
Abstract
As human impacts continue to threaten coastal habitats and ecosystems, marine benthic habitat and substrate mapping has become a key component of many conservation and management initiatives. Understanding the composition and extent of marine habitats can inform marine protected area (MPA) planning and monitoring, help identify vulnerable or rare habitats and support fisheries management. To support conservation planning in Eastern Canada, we mapped the seafloor of Newman Sound, identifying the benthoscape classes (i.e. discrete biophysical seafloor classes) of this ecologically diverse and unique fjord in Newfoundland and Labrador (NL). Mapping was achieved using multibeam echosounder (MBES) data collected using multiple platforms, seafloor videos and an unsupervised pixel-based classification method. Seven benthoscape classes were identified within the extent of the MBES coverages. Multivariate statistical analyses indicate that two benthoscape classes - mixed boulder and mud - support distinct epifaunal communities, and also capture the changes in benthic community composition between hard/shallow substrates and soft/deep substrates. Our results illustrate how benthoscape maps can inform marine spatial planning and conservation in the Newman Sound region, support monitoring and also calls for adaptive management of the adjacent Eastport MPA.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Coastal areas are among the most highly threatened and vulnerable regions of the world (Waycott et al. 2009; Halpern et al. 2015; Li et al. 2018). Climate change, natural resource extraction, coastal development and land-based pollution are just a few of many stressors that can lead to habitat degradation and loss in coastal areas (Halpern et al. 2008; Grech et al. 2011; Lefcheck et al. 2017).
Protecting coastal areas requires knowledge of the habitats and ecosystem characteristics of the region. However, it can be difficult to assess impacts and monitor changes without baseline information on the composition and extent of coastal habitats and ecosystems. As a result, habitat and substrate maps have become central to many coastal conservation and management activities (Brown et al. 2012; Copeland et al. 2013; Buhl-Mortensen et al. 2015; Novaczek et al. 2017a).
Advances in acoustic (sonar) seafloor mapping and sampling techniques have allowed for the production of fine-scale generalized biophysical maps of the seafloor (i.e. benthoscape maps – Brown et al. 2012). Benthoscapes are characterized by dominant substrate types, distinct organisms and/or seabed forms (e.g. ripples, sandwaves; Zajac et al. 2003; Brown et al. 2012). Multibeam echosounders (MBES) offer a survey technique capable of collecting continuous coverage baseline information pertaining to seafloor characteristics (Brown et al. 2011). Seafloor bathymetry measured by MBES can be used to understand the geomorphology of the seafloor through the production of bathymetric derivatives such as slope, rugosity, aspect and curvature (Brown et al. 2011; Lecours et al. 2016). MBES backscatter, a measure of the acoustic signal strength returning from the seafloor, has also proven valuable in understanding seafloor characteristics within a region. Backscatter strength can capture variability in substrate composition, with strong backscatter signals typically representing hard, consolidated substrates such as bedrock, boulder and cobble, while weak signals correspond to soft, muddy substrates (Lurton et al. 2015). Using a combination of bathymetry, bathymetric derivatives and backscatter have proven valuable for predicting substrate and species distribution (Monk et al. 2010; Brown et al. 2011; Lecours et al. 2017).
Recording backscatter intensity is complex. A combination of factors, including system-specific settings and environmental variability can result in backscatter intensity values that are not typically calibrated across surveys (see Lurton et al. 2015, Lacharité et al. 2017 and Brown et al. 2019 for comprehensive discussions of backscatter measurements and challenges). This can present further challenges in terms of extrapolating substrate and habitat predictions across multiple MBES coverages. However, novel methods have been developed and refined for producing seamless biophysical seafloor maps using multiple MBES coverages from an area (Lacharité et al. 2017).
Benthoscape maps and species-specific habitat maps produced using these techniques can inform marine conservation and marine spatial planning activities (Young and Carr 2015; Novaczek et al. 2017b). Specifically, these maps can help identify vulnerable or threatened habitats (Rengstorf et al. 2013), support and monitor restoration activities (Walker and Alford 2016) and inform fisheries management (Brown et al. 2012; Smith et al. 2017; Walton et al. 2017). Identifying the extent, location and composition of seafloor features and biodiversity in the form of habitat maps also provides important baseline information for assessing change, particularly as impacts from climate change and anthropogenic activities continue to threaten the resilience of coastal areas.
Here, we used MBES bathymetry, backscatter and derivatives from two uncalibrated sources to create a benthoscape map of a coastal fjord with high conservation potential. We then discuss how the map provides important baseline information to support conservation planning in the region.
Newman Sound is a fjord located in Bonavista Bay on the northeast coast of the island of Newfoundland, Canada. The western part of the fjord is located within the boundaries of Terra Nova National Park, one of two national (i.e. federal) parks on the island of Newfoundland. Due to the high diversity and abundance of ecologically unique areas, including tidal flats, eelgrass and rhodolith beds, and species rich submerged fjord walls, Newman Sound has been listed in an expert process as a ‘special marine area’ (CPAWS-NL 2017). ‘Special marine areas’ recognize special or representative marine features identified during a series of workshops where scientists, provincial and federal government and community experts together identified areas in the province of Newfoundland and Labrador of higher conservation value. The inner basin of Newman Sound, makes up approximately 70% of the Terra Nova Migratory Bird Sanctuary (11.8 km2), and provides important habitat for a high diversity of migratory shorebirds, seabirds and resident waterfowl (Environment and Climate Change Canada 2017). Newman Sound also supports sizeable eelgrass (Zostera marina) beds, which serve as important refuge and nursery areas for juvenile Atlantic cod (Gadus morhua) (Linehan et al. 2001; Cote et al. 2004; Rao et al. 2014) and other fish and invertebrate species (Joseph et al. 2013; Cote et al. 2013).
The Round Island closure, one of two closed areas of the Eastport Marine Protected Area (MPA), is located in Newman Sound (Fig. 1), adjacent to the area mapped in this study. Eastport is one of two Fisheries and Oceans Canada (DFO) Oceans Act MPAs in the province of Newfoundland and Labrador. This small (2.1 km2) no-take MPA was established in 2005, based on a voluntary fishing closure initiated by the local community in 1997. The primary goal of the Eastport MPA is to protect American lobster (Homarus americanus), a species fished commercially in the region. Reports of high lobster catches informed MPA boundary placement, however no habitat mapping or biodiversity surveys were done prior to MPA establishment. Recent habitat mapping and characterization within the boundaries of the MPA have determined that the MPA does little to conserve habitats and biodiversity representative of the broader region of Newman Sound (Novaczek et al. 2017a). While the Eastport MPA was primarily designed to help protect a single species, its small size offers limited protection of ecologically diverse and unique areas in Newman Sound. A recent ecosystem goods and services study used biological indicators (estimates of lobster abundance; catch per unit effort; estimates of berried females; size distribution) to examine the impact of the MPA on the local lobster fishery (Lewis et al. 2017). The study determined that the Eastport MPA has little to no effect on the enhancement of the local fishery as the majority of biological indicators show no significant improvement when compared to regional scale patterns. The small size of the Eastport MPA is frequently cited as limiting the fisheries enhancement and biodiversity conservation benefits of the MPA (Lewis et al. 2017; Novaczek et al. 2017a; Stanley et al. 2018), leading to calls for adaptive management that could result in expanding or changing the MPA boundaries to include more diverse habitats (Novaczek et al. 2017a). Adaptive management allows regulations and boundaries to be improved as new data and knowledge is gathered and analyzed and as ecological systems change through time (Wilhere 2002). While identifying the appropriate MPA size is complex, general guidelines recommend that if an MPA is intended to conserve biodiversity and support climate change resilience it should be moderate to large in size, ideally 4–20 km across (Green et al. 2014).

Newman Sound, Newfoundland and Labrador
The benthoscape map of Newman Sound presented here provides important baseline information that can support the calls for the potential adaptive management of the Eastport MPA to include representative biodiversity and unique and vulnerable habitats of the wider geographic region. We illustrate scenarios for extending the boundaries of the Round Island closure into the area mapped in this study and the resulting increases in protection of benthoscape classes. The benthoscape map can also be used to identify variability in benthoscape pattern and benthic biodiversity across a range of spatial scales. This information can allow researchers to investigate how seafloor patterns can be most accurately used as surrogates for biodiversity in the region. Additionally, coastal seafloor maps such as the one created in this study can support future research questions and policy development related to habitat limitations for species in response to climate change and for identifying critical nursery, spawning and potential settlement areas for threatened or at-risk species.
Methods
Study area
Newman Sound (Fig. 1) is one of several deep fjords in the region and is separated from Bonavista Bay by a shallow sill (Cumming et al. 1992). The maximum depth of the outer basin is 349 m, while the inner basin has a maximum depth of 63 m. Several rivers and streams flow into the inner basin of Newman Sound where mudflats and estuarine vegetation are common. The adjacent Terra Nova National Park protects 400km2 of sheltered inlets, islands, forest and bog habitat that supports a variety of terrestrial mammals including the endemic Newfoundland marten (Martes americana atrata; Parks Canada 2018).
Multibeam Echosounder surveys
In 2003, an acoustic multibeam echosounder (MBES) survey was conducted in Newman Sound by the Canadian Hydrographic Service (CHS; Fig. 2). Bathymetric and backscatter data were collected using two Kongsberg MBES systems: an EM1002 (95 kHz) system operated aboard the CCGS Matthew, and an EM3000 (300 kHz) system operated aboard the CCGS Plover. The EM1002 system primarily surveyed the deeper regions of the fjord, while the EM3000 system surveyed the shallow and narrow region separating the inner and outer basins, the head of the fjord and the outer margins of the inner basin (Copeland 2006). Bathymetric data were post-processed by the Geological Survey of Canada to generate a 10 m resolution surface for the region. Uncalibrated raw backscatter data (.all format) from the two MBES systems were post-processed independently for this study in QPS FMGT software v7.7.8 using default settings and Adaptive AVG algorithm that reduces noise in terrains with significant slope variation. Both MBES backscatter coverages were rendered as 10 m resolution mosaics (Fig. 3).

Bathymetry data collected during EM1002 (A) and EM3000 (B) multibeam echosounder surveys. Inset maps illustrate areas with interesting bathymetric features: a large change in depth indicating a high slope region of the fjord walls (A) and The Narrows region separating the inner and outer basins of Newman Sound (B)

Backscatter data collected during EM1002 (A) and EM3000 (B) multibeam echosounder surveys. Inset maps illustrate areas with interesting backscatter features: patches of shallow seafloor substrate with low intensity backscatter signals surrounded by relatively high intensity backscatter signals (A) and a region where backscatter intensity abruptly transitions from strong to weak (B)
Benthic ground truthing video surveys
Benthic video surveys were conducted from October 3-13th, 2016 from a 40 ft. inshore fishing vessel, the Jamie Tim N′ Trevor, based out of Happy Adventure, NL. Survey stations were selected using a backscatter stratified random sampling design (N = 28), constrained by the depth range of the camera system (max depth 170 m). Ground truthing efforts focused on the area mapped by the EM1002 system, as extensive archival video data already existed for the shallow region mapped by the EM3000 system (Copeland 2006; Fig. 2B). Although a temporal difference exists between the MBES survey, archived video surveys (2003) and recent video surveys (2016) it is unlikely that the MBES bathymetry, backscatter and derivatives would have changed significantly within the sheltered geographic region at the resolution at which the benthoscape map is generated (10 m).
Video footage was obtained using a custom drop camera system that recorded in both standard and high definition. The camera system was equipped with mounted LED (Light Emitting Diode) lights and two red lasers positioned 5 cm apart to allow for scale measurements. Camera and vessel positions were recorded using a handheld Garmin eTrex10 WAAS (Wide Area Augmentation System) enabled GPS (Global Positioning System), while video transects ran for approximately 4 min as the vessel drifted. Positional accuracy for the WAAS enabled GPS was <3 m (Garmin Ltd. 2005). High definition seafloor video footage was recorded with a downward facing GoPro Hero 3 Black Edition, and an approximate 1 m distance between the camera and the seafloor was maintained through an on-board live feed from a downward facing standard definition 200 m-tethered Deep Blue Pro camera.
Still images were extracted from the high definition GoPro footage at 2 s intervals with each transect yielding between 2 and 56 usable images. Images were deemed unusable if they had an obstructed field of view (e.g., disturbed fine sediment), lacked scaling lasers, were poorly illuminated or were blurred. An additional 29 classified standard definition images from a previous habitat mapping study (Copeland 2006) were incorporated into the analysis. In total, 238 images from 56 stations were used to classify the seafloor of Newman Sound. Image location was recorded using the timestamps from the high definition footage and continuous GPS overlay of vessel position. Image area was measured using Image J Software, and substrates were classified based on biophysical characteristics.
Image classification
Seafloor images were classified based on their biophysical characteristics, including the dominant sediment type and the presence/absence of encrusting coralline algae into benthoscape classes. This approach is comparable to the classification of terrestrial landscapes from remote sensing datasets (see Zajac et al. 2003; Zajac 2008; Brown et al. 2012). Each image was classified into one of eight benthoscape classes that captured the range of variability across ground-truthed images (Table 1; Fig. 4). The eight benthoscape classes were: (A) bedrock, (B) deep pebble/cobble, (C) gravelly muddy sand, (D) mixed boulder, (E) mud, (F) shallow pebble/cobble, (G) rhodolith and (H) sand. The classification mirrored that of the ground truthing surveys (Copeland 2006) to ensure that the two datasets were comparable.

Images representing benthoscape classes in Newman Sound: (A) bedrock, (B) deep pebble/cobble (C) gravelly muddy sand, (D) mixed boulder, (E) mud, (F) shallow pebble/cobble, (G) rhodolith, and (H) sand. Scale bar = 5 cm
Unsupervised segmentation of MBES data
Bathymetry, seafloor slope and backscatter have been used extensively in benthic mapping studies due to their continuous nature in terms of map coverage, and also their important role in separating distinct benthic habitats (Brown et al. 2011; Ierodiaconou et al. 2011; Che Hasan et al. 2014; Hill et al. 2014). These variables have been used specifically to map coastal fjord environments (Cochrane et al. 2011; Copeland et al. 2013), with slope being of particular interest in a context where steep walls and current-winnowed gravel have been shown to support unique and diverse benthic communities (Dale et al. 1989). In this study, the two MBES datasets were segmented separately based on bathymetry, backscatter and slope using the ISO Cluster Unsupervised Classification Tool (ArcGIS 10.3.1). The tool segments a series of input raster bands by combining an iterative-self-organizing (ISO) algorithm and maximum likelihood classification. The ISO segmentation was limited to areas within the depth range of the ground-truthed video surveys (≤170 m). This method has been shown to be an effective way of segmenting MBES data (Calvert et al. 2014). The ISO Cluster unsupervised classification tool allows the user to select the number of output classes. The optimal number of classes for both the EM1002 and EM3000 MBES coverages was determined by using the maximum number of classes possible while maintaining at least two ground-truthed images within each class. This was not possible for one class within the EM3000 coverage due to the limited number of samples within the inner basin of Newman Sound.
The locations of classified ground-truthed images were then overlain on the map derived from MBES segmentation and an error matrix was generated to determine the accuracy of the classification. The error matrix compares observed substrate classes (from image analysis) with the classes derived from MBES segmentation. Three standard measures of accuracy were calculated: (1) overall accuracy measures the percentage of correctly classified reference pixels when overlain on the MBES segmentation, (2) benthoscape class-specific measures of accuracy, measuring the probability that a reference pixel is correctly classified (Producer’s Accuracy) and (3) how likely a map user is to encounter a correctly predicted benthoscape (User’s Accuracy) (Diesing et al. 2016). A methodological diagram (Fig. 5) illustrates the general workflow followed for integrating the two MBES datasets and comparing the final Newman Sound benthoscape classification with epifaunal assemblage analysis (described below).

Methodological diagram representing the workflow followed for producing benthoscape map of Newman Sound using two multibeam echosounder datasets and seafloor video surveys
Epifaunal assemblage analysis
The abundance of benthic fauna was also recorded for the entire transect length using continuous video footage, and organisms were identified to the lowest possible taxonomic level. To explore potential relationships between benthoscape class and organism assemblages, multivariate statistical analyses were performed using the software package PRIMER (Plymouth Routines in Multivariate Ecological Research) v7 (Clarke and Gorley 2015). In order to allow for the integration of the archival data, abundance data were presence-absence transformed and combined for the entire transect, using the mid-point to represent each transect. Transect length in all samples in the archived and recent video footage varied (mean transect length = 110 m). Transect length from the archived video data varied between approximately 30 – 500 m while transect length from recent video footage varied between approximately 60 – 200 m. As such, species presence/absence was not standardized by transect length. Nonetheless, incorporating this archival video footage provides insight into the potential relationship between species composition and benthoscape class, although it is likely that species richness and abundance estimates standardized by transect length would more adequately capture these patterns. Transects were excluded from the analysis if they crossed the boundary between two predicted benthoscape classes. A Bray-Curtis Similarity matrix was generated, and non-metric multi-dimensional scaling (nMDS), analysis of similarity (ANOSIM) and similarity percentage (SIMPER) tests were run in PRIMER to assess the potential impact of benthoscape class on benthic community composition.
Results
Unsupervised substrate segmentation of MBES data
Unsupervised classification of the EM1002 MBES coverage resulted in thirteen substrate classes characterized by their depth, slope and backscatter values. The classes derived from the unsupervised segmentation of the EM1002 coverage which covers the majority of the fjord, aligns with what would be expected in terms of the geomorphology of fjord environments: separate classes that encompass flat deep basins, shallow sills and steep fjord walls (Fig. 6A; Syvitski and Shaw 1995).

Results of ISO unsupervised segmentation using bathymetry, backscatter and slope for EM1002 MBES coverage within the depth range of the drop camera system (max depth 170 m) (A) and EM3000 coverage (B). Dashed line indicates region where two coverages overlap
Four substrate classes were identified in the EM3000 coverage, with segmentation revealing a flat, inner basin with more variable substrate classes along the perimeter and at the shallow sill separating the inner and outer regions of the fjord (Fig. 6B).
The error matrix compared the MBES segmentation against the classified images (Table 2). Ground-truthed images obtained where the two MBES coverages overlapped (the narrow region between the inner and outer basins) were assigned to MBES segmentation classes from the EM1002 coverage, as the EM1002 segmentation provided more variability of classes within the overlapping region (Fig. 7).

Locations of ground truthing in region where EM1002 (left) and EM3000 (right) MBES coverages overlap. Colours on each map represent the same unique substrate classes as Fig. 6
The seventeen classes derived from the two MBES coverages were reduced to seven by merging segmented classes based on their dominant in-situ image class. Several benthoscape classifications from the images corresponded closely with a single MBES segmentation class, while others required merging. Images classified as deep pebble/cobble corresponded to MBES segmentation class 7 (50.0% agreement), which was located at the outer edge of regions classified as mud. Gravelly muddy sand (GMS) images corresponded to MBES segmentation class 11 (75.0% agreement). Images classified as mixed boulder corresponded to MBES segmentation classes 12, 15 and 16 (80.0% agreement) and were predominant in shallow water regions towards the head of the fjord. Images classified as rhodolith were common in MBES segmentation classes 15 and 16, and were only found at the narrow region between the inner and outer fjord basins. Rhodolith was not acoustically distinguishable from mixed boulder and without additional ground truthing surveys in other regions we cannot assume that the rhodolith bed is uniform across the extent of classes 12, 15 and 16. Previous studies have, however, identified the presence of rhodoliths on the shallow sill separating the inner and outer basins of the fjord (Copeland 2006). Mud corresponded to MBES segmentation classes 1, 2, 3, 5, 6, 8, 9 and 14 (98.8% agreement) and was found in the flat inner and outer central basins of the fjord. Substrate class 14 was validated by a single archived image (mud) however the low slope (mean = 1.1 degrees) and low (relative, uncalibrated) backscatter return in the substrate class suggests that mud is an appropriate classification. Bedrock corresponded to MBES segmentation classes 4 and 13 (53.3% agreement) and was predominant in deep, high slope regions toward the periphery of the fjord. Shallow pebble/cobble did not clearly correspond to any MBES segmentation class and was acoustically indistinguishable from gravelly muddy sand (GMS). The number of shallow pebble/cobble and GMS images differed by a single image when overlain on MBES segmentation class 10 (shallow pebble/cobble = 11 images; GMS = 12 images). As a result, MBES segmentation class 10 was retained to capture the transition between GMS and more complex mixed boulder substrates. Images classified as sand corresponded to map class 17 (60.0% agreement) and exhibited a patchy distribution (Fig. 6).
The final benthoscape map of Newman Sound waters shallower than 170 m depth (Fig. 8) was produced by combining the two MBES coverages. In the region where the two MBES coverages overlapped, the EM 1002 coverage was used as it provided a greater number of MBES segmentation classes for ground-truthing comparison. The final map indicates that the region is dominated by mud (54.22%) and mixed boulder (12.85%). Areas classified as mixed boulder occurred in some of the shallowest regions of the MBES coverage. Based solely on the results of this mapping project, the amount of mixed boulder in Newman Sound is likely underestimated because the MBES coverage does not extend to shallow regions adjacent to the shoreline.

Final benthoscape map of Newman Sound (depth < 170 m). Bar chart illustrates the percent contribution of each benthoscape class in the mapped area
The greatest amount of confusion occurred between GMS/shallow pebble/cobble and rhodolith/mixed boulder. Images classified as rhodolith and pebble cobble were acoustically indistinguishable from other benthoscape classes, and were thus not included in the final benthoscape map of Newman Sound, except where shallow pebble/cobble and GMS were combined to capture the transition between GMS and more complex substrates. The overall accuracy of the final map after merging the two MBES coverages was 73.1% (Table 2).
Epifaunal assemblage analyses
A number of distinct patterns emerged in the nMDS plot of benthic community data in Newman Sound (n = 55) (Fig. 9). However, because species data for the entire length of each transect had to be merged into a single point to allow for the incorporation of archival data, the sample size was much smaller and may not fully capture the potential impact of benthoscape class on community composition. The nMDS plot does however suggest a gradual shift from soft substrates (mud) on the right side of the ordination plot (Fig. 9) to more complex and consolidated substrates (mixed boulder) on the left.

nMDS ordination plot of epifaunal assemblage data extracted from seafloor video surveys in Newman Sound
ANOSIM results (Table 3) reveal significantly different species assemblages between mixed boulder and mud (R = 0.619, p < 0.001), mixed boulder and bedrock (R = 0.620, p < 0.004) mixed boulder and GMS (R = 0.648, p = 0.003), mixed boulder and shallow pebble /cobble/GMS (R = 0.606, p = 0.003) and mud and sand (R = 0.549, p = 0.001). Due to the low representativity of some benthoscape classes in the multivariate dataset, pairwise ANOSIM results for several benthoscape classes are not valid because the number of possible permutations was too low.
SIMPER results indicate taxa that contribute most towards the similarity of samples within each benthoscape class and the overall average similarity of epifaunal composition within each class (Table 4). Average similarity within each benthoscape class was generally low, ranging from 33% to 56% (Table 4).
In terms of pairs of benthoscape classes with distinct epifaunal assemblages (Table 3), dissimilarity in faunal composition was driven by the presence of several key species. Dissimilarity in epifaunal assemblages in mixed boulder and mud benthoscape classes (average dissimilarity = 78.1%) was primarily driven by plumose anemones (Metridium senile) and northern seastars (Asterias vulgaris), species commonly found on mixed boulder substrates, and epibenthic trachymedusae (Ptychogastria polaris), common on mud substrates. Plumose anemones, snow crab (Chionoecetes opilio) and daisy brittle stars (Ophiopholis aculeata) were the highest contributors to dissimilarity between mixed boulder and GMS faunal compositions (average dissimilarity = 69.3%). Dissimilarity between mixed boulder and bedrock (average dissimilarity = 62.92%) was primarily driven by the presence of daisy brittle stars, and unidentified white anemones. Differences in faunal composition between mixed boulder and shallow pebble/cobble/GMS (average dissimilarity = 63.6%) was driven by plumose anemones, northern seastars and dahlia anemones (Urticina felina). Common sand dollars (Echinarachnius parma), northern seastars and the hydrozoan P. polaris were the highest contributors to dissimilarity between mud and sand benthoscape classes (average dissimilarity = 83.64%).
Discussion
Seven benthoscape classes and two statistically distinct epifaunal assemblages were identified in Newman Sound, NL by integrating two MBES datasets with new and archived seafloor video surveys. The final predicted benthoscape map (Fig. 8) contributes to seafloor mapping efforts in Newfoundland and Labrador, improves our understanding of regional seafloor substrates and habitats in Bonavista Bay and provides information that can support the potential adaptive management of the Eastport MPA.
Epifaunal assemblage analysis
Mud and mixed boulder benthoscape classes supported statistically distinct epifaunal assemblages. However, due to underrepresented benthoscape classes in the multivariate analyses, it is possible that the species rich bedrock walls and deep pebble/cobble benthoscape classes also support distinct communities. Furthermore, species abundances may be more important for detecting patterns and variation in epifaunal assemblage across benthoscape classes as habitat distinction within the boundaries of the MPA was often driven by the abundance of green urchins (Strongleyocentrotus droebachiensis), northern seastars (Asterias vulgaris) and brittle stars (Ophiopholis sp.; (Novaczek et al. 2017a). Transforming data to presence/absence can make detecting patterns in species assemblages difficult, particularly in the case of Newman Sound where generalist species such as green urchins and brittle stars were found across all substrates. However, these analyses do provide insight into the potential relationships between species composition and benthoscape class. ANOSIM results revealed distinct differences in epifaunal assemblages between mixed boulder and mud benthoscape classes (R = 0.619, p < 0.001) – a pattern likely driven by contrasting sediment composition (hard vs. soft substrates) and depth (shallow vs. deep). This pattern, along with the low similarity of epifaunal assemblage composition within each benthoscape class (33–56%) aligns with other studies that aim to classify seafloor habitats based on benthic community assemblages (McGonigle et al. 2009; Brown et al. 2012). While these results fail to capture gradients of epifaunal assemblages in transitional habitats, a higher density sampling effort measuring species abundance could potentially capture this variation.
Mapping for conservation planning
Seafloor mapping has become increasingly important for informing conservation planning. Design, management and monitoring of MPAs and other area-based conservation measures as well as the identification of Ecologically and Biologically Significant Areas (EBSAs) can all be informed by seafloor maps. Recent habitat characterization and mapping activities in the nearby Eastport MPA determined that the MPA does little to conserve habitats and biodiversity representative of the broader region of Newman Sound (Novaczek et al. 2017a). Novaczek et al. (2017a) identified four distinct benthic substrates within the boundaries of the MPA: “shallow rocky,” “sand and cobble” and “boulder/bedrock” and “sand.” Several of the benthoscape classes identified in this study are either under-represented or unrepresented within the boundaries of the Eastport MPA. The MPA is dominated by shallow rocky substrates, which correspond to the ‘mixed boulder’ classification in this study (86.3% of the Round Island closure). The shallow pebble cobble and GMS substrates identified in this study are similar to the “sand and cobble” habitat identified by Novaczek et al. (2017a). No sandy substrate was predicted to be protected within the boundaries of the Round Island closure, while sand/cobble and bedrock features combined made up only a small portion of substrates within the closure (13.7%; Novaczek et al. 2017a). Furthermore, 95.7% of the Round Island closure is shallower than 80 m and within the photic zone (Novaczek et al. 2017a). Consequently, deep-water substrates identified in this study (mud and deep pebble/cobble) are unrepresented.
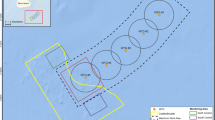
Extending the boundaries of the Round Island closure could help protect nearly all benthoscape classes identified in Newman Sound (Fig. 10). This would increase the protection of benthoscape classes and their associated biodiversity that are under or unrepresented based on the current boundaries of the MPA, and result in a more ecologically representative MPA - a key element of effective conservation planning. Five simple boundary expansion scenarios were tested using buffers of increasing sizes around the existing Round Island MPA closure. The amount (in km2) of each benthoscape class included in each scenario are illustrated in Fig. 10. These incremental boundary expansions are useful for determining the proportion of each benthoscape class that would be included in potential adaptive management scenarios, and how the MPA could better represent benthoscape scale patterns in Newman Sound. In the region, habitats are generally more diverse in shallower waters, and a revised MPA that aims to protect representative habitats would not necessarily have to be very large and could be protect a range of habitats if oriented along contour lines in areas of highest habitat diversity.

Boundary expansion scenarios for the Round Island closure. Bar graph illustrates the amount (km2) of each benthoscape class that would be included in various scenarios. Area measurements only include the region mapped in this study. Current protection levels obtained from Novaczek et al. 2017a
The importance to adaptively manage MPAs has been recognized, as adaptive management allows regulations and boundaries to be improved as new data and knowledge accumulates and as ecological systems change through time (Wilhere 2002). In Canada, however, changing MPA boundaries and regulations proved to be a complex and lengthy process (Morris and Green 2014), making adaptive management of MPAs very rare in practice. In the case of the Eastport MPA, protecting regional biodiversity is not one of the current MPA objectives, as the Eastport MPA was created at a time when many Canadian MPAs focused on single (or few) species, often of commercial interest. Conservation of biodiversity however, has become more important in global conservation initiatives and is now central to Canada’s marine conservation strategy (DFO 2018). This change in context encourages the adoption of adaptive management strategies that could help improve existing Canadian MPAs so they can protect regional biodiversity more efficiently.
Canada committed to protect ecosystems, species and biodiversity through the establishment of an ecologically representative and well-connected MPA network that covers 10% of its coastal and marine areas (DFO 2009). As DFO continues to advance MPA network establishment in its five priority bioregions (DFO 2018), including the ‘Newfoundland and Labrador Shelves’ bioregion, seafloor maps of areas identified as priorities for conservation can help identify habitats that are and are not represented in existing MPAs and MPA networks (Copeland et al. 2013; Novaczek et al. 2017a) as well as future sites to be included in MPA networks. Seafloor maps can also help determine whether habitats are replicated within the reserve or across the reserve network, providing a safeguard against potential disturbances (Young et al. 2017). Newman Sound has already been identified as ‘special marine area’ with high conservation value (CPAWS-NL 2017). As MPA network planning continues across the region, ecologically unique and well-studied coastal areas such as Newman Sound should be considered as future sites for coastal conservation.
The final benthoscape map produced in this study also identifies the location and extent of additional shallow rocky substrate (mixed boulder benthoscape) in Newman Sound that are not included in the boundaries of the Round Island closure. This substrate is commonly associated with juvenile and adult lobster habitat and dominates the MPA (Novaczek et al. 2017a). Increasing the amount of lobster habitat protected within the boundaries of the MPA could potentially result in enhancements to the local fishery by protecting sufficient habitat to potentially support a healthy lobster population in the region.
Although it was not possible to acoustically distinguish rhodolith and mixed boulder benthoscape classes, previous research (Copeland 2006) identified the presence of a rhodolith bed located at the shallow narrow region separating the inner and outer basins of Newman Sound. Rhodolith beds form complex habitats that support a high diversity of invertebrates, fish and algae (Steller et al. 2003; Copeland et al. 2008; Hernandez-Kantun et al. 2017 and references therein) and also provide refuge and feeding grounds for commercially important species such as juvenile Atlantic cod (Kamenos et al. 2004) and juvenile Icelandic scallop (Copeland 2006). Rhodolith beds are sensitive to bottom contact fishing activities (Hall-Spencer and Moore 2000; Kamenos et al. 2003), waste build-up from aquaculture operations (Hall-Spencer et al. 2006) and are particularly vulnerable to impacts from ocean acidification (Martin and Hall-Spencer 2017 and references therein). The rhodolith bed in Newman Sound is also in close proximity to sizeable eelgrass (Zostera marina) beds located in the inner sound (Rao et al. 2014). However, neither eelgrass nor rhodolith beds are protected by the Eastport MPA despite their importance in providing habitat and refuge for a wide range of species.
Newman Sound is an ecologically unique and well-studied region in Newfoundland and Labrador. However, the Eastport MPA protects very little of its diversity despite our knowledge of the importance of conserving representative biodiversity, benthoscape classes and unique habitats such as eelgrass and rhodolith beds. The small size of the Eastport MPA is a clear limitation. However, the benthoscape map produced in this study coupled with extensive research on eelgrass beds in the region and our knowledge of the location of the rhodolith bed provide valuable information that can inform the potential adaptive management and expansion of the MPA.
Mapping to support future research activities in Newman sound
A recent study in Newman Sound used nearshore baited cameras to examine how seabed habitat influences fish, Atlantic rock crab and American lobster community composition (Dalley et al. 2017). The authors identified four common nearshore substrates: bedrock (high relief boulder and bedrock outcrops), sand-pebble, cobble and eelgrass, in addition to one anthropogenic habitat (wharf). Significant differences in species diversity and relative abundance were observed between sand-pebble and bedrock substrates. Dissimilarity was driven by a variety of fish species, including cunner (Tautogolabrus adspersus), age-1 Greenland cod (Gadus microcephalus ogac), winter flounder (Psuedopleuronectes americanus) and Atlantic cod (G. morhua). American lobsters were observed primarily on bedrock substrates, where Atlantic rock crabs were absent (Dalley et al. 2017). The baited cameras used by Dalley et al. (2017) were limited to the depth at which complementary beach seining sampling was done (max depth 3 m), which is shallower than the MBES data used in this study. Although it is difficult to compare the results of baited vs. non-baited sampling methods, the final benthoscape map produced in this study could inform future studies that examine how fish communities change in response to substrate beyond the near-shore environment.
The Atlantic cod population in Newman Sound has been extensively studied in terms of habitat use and movement (Cote et al. 2001, 2004), predator-prey dynamics (Linehan et al. 2001; Gorman et al. 2009), and relationships between cod density and eelgrass patchiness (Thistle et al. 2010). This research was almost exclusively done in shallow regions of the inner basin of Newman Sound. As such, our understanding of cod movement and habitat associations in deeper regions of Newman Sound is limited. The final Newman Sound benthoscape map provides valuable information that can inform future studies on Atlantic cod movement and variability in seasonal and life-stage habitat use.
This benthoscape map provides important baseline information for management and monitoring activities related to the Eastport MPA. An additional objective of this study was to demonstrate the application of a seafloor mapping method that combines MBES backscatter data from two sources, a pixel based unsupervised classification and seafloor image analysis. Methods for integrating MBES backscatter from multiple sources and incorporating archival data are valuable, particularly due to high costs associated with at-sea surveying. Maps that describe seafloor substrates and habitats are crucial for effective marine conservation and management, and can facilitate implementation, design and management of marine conservation objectives, particularly in coastal environments where anthropogenic stressors and threats are concentrated.
References
Brown CJ, Smith SJ, Lawton P, Anderson JT (2011) Benthic habitat mapping: a review of progress towards improved understanding of the spatial ecology of the seafloor using acoustic techniques. Estuar Coast Shelf Sci 92(3):502–520
Brown CJ, Sameoto JA, Smith SJ (2012) Multiple methods, maps, and management applications: purpose made seafloor maps in support of ocean management. J Sea Res 72:1–13
Brown CJ, Beaudoin J, Brissette M, Gazzola V (2019) Multispectral multibeam Echo sounder backscatter as a tool for improved seafloor characterization. Geosciences 9(3):126
Buhl-Mortensen L, Buhl-Mortensen P, Dolan MJF, Gonzalez-Mirelis G (2015) Habitat mapping as a tool for conservation and sustainable use of marine resources: some perspectives from the MAREANO programme, Norway. J Sea Res 100:46–61
Calvert J, Strong JA, Service M, McGonigle C, Quinn R (2014) An evaluation of supervised and unsupervised classification techniques for marine benthic habitat mapping using multibeam echosounder data. ICES J Mar Sci 72(5):1498–1513
Che Hasan R, Ierodiaconou D, Laurenson L, Schimel A (2014) Integrating multibeam backscatter angular response, mosaic and bathymetry data for benthic habitat mapping. PLoS One 9(5):e97339
Clarke, K.R., Gorley, R.N., (2015). PRIMER v7: User Manual/Tutorial. PRIMER-E, Plymouth, 296pp
CPAWS-NL. (2017). Special marine areas in Newfoundland and Labrador - Second Edition. Canadian Parks and Wilderness Society – Newfoundland and Labrador Chapter. Available online at: http://cpaws.org/
Cochrane, G. R., Trusel, L., Harney, J., & Etherington, L. (2011). Habitats and benthos of an evolving fjord, Glacier Bay, Alaska. Seafloor Geomorphology as Benthic Habitat: GeoHAB Atlas of Seafloor Geomorphic Features and Benthic Habitats, 299–308
Copeland, A. (2006). Benthic habitat mapping with multibeam sonar in Newman sound, Terra Nova National Park, Newfoundland. (unpublished Master’s thesis), Memorial University research repository, item ID: 10342. Available online at: http://research.library.mun.ca /10342/
Copeland, A., Edinger, E., Leblanc, P., Bell, T., Devillers, R. & Wroblewski, J. (2008). Marine habitat mapping in Gilbert Bay, Labrador – a marine protected area. Phase III final report. Marine habitat mapping group report # 08–01, Memorial University of Newfoundland, St. John’s, NL
Copeland A, Edinger E, Devillers R, Bell T, LeBlanc P, Wroblewski J (2013) Marine habitat mapping in support of marine protected area management in a subarctic fjord: Gilbert Bay, Labrador, Canada. J Coast Conserv 17(2):225–237
Cote D, Moulton S, Scruton DA, McKinley RS (2001) Microhabitat use of juvenile Atlantic cod in a coastal area of Bonavista Bay, Newfoundland. Trans Am Fish Soc 130(6):1217–1223
Cote D, Moulton S, Frampton P, Scruton D, McKinley R (2004) Habitat use and early winter movements by juvenile Atlantic cod in a coastal area of Newfoundland. J Fish Biol 64(3):665–679
Cote D, Gregory RS, Morris CJ, Newton BH, Schneider DC (2013) Elevated habitat quality reduces variance in fish community composition. J Exp Mar Biol Ecol 440:22–28
Cumming E, Aksu A, Mudie P (1992) Late quaternary glacial and sedimentary history of Bonavista Bay, Northeast Newfoundland. Can J Earth Sci 29(2):222–235
Dale JE, Aitken AE, Gilbert R, Risk MJ (1989) Macrofauna of Canadian arctic fjords. Mar Geol 85(2–4):331–358
Dalley KL, Gregory RS, Morris CJ, Cote D (2017) Seabed habitat determines fish and macroinvertebrate community associations in a subarctic marine coastal nursery. Trans Am Fish Soc 146(6):1115–1125
DFO. (2009). Science guidance on the development of networks of marine protected areas (MPAs). DFO Canadian Science Advisory Secretariat Science Advisory Report 2009/061
DFO. (2018). Marine protected area (MPA) network development. Fisheries and Oceans Canada. Available online at: http://www.dfo-mpo.gc.ca/oceans/networks-reseaux/development-developpement-eng.html
Diesing M, Mitchell P, Stephens D (2016) Image-based seabed classification: what can we learn from terrestrial remote sensing? ICES J Mar Sci 73(10):2425–2441
Environment and Climate Change Canada (2017). Terra Nova Migratory Bird Sanctuary [Online]. Environment and Climate Change Canada – Atlantic Region. Available online at: http://www.ec.gc.ca/ (Accessed February 2018; Modified April 4, 2017)
Garmin Ltd. (2005). Garmin eTrex series specifications. [online]. Garmin International Inc. available online at: https://www8.garmin.com/specs/eTrex_spec_sheet_0105.pdf
Gorman AM, Gregory RS, Schneider DC (2009) Eelgrass patch size and proximity to the patch edge affect predation risk of recently settled age 0 cod (Gadus). J Exp Mar Biol Ecol 371(1):1–9
Grech A, Coles R, Marsh H (2011) A broad-scale assessment of the risk to coastal seagrasses from cumulative threats. Mar Policy 35(5):560–567
Green AL, Fernandes L, Almany G, Abesamis R, McLeod E, Aliño PM, White AT, Salm R, Tanzer J, Pressey RL (2014) Designing marine reserves for fisheries management, biodiversity conservation, and climate change adaptation. Coast Manag 42(2):143–159
Hall-Spencer JM, Moore PG (2000) Scallop dredging has profound, long-term impacts on maerl habitats. ICES J Mar Sci 57(5):1407–1415
Hall-Spencer J, White N, Gillespie E, Gillham K, Foggo A (2006) Impact of fish farms on maerl beds in strongly tidal areas. Mar Ecol Prog Ser 326:1–9
Halpern BS, Walbridge S, Selkoe KA, Kappel CV, Micheli F, D'agrosa C, Bruno JF, Casey KS, Ebert C, Fox HE, Fujita R (2008) A global map of human impact on marine ecosystems. Science 319(5865):948–952
Halpern BS, Frazier M, Potapenko J, Casey KS, Koenig K, Longo C, Lowndes JS, Rockwood RC, Selig ER, Selkoe KA, Walbridge S (2015) Spatial and temporal changes in cumulative human impacts on the world’s ocean. Nat Commun 6(7615):1–7
Hernandez-Kantun J.J. et al. (2017) North Atlantic Rhodolith beds. In: Riosmena-Rodríguez R., Nelson W., Aguirre J. (eds) Rhodolith/Maërl beds: a global perspective. Coastal research library, vol 15. Springer, Cham
Hill NA, Lucieer V, Barrett NS, Anderson TJ, Williams SB (2014) Filling the gaps: predicting the distribution of temperate reef biota using high resolution biological and acoustic data. Estuar Coast Shelf Sci 147:137–147
Ierodiaconou D, Monk J, Rattray A, Laurenson L, Versace VL (2011) Comparison of automated classification techniques for predicting benthic biological communities using hydroacoustics and video observations. Cont Shelf Res 31(2):S28–S38
Joseph, V., Schmidt, A.L., and Gregory, R.S. (2013). Use of eelgrass habitats by fish in eastern Canada DFO Canadian Science Advisory Secretariat Report Science Advisory Report 2012/138
Kamenos NA, Moore PG, Hall-Spencer JM (2003) Substratum heterogeneity of dredged vs un-dredged maerl grounds. J Mar Biol Assoc U K 83(2):411–413
Kamenos NA, Moore PG, Hall-Spencer JM (2004) Small-scale distribution of juvenile gadoids in shallow inshore waters; what role does maerl play? ICES J Mar Sci 61(3):422–429
Lacharité, M., Brown, C. J., & Gazzola, V. (2017). Multisource multibeam backscatter data: developing a strategy for the production of benthic habitat maps using semi-automated seafloor classification methods. Marine Geophysical Research, 1–16
Lecours V, Brown CJ, Devillers R, Lucieer VL, Edinger EN (2016) Comparing selections of environmental variables for ecological studies: a focus on terrain attributes. PLoS One 11(12):e0167128
Lecours V, Devillers R, Edinger EN, Brown CJ, Lucieer VL (2017) Influence of artefacts in marine digital terrain models on habitat maps and species distribution models: a multiscale assessment. Remote Sensing in Ecology and Conservation 3(4):232–246
Lefcheck JS, Wilcox DJ, Murphy RR, Marion SR, Orth RJ (2017) Multiple stressors threaten the imperiled coastal foundation species eelgrass (Zostera marina) in Chesapeake Bay, USA. Glob Chang Biol 23(9):3474–3483
Lewis, S., Coughlan, E., Pepin, P. & Skanes, K. (2017). An assessment of the biological indicators for the Eastport marine protected area (MPA): a case study in support of an ecosystem goods and services valuation of MPAs. DFO Canadian science advisory secretariat report 2017/0001
Li X, Bellerby R, Craft C, Widney SE (2018) Coastal wetland loss, consequences, and challenges for restoration. Anthropocene Coasts 1(0):1–15
Linehan JE, Gregory RS, Schneider DC (2001) Predation risk of age-0 cod (gadus) relative to depth and substrate in coastal waters. J Exp Mar Biol Ecol 263(1):25–44
Lurton, X., Lamarche, G., Brown, C., Lucieer, V. L., Rice, G., Schimel, A., & Weber, T. (2015). Backscatter measurements by seafloor-mapping sonars: guidelines and recommendations. A collective report by members of the GeoHab Backscatter Working Group. Available online at: http://geohab.org/publications/
Martin S., Hall-Spencer J.M. (2017) Effects of ocean warming and acidification on Rhodolith/Maërl beds. In: Riosmena-Rodríguez R., Nelson W., Aguirre J. (eds) Rhodolith/Maërl beds: a global perspective. Coastal research library, vol 15. Springer
McGonigle C, Brown C, Quinn R, Grabowski J (2009) Evaluation of image-based multibeam sonar backscatter classification for benthic habitat discrimination and mapping at Stanton banks, UK. Estuar Coast Shelf Sci 81(3):423–437
Monk J, Ierodiaconou D, Versace VL, Bellgrove A, Harvey E, Rattray A, Laurenson L, Quinn GP (2010) Habitat suitability for marine fishes using presence-only modelling and multibeam sonar. Mar Ecol Prog Ser 420:157–174
Morris CJ, Green JM (2014) MPA regulations should incorporate adaptive management—the case of Gilbert Bay Labrador Atlantic cod (Gadus morhua). Mar Policy 49:20–28
Novaczek E, Howse V, Pretty C, Devillers R, Edinger E, Copeland A (2017a) Limited contribution of small marine protected areas to regional biodiversity: the example of a small Canadian no-take MPA. Front Mar Sci 4:174
Novaczek E, Devillers R, Edinger E, Mello L (2017b) High-resolution seafloor mapping to describe coastal denning habitat of a Canadian species at risk: Atlantic wolffish (Anarhichas lupus). Can J Fish Aquat Sci 74(12):2073–2084
Parks Canada (2018). Terra Nova National Park [Online]. Parks Canada – National Parks. Available online at: http://www.pc.gc.ca/ (Accessed February 2018; Modified January 2, 2018)
Rao AS, Gregory RS, Murray G, Ings DW, Coughlan EJ, Newton BH (2014) Eelgrass (Zostera marina) locations in Newfoundland and Labrador. In: Canadian technical report on fisheries and aquatic sciences 1488–53793113. Canada, Fisheries and Oceans
Rengstorf AM, Yesson C, Brown C, Grehan AJ (2013) High-resolution habitat suitability modelling can improve conservation of vulnerable marine ecosystems in the deep sea. J Biogeogr 40(9):1702–1714
Smith SJ, Sameoto JA, Brown CJ (2017) A novel approach to setting biological reference points for sea scallops Placopectin magellanicus that incorporates the spatial distribution of productivity. Can J Fish Aquat Sci 74(5):650–667
Stanley R, Morris C, Snelgrove P, Metaxas A, Pepin P (2018) The efficacy of small closures: a tale of two marine protected areas in Canada, In The Palgrave Handbook of Sustainability (pp. 207–238). Palgrave Macmillan, Cham
Steller DL, Riosmena-Rodríguez R, Foster MS, Roberts CA (2003) Rhodolith bed diversity in the Gulf of California: the importance of rhodolith structure and consequences of disturbance. Aquat Conserv Mar Freshwat Ecosyst 13(1):5–20
Syvitski, J. P., & Shaw, J. (1995). Sedimentology and geomorphology of fjords. In Developments in sedimentology (Vol. 53, 113-178). Elsevier
Thistle ME, Schneider DC, Gregory RS, Wells NJ (2010) Fractal measures of habitat structure: maximum densities of juvenile cod occur at intermediate eelgrass complexity. Mar Ecol Prog Ser 405:39–56
Walker DJ, Alford JB (2016) Mapping Lake sturgeon spawning habitat in the upper Tennessee River using side-scan sonar. N Am J Fish Manag 36(5):1097–1105
Walton ME, Hayes J, Al-Ansi M, Abdallah M, Al Maslamani I, Al-Mohannadi M, Al-Shaikh I, D’Urban Jackson T, Szostek C, Egerton J, Kaiser MJ (2017) Towards spatial management of fisheries in the Gulf: benthic diversity, habitat and fish distributions from Qatari waters. ICES J Mar Sci 75(1):178–189
Waycott M, Duarte CM, Carruthers TJ, Orth RJ, Dennison WC, Olyarnik S, Calladine A, Fourqurean JW, Heck KL, Hughes AR, Kendrick GA (2009) Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc Natl Acad Sci 106(30):12377–12381
Wilhere GF (2002) Adaptive management in habitat conservation plans. Conserv Biol 16(1):20–29
Young M, Carr M (2015) Assessment of habitat representation across a network of marine protected areas with implications for the spatial design of monitoring. PLoS One 10(3):e0116200
Young, M.A., Wedding, L.M. & Carr, M.H. (2017). Applying landscape ecology for the design and evaluation of marine protected area networks. Seascape Ecology, 429-462
Zajac RN, Lewis RS, Poppe LJ, Twichell DC, Vozarik J, DiGiacomo-Cohen ML (2003) Responses of infaunal populations to benthoscape structure and the potential importance of transition zones. Limnol Oceanogr 48(2):829–842
Zajac RN (2008) Challenges in marine, soft-sediment benthoscape ecology. Landsc Ecol 23(1):7–18
Acknowledgments
This research is sponsored by the NSERC Canadian Healthy Oceans Network and its Partners: Department of Fisheries and Oceans Canada and INREST (representing the Port of Sept-Îles and City of Sept-Îles). We thank Dr. Trevor Bell for contributing to the multibeam data collection. We also thank Bonnell Squire and Sandy Turner of Happy Adventure, NL and Jackie Saturno for support during collection of the 2016 ground truthing data set. This research is dedicated to the memory of Callum Mireault – a dear friend, marine scientist and colleague who is greatly missed.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Proudfoot, B., Devillers, R., Brown, C.J. et al. Seafloor mapping to support conservation planning in an ecologically unique fjord in Newfoundland and Labrador, Canada. J Coast Conserv 24, 36 (2020). https://doi.org/10.1007/s11852-020-00746-8
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11852-020-00746-8