
Abstract
Creating robust datasets of plant–insect interactions is important for understanding ecosystem dynamics, and data on species interactions can be used to evaluate conservation interventions. In the present work, we collected plant–herbivore–parasitoid data on an understudied but critical ecosystem—gallery forests in the Brazilian cerrado. We collected caterpillars on shrubs of Piper (Piperaceae) over the course of a year in seven gallery forests of varying sizes in order to compare seasonal changes in α- and β- diversity and tritrophic interaction networks as well as the role of fragment size in determining species and interaction diversity. Caterpillars were more abundant and diverse in the wet season and also increased with resource availability—the more Piper individuals present, the greater the abundance and richness of herbivores. The number of unique interactions between (i) plants and herbivores and (ii) herbivores and parasitoids did not change across seasons, but there was a high degree of turnover in the herbivore fauna between sites and seasons. Specialization was greatest in the dry-rainy season transition, when new leaves typically flush. Consistent with records of parasitism rates in the cerrado sensu stricto, parasitism in the gallery forests was greatest in the dry-rainy seasons. Forest size was not related to caterpillar richness. Overall, this work demonstrates the conservation value of gallery forests in supporting plant species that span the Amazon and the Atlantic Rainforest as well as diverse and highly seasonal trophic interactions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Plant–insect herbivore interactions are highly diverse and dominate terrestrial ecosystems both in terms of individual actors (plants and herbivores) and the ecological and evolutionary stories they tell (interaction networks; Del-Claro and Torezan-Silingardi 2021; Luna and Dáttilo 2021). Interaction networks provide insight into ecosystem structure and stability that cannot be understood from species richness alone (Poisot et al. 2015; Landi et al. 2018), and preserving network structure preserves ecosystem functioning (Tylianakis et al. 2010). Conservation efforts may therefore be more effective when focused on preserving entire ecological networks rather than just individual species, and more studies are needed to document interactions in both intact and disturbed or threatened areas (Dyer et al. 2010; Del-Claro and Dirzo 2021). Data quantifying and summarizing interactions can inform conservation by highlighting areas in need of protection or by serving as a reference point for restoration. Collecting the natural history data required to describe networks requires long-term studies, so documenting interactions should be an ongoing research objective rather than one that rises out of a need for specific conservation efforts (Tylianakis et al. 2010; Luna and Dáttilo 2021).
The Brazilian cerrado is a highly threatened and undervalued biome (Klink and Machado 2005; Carvalho et al. 2009). It is a biodiversity hotspot (Myers et al. 2000), but only 8.2% of it is protected (MMA 2020). Plant–insect herbivore interactions are well studied in parts of the cerrado (Diniz and Morais 1995, 1997, 2002; Morais et al. 1999; Diniz et al. 2001; Marquis et al. 2002; Scherrer et al. 2010; de Araújo et al. 2013; Diniz et al. 2013; Vilela et al. 2014; Scherrer et al. 2016; Lepesqueur et al. 2018), making it possible to decipher patterns in ecological networks. β-diversity of plant-caterpillar interactions is high in the cerrado, and even though caterpillar communities on closely related host plants in the cerrado tend to be similar to each other, most caterpillars are specialized (Morais et al. 2011), creating a mosaic of diverse caterpillar networks.
Changes in plant–insect interaction networks over space and time have been documented in a number of ecosystems (Lange et al. 2013; López-Carretero et al. 2014; Scherrer et al. 2016; Koch et al. 2018; Lepesqueur et al. 2018; de Araújo et al. 2019; Campos Moreno et al. 2021; Luna and Dáttilo 2021). In the cerrado, there are strong seasonal changes in plant-caterpillar interactions, mostly driven by rewiring of networks rather than changes in community composition (Lepesqueur et al. 2018). Caterpillar diet breadth increases in the cerrado’s dry season (Scherrer et al. 2016) as it also does in the dry season in coastal habitats (López-Carretero et al. 2014). The role of interaction rewiring suggests species with narrowed diet breadth expand their host range in drier months due to changes in host plant nutritional quality and phytochemistry (Scherrer et al. 2016).
Similar changes in host-parasitoid interactions have been documented in other ecosystems, and interactions between the second and third trophic level also vary more across seasons than do species assemblages (Lewis et al. 2002). There are a number of hypothesized mechanisms responsible for this tritrophic rewiring, including the hypothesis that parasitism levels change with the food plant species of a caterpillar host (Lill et al. 2002) and that parasitism can affect an herbivore’s host choice (Smilanich et al. 2011).
The present study examined changes in caterpillar diversity and tritrophic plant–insect interactions across seven gallery forests in the cerrado. There is a paucity of research on plant–insect interactions in the gallery forests within cerrado, and these forests provide an interesting point of comparison for understanding landscape level and seasonal changes in species interactions. While the cerrado is highly seasonal, gallery forests are evergreen, so interaction networks may be less seasonal in gallery forests than they are in the surrounding cerrado. Gallery forests are highly fragmented, and variation in their size may provide information about the effects of fragmentation on diversity. In order to understand seasonality in trophic interaction networks, we examined temporal changes in α- and β-caterpillar diversity and in tritrophic interaction networks. We also examined changes in caterpillar diversity and plant-caterpillar-parasitoid interactions in gallery forests of differing sizes, and explored bottom-up effects of resource availability for herbivore abundance and diversity. We hypothesized that tritrophic interaction networks would be more specialized in the wet season and that rewiring would change more than species composition over time. We also hypothesized that increased host plant availability and host plant richness would be related to increased herbivore diversity. Finally, we hypothesized that species and interactions would be more diverse in gallery forests of greater size.
Materials and methods
Study sites
Plant–herbivore interaction plots were surveyed in seven gallery forests located in the cerrado of the Distrito Federal and Goiás in central Brazil. Five study sites were located in designated conservation units of the Distrito Federal; one was located outside the urban center of Pirenópolis, Goiás, and the last was in Chapada dos Veadeiros National Park, Goiás (Tables S1, S2). The minimum distance between sites was 7.01 km, and the maximum distance was 235.42 km. Sites ranged from 902 to 1247 m a.s.l. The cerrado is the second largest biome in South America after the Amazon and is the most biodiverse savanna in the world with over 4800 endemic species. It once occupied 23% of Brazil, primarily in the country’s central highlands (Ratter et al. 1997); today only ~ 60% of the biome contains natural vegetation (Sano et al. 2010). Cerrado spans a range of habitats from open fields to dense woodlands (in order of increasing crown cover and height: cerrado ralo, cerrado sensu stricto, cerrado denso, and cerradão). These habitats form mosaics with forest and grassland habitats (Ribeiro and Walter 2008). Grasslands include dry (campo limpo, campo sujo, campo cerrado) and seasonal (campo rupestre) habitats. Forests include seasonally dry forests (mata seca) and riverine forests (gallery forest or mata de galeria). Factors such as soil moisture and fertility as well as fire dynamics influence the floristic composition and vegetation structure that characterizes these different habitats. Mean annual monthly temperatures range between 20 and 26 °C, and seasonality is marked by changes in precipitation with 86% of the region receiving between 1000 and 2000 mm of rain per year (Oliveira and Marquis 2002).
Gallery forests historically comprised 5% of the cerrado (Dias 1996). These forests act as biological corridors between the Atlantic Rainforest and the Amazon (Rizzini 1979; Oliveira-Filho and Ratter 1995). Gallery forests house species found in the Atlantic Rainforest, the Amazon, and drainage basins of the Paraná River (Oliveira-Filho and Ratter 1995). While the cerrado is highly seasonal (Espinoza et al. 1982; Alvares et al. 2013), gallery forests are evergreen (Ribeiro and Walter 2008).
Study system
Shrubs of the genus Piper (Piperaceae; Ribeiro and Walter 2008) are among the dominant plants in the understory of gallery forests in the cerrado, the Atlantic Rainforest, and the Amazon. Piper includes ca. 2,600 species, the majority of which are Neotropical (Callejas-Posada 2020), and it is the second most species rich genus of seed plants in the Amazon (Cardoso et al. 2017). Centers of endemism of Piper occur in both the Amazon and the Atlantic Rainforest (Quijano-Abril et al. 2006). Piper requires perennially moist soils, and most species are restricted to the shaded understory of tropical rainforests (Burger 1971). As such, it is absent from the cerrado sensu stricto (Marquis 2004), which is characterized by a marked dry season (Gomes de Moraes et al. 2016). In contrast, gallery forests in the cerrado are characterized by relatively high levels of soil moisture year-round and support Piper species (Oliveira-Filho et al. 1990). Several studies mention the presence of Piper in the cerrado, but the species registered are frequently typical of disturbed habitats (e.g., Piper aduncum L.) and are likely found in gallery forests or along their margins (pers. obs., E.J.T.; Oliveira et al. 2013).
Piper and its associated herbivores have been studied in other parts of Brazil (Shimbori 2009) and Central and South America (Marquis 1991; Gentry and Dyer 2002; Janzen and Hallwachs 2005; Rodríguez-Castañeda et al. 2010; Tepe et al. 2014; Glassmire et al. 2019; Slinn et al. 2018). These studies show Piper hosts a high richness of specialist and generalist caterpillars as well as diverse herbivore-parasitoid interactions. Piper arboreum Aubl. is a species of particular interest because of its broad distribution throughout Central and South America and the occurrence of isolated populations across its range (Yuncker 1972) that can potentially host unique arthropod assemblages. It is possible that populations of this species have diverged to the point they now actually comprise a complex of cryptic species. The present work documents Piper-herbivore-parasitoid interactions in gallery forests.
Collection and rearing
Caterpillars were collected between May 2015 and April 2016 (see Table S3 for collection information). Circular plots measuring 10 m in diameter were established in gallery forests with a randomly selected individual of P. arboreum as the center of each plot. Plots were a minimum of 5 m apart; most were more than 10 m apart. Piper abundance and richness were recorded for each plot, and all leaves of all Piper individuals were exhaustively searched for lepidopteran eggs, larvae, and pupae. Each plot was surveyed only one time, and plots were not evenly distributed across sites or seasons, so different components of the dataset were analyzed to make seasonal comparisons. All stages of immature Lepidoptera were brought to the lab, photographed, and reared to adults or until parasitoids emerged. Insects were reared in plastic containers and were fed leaves of the same species they were originally found on. A total of 243 caterpillars were collected. Fifteen percent of larvae were successfully reared to adulthood, and percent parasitism was 49% (number of parasitized caterpillars/number of adult Lepidoptera + parasitized caterpillars). Adult Lepidoptera were spread and identified by specialists (primarily V. Becker) or with the help of a reference collection. Herbarium samples were made for each species of Piper in the plots and are deposited in the herbarium of the University of Brasilia. Lepidoptera specimens were deposited in the Entomological Collection in the Zoology Department at the University of Brasilia. Parasitoid wasps were identified using a Nikon stereomicroscope (ZM1500). Wasps were identified to the genus level and classified as morphospecies. They were labeled and deposited in the Entomological Collection of the Laboratory of Ecological Interactions and Biodiversity at the Federal University of Goiás, Brazil. Parasitoids that did not eclose were included in the dataset and were assigned unique species names based on unique host associations because of their extreme diversity and specialization (Forbes et al. 2018). Parasitoids that did not eclose but were from the same species of caterpillar collected at the same time were considered the same species.
Data analyses
Descriptive statistics
Caterpillar diversity was calculated for each study site and each season: the rainy season (December–February), rainy to dry season transition (March–May), dry season (June–August), and the dry to rainy season transition (September–November; seasonal assignments following Lepesqueur et al. 2018). Diversity was calculated using Hill numbers (Jost 2006). For calculations of the Shannon–Wiener index (q = 1) and Simpson’s index (q = 2), species abundances were corrected based on the number of Piper plants searched because collections were not standardized across seasons or sites. Diversity was calculated across plots because caterpillars were absent from many plots (values presented are not means, but show total diversity, hence there is no variation around them).
Piper individuals in the plots at each site were summarized according to abundance, richness (q = 0), and Simpson’s index (q = 2). Variation in richness as abundance among plots at each study site was also calculated. Diversity was not calculated for parasitoids because of the low level of parasitism and high mortality of parasitoids, which did not allow all species to be identified.
Caterpillar species richness in the different study sites was also estimated using abundance data with the Chao1 index as a lower bound and the Abundance-based Coverage Estimator modified for heterogeneous communities (ACE-1) as a higher bound estimate using SpadeR (Chao et al. 2016) in R. Rarefaction curves of species richness were also created using the iNEXT package (Chao et al. 2014; Hsieh et al. 2020) in R.
Analyses
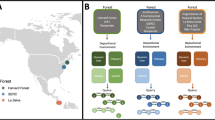
Factors affecting the richness and abundance of caterpillars per plot as well as the number of unique interactions per plot (plants and herbivores plus herbivores and parasitoids) were examined with hierarchical Bayesian models with uninformed priors. Predictor variables included the abundance and richness of Piper per plot and season. For the lowest level of the hierarchy, we included study sites. In these analyses, ‘season’ was defined as the rainy (rainy and rainy-dry season) and dry (dry and dry-rainy season) periods due to the uneven distribution of plots across study sites and seasons. Prior to analyses, the number of unique interactions per plot was standardized by the abundance of Piper in a plot. Models were run using the R2jags package (Su and Yajima 2020) with four chains and 1,000,000 iterations. We evaluated model performance using trace and density plots and the Gelman–Rubin, Geweke, Heidelberger–Welch, and Raftery–Lewis convergence diagnostics with the superdiag package, specifying 1000 burnins (Tsai et al. 2012). Best-fit models were determined with deviance information criterion scores. Caterpillar plots were constructed with ggplot2 (Wickham 2016) to illustrate estimates of beta coefficients and their 90% credible intervals for each location. Locations are arranged along the y-axis of the caterpillar plots in order of increasing gallery forest area where the plots were conducted (calculated in Google Earth). All models converged (Figs. S1–S3).
The Morisita similarity index (MSI) for multiple communities was calculated to describe species turnover across sites and between seasons (Chao et al. 2008). MSI values closer to 1 indicate communities are more similar in their species composition. Calculations were based on species abundances (again corrected for the number of Piper individuals searched per season), and data were bootstrapped 200 times using SpadeR (Chao et al. 2016). The seasonal comparison was only conducted for two of the field sites, ESECAE and FAL, where collections were made during each of the four seasons. Turnover in Piper species between sites was also calculated, correcting for the number of plots per site.
We constructed matrices for each season and for all data combined. Each row consisted of a unique plant species, and each column represented a caterpillar species with which the plant directly interacted or a parasitoid species with which the plant indirectly interacted. We calculated multiple network metrics using the networklevel function in the bipartite package in R (Dormann et al. 2008, 2009). Connectance (the ratio of realized to possible interactions), the weighted nestedness metric based on overlap and decreasing fill (WNODF; Almeida-Neto and Ulrich 2011; scores vary between 0 (no nestedness) and 100 (perfectly nested)), linkage density (weighted diversity of interactions per node), Alatalo interaction evenness (a measure of uniformity of interaction strength), and H2 (a measure of specialization) are reported. We developed null networks to compare with observed values. We applied the ‘r2dtable’ function using 1000 randomizations in the bipartite package. The algorithm constrains marginal totals such that interactions are randomly distributed, but total interactions per species are maintained. Observed values were compared to these null distributions to determine if observed values differed significantly from null values using a two-tailed test with 5% error level. For more valid comparisons across seasons and studies, observed values were z-transformed (z = [x − μ]/σ; where x = observed value, μ = mean of null distribution, σ = standard deviation of null distribution).
Seasonal changes in species and interactions were examined for their contributions to tritrophic network turnover based on Poisot et al. (2012). Dissimilarity of interactions (βWN) was partitioned into its additive components: turnover owed to changes in species composition (βST), and that owed to the rewiring of interactions (βOS). We calculated dissimilarity using the Jaccard index. In this case, a value of 1 indicates complete interaction dissimilarity between two communities. Comparisons were made with the complete dataset and with a randomly created subset of the data including 640 plant records and associated herbivores and parasitoids where relevant per season. Six hundred forty was the minimum number of plants searched across seasons (this was the case for the rainy season). Tritrophic networks were illustrated using the plotweb function in the bipartite package.
Results
Thirteen species of Piper were found across the seven study sites, and species richness per plot was between one and six species (Table S4). A total of 67 species of Lepidoptera were collected; 119 individuals were identified to the species level, and 36 additional individuals were identified to the genus level (Table S5). Variance in plot level Piper richness between sites was between 0.02 and 1.09. The most diverse site was PNCV, followed by JBB and RECOR (Table S4). Piper arboreum and Piper aduncum L. were present at all sites, although P. aduncum was not sampled in all seasons a site was visited. Caterpillars were found on P. arboreum throughout the year, and caterpillars were observed on P. aduncum in all seasons except the rainy season. Piper caldense C.DC., Piper crassinervium Kunth, and Piper xylosteoides (Kunth) Steud. hosted caterpillars in the rainy-dry and dry seasons. Caterpillars were found on five other species surveyed in only one season, although these plants were not as well represented in our samples. Piper communities were fairly similar between sites (MSI = 0.83 ± 0.01).
Hill numbers describing caterpillar diversity were highest in the rainy season (Table 1). This is in spite of the fact that the smallest number of Piper individuals (640) were searched in the rainy season. Overall caterpillar diversity was highest in FAL and JBB (Table 1), which had the largest areas of gallery forest (Table 1; Fig. S4). Chao1 and ACE1 estimates of caterpillar richness suggest Pirenópolis, JBB, and FAL were the most species rich sites (Table S6). The site with the smallest amount of gallery forest (IBGE) had caterpillar diversity comparable with sites with 5–19 times more forest (Table 1; Table S1). Parasitism levels were slightly higher in the dry (16.2%) and dry-rainy seasons (16.7%) than in the rainy (13.9%) and rainy-dry seasons (12.9%).
Bayesian models indicate caterpillar abundance and richness were higher in the wet relative to the dry season, and caterpillar abundance and richness were higher when Piper was more abundant. Posterior distributions of beta coefficients describing interaction richness (the number of unique plant–herbivore or herbivore-parasitoids interactions per plot) were very small and credible intervals were mostly centered on zero in response to seasonal change and Piper abundance (Table 2). No patterns between gallery forests differing in size emerged (Fig. 1).

Beta values values and 90% credible intervals resulting from hierarchical Bayesian analyses of caterpillar abundance (a, b), caterpillar richness (c, d), and the number of unique interactions per plot (e, f). Increases in caterpillar abundance a and richness c were related to the abundance of Piper in a plot. Caterpillar abundance b and richness d were greater in the wet season. Increases in Piper richness were typically related to decreases in the number of unique interactions in a plot (e). The number of unique interactions was greater in the dry season (f). Study sites are shown in increasing order of the size of gallery forests where plots were conducted; no clear relationship with forest size emerged
Caterpillar communities changed across seasons (MSI = 0.20 ± 0.06). No two seasons were the same in species composition, but the rainy-dry and dry seasons were more similar than any other combination of seasons (Table 3). Differences between seasons were greater than differences between study sites (MSI = 0.37 ± 0.03; Table S6). Between site differences were not related to the distance between study sites or turnover in Piper species (Tables S7, S8). When comparing caterpillar communities on P. arboreum alone, similarity was only slightly greater between sites, indicating differences across forests in spite of the identity of the host plant (MSI = 0.42 ± 0.05).
These patterns are further reflected in seasonal interaction networks (Figs. 2, 3). There was a high degree of turnover in networks between seasons, driven primarily by changes in community composition (βST > βOS; Table 4). Results were the same when comparing the full dataset and the sub-sampled dataset (Table 4). Changes in interactions between species shared across seasons only contributed to network dissimilarity when comparing the (i) rainy-dry and dry seasons and (ii) the rainy and dry seasons (see Table S5 for a list of caterpillars and their parasitoids by season).

Complete network of all Piper-caterpillar-parasitoid interactions across seasons and sites

Seasonal networks of Piper-caterpillar-parasitoid interactions. The size of the parasitoid boxes in the dry-rainy season are not drawn to scale; each parasitoid species was represented by only one individual
Network parameters varied across seasons (Tables 5, S9; Fig. S5). Connectance in the rainy season was the highest (0.52). The rainy-dry season was the least connected (0.14) and most nested (WNODF = 10.46) network. The dry season was the least nested (WNODF = 0) network and had the lowest value of interaction evenness (0.44). The dry-rainy season did not have any values that differed from null expectations, although it was the most specialized and had the highest interaction evenness. The rainy season was the least modular (0.18) while the dry-rainy season was the most modular (0.37).
Discussion
Studies of seasonal variation in plant–insect interactions in the cerrado demonstrate a high degree of specialization and seasonal turnover. Only about 40% of plants typically host caterpillars across multiple seasons (Scherrer et al. 2016; Lepesqueur et al. 2018). In our study, focused on a single genus in cerrado gallery forests, caterpillars were recorded on our most abundant host plant throughout the year and on other common host plants for multiple seasons. These patterns suggest gallery forest species may be more consistent hosts across seasons, even as caterpillar species experience substantial turnover.
Another study from the cerrado that divided the year into four seasons found 70% of caterpillars were only present in a single season, while only 3.5% of species were found year-round (Lepesqueur et al. 2018). We similarly found 14 of 67 Lepidoptera species (20%) in more than one season, and only one species, Memphis moruus (Nymphalidae), occurred in all four seasons. The greatest caterpillar richness occurs in the rainy-dry season in the cerrado; in our study in gallery forests, the highest richness was similarly observed in the rainy season.
In the cerrado, plant–herbivore interactions are more generalized in the dry season with caterpillars consuming a wider variety of plants (Scherrer et al. 2016). In contrast, our data demonstrate specialization in plant–herbivore–parasitoid interactions was lowest in the rainy season. This could be because only two host plant species were included in rainy season plots, while four were present in the dry season plots. Cerrado plants flush new leaves in the dry-rainy season, and leaf production of deciduous and evergreen species alike peaks at the end of the dry-rainy season (Gutierrez de Camargo et al. 2018). This is also the time of year when the greatest herbivory is observed in the cerrado (Marquis et al. 2001). The abundance of new leaves and the fact that young leaves are often highly defended (Kursar and Coley 2003; Massad et al. 2014; Wiggins et al. 2016; Coley et al. 2018) may promote the specialization we observed in dry-rainy season and the greater abundance and α-diversity of caterpillars observed in the rainy season as resources become abundant. Studies of plant–insect interactions in the cerrado focused on other insect orders similarly show important temporal changes. Changes in plant phenology and the expression of traits like extrafloral nectaries change ant-plant networks (Lange et al. 2013; Vilela et al. 2014; Moura et al. 2021). In addition, as the climate warms, floral resources become more concentrated temporally, resulting in more herbivory (Vilela et al. 2018).
Bottom-up forces do not operate in isolation, and the presence of herbivores may also be influenced from the top-down, regardless of plant quality (Myers 1981; Janzen 1993). In our study, plant–herbivore–parasitoid interactions were most specialized in the dry-rainy season. Percent parasitism was slightly higher in the dry and dry-rainy seasons, and connectance was highest in the dry-rainy seasons, indicating herbivore species were more evenly attacked by parasitoids at that time. Herbivore seasonality may therefore be driven by bottom-up changes in resource availability or by top-down limitations. This requires further study in gallery forests. In tropical dry forest the presence of young leaves seems to be more important than the risk of parasitism in determining leaf miner phenology (Hopkins and Memmot 2003).
Gallery forests provide important islands of habitat within a sea of cerrado. The forests support Piper species and caterpillars shared with the Amazon and the Atlantic Rainforest (authors, pers. obs.). The size of the gallery forests where our plots were located varied from 0.04 to 1.25 km2, but there were no patterns of association between forest area and the abundance or richness of plants or herbivores. Rarefaction curves suggest that the two largest sites may have the highest caterpillar richness, as predicted by the well-established species-area relationship (Fig. S4; Preston 1962). Although there was a high degree of dissimilarity between caterpillar assemblages collected in our different study sites, some species of caterpillars were widespread. Other studies have also recorded Lepidoptera with broad distributions, overlapping the cerrado and the Amazon or Atlantic Rainforest (Camargo and Becker 1999; Amorim et al. 2009). Patterns of herbivore biogeography are therefore similar to those of plants, with gallery forests housing multiple species found in both the Amazon and the Atlantic Rainforest (Oliveira-Filho and Ratter 1995).
In other regions of South and Central America, higher than expected plant richness occurs in gallery forests because they house plant species typically found in tropical moist forests (Meave et al. 1991). Predictions based on island biogeography (MacArthur and Wilson 1967) may therefore not be met for gallery forest islands because they effectively act as corridors for Amazonian and Atlantic Rainforest species (Oliveira-Filho and Ratter 1995). The role of gallery forests as corridors is of great value to conservation and may become more important as climate change affects both the Amazon and the Atlantic Rainforest (Sörensson et al. 2010; Penalba and Rivera 2013). The specificity of herbivore communities as well as the inclusion of widespread species contribute to the substantive conservation value of cerrado gallery forests.
In spite of their importance as corridors, our data also showed considerable turnover in the herbivore fauna between gallery forests, even when sampling on the same host plant species. These results emphasize the importance of creating robust natural history databases to document biodiversity and species interactions across landscapes. Protecting gallery forests is necessary to maintain their roles as corridors for widespread species and as islands where species with narrow ranges are isolated. Natural history data from gallery forests could potentially be used to support an expansion of the size of Permanent Protected Areas (Áreas de Preservação Permanentes; https://www.embrapa.br/en/codigo-florestal/entenda-o-codigo-florestal/area-de-preservacao-permanente) in the cerrado.
Data availability
Data will be shared upon request and will be made available on Dryad (datadryad.org) upon acceptance of the manuscript.
References
Almeida-Neto M, Ulrich W (2011) A straightforward computational approach for measuring nestedness using quantitative matrices. Environ Model Softw 26:173–178. https://doi.org/10.1016/j.envsoft.2010.08.003
Alvares CA, Stape JL, Sentelhas PC, Moraes G, Leonardo J, Sparovek G (2013) Köppen’s climate classification map for Brazil. Meteorol Z 22:711–728
Amorim FW, de Avila RS, de Camargo AJA et al (2009) A hawkmoth crossroads? Species richness, seasonality and biogeographical affinities of Sphingidae in a Brazilian Cerrado. J Biogeogr 36:662–674. https://doi.org/10.1111/j.1365-2699.2008.02033.x
Bersier LF, Banasek-Richter C, Cattin MF (2002) Quantitative descriptors of food-web matrices. Ecol Lett 83:2394–2407
Blüthgen N, Menzel F, Blüthgen N (2006) Measuring specialization in species interaction networks. BMC Ecol 6:1–12
Burger WC (1971) Piperaceae. In: Flora costaricensis. Fieldiana Bot 35:5–227
Callejas-Posada R (2020) Piperaceae. In: Davidse G, Ulloa Ulloa C, Hernández Macías HM, Knapp S (eds) Flora mesoamericana, vol 2. Missouri Botanical Garden Press, St. Louis, pp 1–590
Camargo AJA, Becker VO (1999) Saturniidae (Lepidoptera) from the Brazilian Cerrado: composition and biogeographic relationships. Biotropica 31:696–705
Campos-Moreno DF, Dyer LA, Salcido D, Massad TJ, Pérez-Lachaud G, Tepe EJ, Whitfield JB, Pozo C (2021) Importance of interaction rewiring in determining spatial and temporal turnover of tritrophic (Piper-caterpillar-parasitoid) metanetworks in the Yucatán Península, México. Biotropica. https://doi.org/10.1111/btp.12946
Cardoso D, Särkinen T, Alexander S et al (2017) Amazon plant diversity revealed by a taxonomically verified species list. PNAS 114:10695–10700. https://doi.org/10.1073/pnas.1706756114
Carvalho FMV, De Marco P, Ferreira LG (2009) The Cerrado into-pieces: habitat fragmentation as a function of landscape use in the savannas of central Brazil. Biol Conserv 142:1392–1403. https://doi.org/10.1016/j.biocon.2009.01.031
Chao A, Jost L, Chiang SC et al (2008) A two-stage probabilistic approach to multiple-community similarity indices. Biometrics 64:1178–1186. https://doi.org/10.1111/j.1541-0420.2008.01010.x
Chao A, Gotelli NH, Hsieh TC, Sander EL, Ma KH, Colwell RK, Ellison AM (2014) Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecol Monogr 84:45–67
Chao A, Ma KH, Hsieh TC, Chiu CH (2016) SpadeR (species-richness prediction and diversity estimation in R): an R package in CRAN. Program and user’s guide also published at https://cran.r-project.org/web/packages/SpadeR/SpadeR.pdf. Accessed 24 July 2021
Coley PD, Endara M-J, Kursar TA (2018) Consequences of interspecific variation in defenses and herbivore host choice for the ecology and evolution of Inga, a speciose rainforest tree. Oecologia 187:361–376. https://doi.org/10.1007/s00442-018-4080-z
de Araújo WS, Scareli-Santos C, Guimaraes Guilherme FA, Cuevas-Reyes P (2013) Comparing galling insect richness among neotropical savannas: effects of plant richness, vegetation structure and super-host presence. Biodivers Conserv 22:1083–1094. https://doi.org/10.1007/s10531-013-0474-8
de Araújo WS, Moreira LT, Falcão LAD et al (2019) Superhost plants alter the structure of plant-galling insect networks in neotropical savannas. Plants (Basel) 8:369. https://doi.org/10.3390/plants8100369
de Oliveira-Filho AT, Ratter JA, Shepherd GJ (1990) Floristic composition and community structure of a central Brazilian gallery forest. Flora 184:103–117
Del-Claro K, Dirzo R (2021) Impacts of Anthropocene defaunation on plant-animal interactions. In: Del-Claro K, Torezan-Silingardi (eds) Plant-animal interactions. Springer Nature Switzerland, Switzerland, pp 333–348
Del-Claro K, Torezan-Silingardi HM (2021) An evolutionary perspective on plant-animal interactions. In: Del-Claro K, Torezan-Silingardi (eds) Plant-animal interactions. Springer Nature Switzerland, Switzerland, pp 93–118
Dias BFS (1996) Cerrados: Uma caracterização. In: Dias BFS (ed) Alternativas de desenvolvimento dos Cerrados: manejo e conservação dos recursos naturais renováveis. Fundação Pró-Natureza, Brasília, pp 11–25
Diniz IR, Morais HC (1995) Larvas de Lepidoptera e suas plantas hospedeiras em um cerrado de Brasília, Distrito Federal, Brasil. Rev Bras Entomol 39:755–770
Diniz IR, Morais HC (1997) Lepidopteran caterpillar fauna of cerrado host plants. Biodivers Conserv 6:817–836. https://doi.org/10.1023/B:BIOC.0000010404.17467.6c
Diniz IR, Morais HC (2002) Local pattern of host plant utilization by lepidopteran larvae in the cerrado vegetation. Entomotropica 17:115–119
Diniz IR, Morais HC, Camargo AJA (2001) Host plant of lepidopteran caterpillar in the cerrado of Distrito Federal, Brazil. Rev Bras Entomol 45:107–122
Diniz IR, Braga L, Lepesqueur C, Silva N, Morais HC (2013) Lagartas do Cerrado—Guia de Campo. Technical Books. Rio de Janeiro, Brazil
Dormann CF, Gruber B, Fründ J (2008) Introducing the bipartite package: analysing ecological networks. R News 8:8–11
Dormann CF, Fründ J, Blüthgen N, Gruber B (2009) Indices, graphs and null models: analyzing bipartite ecological networks. Open Ecol J 2:7–24
Dunne JA, Williams RJ, Martinez ND (2002) Network structure and biodiversity loss in food webs: robustness increases with connectance. Ecology 5:558–567
Dyer LA, Walla TR, Greeney HF et al (2010) Diversity of interactions: a metric for studies of biodiversity. Biotropica 42:281–289. https://doi.org/10.1111/j.1744-7429.2009.00624.x
Espinoza CW, Azevedo LG, Jarreta M (1982) O clima da região dos cerrados em relação à agricultura. Planaltina, Brasil: Embrapa-CPAC. Circ Téc 9:15
Forbes AA, Bagley RK, Beer MA et al (2018) Quantifying the unquantifiable: why Hymenoptera, not Coleoptera, is the most speciose animal order. BMC Ecol 18:21. https://doi.org/10.1186/s12898-018-0176-x
Gentry GL, Dyer LA (2002) On the conditional, nature of neotropical caterpillar defenses against their natural enemies. Ecology 83:3108–3119
Glassmire AE, Philbin C, Richards LA et al (2019) Proximity to canopy mediates changes in the defensive chemistry and herbivore loads of an understory tropical shrub, Piper kelleyi. Ecol Lett 22:332–341. https://doi.org/10.1111/ele.13194
Gomes de Moraes M, Machado de Carvalho MA, Franco AC, Pollock CJ, Figueiredo-Ribeiro RCL (2016) Fire and drought: soluble carbohydrate storage and survival mechanisms in herbaceous plants from the Cerrado. Bioscience 66:107–117. https://doi.org/10.1093/biosci/biv178
Gutierrez de Camargo MG, de Carvalho GH, de Costa AB et al (2018) Leafing patterns and leaf exchange strategies of a Cerrado woody community. Biotropica 50:442–454. https://doi.org/10.1111/btp.12552
Hopkins GW, Memmott J (2003) Seasonality of a tropical leaf-mining moth: leaf availability versus enemy-free space. Ecol Entomol 28:687–693. https://doi.org/10.1111/j.1365-2311.2003.00552.x
Hsieh TC, Ma KH, Chao A (2020) iNEXT: iNterpolation and EXTrapolation for species diversity. R package version 2.0.20. https://doi.org/10.1111/2041-210X.12613. Accessed 24 July 2021
Janzen DH, Hallwachs W (2005) Caterpillars, pupae, butterflies & moths of the ACG. http://janzen.sas.upenn.edu/. Accessed 24 July 2021
Janzen DH (1993) Caterpillar seasonality in a Costa Rican dry forest. In: Stamp NE, Casey TM (eds) Caterpillars. Ecological and evolutionary constraints on foraging. Chapman and Hall, New York, pp 448–477
Jost L (2006) Entropy and diversity. Oikos 113:363–375. https://doi.org/10.1111/j.2006.0030-1299.14714.x
Klink CA, Machado RB (2005) Conservation of the Brazilian Cerrado. Conserv Biol 19:707–713. https://doi.org/10.1111/j.1523-1739.2005.00702.x
Koch EBA, Dáttilo W, Camarota F, Vasconcelos HL (2018) From species to individuals: does the variation in ant-plant networks scale result in structural and functional changes? Popul Ecol 60:309–318. https://doi.org/10.1007/s10144-018-0634-5
Kursar TA, Coley PD (2003) Convergence in defense syndromes of young leaves in tropical rainforests. Biochem Syst Ecol 31:929–949. https://doi.org/10.1016/S0305-1978(03)00087-5
Landi P, Minoarivelo HO, Brannstrom A et al (2018) Complexity and stability of ecological networks: a review of the theory. Popul Ecol 60:319–345. https://doi.org/10.1007/s10144-018-0628-3
Lange D, Dáttilo W, Del-Claro K (2013) Influence of extrafloral nectary phenology on ant-plant mutualistic networks in a neotropical savanna. Ecol Entomol 38:463–469. https://doi.org/10.1111/een.12036
Lepesqueur C, Scherrer S, Vieira MC et al (2018) Changing interactions among persistent species as the major driver of seasonal turnover in plant-caterpillar interactions. PLoS ONE 13:1–10. https://doi.org/10.1371/journal.pone.0203164
Lewis OT, Memmott J, Lasalle J et al (2002) Structure of a diverse tropical forest insect-parasitoid community. J Anim Ecol 71:855–873
Lill JT, Marquis RJ, Ricklefs RE (2002) Host plants influence parasitism of forest caterpillars. Nature 417:170. https://doi.org/10.1038/417170a
López-Carretero A, Díaz-Castelazo C, Boege K, Rico-Gray V (2014) Evaluating the spatio-temporal factors that structure network parameters of plant-herbivore interactions. PLoS ONE 9:e110430. https://doi.org/10.1371/journal.pone.0110430
Luna P, Dáttilo W (2021) Dientangling plant-animal interactions into complex networks: a multi-view approach and perspectives. In: Del-Claro K, Torezan-Silingardi (eds) Plant-animal interactions. Springer Nature Switzerland, Switzerland, pp 261–282
MacArthur R, Wilson EO (1967) The theory of Island biogeography. Princeton University Press, Princeton
Marquis RJ (1991) Herbivore fauna of Piper (Piperaceae) in a Costa Rican wet forest: diversity, specificity, and impact. In: Price PW, Lewinsohn TM, Fernandes GM, Benson WW (eds) Plant-animal interactions: evolutionary ecology in tropical and temperate regions. Wiley, New York, pp 179–203
Marquis RJ (2004) Biogeography of neotropical Piper. In: Dyer L, Palmer ADN (eds) Piper: a model genus for studies of phytochemistry, ecology, and evolution. Springer, Boston, pp 78–96
Marquis RJ, Diniz IR, Morais HC (2001) Patterns and correlates of interspecific variation in foliar insect herbivory and pathogen attack in Brazilian Cerrado. J Trop Ecol 17:127–148. https://doi.org/10.1017/S0266467401001080
Marquis RJ, Morais HC, Diniz IR (2002) Interactions among Cerrado plants and their herbivores: unique or typical? In: Oliveira PS, Marquis RJ (eds) The Cerrados of Brazil. Columbia University Press, New York, pp 306–328
Massad TJ, Trumbore SE, Ganbat G et al (2014) An optimal defense strategy for phenolic glycoside production in Populus trichocarpa—isotope labeling demonstrates secondary metabolite production in growing leaves. New Phytol 203:607–619
Meave J, Kellman M, Macdougall A, Rosales J (1991) Riparian habitats as tropical forest Refugia. Glob Ecol Biogeogr 1:69–76. https://doi.org/10.2307/2997492
MMA—Ministério do Meio. Ambiente (2020) O Bioma Cerrado. https://www.mma.gov.br/biomas/cerrado
Morais HC, Diniz IR, Silva DMS (1999) Caterpillar seasonality in a central Brazilian Cerrado. Rev Biol Trop 47:1025–1033
Morais HC, Sujii ER, Almeida-Neto M et al (2011) Host plant specialization and species turnover of caterpillars among hosts in the Brazilian Cerrado. Biotropica 43:467–472. https://doi.org/10.1111/j.1744-7429.2010.00736.x
Moura RF, Colberg E, Alves-Silva E et al (2021) Biotic defenses against herbivory. In: Del-Claro K, Torezan-Silingardi (eds) Plant-animal interactions. Springer Nature Switzerland, Switzerland, pp 93–118
Muller CB, Adriaanse ICT, Belshaw R, Godfray HCJ (1999) The structure of an aphid–parasitoid community. J Anim Ecol 68:346–370
Myers JH (1981) Interactions between western tent caterpillars and wild rose: a test of some general plant herbivore hypotheses. J Anim Ecol 50:11–25. https://doi.org/10.2307/4028
Myers N, Mittermeier RA, Mittermeier CG et al (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858. https://doi.org/10.1038/35002501
Oliveira, PS, Marquis, RJ (eds) (2002) The cerrados of Brazil: ecology and natural history of a neotropical savanna. Columbia University Press, New York
Oliveira GL, Moreira DD, Mendes AD, Guimarães EF, Figueiredo LS, Kaplan MA, Martins ER (2013) Growth study and essential oil analysis of Piper aduncum from two sites of Cerrado biome of Minas Gerais State, Brazil. Rev Bras 23:743–753. https://doi.org/10.1590/S0102-695X2013000500005
Oliveira-Filho AT, Ratter JA (1995) A study of the origin of central Brazilian forests by the analysis of plant species distribution patterns. Edinb J Bot 52:141–194. https://doi.org/10.1017/S0960428600000949
Penalba OC, Rivera JA (2013) Future changes in drought characteristics over Southern South America projected by a CMIP5 multi-model ensemble. Am J Clim Chang 2:173–182. https://doi.org/10.4236/ajcc.2013.23017
Poisot T, Canard E, Mouillot D et al (2012) The dissimilarity of species interaction networks. Ecol Lett 15:1353–1361. https://doi.org/10.1111/ele.12002
Poisot T, Stouffer DB, Gravel D (2015) Beyond species: why ecological interaction networks vary through space and time. Oikos 124:243–251. https://doi.org/10.1111/oik.01719
Preston FW (1962) The canonical distribution of commonness and rarity: part II. Ecology 43:410–432. https://doi.org/10.2307/1933371
Quijano-Abril MA, Callejas-Posada R, Miranda-Esquivel DR (2006) Areas of endemism and distribution patterns for neotropical Piper species (Piperaceae). J Biogeogr 33:1266–1278. https://doi.org/10.1111/j.1365-2699.2006.01501.x
Ratter JA, Ribeiro JF, Bridgewate S (1997) The Brazilian Cerrado vegetation and threats to its biodiversity. Ann Bot-London 80:223–230
Ribeiro JF, Walter BMT (2008) As Principais Fitofisionomias do Bioma Cerrado. In: Sano SM, de Almeida SP, Ribeiro JF (eds) Cerrado: Ecologia e Flora v. 2. EMBRAPA, Brasília, pp 151–212
Rizzini CT (1979) Tratado de fitogeografia do Brasil. Hucitec Edusp, São Paulo
Rodríguez-Castañeda G, Dyer LA, Brehm G et al (2010) Tropical forests are not flat: how mountains affect herbivore diversity. Ecol Lett 13:1348–1357. https://doi.org/10.1111/j.1461-0248.2010.01525.x
Sano EE, Rosa R, Brito JLS, Ferreira LG (2010) Land cover mapping of the tropical savanna region in Brazil. Environ Monit Assess 166:113–124. https://doi.org/10.1007/s10661-009-0988-4
Scherrer S, Diniz IR, Morais HC (2010) Climate and host plant characteristics effects on lepidopteran caterpillar abundance on Miconia ferruginata DC. and Miconia pohliana Cogn (Melastomataceae). Braz J Biol 70:103–109. https://doi.org/10.1590/S1519-69842010000100014
Scherrer S, Lepesqueur C, Vieira MC et al (2016) Seasonal variation in diet breadth of folivorous Lepidoptera in the Brazilian Cerrado. Biotropica 48:491–498. https://doi.org/10.1111/btp.12325
Shimbori EM (2009) Sistema hospedeiro-parasitoide associado à Piper glabratum Künth e P. mollicomum Künth (Piperaceae) no município de São Carlos, SP (Doctoral dissertation). Universidade Federal de São Carlos, São Carlos
Slinn HL, Richards LA, Dyer LA et al (2018) Across multiple species, phytochemical diversity and herbivore diet breadth have cascading effects on herbivore immunity and parasitism in a tropical model system. Front Plant Sci. https://doi.org/10.3389/fpls.2018.00656
Smilanich AM, Mason PA, Sprung L et al (2011) Complex effects of parasitoids on pharmacophagy and diet choice of a polyphagous caterpillar. Oecologia 165:995–1005. https://doi.org/10.1007/s00442-010-1803-1
Sörensson AA, Menendez CG, Ruscica R et al (2010) Projected precipitation changes in South America: a dynamical downscaling within CLARIS. Meteorol Z 19:347–355. https://doi.org/10.1127/0941-2948/2010/0467
Su YS, Yajima M (2020) R2jags: Using R to Run 'JAGS'. R package version 0.6-1. https://CRAN.R-project.org/package=R2jags. Accessed 24 July 2021
Tepe EJ, Rodriguez-Castaneda G, Glassmire AE, Dyer LA (2014) Piper kelleyi, a hotspot of ecological interactions and a new species from Ecuador and Peru. PhytoKeys 34:19–32. https://doi.org/10.3897/phytokeys.34.6376
Tsai T-H, Gill J, Rapkin J (2012) Superdiag: R code for testing Markov chain nonconvergence. R package version 1.1. https://CRAN.R-project.org/package=superdiag. Accessed 24 July 2021
Tylianakis JM, Laliberté E, Nielsen A, Bascompte J (2010) Conservation of species interaction networks. Biol Conserv 143:2270–2279. https://doi.org/10.1016/j.biocon.2009.12.004
Vilela AA, Torezan-Silingardi HM, Del-Claro K (2014) Conditional outcomes in ant–plant–herbivore interactions influenced by sequential flowering. Flora—Morphol, Distrib, Funct Ecol Plants 209:359–366. https://doi.org/10.1016/j.flora.2014.04.004
Vilela AA, Del Claro VTS, Torezan-Silingardi HM, Del-Claro K (2018) Climate changes affecting biotic interactions, phenology, and reproductive success in a savanna community over a 10-year period. Arthropod-Plant Interact 12:215–227. https://doi.org/10.1007/s11829-017-9572-y
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer, New York
Wiggins NL, Forrister DL, Endara M-J et al (2016) Quantitative and qualitative shifts in defensive metabolites define chemical defense investment during leaf development in Inga, a genus of tropical trees. Ecol Evol 6:478–492. https://doi.org/10.1002/ece3.1896
Yuncker TG (1972) The Piperaceae of Brazil: 1. Piper-group 1, 2, 3, 4. Hoehnea 2:21–366
Acknowledgements
We thank Vanessa Morais, Neuza A. P. Silva and Priscila M. C. da Luz for field and laboratory support and Vitor Becker for help in identifying the lepidopteran species.
Funding
RSR was supported by a master’s scholarship from CAPES; LAD and EJT were supported by NSF grant DEB-1442103 and DEB-1442075, respectively. IRD received financial support from CNPq and FAPDF, and CL was supported by a post-doctoral scholarship from CNPq.
Author information
Authors and Affiliations
Contributions
RSR designed the study, collected data, identified Lepidoptera, and helped write the manuscript; LAD designed the study, analyzed data, and wrote and edited the manuscript; CL collected data, identified Lepidoptera and edited the manuscript; TPS, PAR, and TBT collected data; IRD designed the study, identified Lepidoptera, and edited the manuscript; ARN identified parasitoids; DMS analyzed data and helped write and edit the manuscript; EJP identified Piper and edited the manuscript; TJM analyzed data and wrote and edited the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
All authors declare they have no conflict of interest or competing interests.
Consent for publication
All authors consent to publication.
Additional information
Handling Editors: Miriama Malcicka and Heikki Hokkanen.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Rabelo, R.S., Dyer, L.A., Lepesqueur, C. et al. Tritrophic interaction diversity in gallery forests: A biologically rich and understudied component of the Brazilian cerrado. Arthropod-Plant Interactions 15, 773–785 (2021). https://doi.org/10.1007/s11829-021-09856-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11829-021-09856-y