015) Antioxidant therapy for treatment of inflammatory bowel disease: does it work? Redox Biol 6:617–639. https://doi.org/10.1016/j.redox.2015.10.006
Choi R, Sun J, Yoo H, Kim S, Cho YY, Kim HJ, Kim S, Chung J, Oh SY, Lee SY (2016) A prospective study of serum trace elements in healthy Korean pregnant women. Nutrients 8. https://doi.org/10.3390/nu8110749
Bhattacharya PT, Misra SR, Hussain M (2016) Nutritional aspects of essential trace elements in oral health and disease: an extensive review. Scientifica (Cairo) 2016:5464373. https://doi.org/10.1155/2016/5464373
Goldhaber SB (2003) Trace element risk assessment: essentiality vs. toxicity. Regul Toxicol Pharmacol 38:232–242
Sapkota M, Knoell DL (2018) Essential role of zinc and zinc transporters in myeloid cell function and host defense against infection. J Immunol Res 2018:4315140. https://doi.org/10.1155/2018/4315140
Escobedo Monge MF, Barrado E, Alonso Vicente C, Redondo Del Rio MP, Marugan de Miguelsanz JM (2019) Zinc nutritional status in patients with cystic fibrosis. Nutrients 11. https://doi.org/10.3390/nu11010150
Evans GW (1986) Zinc and its deficiency diseases. Clin Physiol Biochem 4:94–98
Gammoh NZ, Rink L (2017) Zinc in infection and inflammation. Nutrients 9. https://doi.org/10.3390/nu9060624
Gilca-Blanariu GE, Diaconescu S, Ciocoiu M, Stefanescu G (2018) New insights into the role of trace elements in IBD. Biomed Res Int 2018:1813047. https://doi.org/10.1155/2018/1813047
Stoffaneller R, Morse NL (2015) A review of dietary selenium intake and selenium status in Europe and the Middle East. Nutrients. 7:1494–1537. https://doi.org/10.3390/nu7031494
Avery JC, Hoffmann PR (2018) Selenium, selenoproteins, and immunity. Nutrients 10. https://doi.org/10.3390/nu10091203
Bera S, De Rosa V, Rachidi W, Diamond AM (2013) Does a role for selenium in DNA damage repair explain apparent controversies in its use in chemoprevention? Mutagenesis. 28:127–134. https://doi.org/10.1093/mutage/ges064
Zeng H (2009) Selenium as an essential micronutrient: roles in cell cycle and apoptosis. Molecules. 14:1263–1278. https://doi.org/10.3390/molecules14031263
Flores-Mateo G, Navas-Acien A, Pastor-Barriuso R, Guallar E (2006) Selenium and coronary heart disease: a meta-analysis. Am J Clin Nutr 84:762–773. https://doi.org/10.1093/ajcn/84.4.762
Jaworska K, Gupta S, Durda K, Muszynska M, Sukiennicki G, Jaworowska E et al (2013) A low selenium level is associated with lung and laryngeal cancers. PLoS One 8:e59051. https://doi.org/10.1371/journal.pone.0059051
Pizzulli A, Ranjbar A (2000) Selenium deficiency and hypothyroidism: a new etiology in the differential diagnosis of hypothyroidism in children. Biol Trace Elem Res 77:199–208. https://doi.org/10.1385/bter:77:3:199
Fritz H, Kennedy D, Fergusson D, Fernandes R, Cooley K, Seely A et al (2011) Selenium and lung cancer: a systematic review and meta analysis. PLoS One 6:e26259. https://doi.org/10.1371/journal.pone.0026259
Vinceti M, Filippini T, Del Giovane C, Dennert G, Zwahlen M, Brinkman M et al (2018) Selenium for preventing cancer. Cochrane Database Syst Rev 1:CD005195. https://doi.org/10.1002/14651858.CD005195.pub4.
Castro Aguilar-Tablada T, Navarro-Alarcon M, Quesada Granados J, Samaniego Sanchez C, Rufian-Henares JA, Nogueras-Lopez F (2016) Ulcerative colitis and Crohn's disease are associated with decreased serum selenium concentrations and increased cardiovascular risk. Nutrients 8. https://doi.org/10.3390/nu8120780
Moein S, Vaghari-Tabari M, Qujeq D, Majidinia M, Nabavi SM, Yousefi B (2019) MiRNAs and inflammatory bowel disease: an interesting new story. J Cell Physiol 234:3277–3293. https://doi.org/10.1002/jcp.27173
Hendrickson BA, Gokhale R, Cho JH (2002) Clinical aspects and pathophysiology of inflammatory bowel disease. Clin Microbiol Rev 15:79–94. https://doi.org/10.1128/cmr.15.1.79-94.2002
Tulewicz-Marti E, Moniuszko A, Rydzewska G (2017) Management of anemia in inflammatory bowel disease: a challenge in everyday clinical practice. Prz Gastroenterol 12:239–243. https://doi.org/10.5114/pg.2017.72096
Manganiotis AN, Banner MP, Malkowicz SB (2001) Urologic complications of Crohn's disease. Surg Clin North Am 81:197–215, x. https://doi.org/10.1016/s0039-6109(05)70281-4
Bopanna S, Ananthakrishnan AN, Kedia S, Yajnik V, Ahuja V (2017) Risk of colorectal cancer in Asian patients with ulcerative colitis: a systematic review and meta-analysis. Lancet Gastroenterol Hepatol 2:269–276. https://doi.org/10.1016/s2468-1253(17)30004-3
Ng WK, Wong SH, Ng SC (2016) Changing epidemiological trends of inflammatory bowel disease in Asia. Intest Res 14:111–119. https://doi.org/10.5217/ir.2016.14.2.111
Kaser A, Zeissig S, Blumberg RS (2010) Inflammatory bowel disease. Annu Rev Immunol 28:573–621. https://doi.org/10.1146/annurev-immunol-030409-101225
Guan G, Lan S (2018) Implications of antioxidant systems in inflammatory bowel disease. Biomed Res Int 2018:1290179. https://doi.org/10.1155/2018/1290179
Tambuwala MM (2016) Natural nuclear factor kappa beta inhibitors: safe therapeutic options for inflammatory bowel disease. Inflamm Bowel Dis 22:719–723. https://doi.org/10.1097/mib.0000000000000655
Rosenfeld G, Bressler B (2012) The truth about cigarette smoking and the risk of inflammatory bowel disease. Am J Gastroenterol 107:1407–1408. https://doi.org/10.1038/ajg.2012.190.
Ananthakrishnan AN, Khalili H, Song M, Higuchi LM, Richter JM, Chan AT (2015) Zinc intake and risk of Crohn's disease and ulcerative colitis: a prospective cohort study. Int J Epidemiol 44:1995–2005. https://doi.org/10.1093/ije/dyv301
Ak T, Gülçin I (2008) Antioxidant and radical scavenging properties of curcumin. Chem Biol Interact 174:27–37. https://doi.org/10.1016/j.cbi.2008.05.003
Gülçin I (2009) Antioxidant activity of L-adrenaline: a structure-activity insight. Chem Biol Interact 179:71–80. https://doi.org/10.1016/j.cbi.2008.09.023
Gülçin İ (2012) Antioxidant activity of food constituents: an overview. Arch Toxicol 86:345–391. https://doi.org/10.1007/s00204-011-0774-2
Gülçin İ (2010) Antioxidant properties of resveratrol: a structure–activity insight. Innovative Food Sci Emerg Technol 11:210–218. https://doi.org/10.1016/j.ifset.2009.07.002
Gülçin İ (2011) Antioxidant activity of eugenol: a structure-activity relationship study. J Med Food 14:975–985. https://doi.org/10.1089/jmf.2010.0197
Taslimi P, Gulçin İ (2018) Antioxidant and anticholinergic properties of olivetol. J Food Biochem 42:e12516. https://doi.org/10.1111/jfbc.12516
Gulcin İ (2020) Antioxidants and antioxidant methods: an updated overview. Arch Toxicol 94:651–715. https://doi.org/10.1007/s00204-020-02689-3
Balmus IM, Ciobica A, Trifan A, Stanciu C (2016) The implications of oxidative stress and antioxidant therapies in inflammatory bowel disease: clinical aspects and animal models. Saudi J Gastroenterol 22:3–17. https://doi.org/10.4103/1319-3767.173753
Phaniendra A, Jestadi DB, Periyasamy L (2015) Free radicals: properties, sources, targets, and their implication in various diseases. Indian J Clin Biochem 30:11–26. https://doi.org/10.1007/s12291-014-0446-0
Wong-Ekkabut J, Xu Z, Triampo W, Tang IM, Tieleman DP, Monticelli L (2007) Effect of lipid peroxidation on the properties of lipid bilayers: a molecular dynamics study. Biophys J 93:4225–4236. https://doi.org/10.1529/biophysj.107.112565
Achitei D, Ciobica A, Balan G, Gologan E, Stanciu C, Stefanescu G (2013) Different profile of peripheral antioxidant enzymes and lipid peroxidation in active and non-active inflammatory bowel disease patients. Dig Dis Sci 58:1244–1249. https://doi.org/10.1007/s10620-012-2510-z
Yasukawa K, Tokuda H, Tun X, Utsumi H, Yamada K (2012) The detrimental effect of nitric oxide on tissue is associated with inflammatory events in the vascular endothelium and neutrophils in mice with dextran sodium sulfate-induced colitis. Free Radic Res 46:1427–1436. https://doi.org/10.3109/10715762.2012.732698
Al-Sadi R, Guo S, Ye D, Ma TY (2013) TNF-alpha modulation of intestinal epithelial tight junction barrier is regulated by ERK1/2 activation of Elk-1. Am J Pathol 183:1871–1884. https://doi.org/10.1016/j.ajpath.2013.09.001
Kuwano Y, Tominaga K, Kawahara T, Sasaki H, Takeo K, Nishida K et al (2008) Tumor necrosis factor alpha activates transcription of the NADPH oxidase organizer 1 (NOXO1) gene and upregulates superoxide production in colon epithelial cells. Free Radic Biol Med 45:1642–1652. https://doi.org/10.1016/j.freeradbiomed.2008.08.033
Rao RK, Basuroy S, Rao VU, Karnaky KJ Jr, Gupta A (2002) Tyrosine phosphorylation and dissociation of occludin-ZO-1 and E-cadherin-beta-catenin complexes from the cytoskeleton by oxidative stress. Biochem J 368:471–481. https://doi.org/10.1042/bj20011804
Baranipour S, Amini Kadijani A, Qujeq D, Shahrokh S, Haghazali M, Mirzaei A et al (2018) Inducible nitric oxide synthase as a potential blood-based biomarker in inflammatory bowel diseases. Gastroenterol Hepatol Bed Bench 11:S124–S1S8
Reimund JM, Allison AC, Muller CD, Dumont S, Kenney JS, Baumann R et al (1998) Antioxidants inhibit the in vitro production of inflammatory cytokines in Crohn's disease and ulcerative colitis. Eur J Clin Investig 28:145–150. https://doi.org/10.1046/j.1365-2362.1998.00257.x
Sutti S, Jindal A, Locatelli I, Vacchiano M, Gigliotti L, Bozzola C et al (2014) Adaptive immune responses triggered by oxidative stress contribute to hepatic inflammation in NASH. Hepatology. 59:886–897. https://doi.org/10.1002/hep.26749
Schreck R, Rieber P, Baeuerle PA (1991) Reactive oxygen intermediates as apparently widely used messengers in the activation of the NF-kappa B transcription factor and HIV-1. EMBO J 10:2247–2258
Pan H, Wang H, Wang X, Zhu L, Mao L (2012) The absence of Nrf2 enhances NF-kappaB-dependent inflammation following scratch injury in mouse primary cultured astrocytes. Mediat Inflamm 2012:217580. https://doi.org/10.1155/2012/217580
Anrather J, Racchumi G, Iadecola C (2006) NF-kappaB regulates phagocytic NADPH oxidase by inducing the expression of gp91phox. J Biol Chem 281:5657–5667. https://doi.org/10.1074/jbc.M506172200
Morgan MJ, Liu ZG (2011) Crosstalk of reactive oxygen species and NF-kappaB signaling. Cell Res 21:103–115. https://doi.org/10.1038/cr.2010.178
Tuzun A, Erdil A, Inal V, Aydin A, Bagci S, Yesilova Z et al (2002) Oxidative stress and antioxidant capacity in patients with inflammatory bowel disease. Clin Biochem 35:569–572
Sido B, Hack V, Hochlehnert A, Lipps H, Herfarth C, Droge W (1998) Impairment of intestinal glutathione synthesis in patients with inflammatory bowel disease. Gut. 42:485–492. https://doi.org/10.1136/gut.42.4.485
Kuroki F, Iida M, Tominaga M, Matsumoto T, Hirakawa K, Sugiyama S et al (1993) Multiple vitamin status in Crohn's disease. Correlation with disease activity. Dig Dis Sci 38:1614–1618. https://doi.org/10.1007/bf01303168
Isik B, Ceylan A, Isik R (2007) Oxidative stress in smokers and non-smokers. Inhal Toxicol 19:767–769. https://doi.org/10.1080/08958370701401418
Mangiapane E, Pessione A, Pessione E (2014) Selenium and selenoproteins: an overview on different biological systems. Curr Protein Pept Sci 15:598–607. https://doi.org/10.2174/1389203715666140608151134
Tinggi U (2008) Selenium: its role as antioxidant in human health. Environ Health Prev Med 13:102–108. https://doi.org/10.1007/s12199-007-0019-4
Stadtman TC (2005) Selenoproteins--tracing the role of a trace element in protein function. PLoS Biol 3:e421. https://doi.org/10.1371/journal.pbio.0030421
Han YM, Yoon H, Lim S, Sung MK, Shin CM, Park YS et al (2017) Risk factors for vitamin D, zinc, and selenium deficiencies in Korean patients with inflammatory bowel disease. Gut Liver 11:363–369. https://doi.org/10.5009/gnl16333
Labunskyy VM, Hatfield DL, Gladyshev VN (2014) Selenoproteins: molecular pathways and physiological roles. Physiol Rev 94:739–777. https://doi.org/10.1152/physrev.00039.2013
Kudva AK, Shay AE, Prabhu KS (2015) Selenium and inflammatory bowel disease. Am J Physiol Gastrointest Liver Physiol 309:G71–G77. https://doi.org/10.1152/ajpgi.00379.2014
Tian T, Wang Z, Zhang J (2017) Pathomechanisms of oxidative stress in inflammatory bowel disease and potential antioxidant therapies. Oxidative Med Cell Longev 2017:4535194. https://doi.org/10.1155/2017/4535194
Combs GF Jr, Watts JC, Jackson MI, Johnson LK, Zeng H, Scheett AJ et al (2011) Determinants of selenium status in healthy adults. Nutr J 10:75. https://doi.org/10.1186/1475-2891-10-75
Esworthy RS, Aranda R, Martin MG, Doroshow JH, Binder SW, Chu FF (2001) Mice with combined disruption of Gpx1 and Gpx2 genes have colitis. Am J Physiol Gastrointest Liver Physiol 281:G848–G855. https://doi.org/10.1152/ajpgi.2001.281.3.G848
Krehl S, Loewinger M, Florian S, Kipp AP, Banning A, Wessjohann LA et al (2012) Glutathione peroxidase-2 and selenium decreased inflammation and tumors in a mouse model of inflammation-associated carcinogenesis whereas sulforaphane effects differed with selenium supply. Carcinogenesis. 33:620–628. https://doi.org/10.1093/carcin/bgr288
Barrett CW, Ning W, Chen X, Smith JJ, Washington MK, Hill KE et al (2013) Tumor suppressor function of the plasma glutathione peroxidase gpx3 in colitis-associated carcinoma. Cancer Res 73:1245–1255. https://doi.org/10.1158/0008-5472.can-12-3150
Nunes C, Teixeira N, Serra D, Freitas V, Almeida L, Laranjinha J (2016) Red wine polyphenol extract efficiently protects intestinal epithelial cells from inflammation via opposite modulation of JAK/STAT and Nrf2 pathways. Toxicol Res (Camb) 5:53–65. https://doi.org/10.1039/c5tx00214a
Khor TO, Huang MT, Kwon KH, Chan JY, Reddy BS, Kong AN (2006) Nrf2-deficient mice have an increased susceptibility to dextran sulfate sodium-induced colitis. Cancer Res 66:11580–11584. https://doi.org/10.1158/0008-5472.can-06-3562
Cebula M, Schmidt EE, Arner ES (2015) TrxR1 as a potent regulator of the Nrf2-Keap1 response system. Antioxid Redox Signal 23:823–853. https://doi.org/10.1089/ars.2015.6378
Brigelius-Flohe R, Muller M, Lippmann D, Kipp AP (2012) The yin and yang of nrf2-regulated selenoproteins in carcinogenesis. Int J Cell Biol 2012:486147. https://doi.org/10.1155/2012/486147
Reszka E, Wieczorek E, Jablonska E, Janasik B, Fendler W, Wasowicz W (2015) Association between plasma selenium level and NRF2 target genes expression in humans. J Trace Elem Med Biol 30:102–106. https://doi.org/10.1016/j.jtemb.2014.11.008
Prabhu KS, Zamamiri-Davis F, Stewart JB, Thompson JT, Sordillo LM, Reddy CC (2002) Selenium deficiency increases the expression of inducible nitric oxide synthase in RAW 264.7 macrophages: role of nuclear factor-kappaB in up-regulation. Biochem J 366:203–209. https://doi.org/10.1042/bj20020256
Kim SH, Johnson VJ, Shin TY, Sharma RP (2004) Selenium attenuates lipopolysaccharide-induced oxidative stress responses through modulation of p38 MAPK and NF-kappaB signaling pathways. Exp Biol Med (Maywood) 229:203–213. https://doi.org/10.1177/153537020422900209
Zhu C, Zhang S, Song C, Zhang Y, Ling Q, Hoffmann PR et al (2017) Selenium nanoparticles decorated with Ulva lactuca polysaccharide potentially attenuate colitis by inhibiting NF-kappaB mediated hyper inflammation. J Nanobiotechnology 15:20. https://doi.org/10.1186/s12951-017-0252-y
Nelson SM, Lei X, Prabhu KS (2011) Selenium levels affect the IL-4-induced expression of alternative activation markers in murine macrophages. J Nutr 141:1754–1761. https://doi.org/10.3945/jn.111.141176
Abron JD, Singh NP, Price RL, Nagarkatti M, Nagarkatti PS, Singh UP (2018) Genistein induces macrophage polarization and systemic cytokine to ameliorate experimental colitis. PLoS One 13:e0199631. https://doi.org/10.1371/journal.pone.0199631
Vunta H, Belda BJ, Arner RJ, Channa Reddy C, Vanden Heuvel JP, Sandeep Prabhu K (2008) Selenium attenuates pro-inflammatory gene expression in macrophages. Mol Nutr Food Res 52:1316–1323. https://doi.org/10.1002/mnfr.200700346
Kaushal N, Kudva AK, Patterson AD, Chiaro C, Kennett MJ, Desai D et al (2014) Crucial role of macrophage selenoproteins in experimental colitis. J Immunol 193:3683–3692. https://doi.org/10.4049/jimmunol.1400347
Gandhi UH, Kaushal N, Ravindra KC, Hegde S, Nelson SM, Narayan V et al (2011) Selenoprotein-dependent up-regulation of hematopoietic prostaglandin D2 synthase in macrophages is mediated through the activation of peroxisome proliferator-activated receptor (PPAR) gamma. J Biol Chem 286:27471–27482. https://doi.org/10.1074/jbc.M111.260547
Vong L, Ferraz JG, Panaccione R, Beck PL, Wallace JL (2010) A pro-resolution mediator, prostaglandin D(2), is specifically up-regulated in individuals in long-term remission from ulcerative colitis. Proc Natl Acad Sci U S A 107:12023–12027. https://doi.org/10.1073/pnas.1004982107
Choi JM, Bothwell AL (2012) The nuclear receptor PPARs as important regulators of T-cell functions and autoimmune diseases. Mol Cell 33:217–222. https://doi.org/10.1007/s10059-012-2297-y
Le Loupp AG, Bach-Ngohou K, Bourreille A, Boudin H, Rolli-Derkinderen M, Denis MG et al (2015) Activation of the prostaglandin D2 metabolic pathway in Crohn's disease: involvement of the enteric nervous system. BMC Gastroenterol 15:112. https://doi.org/10.1186/s12876-015-0338-7
Choo J, Lee Y, Yan XJ, Noh TH, Kim SJ, Son S et al (2015) A novel peroxisome proliferator-activated receptor (PPAR)gamma agonist 2-hydroxyethyl 5-chloro-4,5-didehydrojasmonate exerts anti-inflammatory effects in colitis. J Biol Chem 290:25609–25619. https://doi.org/10.1074/jbc.M115.673046
Sheibanie AF, Yen JH, Khayrullina T, Emig F, Zhang M, Tuma R et al (2007) The proinflammatory effect of prostaglandin E2 in experimental inflammatory bowel disease is mediated through the IL-23-- > IL-17 axis. J Immunol 178:8138–8147. https://doi.org/10.4049/jimmunol.178.12.8138
Kaur R, Thakur S, Rastogi P, Kaushal N (2018) Resolution of Cox mediated inflammation by Se supplementation in mouse experimental model of colitis. PLoS One 13:e0201356. https://doi.org/10.1371/journal.pone.0201356
Dhanjal NI, Sharma S, Skalny AV, Skalnaya MG, Ajsuvakova OP, Tinkov AA et al (2019) Selenium-rich maize modulates the expression of prostaglandin genes in lipopolysaccharide-stimulated RAW264.7 macrophages. Food Funct 10:2839–2846. https://doi.org/10.1039/c9fo00186g
Kim W, Lee Y, Jeong S, Nam J, Lee S, Jung Y (2015) Colonic delivery of celecoxib is a potential pharmaceutical strategy for repositioning the selective COX-2 inhibitor as an anti-colitic agent. Arch Pharm Res 38:1830–1838. https://doi.org/10.1007/s12272-015-0602-y
Sang L, Chang B, Zhu J, Yang F, Li Y, Jiang X et al (2016) Dextran sulfate sodium-induced acute experimental colitis in C57BL/6 mice is mitigated by selenium. Int Immunopharmacol 39:359–368. https://doi.org/10.1016/j.intimp.2016.07.034
Sang LX, Chang B, Zhu JF, Yang FL, Li Y, Jiang XF et al (2017) Sodium selenite ameliorates dextran sulfate sodium-induced chronic colitis in mice by decreasing Th1, Th17, and gammadeltaT and increasing CD4(+)CD25(+) regulatory T-cell responses. World J Gastroenterol 23:3850–3863. https://doi.org/10.3748/wjg.v23.i21.3850
Thorsteinsdottir S, Gudjonsson T, Nielsen OH, Vainer B, Seidelin JB (2011) Pathogenesis and biomarkers of carcinogenesis in ulcerative colitis. Nat Rev Gastroenterol Hepatol 8:395–404. https://doi.org/10.1038/nrgastro.2011.96
El-Demerdash FM, Nasr HM (2014) Antioxidant effect of selenium on lipid peroxidation, hyperlipidemia and biochemical parameters in rats exposed to diazinon. J Trace Elem Med Biol 28:89–93. https://doi.org/10.1016/j.jtemb.2013.10.001
Akil M, Bicer M, Menevse E, Baltaci AK, Mogulkoc R (2011) Selenium supplementation prevents lipid peroxidation caused by arduous exercise in rat brain tissue. Bratisl Lek Listy 112:314–317
Short SP, Pilat JM, Williams CS (2018) Roles for selenium and selenoprotein P in the development, progression, and prevention of intestinal disease. Free Radic Biol Med 127:26–35. https://doi.org/10.1016/j.freeradbiomed.2018.05.066
Bjornstedt M, Hamberg M, Kumar S, Xue J, Holmgren A (1995) Human thioredoxin reductase directly reduces lipid hydroperoxides by NADPH and selenocystine strongly stimulates the reaction via catalytically generated selenols. J Biol Chem 270:11761–11764. https://doi.org/10.1074/jbc.270.20.11761
Rock C, Moos PJ (2010) Selenoprotein P protects cells from lipid hydroperoxides generated by 15-LOX-1. Prostaglandins Leukot Essent Fat Acids 83:203–210. https://doi.org/10.1016/j.plefa.2010.08.006
Lee HH, Prasad AS, Brewer GJ, Owyang C (1989) Zinc absorption in human small intestine. Am J Physiol 256:G87–G91
Prasad AS (2014) Zinc is an antioxidant and anti-inflammatory agent: its role in human health. Front Nutr 1:14
Siva S, Rubin DT, Gulotta G, Wroblewski K, Pekow J (2016) Zinc deficiency is associated with poor clinical outcomes in patients with inflammatory bowel disease. Inflamm Bowel Dis 23:152–157
Vagianos K, Bector S, McConnell J, Bernstein CN (2007) Nutrition assessment of patients with inflammatory bowel disease. J Parenter Enter Nutr 31:311–319
Alkhouri RH, Hashmi H, Baker RD, Gelfond D, Baker SS (2013) Vitamin and mineral status in patients with inflammatory bowel disease. J Pediatr Gastroenterol Nutr 56:89–92
Skrovanek S, DiGuilio K, Bailey R, Huntington W, Urbas R, Mayilvaganan B et al (2014) Zinc and gastrointestinal disease. World J Gastrointest Pathophysiol 5:496
Ranaldi G, Ferruzza S, Canali R, Leoni G, Zalewski PD, Sambuy Y et al (2013) Intracellular zinc is required for intestinal cell survival signals triggered by the inflammatory cytokine TNFα. J Nutr Biochem 24:967–976
Mayer LS, Uciechowski P, Meyer S, Schwerdtle T, Rink L, Haase H (2014) Differential impact of zinc deficiency on phagocytosis, oxidative burst, and production of pro-inflammatory cytokines by human monocytes. Metallomics. 6:1288–1295
Kitabayashi C, Fukada T, Kanamoto M, Ohashi W, Hojyo S, Atsumi T et al (2010) Zinc suppresses Th17 development via inhibition of STAT3 activation. Int Immunol 22:375–386. https://doi.org/10.1093/intimm/dxq017
Mohammadi E, Qujeq D, Taheri H, Hajian-Tilaki K (2017) Evaluation of serum trace element levels and superoxide dismutase activity in patients with inflammatory bowel disease: translating basic research into clinical application. Biol Trace Elem Res 177:235–240
Boztaş M, Çetinkaya Y, Topal M (2015) Gülçin İ, Menzek A, Şahin E, et al. Synthesis and carbonic anhydrase isoenzymes I, II, IX, and XII inhibitory effects of dimethoxybromophenol derivatives incorporating cyclopropane moieties. J Med Chem 58:640–650. https://doi.org/10.1021/jm501573b.
Arabaci B, Gulcin I, Alwasel S (2014) Capsaicin: a potent inhibitor of carbonic anhydrase isoenzymes. Molecules. 19:10103–10114. https://doi.org/10.3390/molecules190710103
Topal F, Gulcin I, Dastan A, Guney M (2017) Novel eugenol derivatives: potent acetylcholinesterase and carbonic anhydrase inhibitors. Int J Biol Macromol 94:845–851. https://doi.org/10.1016/j.ijbiomac.2016.10.096
Gülçin İ, Trofimov B, Kaya R, Taslimi P, Sobenina L, Schmidt E et al (2020) Synthesis of nitrogen, phosphorus, selenium and sulfur-containing heterocyclic compounds - determination of their carbonic anhydrase, acetylcholinesterase, butyrylcholinesterase and α-glycosidase inhibition properties. Bioorg Chem 103:104171. https://doi.org/10.1016/j.bioorg.2020.104171
Oteiza PI (2012) Zinc and the modulation of redox homeostasis. Free Radic Biol Med 53:1748–1759
Ruz M, Carrasco F, Rojas P, Codoceo J, Inostroza J, Basfi-Fer K et al (2013) Zinc as a potential coadjuvant in therapy for type 2 diabetes. Food Nutr Bull 34:215–221
Chasapis CT, Loutsidou AC, Spiliopoulou CA, Stefanidou ME (2012) Zinc and human health: an update. Arch Toxicol 86:521–534
Sato M, Bremner I (1993) Oxygen free radicals and metallothionein. Free Radic Biol Med 14:325–337
Bogani D, Morgan MA, Nelson AC, Costello I, McGouran JF, Kessler BM et al (2013) The PR/SET domain zinc finger protein Prdm4 regulates gene expression in embryonic stem cells but plays a nonessential role in the developing mouse embryo. Mol Cell Biol 33:3936–3950
Alam S, Kelleher SL (2012) Cellular mechanisms of zinc dysregulation: a perspective on zinc homeostasis as an etiological factor in the development and progression of breast cancer. Nutrients. 4:875–903
Choi S, Liu X, Pan Z (2018) Zinc deficiency and cellular oxidative stress: prognostic implications in cardiovascular diseases. Acta Pharmacol Sin 39:1120
Tsuji T, Naito Y, Takagi T, Kugai M, Yoriki H, Horie R et al (2013) Role of metallothionein in murine experimental colitis. Int J Mol Med 31:1037–1046
Westin G, Schaffner W (1988) A zinc-responsive factor interacts with a metal-regulated enhancer element (MRE) of the mouse metallothionein-I gene. EMBO J 7:3763–3770
Itoh N, Shibayama H, Kanekiyo M, Namphung D, Nakanishi T, Matsuyama A et al (2005) Reduced bactericidal activity and nitric oxide production in metallothionein-deficient macrophages in response to lipopolysaccharide stimulation. Toxicology. 216:188–196
Mulder T, Veer AS, Verspaget H, Griffioen G, Peña A, Janssens A et al (1994) Effect of oral zinc supplementation on metallothionein and superoxide dismutase concentrations in patients with inflammatory bowel disease. J Gastroenterol Hepatol 9:472–477
Di Leo V, D'Incà R, Barollo M, Tropea A, Fries W, Mazzon E et al (2001) Effect of zinc supplementation on trace elements and intestinal metallothionein concentrations in experimental colitis in the rat. Dig Liver Dis 33:135–139
Tran C, Ball J, Sundar S, Coyle P, Howarth G (2007) The role of zinc and metallothionein in the dextran sulfate sodium-induced colitis mouse model. Dig Dis Sci 52:2113–2121
Tran C, Butler R, Philcox J, Rofe A, Howarth G, Coyle P (1998) Regional distribution of metallothionein and zinc in the mouse gut. Biol Trace Elem Res 63:239–251
Kruidenier L, Kuiper I, Lamers CB, Verspaget HW (2003) Intestinal oxidative damage in inflammatory bowel disease: semi-quantification, localization, and association with mucosal antioxidants. J Pathol 201:28–36
Lih-Brody L, Powell SR, Collier KP, Reddy GM, Cerchia R, Kahn E et al (1996) Increased oxidative stress and decreased antioxidant defenses in mucosa of inflammatory bowel disease. Dig Dis Sci 41:2078–2086
Kruidenier L, Kuiper I, van Duijn W, Mieremet-Ooms MA, van Hogezand RA, Lamers CB et al (2003) Imbalanced secondary mucosal antioxidant response in inflammatory bowel disease. J Pathol 201:17–27
Seguí J, Gironella M, Sans M, Granell S, Gil F, Gimeno M et al (2004) Superoxide dismutase ameliorates TNBS-induced colitis by reducing oxidative stress, adhesion molecule expression, and leukocyte recruitment into the inflamed intestine. J Leukoc Biol 76:537–544
Matias JP, Costa DM, Cruz KJC, Silva KG, Feitosa MM, Medeiros LGO et al (2015) Effect of zinc supplementation on superoxide dismutase activity in patients with ulcerative rectocolitis. Nutr Hosp 31:1434–1437
Bedard K, Krause K-H (2007) The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol Rev 87:245–313
Szanto I, Rubbia-Brandt L, Kiss P, Steger K, Banfi B, Kovari E et al (2005) Expression of NOX1, a superoxide-generating NADPH oxidase, in colon cancer and inflammatory bowel disease. J Pathol 207:164–176
Ramonaite R, Skieceviciene J, Kiudelis G, Jonaitis L, Tamelis A, Cizas P et al (2013) Influence of NADPH oxidase on inflammatory response in primary intestinal epithelial cells in patients with ulcerative colitis. BMC Gastroenterol 13:159
Kim J, Cha Y-N, Surh Y-J (2010) A protective role of nuclear factor-erythroid 2-related factor-2 (Nrf2) in inflammatory disorders. Mutat Res 690:12–23
Khor TO, Huang M-T, Prawan A, Liu Y, Hao X, Yu S et al (2008) Increased susceptibility of Nrf2 knockout mice to colitis-associated colorectal cancer. Cancer Prev Res 1:187–191
Lee K-M, Kang K, Lee SB, Nho CW (2013) Nuclear factor-E2 (Nrf2) is regulated through the differential activation of ERK1/2 and PKC α/βII by Gymnasterkoreayne B. Cancer Lett 330:225–232
Li J, Chen H, Wang B, Cai C, Yang X, Chai Z et al (2017) ZnO nanoparticles act as supportive therapy in DSS-induced ulcerative colitis in mice by maintaining gut homeostasis and activating Nrf2 signaling. Sci Rep 7:43126
Gîlcă-Blanariu G-E, Diaconescu S, Ciocoiu M, Ștefănescu G (2018) New insights into the role of trace elements in IBD. Biomed Res Int 2018:1813047.
Lee H, Kim B, Choi YH, Hwang Y, Kim DH, Cho S et al (2015) Inhibition of interleukin-1beta-mediated interleukin-1 receptor-associated kinase 4 phosphorylation by zinc leads to repression of memory T helper type 17 response in humans. Immunology. 146:645–656. https://doi.org/10.1111/imm.12536
Chiba H, Kojima T, Osanai M, Sawada N (2006) The significance of interferon-gamma-triggered internalization of tight-junction proteins in inflammatory bowel disease. Sci STKE 2006:pe1. https://doi.org/10.1126/stke.3162006pe1
Hayashi K, Ishizuka S, Yokoyama C, Hatae T (2008) Attenuation of interferon-gamma mRNA expression in activated Jurkat T cells by exogenous zinc via down-regulation of the calcium-independent PKC-AP-1 signaling pathway. Life Sci 83:6–11. https://doi.org/10.1016/j.lfs.2008.04.022
Wong CP, Rinaldi NA, Ho E (2015) Zinc deficiency enhanced inflammatory response by increasing immune cell activation and inducing IL6 promoter demethylation. Mol Nutr Food Res 59:991–999. https://doi.org/10.1002/mnfr.201400761
Rosenkranz E, Maywald M, Hilgers RD, Brieger A, Clarner T, Kipp M et al (2016) Induction of regulatory T cells in Th1-/Th17-driven experimental autoimmune encephalomyelitis by zinc administration. J Nutr Biochem 29:116–123. https://doi.org/10.1016/j.jnutbio.2015.11.010
Ueno A, Ghosh A, Hung D, Li J, Jijon H (2015) Th17 plasticity and its changes associated with inflammatory bowel disease. World J Gastroenterol 21:12283–12295. https://doi.org/10.3748/wjg.v21.i43.12283
Kido T, Ishiwata K, Suka M, Yanagisawa H (2019) Inflammatory response under zinc deficiency is exacerbated by dysfunction of the T helper type 2 lymphocyte-M2 macrophage pathway. Immunology. 156:356–372. https://doi.org/10.1111/imm.13033
Kilby K, Mathias H, Boisvenue L, Heisler C, Jones JL (2019) Micronutrient absorption and related outcomes in people with inflammatory bowel disease: a review. Nutrients. 11:1388
Ma A, Malynn BA (2012) A20: linking a complex regulator of ubiquitylation to immunity and human disease. Nat Rev Immunol 12:774
Morgan CI, Ledford JR, Zhou P, Page K (2011) Zinc supplementation alters airway inflammation and airway hyperresponsiveness to a common allergen. J Inflamm 8:36
Jarosz M, Olbert M, Wyszogrodzka G, Młyniec K, Librowski T (2017) Antioxidant and anti-inflammatory effects of zinc. Zinc-dependent NF-κB signaling. Inflammopharmacology. 25:11–24
Liu M-J, Bao S, Gálvez-Peralta M, Pyle CJ, Rudawsky AC, Pavlovicz RE et al (2013) The zinc transporter SLC39A8 is a negative feedback regulator of NF-κB through zinc-mediated inhibition of IKK. Cell Rep 3:386
Miyoshi Y, Tanabe S, Suzuki T (2016) Cellular zinc is required for intestinal epithelial barrier maintenance via the regulation of claudin-3 and occludin expression. Am J Physiol Gastrointest Liver Physiol 311:G105–G116. https://doi.org/10.1152/ajpgi.00405.2015
Younus MM, Taher MA, Askar BA, Kurmanji JM (2015) Serum selenium correlations with C-reactive protein, serum calprotectin and disease activity in IBD patients treated with infliximab. World J Pharm Pharm Sci 4(9):165–179
Gentschew L, Bishop KS, Han DY, Morgan AR, Fraser AG, Lam WJ et al (2012) Selenium, selenoprotein genes and Crohn's disease in a case-control population from Auckland, New Zealand. Nutrients. 4:1247–1259. https://doi.org/10.3390/nu4091247
Poursadegh F, Ahadi M, Vosoughinia H, Salehi M, Beheshti Namdar A, Farzanehfar MR et al (2018) A STROBE compliant observational study on trace elements in patients with ulcerative colitis and their relationship with disease activity. Medicine (Baltimore) 97:e13523. https://doi.org/10.1097/md.0000000000013523
Stedman JD, Spyrou NM, Millar AD, Altaf WJ, Akanle OA, Rampton DS (1997) Selenium supplementation in the diets of patients suffering from ulcerative colitis. J Radioanal Nucl Chem 217:189–191. https://doi.org/10.1007/BF02034441
Younus MM, Taher MA, ALMaliki JM, AlKhalidi NM (2015) Selenium supplementation may decrease the rate of infliximab ADRS in IBD patients. World J Pharm Pharm Sci 4(9):193–203
Stochel-Gaudyn A, Fyderek K, Koscielniak P (2019) Serum trace elements profile in the pediatric inflammatory bowel disease progress evaluation. J Trace Elem Med Biol 55:121–126. https://doi.org/10.1016/j.jtemb.2019.06.016
Kobayashi Y, Ohfuji S, Kondo K, Fukushima W, Sasaki S, Kamata N et al (2019) Association between dietary iron and zinc intake and development of ulcerative colitis: a case-control study in Japan. J Gastroenterol Hepatol. https://doi.org/10.1111/jgh.14642
Mechie NC, Mavropoulou E, Ellenrieder V, Petzold G, Kunsch S, Neesse A et al (2019) Serum vitamin D but not zinc levels are associated with different disease activity status in patients with inflammatory bowel disease. Medicine (Baltimore) 98:e15172. https://doi.org/10.1097/md.0000000000015172
Acknowledgments
We are very grateful to Prof. Gordon Ferns for his helpful comments.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Ethical Approval and Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Vaghari-Tabari, M., Jafari-Gharabaghlou, D., Sadeghsoltani, F. et al. Zinc and Selenium in Inflammatory Bowel Disease: Trace Elements with Key Roles?. Biol Trace Elem Res 199, 3190–3204 (2021). https://doi.org/10.1007/s12011-020-02444-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12011-020-02444-w