
Abstract
The objective of this work was to study the influence of carob flours from seed germ (G) and from fruit pulp (P) fractions (from 10 to 30 % w/w) on hydration (moisture, water activity, and relaxation time) and rheological properties (farinograph, rheometry, texture) of wheat dough. Bread quality (specific volume (V s), protein content, crumb texture, and moisture) was also analyzed. Farinogram shape of G and P samples were different. Water absorption of control flour (C) was lower than that obtained for carob germ-wheat flour blends, presenting wheat flour the highest development time. Dough hardness and adhesiveness increased with carob flour, mainly with G. Cohesiveness and springiness of C were higher than those observed for wheat-carob dough. Differences in dough microstructure for the different formulations were predicted by a G’ versus G” plot; heterogeneity in polymer morphology was detected with high contents of carob flours. Even though breads with increasing levels of carob flour presented lower V s and higher crumb firmness and chewiness than wheat bread, protein content for G breads was higher, converting this flour in an interesting ingredient for increasing protein nutrition. Protein value was slightly low for breads with pulp carob flour; nevertheless, fiber contribution would be high.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Carob tree (Ceratonia siliqua L., Fabaceae) belongs to the Leguminosae family and is widely cultivated in the Mediterranean area (Yousif and Alghzawi 2000; Dakia et al. 2007). The carob tree is an important component of the Mediterranean vegetation, and its cultivation in marginal and prevailing calcareous soils of the Mediterranean region is important from the environmental and economical point of view (Battle and Tous 1997).
The carob fruit is a coriaceous dark brown pod that contains a sweet rubbery pulp surrounding palatable seeds; it contains two major parts: the pod (80–90 % w/w) and the seeds (10–20 % w/w) (Naghmouchi et al. 2009; Tetik et al. 2011). The whole fruit contains about 50–65 % w/w sugars (more than 30 % sucrose and glucose; fructose and maltose in low proportions), 1–5 % w/w proteins, and 0.2–0.8 % w/w lipids as well as crude fiber (approximately 9–13 % w/w) and significant amounts of minerals (1–6 % w/w, mainly calcium, potassium, magnesium, sodium, phosphorus, and iron) (Avallone et al. 1997; Yousif and Alghzawi 2000; Ozcan et al. 2007), and also a variable amount of tannins (Yousif and Alghzawi 2000; Makris and Kefalas 2004).
When carob pods maturate in the plant, moisture content is variable (10–20 % w/w) depending on harvesting conditions. After harvesting, in carob industry, moisture content is reduced to values around 8–10 % w/w by drying under shelter in dry and ventilated places to avoid rotting. Carob pods are then crushed mechanically using kibbler and separated from the seeds. Carob pods, due to the high content of sugars, have been also used as a sweetener food ingredient in the production of confectionery, beverages, bread, or pastries (Avallone et al. 1997).
Carob seeds are extremely hard and consist of coat (30–33 % w/w) rich in antioxidants (Batista et al. 1996), endosperm (42–46 %), and germ (23–25 %) (Neukom 1988). The processing of the seeds to yield the corresponding endosperm involves the removal of the husks and of the germ fractions, either by chemical or by thermo-mechanical treatment (Ensminger et al. 1994). The seeds are a source of gum (Rizzo et al. 2004); locust bean gum (LBG), also called carob gum or E410 (Hoefler 2004), is obtained from seed endosperm. The 30–40 % of endosperm consists of galactomannan that is a polysaccharide molecule composed of mannose and galactose sugar units. Therefore, this product is mostly used in the food industry (Catarino 1993) as thickener. The germs recovered as by-products of the seed processing for gum production, are mainly used after milling and heat treatment, as animal feed or as human dietetic food (Battle and Tous 1997).
The literature data on chemical composition indicate that the nutritive value of carob seed germ is high, due to the protein content (>50 % w/w), being more than 50 % of these proteins soluble in water (Bengoechea et al. 2008). These authors found for carob seed germ high content of aspartic and glutamic acids and arginine amino acids, being the limiting amino acids sulphured.
The germ oil (approximately 5–8 %) contains high proportion of highly unsaturated fatty acids (iodine value >100) (Maza et al. 1989).
Germ flour, due to its high nutritional value, is used for dietetic supplements for humans (Dakia et al. 2007) or as an ingredient in cereal-derived foods for celiac people (Feillet and Roulland 1998). However, a carob pulp product enriched in insoluble dietary fiber showed promising cholesterol-lowering properties in animal (Pérez-Olleros et al. 1999) as well as human trials (Zunft et al. 2003).
Wheat proteins are deficient in the essential amino acid lysine; also, wheat flour, due to milling process of grain, lacks of high proportion of minerals and vitamins, resulting in a reduction in the nutritional value of the flour. Therefore, supplementation of wheat flour with carob flour could improve the nutritional value of bakery products.
The aims of this work were to study the hydration and rheological behavior of the dough made with wheat flour complemented with carob germ or pulp flours and to evaluate bread quality.
Materials and Methods
Materials
Wheat flour (type 000, Molino Campodónico Ltda., Argentina) (AAC 2014) for breadmaking with 14.3 % w/w of protein and 13.0 % w/w of moisture contents was used. Carob (C. siliqua L.) flours—pulp (P) with 4.9 % w/w and germen (G) with 55.7 % w/w of protein—were utilized. Also, the P contained 7.2 % w/w moisture, 0.57 % w/w lipids, and 47.0 % w/w of total dietary fiber while G presented 8.0 % w/w moisture, 6.81 % w/w lipids, and 9.3 % w/w of total dietary fiber. Other ingredients used were sodium chloride (CELUSAL, Argentina), fresh yeast (CALSA, Argentina), and distilled water. Blends with wheat flour complemented with both carob flours, P or G (10, 20, and 30 % w/w), were prepared. A control (C) sample without carob flour was included.
Methods
Farinograph Assays
The water absorption (W abs), development time (t d), stability (St), and softening degree (DS) was determinate using a 300 g Brebander farinograph (Duisburg, Germany) according to AACC 2000.
Dough Formulation
Each flour blend consisted of wheat flour, NaCl (2 % w/w), carob germ or pulp flour in levels of 10, 20, and 30 % w/w, and fresh yeast 3 % w/w. Solid ingredients were mixed in a planetary small scale kneader (Kenwood Major, Milano, Italy) for 1 min, and the optimum quantity of water (W abs) was added to the solids. The first minute was kneaded at 50 rpm (speed 1) and the rest of the time to 90 rpm (speed 2) until reaching t d. Final dough temperature was 24 ± 1 °C. Dough was laminated four times before leading to rest for 15 min at 25 °C, and then it was covered with a plastic film to avoid water loss. Hydration and rheological assays on dough were performed without fresh yeast.
Hydration Properties of Dough
Moisture Content and Water Activity
Moisture content of dough was determined according to AACC Method 44-19 (AACC 2000) using a stove (San Jor, Buenos Aires, Argentina). Water activity was measured with an equipment Meter Aqualab series 3 (Decagon Devices Inc., Washington, USA).
Values correspond to the average of three determinations in both cases.
Molecular Mobility
The molecular mobility of the different dough was analyzed by relaxation assays with NMR Brüker Minispec equipment (Brüker, USA). A portion of dough was placed in glass tubes (10-mm diameter) up to 3-cm height, and tubes were closed to avoid dehydration. 1H spin-spin relaxation times (λ) were measured using the Carr-Purcell-Meiboom-Gill pulse sequence. Assays were performed in quadruplicate. Molecular mobility has been studied in cereal systems using the 1H NMR relaxation technique (Leung et al. 1979; Chen et al. 1997; Ruan et al. 1997). Nuclei are excited for a few miliseconds, and when the pulse stops, they return to ground state emitting a signal. Relaxation curves of the proton (1H) signal intensity versus time have exponential decays and can be fitted with equations having one, two, or more exponential terms. Each term, represented by a relaxation time (λ1, λ2, λ3… λi), can be associated to distinct populations of molecules having different mobility. Species with shorter relaxation times are less mobile (solid-like state) than those with longer relaxation times (liquid-like state). The spin echo signal at t = 0 is proportional to the number of hydrogen nuclei of each species. In the present work, the decay curves were fitted to a one-term exponential model according to Eq. 1:
where I(t) represents the 1H signal intensity (proportional to mobile water fraction in the sample), t is the time, λ is the relaxation time (a constant parameter), and A is the signal intensity of protons at t = 0.
Rheological Properties of Dough
Dough without fresh yeast was laminated (1 cm in height) before cutting. Cylindrical dough pieces (diameter = 30 mm) cut with an aluminum cylindrical puncher were obtained for assays.
Dough Texture
Dough texture was analyzed performing a Texture Profile Analysis using a TA.XT2i Texture Analyzer (STABLE MICRO SYSTEMS, Surrey, U.K.) with a load cell of 25 kg and a Texture Expert for Windows version 1.2 Software. Dough samples (15) were utilized. Each sample was subjected to two cycles of compression (deformation = 40 %) with a cylindrical probe (P/75). Force-time curves were obtained at a crosshead speed of 0.5 mm/s. Hardness (Hard), cohesiveness (Cohes), adhesiveness (Adh), and springiness (Sprin) of dough were calculated. Assays were performed in duplicate.
Dough Viscoelasticity
Measurements were performed in a Haake RS600 (Thermoelectron, Karlsruhe, Germany) at 25 ± 0.1 °C, using a plate-plate serrated surface sensor system (35-mm diameter) with 1.5-mm gap between plates. The upper plate was lowered and the excess of sample was trimmed off. The exposed surface was covered with a thin layer of semisolid silicone to prevent moisture loss during testing. Samples were rested for 15 min for relaxing before testing. Frequency sweeps (from 0.005 to 100 Hz) at constant deformation (5 Pa) within the linear viscoelastic range were performed. Mechanical spectra were obtained by recording the dynamic moduli G’ and G” and tan δ as frequency function.
Breadmaking Process
Portions of dough prepared with fresh yeast (50 g) were placed in individual cone aluminum mold (upper width = 65 mm, bottom width = 40 mm, height = 50 mm). Molds were placed for 80 min in a fermentation chamber (Brito Hnos, Buenos Aires, Argentina) at 30 °C. Baking was then carried out for 23 min at 210 °C in the oven (Ariston, Buenos Aires, Argentina). After baking, breads were removed from the pans and cooled to room temperature (2 h) before testing.
Bread Quality Evaluation
Specific Volume
Five breads of each formula were analyzed. Volume of loaves was evaluated through the rapeseed displacement method. The specific volumes (ratio between volume and weight) were calculated.
Moisture and Protein Content
Crumb moisture and protein content of bread were determined according to AACC Method (AACC 2000). Values obtained were the mean of three replicates.
Molecular Mobility
The molecular mobility of the different bread crumb was analyzed by relaxation assays in a similar way to that performed in dough.
Crumb Image
Images of crumbs (10 × 10 cm2) were acquired at 350 dpi with a HP scanner 4070 model using a digital image analysis system.
Texture Properties
Texture Profile Analysis of bread crumbs was performed using a TA.XT2i Texture Analyzer (STABLE MICRO SYSTEMS, Surrey, UK) with a load cell of 25 kg. Middle bread slices of 20-mm thickness underwent a double compression cycle (two-bite texture profile) up to 40 % deformation of its original height with a disc probe (SMS/35). Eight replicates were analyzed for each kind of bread. Slices were compressed two times, and firmness, cohesiveness, springiness, and chewiness were obtained as textural parameters.
Statistical Analysis
Results were analyzed using Statgraphics Plus for Windows 5.1 software. Fisher’s least significant differences test (p < 0.05) was used to define differences between means. Mean and standard deviation were calculated for each parameter.
Results and Discussion
Dough Farinograph Assays
Farinograms of the different formulations are shown in Fig. 1. The farinogram shape (Fig. 1a) of blends with 10 % w/w of germ flour (G10) was similar to that obtained for control wheat flour (data not shown). Formulations with 20 and 30 % w/w (G20 and G30) presented the first peak more pronounced than the second one (Fig. 1b, c). Nevertheless, some differences were observed; G20 unlike G30 did not develop the second peak. Instead, farinogram with P presented a defined and pronounced second peak (Fig. 1d–f), a behavior that could be attributed to the presence of high content of fiber in carob pulp flour. Proteins and fiber contained in G and P, respectively, interact with water in a distinct way, leading to dough with different consistencies. Salinas and Puppo (2014) found changes in the farinogram shape for wheat flour with 12 % w/w inulin as fiber. These authors proposed that these changes in farinogram pattern would be related to the type of gluten structure that is formed during mixing in the presence of fiber and also to the nature of fiber–water interaction in dough. Miś et al. (2012) studied the effect of carob fiber (0, 1, 2, 3, 4, and 5 % w/w, wheat flour basis) and oat whole meal (0, 5, 10, 15, 20, and 25 % w/w, flour basis) on the rheological properties of wheat flour dough, through farinograph and extensograph assays. They found that the farinograph of wheat-carob blends presented two peaks, the second peak being lower than the first peak, an opposite behavior comparing to our results, probably due to the low quantity of carob fiber used by these authors. In the case of oat, as in our case, these authors observed a second peak higher than the first peak. They attributed this behavior to the velocity of the hydration process of dough components, suggesting lower hydration rate in the presences of high amounts of fiber.

Farinograph curves of wheat flour supplemented with a 10 % w/w, b 20 % w/w, and c 30 % w/w of germ carob flour and with d 10 % w/w, e 20 % w/w, and f 30 % w/w of pulp carob flour. W abs, water absorption; t d, development time; St, stability; DS, softening degree
W abs of control (C) was 60.3 (%) and this sample presented the highest t d (19 min) compared with values obtained for carob-wheat flour blends. Systems with carob germ and carob pulp presented higher and lower W abs than control sample, respectively. These results suggest that interaction of carob proteins of germ flour with wheat components would lead to high water absorption due to water-protein interactions. Insoluble fiber absorb less quantity of water; therefore, up to 20 % W abs decreased. Nevertheless, high amount of carob pulp flour incorporate fiber and also proteins to the mix, leading probably to a water absorption value similar to that observed for control sample.
An increase in G content resulted in higher W abs and the dough took more time to reach the optimal consistency.
Stability (St) and softening degree (DS) of control sample were 29 min and 40 FU, respectively. In comparison, all samples with carob flours presented lower St and higher DS. Comparing to low levels of G (≤20 % w/w), a 30 % w/w quantity developed dough at longer time and was more stable with less softening degree. Despite that there is an effect of dilution of wheat gluten, the incorporation of lipids and mainly carob proteins with G30, predominantly globulins (Bengoechea et al. 2008), leads to the stabilization of dough matrix. It is evident that some types of carob protein-wheat protein, carob protein-water, and lipid-protein interactions are being established.
W abs values of blends with P were slightly lower than value obtained for C. Also, W abs and t d slightly increased with the increment of P. Comparing to samples with 10 and 20 % w/w of P, dough with relatively high stability and less degree of softening was obtained with 30 % w/w of wheat flour replacement (P30).
Hydration Properties of Dough
Dough with G and P showed higher and lower moisture content than C, respectively (Table 1). This behavior is in accordance with W abs; dough with greater W abs presented higher moisture values.
No significant differences in water activity (a w) values between dough prepared with G and C dough were obtained. In contrast, dough prepared with addition of P showed significant differences in a w when using 30 % w/w of P (P30), and dough presented the greatest a w (Table 1) suggesting that water is in a more available state. In addition, a w tendency for P was the same as those observed for water absorption and dough moisture content.
Results suggest distinct interactions between components of wheat and carob flours with water. Different compositions of flours could result in mixtures that require different quantity of water and different times to develop dough. Germ flour has higher protein content than pulp flour; these globular proteins may have greater affinity for water than components of pulp flour, the last one containing high amounts of insoluble compounds belonging to the outermost layers of the pod (i.e., fiber). Leung et al. (1979) reported for dough a double-exponential decay assigning two different mobile water fractions, although other authors (Lopes-da-Silva et al. 2007) have applied a simple exponential decay equation for modeling dough NMR relaxation curves. In dough, λ is a parameter related to the water mobility of the system. Higher λ values indicate higher molecular mobility, where water is more loosely linked to the other molecules and consequently in a high energy state. Species with longer relaxation times seem to be in a liquid-like state, a phenomenon that is also associated to the formation of a more viscous network.
This phenomenon depends on the molecular structure of dough components; for example, molecular mobility of water in dough was found to be affected by interactions of inulin and calcium with water (Salinas et al. 2012).
Values of λ of wheat-carob dough were higher than those of C dough, suggesting less structured dough, mainly in the case of germ flour. This behavior could be due to the presence of carob flour components that would interfere with an adequate formation of gluten network by two mechanisms, directly changing dough structure or through the phenomenon of dilution of gluten proteins (Wang et al. 2002).
Species with shorter relaxation times (λ) are less mobile and related with a solid-like state. When germ flour was used, an increment in G at a level of 30 % w/w produced dough with minor value, suggesting that a more structured dough was formed, coincident with the high stability and low softening degree obtained by farinographic assays. This behavior could be attributed to the important role in the interaction of carob globular proteins with water.
Rheological Properties of Dough
The effects of carob flours on the rheological properties of wheat flour dough are summarized in Table 1. The hardness and adhesiveness of dough increased with the addition of carob flour, with a more pronounced effect when G was used. It is important to mark out the behavior of G30 sample; this dough presented significantly higher Hard, Adh, and Sprin than G10 and G20 dough. Values were also higher than those obtained for P30, suggesting the evident effect of carob proteins on structuring dough matrix.
Flour blends absorbed more quantity of water in the presence of carob germ flour; however, the respective dough became harder. Miś et al. (2012) proposed the establishment of a great interaction between gluten and carob proteins that counteracts the effect of network weakening due to gluten dilution. In addition, these flours contain gums (galactomanans) that contribute to dough structure. Hydrocolloids due to their hydrophilic nature, as in the case of proteins, can also absorb high amounts of water, improving dough texture (Rizzo et al. 2004).
Cohes and Sprin of C were higher than those observed for wheat-carob dough, and the elastic component G' of C dough was the lowest. Only germ carob flours produced significant changes in viscoelasticity (tan δ = G”/G’) of dough. G30 presented the highest G' value leading to more elastic dough as it can be deduced from tan δ (tan δ = 0.25).
Mechanical spectra of all dough showed a typical response of a cross-linked polymer network with predominant solid-like behavior, that is, G’ > G” in all frequency range, belonging to viscoelastic network response.
Another form of analyzing mechanical spectra (G’ and G” vs. frequency) of dough is evaluating the dependence between G” and G” as it was stated in Fig. 2. Slopes of the G' versus G” curves have been used as an indicator of change in morphology of polymer blends (Ahmed et al. 2013). Superimposed curves indicate no differences in the morphology of polymers while non-superimposed behavior suggests the formation of rheological heterogeneous matrix. Shift of curves from G” toward G’ corresponding to a high slope, refers to a more elastic network.

Elastic modulus G’ as function of viscous modulus G” for wheat-carob flour dough supplemented with a germ carob (G) and b pulp carob (P) flours
All curves presented almost the same slope (Fig. 2). Two groups for both types of carob flours were detected. Curves of C and G10 were slightly below the G20 and G30 curves and superposed each other (Fig. 2a). Therefore, when 20 or 30 % of G was utilized, the gluten network was not morphologically affected, forming a homogeneous matrix (overlapped curves) suggesting a compatibility between the two source of protein.
However, P presented a different behavior: curves C, P10, and P20 were superposed over the red line of 45° (Fig. 2b). On the other hand, although tan δ of P30 was similar to C, curve of this dough fitted the red line under the other curves, suggesting that a heterogeneous matrix was formed, probably due to the presence of high amount of fiber.
Bread Quality
The specific volume (V s) of control bread was the highest (2.50 cm3/g). With the increment of G, the V s of breads gradually diminished. Plaut et al. (1953) observed a decrease in volume in breads which included carob germ, when they were compared with wheat bread. Instead, breads with P presented also lower V s values than C, independently of the quantity of pulp flour used (Table 2).
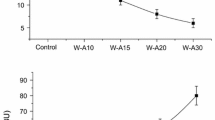
While breads with G were smaller, they presented higher protein content with values ranging from 7.27 to 12.73 % w/w (Table 2). Furthermore, the protein from carob germ has good nutritional value with a complementary amino acid profile to that of wheat protein, with a high content of glutamic acid, aspartic acid, and arginine (Bengoechea et al. 2008). Therefore, this flour is useful for use as food ingredient for athletes’ diet because it increases muscle mass, collagen synthesis, and production of glycogen (Varnier et al. 1995; Flynn et al. 2002). The opposite effect was observed with the addition of P. Protein percentage diminished until 6.35 % w/w with 30 % of P (P30). Results were expected considering the different components of carob flours: the germ flour contains 55.7 % w/w protein, and the pulp flour 47.0 % w/w of total dietary fiber. Another advantage resulting from complementation of wheat flour with carob germ is that the crumbs retained more moisture with higher levels of G, which would contribute to a softer texture. The opposite effect was observed for the crumbs with P, showing generally lower moisture values than the C crumb. In general, the change in moisture of crumb was in accordance with variation of W abs (Table 2).
The relaxation time (λ) of bread crumbs with G was greater than λ of control crumb. A high value of λ is associated with a less structured and less compact crumb. With the increment in G, λ value was significantly decreased, suggesting the formation of a slightly structured matrix, perhaps due to the gelling effect of carob globulin proteins after heating and cooling (Bengoechea et al. 2009). This behavior followed the same tendency observed with farinograph stability and molecular mobility of dough.
On the other hand, crumb with P presented lower crumb λ and an opposite behavior of that observed for dough λ, although no significant differences with level of P was detected.
Figure 3 shows photographs of slices of different breads. Crumb of control bread presented a nearly white color with a great alveolar area. Breads with G flour were of a yellowish-orange color, while breads with carob pulp flour were brown; the latter case is due to the drying processing of carob pod. An increase in level of carob flour the alveoli became smaller and would have consequences for the crumb firmness. Harder (Fig. 4a), less elastic (Fig. 4b), and more chewable (Fig. 4c) crumbs were obtained with high levels of carob flours. Control crumb was the most cohesive. At low levels (G10), a significant decrease in cohesiveness was observed, which was recovered by the addition of more quantity of G, resulting in more integrated crumb (Fig. 4d) corresponding to the lower molecular mobility at 30 % G (Table 2). Although the specific volume and texture of bread crumbs with G and P were not substantially different, formulations were distinct in appearance and nutritional quality; G provided a good proportion of protein, while P contributed with fewer protein but with high level of dietary fiber.

Slice of breads. Control bread, C; breads with carob germ flour, G10, G20, and G30; breads with carob pulp flour, P10, P20, and P30

Texture parameters of bread with carob flour: a hardness, b springiness, c chewiness, and d cohesiveness. Errors bars: standard deviations. Different letters in the same flour indicate significant differences (p < 0.05)
Conclusions
Hydration properties and rheological behavior of wheat dough were affected according to the fraction type of carob flour used. Carob flour changed farinogram profiles of wheat flour. Incorporation of carob germ flour that contains 56 % w/w of protein at a level of 30 % (G30) produced, comparing to G10 and G20, a pronounced second peak with high water absorption, development time, and dough stability, suggesting the formation of structured and heterogeneous dough network due to carob protein-wheat protein or carob protein-water interactions, supported by the low value of molecular mobility and high value of hardness and elasticity. This dough generated yellowish breads with viable increased protein nutrition claim labeling as the content is 12.73 and 75 % higher than that obtained for wheat bread. Dough structuring was maintained after cooking; crumbs with low molecular mobility, high firmness and cohesiveness, and low springiness were also obtained.
On the other hand, low amounts of carob pulp flour (P ≤20 %) with high content of fiber (47 % w/w) produced the same changes in dough parameters as carob germ flour did, leading to softening of dough, although form of farinogram did not substantially change with this carob pulp flour. Carob pulp flour (P30) formed a heterogeneous laxer dough matrix with high molecular mobility, although less harder and elastic than G30 dough. This dough led to brownish breads with only 13 % less protein content than that observed for wheat bread with a more compact crumb with low molecular mobility and high firmness.
References
AAC (2014). Argentinean Alimentarius Codex. Chapter IX. (Art 661-Res.167) http://www.anmat.gov.ar/alimentos/normativas_alimentos_caa.asp. Acceded in October 2014.
AACC. (2000). American association of cereal chemists (10th ed.). St. Paul: The Association.
Ahmed, J., Almusallam, A. S., Al-Salman, F., AbdulRahman, M. H., & Al-Salem, E. (2013). Rheological properties of water insoluble date fiber incorporated wheat flour dough. LWT--Food Science and Technology, 51(2), 409–416.
Avallone, R., Plessi, M., Baraldi, M., & Monzani, A. (1997). Determination of chemical composition of carob (Ceratonia siliqua): protein, Fat, carbohydrates, and tannins. Journal of Food Composition and Analysis, 10(2), 166–172.
Batista, M. T. Amaral, M. T., & Proença Da Cunha, A. (1996). Carob fruits as a source of natural antioxidants. III International Carob Symposium. Cabanas-Tavira, Portugal.
Battle, I., & Tous, J. (1997). Carob tree. Ceratonia siliqua L. Promoting the conservation and use of underutilized and neglected crops. Rome, Italy: Institute of Plant Genetics and Crop Plant Research, Gatersleben/International Plant Genetic Resources Institute, pp 1–92.
Bengoechea, C., Romero, A., Villanueva, A., Moreno, G., Alaiz, M., Millán, F., Guerrero, A., & Puppo, M. C. (2008). Composition and structure of carob (Ceratonia siliqua L.) germ proteins. Food Chemistry, 107, 675–683.
Bengoechea, C., Molina, S., Guerrero, A., & Puppo, M. C. (2009). Influence of functional properties on microstructure, rheology and texture of carob protein isolate (CPI) gel. International Symposium on Food Rheology and Structure, 630-631. Zurich, Switzerland.
Catarino, F. (1993). The carob tree: an exemplary plant. Naturopa, 73, 14–15.
Chen, P. L., Long, Z., Ruan, R., & Labuza, T. P. (1997). Nuclear magnetic resonance studies of water mobility in bread during storage. Lebensmittel-Wissenschaft und-Technologie, 30, 178–183.
Dakia, P. A., Wathelet, B., & Paquot, M. (2007). Isolation and chemical evaluation of carob (Ceratonia siliqua L.) seed germ. Food Chemistry, 102, 1368–1374.
Ensminger, A. H., Ensminger, M. E., Konlande, J. E., & Robson, J. R. K. (1994). Foods and Nutrition Encyclopedia (2nd ed.). Florida: CRC Press, 1, pp 346–348.
Feillet, P., & Roulland, T. M. (1998). Caroubin: a gluten-like protein isolate from carob bean germ. Cereal Chemistry, 75, 488–492.
Flynn, N. E., Meininger, C. J., Haynes, T. E., & Wu, G. (2002). The metabolic basis of arginine nutrition and pharmacotherapy. Biomedicine Pharmacotherapy, 56, 427–438.
Hoefler, A. C. (2004). Hydrocolloids. In Eagan press handbook series. St. Paul, Minnesota: Eagan Press.
Leung, H. K., Magnuson, J. A., & Bruinsma, B. L. (1979). Pulsed NMR study of water mobility in flour dough. Journal of Food Science, 44, 1408–1411.
Lopes-da-Silva, J. A., Santos, D. M. J., Freitas, A., Brites, C., & Gil, A. M. (2007). Rheological and nuclear magnetic resonance (NMR) study of hydration and heating of undeveloped wheat doughs. Journal of Agricultural and Food Chemistry, 55, 5636–5644.
Makris, D. P., & Kefalas, P. (2004). Carob Pod Polyphenolic antioxidants. Food Technology and Biotechnogy, 42(2), 105–108.
Maza, M. P., Zamora, R., Alaiz, M., Hidalgo, F. J., Millan, F., & Vioque, E. (1989). Carob been germ seed (Ceratonia Siliqua): study of the oil and proteins. Journal of the Science of Food and Agriculture, 46, 495–502.
Miś, A., Grundas, S., Dziki, D., & Laskowski, J. (2012). Use of farinograph measurements for predicting extensograph traits of bread dough enriched with carob fibre and oat wholemeal. Journal of Food Engineering, 108(1), 1–12.
Naghmouchi, S., Khouja, M. L., Romero, A., Tous, J., & Boussaid, M. (2009). Tunisian carob (Ceratonia siliqua L.) populations: Morphological variability of pods and kernel. Scientia Horticulturae—Amsterdam 121, 125–130.
Neukom, H. (1988). Carob bean gum: properties and applications. In: P. Fito & A. Mulet (Ed.), II International Carob Symposium, Valencia, Spain, pp. 551–555.
Ozcan, M. M., Arslan, D., & Gokcalik, H. (2007). Some compositional properties and mineral contents of carob (Ceratonia siliqua) fruit, flour and syrup. International Journal of Food Sciences and Nutrition, 58, 652–658.
Pérez-Olleros, L., Garcia-Cuevas, M., & Ruiz-Roso, B. (1999). Note: influence of pulp and natural carob fiber on some aspects of nutritional utilization and cholesterolemia in rats. Food Science and Technology International, 5(5), 425–430.
Plaut, M., Zelcbuch, B., & Guggebheim, K. (1953). Nutritive and baking properties of carob germ flour. Bulletin of the Research Council of Israel, 3, 129–131.
Rizzo, V., Tomaselli, F., Gentile, A., La Malfa, S., & Maccarone, E. (2004). Rheological properties and sugar composition of locust bean gum from different carob varieties (Ceratonia siliqua L.). Journal of Agricultural and Food Chemistry, 52(26), 7925–7930.
Ruan, R. R., Zou, C., Wadhawan, C., Martinez, B., Chen, P. L., & Addis, P. (1997). Studies of hardness and water mobility of cooked wild rice using nuclear magnetic resonance. Journal of Food Process Preservation, 21, 91–104.
Salinas, M. V., & Puppo, M. C. (2014). Rheological properties of bread dough formulated with wheat flour–organic calcium salts–FOS-enriched inulin systems. Food and Bioprocess Technology, 7(6), 1618–1628.
Salinas, M. V., Zuleta, A., Ronayne, P., & Puppo, M. C. (2012). Wheat flour enriched with calcium and inulin: a study of hydration and rheological properties of dough. Food and Bioprocess Technology, 5(8), 3129–3141.
Tetik, N., Turhan, I., Oziyci, H. R., Gubbuk, H., Karhan, M., & Ercisli, S. (2011). Physical and chemical characterization of Ceratonia siliqua L. germplasm in Turkey. Scientia Horticulturae —Amsterdam, 129, 583–589.
Varnier, M., Leese, G. P., Thompson, J., & Rennie, M. J. (1995). Stimulatory effect of glutamine on glycogen accumulation in human skeletal muscle. American Journal of Physiology Endocrinology and Metabolism, 269, E309–E315.
Wang, J., Rosell, C. M., & de Barber, C. B. (2002). Effect of the addition of different fibres on wheat dough performances and bread quality. Food Chemistry, 79, 221–226.
Yousif, A. K., & Alghzawi, H. M. (2000). Processing and characterization of carob powder. Food Chemistry, 69, 283–287.
Zunft, H. J. F., Lueder, W., Harde, A., Haber, B., Graubaum, H. J., Koebnick, C., & Gruenwald, J. (2003). Carob pulp preparation rich in insoluble fibre lowers total and LDL cholesterol in hypercholesterolemic patients. European Journal of Nutrition, 42, 235–242.
Acknowledgments
Authors would like to acknowledge UNLP, CONICET, MINCYT of Argentina, and FCT of Portugal for the financial support (MINCYT-FCT PO928-2009).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Salinas, M.V., Carbas, B., Brites, C. et al. Influence of Different Carob Fruit Flours (Ceratonia siliqua L.) on Wheat Dough Performance and Bread Quality. Food Bioprocess Technol 8, 1561–1570 (2015). https://doi.org/10.1007/s11947-015-1527-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11947-015-1527-7