
Abstract
Grifola frondosa is a medicinal macro-fungus with a wide range of biological activities. Polysaccharides from Grifola frondosa (PGF) play a positive role in regulating blood glucose and alleviating kidney injury. Here, we investigated the exact mechanism of action by which PGF ameliorates diabetic nephropathy. Our results showed that PGF effectively improved glucose tolerance and insulin sensitivity in streptozocin (STZ)-induced DN mice. Additionally, administration of PGF also ameliorated renal function and inflammatory response in STZ-induced DN mice. Consistent with the in vitro results, the high glucose-induced inflammatory response and apoptosis of renal tubular epithelial cells were decreased by PGF treatment. Furthermore, PGF not only suppressed the expression of TLR4, but also more effectively protected the kidney and reduced the inflammatory response when TLR4 was inhibited. All these data revealed that PGF alleviates diabetic nephropathy by blocking the TLR4/NF-κB pathway.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Diabetic nephropathy (DN) is an important complication of diabetes mellitus and is the primary contributor to end-stage renal disease and kidney failure worldwide [1]. DN is characterized by rapid decline in renal blood filtration capacity, which causes increased albuminuria, decreased glomerular filtration rate and hypertrophied renal tubule epithelium [2]. Although DN is a non-immune disease, some studies suggested that inflammatory responses are in correlation with the pathogenesis and progression of this disease. This means that DN progression is accompanied by elevated levels of inflammatory cytokines, namely, IL-6, TNF-α, and TGF-β1 [3]. Moreover, the activation of inflammatory cytokines promotes abnormal secretion of extracellular matrix, causing diffuse thickening of basement membrane and glomerulosclerosis in the kidney [4, 5].
Toll-like receptors (TLRs), a kind of transmembrane receptor, can recognize exogenous pathogens or cell damage. Therefore, they are essential for immune response, cell signal transduction, and inflammatory response. TLR4 expression is increased in various cell types and organs of diabetic patients. TLR4 is activated by inducing innate immune response when tissue injury occurs, and the downstream myeloid differentiation factor 88 (Myd88) is recruited for intracellular signal transducing, which then activates nuclear factor kappa-B (NF-κB) and upregulates inflammatory cytokines [6, 7]. Therefore, the positive regulation of the TLR4/NF-κB network may improve DN pathogenesis.
Grifola frondosa, a large medicinal and edible fungus, has been extensively studied [8]. Polysaccharide from Grifola frondosa (PGF) has been demonstrated to have a number of pharmacological properties, including anti-tumor [9], anti-oxidant [10], and anti-hyperlipidemic effects [11], suggesting the clinical and research value of PGF. In addition, our previous study revealed that PGF regulated glucose metabolism and reduced the level of proteinuria and prevented renal fibrosis [12, 13]. Unfortunately, the exact mechanism by which PGF alleviates renal injury in the progression of DN is still uncertain. Here, we delineated the potential molecular mechanism whereby PGF protects the kidneys against DN and examined whether this mechanism involves the TLR4/NF-κB network.
Materials and Methods
PGF Preparation
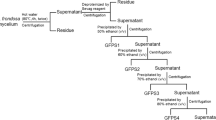
Grifola frondosa was obtained from the China General Microbiological Culture Collection Center (Beijing, China). PGF was isolated from the water extraction of Grifola frondosa as described in our previous studies [12]. In brief, the powder of Grifola frondosa was extracted thrice with boiling water, and the extract was filtered and concentrated. The precipitate was washed with ethanol and lyophilized to obtain PGF.
Experimental Animals and DN Induction
All animal protocols received ethical approval from the Zhuhai College of Science and Technology. Male C57BL/6 J mice (8-week-old) were obtained from Guangdong Laboratory Animal Institution. After acclimatization for 7 days, eight mice were randomly selected as the negative control group (NC). The other mice were intraperitoneally administered with 60 mg/kg (dose/weight) streptozocin (STZ). The mice with blood glucose (BG) readings above 11.2 mmol/L and the level of urine microalbumin exceeded 30 mg/day were selected as the DN model. The DN mice were then divided into 4 groups of 8 mice per group: the diabetic nephropathy group (DN), low dosage of PGF (100 mg/kg, PGF100), medium dosage of PGF (300 mg/kg, PGF300), and high dosage of PGF (500 mg/kg, PGF500). The dose of PGF was used according to the previous study [12]. Physiological parameters, including BG and body weight (BW), were recorded every week. All mice were executed after 8 weeks of administration. Before killing, serum and kidney samples were collected and maintained at 80 °C until further analysis.
Glucose and Insulin Tolerance Test
The mice were fasted at least 12 h with free access to water before oral glucose tolerance testing, glucose solution (2 g/kg) was administered directly into the stomach, and then the BG levels were recorded with the glucometer at 0, 15, 30, 60, and 120 min. The mice were also fasted before the insulin tolerance evaluation, insulin (0.75 U/kg) was intraperitoneally administered with a sterile gauge needle, and then the BG levels were recorded with a glucometer at 0, 30, 60, and 90 min.
Biochemical Indexes and Inflammatory Cytokine Analyses
All experimental mice were housed in separate metabolic cages for 24 h before killing. Urine was collected to measure 24-h albuminuria. Serum creatinine (Scr) and blood urea nitrogen (BUN) were detected with the full-automatic biochemical analyzer (Hitachi, Japan). Inflammatory cytokines like IL-6, IL-1β, TGF-β1, and TNF-α in the serum were evaluated using ELISA kits (Invitrogen, USA), following kit directions.
Cell Culture and Treatment
For NRK-52E cells, a renal tubular epithelial cell line was acquired from The Cell Bank of Type Culture Collection (Shanghai, China) and grown in DMEM medium containing 5.5 mM d-glucose, 10% fetal bovine serum, and 1% penicillin–streptomycin. The cells were pretreated with 30 mM high glucose (HG) for 24 h before exposure to varying PGF doses (10, 30 and 50 μM) for 24 h or 1 μM of the TLR-4 antagonist resatorvid (TAK242) 12 h prior to PGF treatment.
qRT-PCR
Total RNA was isolated from cells and renal tissues with the RNAiso plus reagent (Takara, Japan), before conversion into cDNA using PrimeScript RT master mix (Takara, Japan). Quantitative PCR was conducted with the SYBR Premix Ex Taq (Takara, Japan) on a CFX96 real-time PCR system (Bio-Rad, USA). Gene expression was adjusted to β-actin levels, with the ΔΔCt formula. Table 1 lists the primer sequences used in this study.
Western Blot Analysis
Kidney and cell lysate homogenates were prepared for protein extraction. Equal amounts of proteins were loaded into 10% SDS–polyacrylamide gels before transfer to PVDF membranes (Bio-Rad, USA), which were then placed in 5% non-fat dry milk in PBS for 1 h, before an overnight 4 °C incubation with primary antibodies, followed by an additional incubation with secondary antibody, before protein visualization with a chemiluminescence imaging analysis system (Tanon 5200, China). The grey value of protein bands was quantified by ImageJ program. All employed antibodies were obtained from Proteintech Group, China.
Apoptosis Analysis
The apoptosis of NRK-52E renal tubular epithelial cells was assessed via the Annexin V-FITC/PI Apoptosis Detection kit (Beyotime, China), as per kit directions. Briefly, cells were collected and stained with Annexin V-FITC for 10 min in binding buffer, and then PI was added 10 min prior to testing. The apoptosis rate of cells was determined using Guava EasyCyte Flow Cytometer (Merck Millipore, Germany).
Statistical Analysis
The presented data are mean ± standard deviation (SD) of 3 separate experiments, unless otherwise specified. One-way ANOVA and Tukey’s post hoc test were employed for the determination of multiple group differences. GraphPad Prism Software version 8.0 (GraphPad Software Inc., USA) was employed for statistical analyses. p < 0.05 was the threshold for significance.
Results
PGF Treatment Attenuates Fasting BG Levels and BW
To confirm whether PGF can alleviate typical symptoms in DN mice, we first measured fast BG levels and BW. Relative to the healthy NC mice, all STZ-induced mice displayed an apparent increase in BG levels at week 0, suggesting the successful establishment of the DN model (Fig. 1a). Then, the BG levels in DN mice remained persistently high in the subsequent weeks, while PGF treatment slowly decreased the BG levels in all SZT-treated DN mice. Moreover, compared with the DN group, administration of medium-dose PGF (300 mg/kg) and high-dose PGF (500 mg/kg) significantly lowered the BG levels of mice. The BW in the NC group continuously increased, while it gradually decreased in DN mice (Fig. 1b). Relative to DN mice, treatment with PGF (500 mg/kg) significantly increased the BW of mice. However, no obvious differences were observed in BW between the PGF300 (or PGHF100) and DN groups. Collectively, these data demonstrated that high-dose PGF treatment could effectively attenuate BG levels and improve BW in STZ-treated DN mice.

The physiological outcomes of PGF in DN mice. a Levels of fasting blood glucose; b body weight. n = 8 per group. Data presented as mean ± SD. ***P < 0.001; ****P < 0.0001 vs. NC mice, #P < 0.05; ##P < 0.01; ###P < 0.001 vs. DN mice
PGF Treatment Improves the Tolerance of Glucose and Insulin in DN Mice
To assess the effect of PGF on the regulation of glucose and insulin intolerance, we performed glucose and insulin tolerance test in DN mice after 8 weeks of administration. The BG levels of mice in all groups were elevated immediately after administration of 25% glucose solution, and then began to decrease after 15 min, while this downward trend was not obvious in the DN group (Fig. 2a). Notably, when compared with DN group, the BG levels of DN mice treated with PGF (500 mg/kg) were significantly reduced. Additionally, the BG levels of healthy mice in the NC group after insulin injection decreased immediately, and then slowly increased to a normal level, while those of the DN group exhibited a persistent and dramatic decrease (Fig. 2b). As expected, the BG levels of DN mice administered PGF (500 mg/kg) gradually stabilized after 60 min. All these results indicated that PGF effectively regulates glucose and insulin intolerance.

Evaluation of the PGF-induced glucose and insulin tolerance in DN mice. a Oral glucose tolerance; b insulin tolerance. n = 8 per group. Data presented as mean ± SD. ***P < 0.001; ****P < 0.0001 vs. NC mice, #P < 0.05; ###P < 0.001 vs. DN mice
PGF Treatment Ameliorates Renal Function and Attenuates Renal Inflammatory Responses in DN Mice
Biochemical indexes related to renal function were measured to evaluate the PGF-mediated renal protection in DN mice. Relative to the NC mice, the levels of 24 h albuminuria (Fig. 3a), BUN (Fig. 3b), and Scr (Fig. 3c) in STZ-induced DN mice were significantly increased, which indicated that STZ injection impaired renal function. However, treatment with PGF (500 mg/kg) significantly decreased the levels of 24-h albuminuria, BUN, and Scr compared with the DN group. Therefore, PGF treatment effectively improves renal function in DN mice. To assess whether PGF treatment inhibits inflammatory reactions in DN mice, the levels of inflammatory cytokines and fibrosis factors were evaluated. The results revealed that the levels of IL-6 (Fig. 3d), IL-1β (Fig. 3e), TNF-α (Fig. 3f), and TGF-β1 (Fig. 3g) in DN mice were markedly higher, compared to the NC mice. However, 500 mg/kg and 300 mg/kg PGF exposure significantly decreased IL-6, IL-1β, TGF-β1, and TNF-α levels. Treatment with low dose PGF (100 mg/kg) did not alter the level of inflammatory cytokines. In conclusion, PGF treatment can effectively attenuate renal inflammatory responses in STZ-treated DN mice.

Evaluation of PGF-mediated renal protection and renal inflammatory response in DN mice. Levels of a 24-h albuminuria, b BUN, c Scr, d IL-6, e IL-1β, f TNF-α, g TGF-β1. n = 8 per group. Data presented as mean ± SD. ****P < 0.0001 vs. NC mice; #P < 0.05; ##P < 0.01; ###P < 0.001 vs. DN mice
PGF Treatment Decreased HG-Driven Apoptosis in NRK-52E Cells
To assess the renal protective effect of PGF in vitro, we performed apoptosis analysis on HG-induced renal tubular epithelial cells. The quantity of apoptotic cells was significantly elevated in renal tubular epithelial cells that had undergone HG induction for 24 h (14.53% in the HG group vs. 3.65% in the NC group: Fig. 4a, b). However, HG-induced apoptosis was inhibited by PGF (12.16% in the PGF 10 group, 10.58% in the PGF 30 group, 7.14% in the PGF 50 group). Cell apoptotic proteins, namely, Bax, cleaved caspase-3, and Bcl-2, were also determined. Based on our results, the pro-apoptotic factor proteins Bax and cleaved caspase-3 were obviously increased through HG stimulation, whereas a significant reversal was seen with PGF treatment (Fig. 4c, d). Consistently, HG induction reduced Bcl-2 levels, and this effect was also significantly reversed by administration of PGF. Therefore, PGF treatment can reduce the HG-induced apoptosis of tubular epithelial cells.

Evaluation of the PGF-mediated cell apoptosis in HG-induced NRK-52E cells. a Levels of apoptosis were determined using flow cytometry. b Apoptosis rate. c Western blotting for the assessment of Bax, Bcl-2, and cleaved caspase-3. d Quantitative analyses of western blot bands. Data presented as mean ± SD. ****P < 0.0001 vs. NC mice; #P < 0.05; ##P < 0.01; ###P < 0.001; ####P < 0.0001 vs. HG mice, n = 3
PGF Treatment Inhibits the TLR4/NF-κB Network in NRK-52E Cells
To investigate whether PGF regulates the TLR4/NF-κB network in HG-induced tubular epithelial cells, we first tested the protein and mRNA expression of TLR4. Based on our data, the TLR4 mRNA and protein expressions were markedly increased compared with those in the NC group, which could be markedly reversed by PGF administration (Fig. 5a, b, c). Furthermore, p65, p-p65, IκBα, and p-IκBα are also key signaling molecules in TLR4/NF-κB pathway. Western blot analysis showed that the ratio of p-IκBα/IκBα and p-p65/p65 (Fig. 5b, c) was significantly increased in HG-treated tubular epithelial cells, while these ratios were markedly decreased by PGF treatment. Collectively, PGF could accurately suppress the TLR4/NF-κB network in HG-treated tubular epithelial cells.

Effects of PGF on the inhibition of the TLR4/NF-κB network in HG-induced NRK-52E cells. a TLR4 mRNA levels. b Western blotting for the expression of TLR4, p-IκBα, IκBα, p-p65, and p65. c Quantitative analyses of western blot bands. Data presented as mean ± SD. ***P < 0.001; ****P < 0.0001 vs. NC cells; #P < 0.05; ###P < 0.001; ####P < 0.0001 vs. HG cells, n = 3
PGF Treatment Attenuates Renal Inflammatory Responses and Apoptosis in HG-Treated NRK-52E Cells by Blocking the TLR4/NF-κB Pathway
We then investigated whether there is an association between the renal protective effect of PGF and TLR4/NF-κB network. NRK-52E cells were stimulated with high glucose to induce inflammatory response, and then treated with 1 μM TAK-242, which is an inhibitor of TLR4 signaling pathway. qRT-PCR data suggested that HG induction markedly enhanced the expression of IL-6 (Fig. 6a), IL-1β (Fig. 6b), TGF-β1 (Fig. 6c), and TNF-α (Fig. 6d) in tubular epithelial cells compared to uninduced cells. However, TAK-242 or PGF treatment significantly decreased the levels of these inflammatory cytokines. Combination treatment with PGF and TAK-242 more effectively suppressed the TLR4/NF-κB network and downregulated the amounts of IL-6, IL-1β, TGF-β1, and TNF-α in HG-induced NRK-52E cells. In addition, flow cytometry analysis showed that TAK-242 or PGF treatment markedly reduced the apoptosis rate in HG-induced NRK-52E cells (Fig. 6e), while combination treatment with PGF and TAK-242 intensified the PGF-mediated antiapoptotic effect. Consistent with the flow cytometry results, the western blot results indicated that TAK-242 not only inhibited apoptosis-related protein expression, including BAX and cleaved caspase-3, but also enhanced the protein expression of Bcl-2 in HG-treated NRK-52E cells, while these effects were stronger in combination treatment with PGF and TAK-242 (Fig. 6f, g). In addition, TAK-242 or PGF treatment significantly decreases the protein expression of TLR4, p-IκBα/IκBα, and p-p65/p65 in HG-induced cells, which indicated that both PGF and TAK-242 blocked the TLR4/NF-κB network (Fig. 6h, i). Collectively, the above research results demonstrated that PGF treatment attenuated the inflammatory response and cell apoptosis by inhibiting the TLR4/NF-κB network.

Effects of TAK-242 and PGF on the inflammatory response and apoptosis in HG-induced NRK-52E cells. The transcript levels of IL-6 (a), IL-1β (b), TGF-β1 (c), and TNF-α (d). e Apoptosis rate. f Western blotting for the expression of Bax, Bcl-2, and cleaved caspase-3. g Quantitative analyses of western blot bands. h Western blotting for the expression of TLR4, p-IκBα, IκBα, p-p65, and p65. i Quantitative analyses of western blot bands. Data presented as mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001, n = 3
Discussion
DN is a severe microvascular complication of diabetes mellitus and is a major cause of death among diabetic patients. The etiology and pathogenesis of DN are still not very clear, and it is generally acknowledged that the disorder of glucose and lipid metabolism may be the major factor in the pathogenesis of DN. In addition, abnormal hemorheology, podocytopathy, glycosylation products, and endothelium oxidative stress injury are also important factors leading to the occurrence and development of diabetic nephropathy [14]. Although DN is difficult to cure once it has developed to an advanced stage, most people restore to optimal health in the early stages of DN if treated promptly and properly. Increasing evidences reveal that renal inflammation responses play a promoting role in the occurrence and progression of DN, and the activation of inflammatory signaling pathways leads to renal injury in DN patients [15]. Established treatments for DN patients include control of glucose, hypertension, and dietary protein intake, but there are few drugs available to reduce inflammation and delay the development of DN. It is imperative to apply more effective therapeutic strategies targeting inflammation to prevent renal injury.
PGF, extracted from Grifola frondosa, has numerous bio-activities, including antitumor, anti-inflammatory, and antiviral activities [16]. Moreover, increasing evidence has demonstrated that PGF possesses hypoglycemic and hypolipidemic activities in obese mice with diabetes [17, 18]. Additionally, PGF also acts as an activator of PPARδ to improve lipid metabolism disorders and glucose intolerance in obese mice [19]. Consistent with these findings, our study indicated that PGF not only effectively attenuated fasting BG levels but also improved glucose and insulin tolerance in STZ-induced DN mice. Recent research showed that biochemical indexes of kidney function, including Scr, BUN, and albuminuria, were all returned to normal level by PGF treatment, confirming their anti-nephropathy activities. These effects of PGF in STZ-induced diabetic rats could be achieved by modulating oxidative stress [20]. However, excess oxidative stress may induce the nuclear translocation of NF-κB by phosphorylating IκB [21]. Therefore, our study supplemented the mechanism of PGF in the treatment of diabetic nephropathy. Once renal injury occurs, biochemical markers, including albuminuria, BUN, and Scr, are immediately elevated [22, 23]. Administration of PGF ameliorates renal dysfunction since biochemical markers recover to the normal range during treatment. The inflammatory response affects kidney cells and accelerates DN deterioration [24, 25]. Although various inflammatory pathways contribute to DN development, TLR4/NF-κB is one of the most important pathways for alleviating renal inflammation. TLR4 agonist exacerbates renal inflammatory response and fibrosis through the NF-κB signaling pathway [26, 27]. Accumulating studies have shown that administration of a TLR4 antagonist significantly decreases albuminuria, BUN, glomerular hypertrophy, glomerulosclerosis, and tubulointerstitial injury [28]. Moreover, NF-κB, as a nuclear transcription factor widely occurring in cells, is not only associated with cell apoptosis but also regulates the expression of chemokines, growth factor, and cytokines [29]. Reduction of NF-κB expression can improve renal injury, inflammatory response, and BG levels in DN mice [30]. Phosphorylation of IκB and NF-κB p65 subunits regulates the transcriptional activity of NF-κB. High glucose drove serine phosphorylation of the IκB subunit, resulting in the formation of NF-κB dimer and the activation of p65 (31, 32). Our study demonstrated that PGF decreased p-p65 and p-IκBα levels in HG-induced tubular epithelial cells, while there were no significant differences in p65 and IκBα. In addition, administration of PGF- to HG-induced tubular epithelial cells not only inhibited the inflammatory response but also decreased cell apoptosis, which was achieved by the TLR inhibiting 4/NF-κB network. Furthermore, a combination treatment with PGF and TAK-242 had more significant inhibitory effects on the expression of inflammatory cytokines and cell apoptosis than the individual treatment. Therefore, PGF inhibits inflammatory response and improves symptoms of DN via the TLR4/NF-κB pathway.
Conclusions
In conclusion, we demonstrated that PGF can alleviate renal injury, the inflammatory response, and renal tubular epithelial cell apoptosis via inhibition of the TLR4/NF-κB network. Our findings thus provide more accurate information about the clinical value of PGF in the treatment of early DN patients.
Data Availability
Not applicable.
References
Zheng, C., Huang, L., Luo, W., et al. (2019). Inhibition of STAT3 in tubular epithelial cells prevents kidney fibrosis and nephropathy in STZ-induced diabetic mice. Cell Death & Disease, 10(11), 848.
Cai, S., Chen, J., & Li, Y. (2020). Dioscin protects against diabetic nephropathy by inhibiting renal inflammation through TLR4/NF-kappaB pathway in mice. Immunobiology, 225(3), 151941.
Navarro-Gonzalez, J. F., & Mora-Fernandez, C. (2008). The role of inflammatory cytokines in diabetic nephropathy. Journal of the American Society of Nephrology, 19(3), 433–442.
Stewart, A. G., Thomas, B., & Koff, J. (2018). TGF-beta: master regulator of inflammation and fibrosis. Respirology, 23(12), 1096–1097.
Schena, F. P., & Gesualdo, L. (2005). Pathogenetic mechanisms of diabetic nephropathy. Journal of the American Society of Nephrology, 16(Suppl 1), S30-33.
Lu, S. Y., Zhang, H. L., Wei, X. J., et al. (2019). 2-dodecyl-6-methoxycyclohexa-2,5-diene-1,4-dione isolated from Averrhoa carambola L. root ameliorates diabetic nephropathy by inhibiting the TLR4/MyD88/NF-kappa B pathway. Diabetes, Metabolic Syndrome and Obesity: Targets and Therapy, 12, 1355–1363.
Lin, M., Yiu, W. H., Wu, H. J., et al. (2012). Toll-like receptor 4 promotes tubular inflammation in diabetic nephropathy. Journal of the American Society of Nephrology, 23(1), 86–102.
Lee, B. C., Bae, J. T., Pyo, H. B., et al. (2003). Biological activities of the polysaccharides produced from submerged culture of the edible Basidiomycete Grifola frondosa. Enzyme and Microbial Technology, 32(5), 574–581.
Cui, F. J., Tao, W. Y., Xu, Z. H., et al. (2007). Structural analysis of anti-tumor heteropolysaccharide GFPS1b from the cultured mycelia of Grifola frondosa GF9801. Bioresource Technology, 98(2), 395–401.
Fan, Y., Wu, X., Zhang, M., et al. (2011). Physical characteristics and antioxidant effect of polysaccharides extracted by boiling water and enzymolysis from Grifola frondosa. International Journal of Biological Macromolecules, 48(5), 798–803.
Ding, Y., Xiao, C., Wu, Q., et al. (2016). The mechanisms underlying the hypolipidaemic effects of Grifola frondosa in the liver of rats. Frontiers in Microbiology, 7, 1186.
Jiang, T., Wang, L., Ma, A., et al. (2020). The hypoglycemic and renal protective effects of Grifola frondosa polysaccharides in early diabetic nephropathy. Journal of Food Biochemistry, 44(12), e13515.
Liu, H., Takagaki, Y., Kumagai, A., Kanasaki, K., & Koya, D. (2021). The PKM2 activator TEPP-46 suppresses kidney fibrosis via inhibition of the EMT program and aberrant glycolysis associated with suppression of HIF-1alpha accumulation. Journal of Diabetes Investigation, 12(5), 697–709.
Nagy, C., & Einwallner, E. (2018). Study of in vivo glucose metabolism in high-fat diet-fed mice using oral glucose tolerance test (OGTT) and insulin tolerance test (ITT). Journal of Visualized Experiments, 7(131), 56672.
Xu, B. H., Sheng, J., You, Y. K., et al. (2020). Deletion of Smad3 prevents renal fibrosis and inflammation in type 2 diabetic nephropathy. Metabolism, 103, 154013.
He, Y. L., Li, X. L., Hao, C., et al. (2018). Grifola frondosa polysaccharide: a review of antitumor and other biological activity studies in China. Discovery Medicine, 25(138), 159–176.
Guo, W. L., Deng, J. C., Pan, Y. Y., et al. (2020). Hypoglycemic and hypolipidemic activities of Grifola frondosa polysaccharides and their relationships with the modulation of intestinal microflora in diabetic mice induced by high-fat diet and streptozotocin. International Journal of Biological Macromolecules, 153, 1231–1240.
Chen, Y., Liu, D., Wang, D., et al. (2019). Hypoglycemic activity and gut microbiota regulation of a novel polysaccharide from Grifola frondosa in type 2 diabetic mice. Food and Chemical Toxicology, 126, 295–302.
Aoki, H., Hanayama, M., Mori, K., & Sato, R. (2018). Grifola frondosa (Maitake) extract activates PPARdelta and improves glucose intolerance in high-fat diet-induced obese mice. Bioscience, Biotechnology, and Biochemistry, 82(9), 1550–1559.
Kou, L., Du, M., Liu, P., et al. (2019). Anti-diabetic and anti-nephritic activities of grifola frondosa mycelium polysaccharides in diet-streptozotocin-induced diabetic rats via modulation on oxidative stress. Applied Biochemistry and Biotechnology, 187(1), 310–322.
Rashid, S., Nafees, S., Siddiqi, A., et al. (2017). Partial protection by 18beta glycyrrhetinic acid against cisplatin induced oxidative intestinal damage in wistar rats: Possible role of NFkB and caspases. Pharmacological Reports, 69(5), 1007–1013.
Kishore, L., Kaur, N., & Singh, R. (2017). Distinct biomarkers for early diagnosis of diabetic nephropathy. Current Diabetes Review, 13(6), 598–605.
Adler, A. I., Stevens, R. J., Manley, S. E., et al. (2003). Development and progression of nephropathy in type 2 diabetes: the United Kingdom Prospective Diabetes Study (UKPDS 64). Kidney International, 63(1), 225–232.
Bhalla, V., Zhao, B., Azar, K. M., et al. (2013). Racial/ethnic differences in the prevalence of proteinuric and nonproteinuric diabetic kidney disease. Diabetes Care, 36(5), 1215–1221.
Wang, F., Li, R., Zhao, L., Ma, S., & Qin, G. (2020). Resveratrol ameliorates renal damage by inhibiting oxidative stress-mediated apoptosis of podocytes in diabetic nephropathy. European Journal of Pharmacology, 885, 173387.
Rayego-Mateos, S., Morgado-Pascual J. L., Opazo-Rios L., et al. (2020). Pathogenic pathways and therapeutic approaches targeting inflammation in diabetic nephropathy. International Journal of Molecular Sciences, 21(11), 3798.
Lin, M., & Tang, S. C. (2014). Toll-like receptors: sensing and reacting to diabetic injury in the kidney. Nephrology, Dialysis, Transplantation, 29(4), 746–754.
Lin, M., Yiu, W. H., Li, R. X., et al. (2013). The TLR4 antagonist CRX-526 protects against advanced diabetic nephropathy. Kidney International, 83(5), 887–900.
Fernandes, M. H. V., Maggioli, M. F., Otta, J., Joshi, L. R., Lawson, S., & Diel, D. G. (2019). Senecavirus A 3C protease mediates host cell apoptosis late in infection. Frontiers in Immunology, 10, 363.
Kim, J. E., Lee, M. H., Nam, D. H., et al. (2013). Celastrol, an NF-kappaB inhibitor, improves insulin resistance and attenuates renal injury in db/db mice. PLoS ONE, 8(4), e62068.
Manna, P., Das, J., Ghosh, J., & Sil, P. C. (2010). Contribution of type 1 diabetes to rat liver dysfunction and cellular damage via activation of NOS, PARP, I kappa B alpha/NF-kappa B, MAPKs, and mitochondria-dependent pathways: prophylactic role of arjunolic acid. Free Radical Biology and Medicine, 48(11), 1465–1484.
Sun, H. J., Xiong, S. P., Cao, X., et al. (2021). Polysulfide-mediated sulfhydration of SIRT1 prevents diabetic nephropathy by suppressing phosphorylation and acetylation of p65 NF-kappaB and STAT3. Redox Biology, 38, 101813.
Funding
This research was supported by the Innovation Ability Cultivation Project of Zhuhai College of Science and Technology (No. 2020XJCQ020).
Author information
Authors and Affiliations
Contributions
Conceptualization: S. Shen; methodology: M. Zhao; methodology: L. Wang; formal analysis and investigation: Y. Li; resources: S. Huang; supervision and writing: T. Jiang.
Corresponding author
Ethics declarations
Ethics Approval
This study was approved by the ethical committee of Zhuhai College of Science and Technology.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Jiang, T., Shen, S., Wang, L. et al. Grifola frondosa Polysaccharide Ameliorates Early Diabetic Nephropathy by Suppressing the TLR4/NF-κB Pathway. Appl Biochem Biotechnol 194, 4093–4104 (2022). https://doi.org/10.1007/s12010-022-03976-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-022-03976-8