
Abstract
The current study deals with the diversity and distribution of culturable cold-adapted bacteria in Himalayan Thajwas glacial soil. The total bacterial count of the seven soil samples collected near glacier snout at different altitudes was investigated through a culture-dependent method. The highest cfu/g (6.8±2 × 107) was recorded at site I, and the lowest cfu/g (4.6±2 × 103) was recorded at site VII on nutrient agar (NA) media. The 31 isolated strains were identified using 16S rRNA gene sequencing. Based on their optimum growth at different temperatures, these bacterial isolates were categorized into two overlapping groups: psychrophiles (optimum growth ≥15°C) and psychrotrophs (optimum growth ≥20°C). Besides, these bacterial strains were screened for their cold-active enzyme activity. Further, the microbial diversity was affected by higher incubation temperature, which reduced the richness due to the selection of psychrotrophic microorganisms. Variation in microbial diversity may be related to soil physicochemical properties since a high positive correlation was observed with water content, total nitrogen and organic carbon. This is the first research to look at microbial communities from the Thajwas Glacier, and it adds to our knowledge of the microbial diversity found in Himalayan cold ecosystems.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The cryosphere is an important part of the global climate system and one of the earth’s largest habitable ecosystems. From polar regions to non-polar mountain glaciers and deep oceans, the cold environments cover approximately 75% of the earth’s surface (Margesin and Collins 2019; Bhatia et al. 2020). Cold habitats at permanently low temperatures (below 5°C) represent the largest portion of the earth’s biosphere (Margesin and Collins 2019). Among these glacial soil, permafrost and glaciers exhibit very extreme weather conditions and harbor the oldest microorganisms on Earth (Willerslev et al. 2004). Between these cold environments, the non-polar and polar glaciers and ice sheets are significant, as they almost cover about 10% of the earth’s surface (Anesio and Laybourn-Parry 2012) and host vast reservoir (9.61 × 1025 cells) of microorganisms (Priscu and Christner 2004; Priscu et al. 2007). Microbes that survive in these conditions near freezing point face several challenges, including persistently low temperatures, minimal nutrient availability, regular freeze-thaw cycles, desiccation, salinity fluctuations and different light conditions (Varin et al. 2012).
Glacial and high-altitude soils remain primarily understudied due to restricted accessibility and considerable heterogeneity hampers the comparability of different alpine studies. Adaptation of psychrophilic microorganisms enables them to thrive in harsh environmental conditions of alpine environments. Microbes in glacial soil are thought to be pioneers in the initial build-up of biomass, with allochthonous deposition, input from landslides, avalanches, ancient organic matter deposited in the earth and autotrophic fixation as possible sources of C and N (Schmidt et al. 2008; Brankatschk et al. 2011). The potential source of nutrients, such as phosphorus and sulfur, could be the weathered bedrocks (Frey et al. 2010). However, these different sources’ relative contributions remain a matter of ongoing debate for alpine glacier soils (Duc et al. 2009; Zumsteg et al. 2012; Frey et al. 2013; Rime et al. 2016; Donhauser and Frey 2018). The community structures of psychrophilic and psychrotrophic bacteria strongly correlate with climatic and soil properties and thus closely mirror the complexity and small-scale heterogeneity of alpine glacial soils (Srinivas et al. 2011; Donhauser and Frey 2018; Kumar et al. 2019).
Covering about 33,000 km2, the Himalayas is home to many glaciers outside the polar region. These glaciers prove extremely inhospitable to every life form, as the temperature over here falls well below -70°C during winters and rarely exceeds 0°C during summers (Gangwar et al. 2009; Shivaji et al. 2011). This seasonal temperature shift makes the region more intense for microbes, including the psychrophilic and psychrotrophic bacteria. The persistent UV radiation, low nutrient availability and metabolite transfer further add to the region's environmental hostility (Johnson et al. 2007; Steven et al. 2007; Gilichinsky et al. 2008; Zhang et al. 2013). However, notwithstanding such extreme conditions, the psychrophiles, owing to their adaptive mechanisms, still inhabit the Himalayan glaciers and other cold environments like ice and sediments (Srinivas et al. 2011; Rafiq et al. 2017). Apart from mere isolation, these bacteria have also been investigated for extracellular hydrolytic enzymes like proteases, amylases, lipases, cellulases and esterases (Al-Ghanayem and Joseph 2020; Bhatia et al. 2020). More recently, these life forms have been extensively investigated using an amalgamation of culture-dependent and culture-independent methods (Nemergut et al. 2007; Shivaji et al. 2011; Pradhan et al. 2010; Rafiq et al. 2017; Lauro et al. 2011; Sułowicz et al. 2020). In this study, the psychrophilic and psychrotrophic bacterial diversity of soil samples collected near the Thajwas glacier in the Northwestern Himalayas, India, was determined using a culture-dependent approach. Besides, the cold-active enzyme activity of these bacterial isolates was also tested. Based on their optimum growth temperature (OGT), these microorganisms were classified (Morita 1975) into two groups viz., psychrophiles (OGT of ≥15°C) and psychrotrophs (OGT of ≥20°C).
Materials and methods
Sample collection
Soil samples were collected from the Thajwas glacier, located in the Northwestern Himalayan region of Kashmir, between altitudes of 2944-3909 m (Fig. S1). The soil samples were collected near glacier snout in sterile poly bags and plastic vials (Shivaji et al. 2011). The soil samples were transported to the laboratory and stored at -20°C until further analysis. Besides, with the mercury bulb soil thermometer’s assistance, the temperature of the soil sample sites was recorded on site.
Physicochemical characteristics of soil
Standard methods were used for the physiochemical analysis of soil samples which included; soil moisture (Gardner 1986), soil pH (Watson and Brown 1998), total organic carbon (Walkley and Black 1934), and total Kjeldahl nitrogen (Bremner 1965). Each experiment was run in triplicates, and the average value was used.
Isolation of bacteria
Fourfold dilution was prepared in sterilized normal saline for every soil sample and three sets of triplicate were prepared for each dilution. The spread plate technique was used to spread 0.1 mL of inoculum on media plates. Each set of triplicate plates were incubated at three different temperatures (4, 20, and 37°C) on nutrient agar (NA) and Reasoner’s 2 agar (R2A) for 2-30 days, growth was observed after every 24 h for the new bacterial colony (Zhang et al. 2013). The number of culturable bacteria was estimated by counting the average colony-forming units (cfu/g) individually on NA and R2A agar plates. Morphologically distinct colonies were purified by streaking and maintained on NA plates (Shivaji et al. 2011). For determining the optimum growth temperatures, the isolate was inoculated on pre-prepared nutrient media plates followed by incubation at five different temperatures (4, 10, 15, 20, 37°C) and growth was observed every 24 h (Zhang et al. 2013).
Morphological characterization
The colony was observed for macro morphological characteristics like whole colony appearance (WCA), margin, elevation and shape (Rafiq et al. 2017). After that, the pure isolates were stained by Grams Stain-Kit K001 (Himedia® India) to determine their Grams reaction and the cell morphology was then studied under the light microscope (Olympus IX71).
Genomic DNA extraction and PCR amplification
The bacterial genomic DNA was extracted using the GenEluteTM bacterial genomic DNA kit (Sigma-Aldrich, USA). The molecular identification was done using 16S rRNA gene markers, F: 5ˈ- AGA GTT TGA TCC TGG CTC AG -3ˈand R: 5ˈ- GGT TAC CTT GTT ACG ACT T -3ˈ. Amplification was done by polymerase chain reaction (PCR) as described previously by Shivaji et al. (2004).
Sequencing and phylogenetic analysis
Sequencing of the amplified PCR products was done at Agrigenome laboratory Kerala, India, using ABI 3730XL sequencer. The sequenced data thus obtained was compared with the database available in the NCBI database using BLASTn. The phylogenetic tree was constructed in MEGA 7.0 software (Kumar et al. 2016), employing the neighbor-joining method.
Screening for different hydrolytic enzymes
The 31 identified isolates were screened for their extracellular enzymatic activity on different media. The isolates were cultured on pre-prepared media plates containing NA with one of the following substrates; skim milk 1% (w/v), starch 0.2% (w/v), carboxymethyl cellulose (CMC) 1% (w/v), tributyrin 1% (v/v) and urea 1% (w/v), for protease, amylase, cellulase, lipase and urease respectively. The inoculated plates were incubated at 10, 15, and 20°C for 5-20 days (Alam and Singh 2002). After incubation, the colonies of isolates were examined for zones of hydrolysis (clear halos) on skim milk, tributyrin, CMC (after being flooded with congo-red solution) and starch (after being flooded with iodine solution) agar plates for protease, lipase, cellulase and amylase activities, respectively. Urease activity was determined by observing the development of pink color around the bacterial colonies.
Statistical analysis
The computation of Shannon’s diversity index was done by using the PAST 4.03 statistical application (Hammer et al. 2001). SPSS software was used to determine the relationship between soil physicochemical parameters and bacteria diversity in glacial soil samples using the Pearson correlation (rp) matrix. Further, one-way ANOVA was used to calculate the significance of difference among different sites for physicochemical parameters.
GenBank accession numbers
The nucleotide sequences of the 16S rRNA gene were deposited at NCBI GenBank under accession numbers: MN102377 to MN102391, MT478140, MT478142 to MT478150, MT477765, MT478875 and MT495407 to MT495410.
Results
Isolation and enumeration of cultivable psychrophilic bacteria
Enumeration of viable bacteria was done after analyzing NA plate’s growth at 4, 20 and 37°C, while as only at 4°C on R2A agar (Table 1). The highest bacterial count (6.8±2 × 107 cfu/g) was recorded at site I at 4°C and the lowest bacterial count (4.6±2 × 103 cfu/g) was recorded for site VII at 37°C (incubation temperature) on NA. The highest cfu/g (5.2±2 × 107) on R2A agar was recorded at site I and the lowest (1.8±1 × 105) at site VII. Isolates able to grow at 4°C preferred R2A media while NA was preferred at all three (4, 20, and 37°C) incubation temperatures. Besides, these bacterial strains showed growth at a different range of temperatures. Out of 31 isolates, 27 (79.41%) isolates were able to grow at 4°C while only 11 could grow at 37°C. All those isolates that could grow at 4°C also showed growth at 20°C. Based on their growth at different temperatures, these bacterial isolates were categorized into two overlapping groups: psychrophiles (optimum growth ≥15°C) and psychrotrophs optimum growth ≥20°C) (Supplementary table S1).
Morphological and microscopic identification of bacterial strains
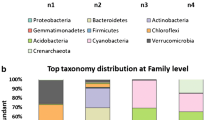
The isolated bacterial strains were first categorized based on morphological and physiological characteristics like Gram’s stain, WCA, margin, elevation and shape (Supplementary table S1). Further, the bacterial isolates were placed in two distinct groups: Gram-positive and Gram-negative bacteria. Gram-positive were more prominent with 25 (81%) isolates than Gram-negative with only six isolates (19%). The psychrophilic and psychrotrophic bacterial diversity of Thajwas glacial soil comprised of 4 phyla viz., Firmicutes, ϒ-Proteobacteria, Actinobacteria, and β-Proteobacteria. Firmicutes dominated the overall bacterial diversity, followed by ϒ-Proteobacteria, Actinobacteria and β-Proteobacteria (Fig. 1a). In Firmicutes phyla, the maximum number of species was recorded at all seven sites that showed a slight decrease with increasing altitude. Three (Firmicutes, ϒ-Proteobacteria, and Actinobacteria) of the four phyla were present at all sites, while β-Proteobacteria was absent at sites IV, V, and VII (Fig. 1c). A gradual decline in the number of species from site I to site V was observed, with a rise in altitude.

a Overall distribution of phyla b Screening of bacterial phyla for their cold-active enzymes c Site wise distribution of phyla.
Molecular characterization
The 16S rRNA nucleotide sequences of all the 31 isolates displayed close similarity (100-98%) with respective species after BLASTn search at NCBI. Phylogenetic analysis of 31 isolates based on 16S rRNA gene nucleotide sequences revealed substantial similarities with other highly homologous species by sharing the same branch in the phylogenetic tree. Each similar specie's strain name is in front of the species name (Figs. 2 and 3).

16S rRNA phylogenetic tree (Neighbor-joining) showing the phylogenetic relationship of Proteobacteria affiliated species, Actinobacteria affiliated species and bootstrap values were 1000

16S rRNA phylogenetic tree (Neighbor-joining) showing the phylogenetic relationship of phyla Firmicutes affiliated species and bootstrap values were 1000
Screening of psychrophilic and psychrotrophic bacteria for their cold-active enzyme activity
The enzymatically positive species were dominated by Firmicutes, which exhibited activity for all 5 enzymes tested (Fig. S2). It was followed by species belonging to ϒ-Proteobacteria, which were found positive for protease, lipase and cellulose at their respective optimal temperatures. The species of phyla Actinobacteria showed the protease and urease activity while no enzyme activity was observed in β-Proteobacteria phyla, which consisted of only single species. Proteases and lipases were found to be equally dominant (Fig. 1b).
Shannon-Wiener diversity index
Shannon-Wiener bacterial diversity index of seven sampling sites (Fig. 4a) showed the highest diversity (2.95) at site I followed by site II (2.77) and the lowest diversity was observed at site V (1.99). As shown in Table 2, temperature-dependent diversity indices showed low Shannon diversity and evenness at 37°C and highest at 20°C, so it could be concluded that higher temperatures restricted the growth of psychrophilic microorganisms (unable to develop above 20°C) and only the psychrotrophic portion of the population was active at 37°C, resulting in lower diversity at 37°C.

a Shannon-Wiener diversity index of seven sampling sites b Correlation between different physicochemical parameters and bacterial count obtained from glacial soil of Thajwas glacier where size and numerical values inscribed inside the bubble determines the type of correlation (numerical values inscribed on horizontal and vertical axis designate 1: Temperature; 2: Organic carbon; 3: Total nitrogen; 4: pH; 5: Water content; 6: Number of species)
Bacterial diversity influencing factors
The number of psychrophilic and psychrotrophic bacterial species isolated from glacial soil showed a high positive correlation with water content (0.841), total nitrogen (0.918) and organic carbon (0.898), respectively. Temperature showed a medium positive correlation (0.406); however, pH showed a negative correlation (-0.772) with bacterial diversity of glacial soil (Fig. 4b).
Soil physicochemical characteristics
The average on-site temperature for the seven sampling sites varied between 0-1.3°C. The lowest average temperature of 0°C was recorded at site III, while the highest on-site average temperature of 1.3°C was recorded at sites II, V and VI (Fig. 5a). The physicochemical characteristics of seven glacial soil samples showed variation except for the soil pH, where a slight variation was observed. The pH value ranged from 7.44 to 8.04 (Fig. 5b). The highest pH was recorded for site VII (8.04) and the lowest was recorded for site II (7.35). The water content showed a greater variation between 61.37 to 11.44%. It was recorded highest for sites I, II and V; lowest for sites III, IV, VI and VII (Fig. 5c). Total nitrogen and organic carbon assessment showed that the soil overall had a lower content. Further, the total nitrogen in the glacial soil samples from higher altitudes was found below the detection level (Fig. 5d). Organic carbon concentration was comparatively higher for low altitude sites, which decreased swiftly with increasing altitude (Fig. 6). Physiochemical parameters were analyzed for seven different sampling sites of Thajwas glacial soil by one-way ANOVA. pH (p<0.005), water content (%) (p<0.001), total nitrogen (%) (p<0.001), organic carbon (p<0.001) showed significant difference among different sites however, temperature (P>0.1) was not significantly different among all sites (Supplementary table S2).

Site-wise average a On-site temperature b pH c Water content (%) d Total nitrogen (%) in the Thajwas glacial soils

Site-wise mean organic carbon (%) content of glacial soil samples
Discussion
The bacterial abundance of Himalayan glacial soils based on culture-independent technique is reported in the range 0.9 × 107 to 30.71 × 108 bacteria/g (Pradhan et al. 2010; Shivaji et al. 2011; Srinivas et al. 2011). Based on the culture-dependent technique, the viable count is reported in the range 2.0 × 101 to 1.53 × 109 cfu/g (Gangwar et al. 2009; Rafiq et al. 2017; Sherpa et al. 2018). The soil samples of the Thajwas glacier had a viable count ranged from 6.8±2 × 107 to 4.6±2 × 103 cfu/g at 4 and 37°C, respectively. When using a culture-dependent technique, the cfu/g varies with incubation temperature; for example, a high viable count was recorded below 20°C, which decreased above 20°C. So far, the number of culturable species found in the soils of Himalayan glaciers has been limited. Some studies have reported 20 (Shivaji et al. 2011), 11 (Srinivas et al. 2011) and 24 (Rafiq et al. 2017), ours is the highest with 31 different species. Further, among 4 genera (Firmicutes, ϒ-Proteobacteria, β-Proteobacteria, and Actinobacteria), Firmicutes and Proteobacteria were seen to be dominant and similar observations were made in other culture-based studies on Himalayan glacial soils (Gangwar et al. 2009; Shivaji et al. 2011; Srinivas et al. 2011; Rafiq et al. 2017).
Studies on psychrophilic and psychrotrophic bacterial isolation from cold-habitats show that NA and R2A are the more suitable media for their isolation (Zhang et al. 2013; Rafiq et al. 2017; Hassan et al. 2020). Incubation on R2A at lower temperatures has already been shown to facilitate the development of rare, slow-growing bacteria, resulting in higher levels of diversity (Reasoner and Geldreich 1985; Wernicke et al. 1990; Reasoner 2004; Zhang et al. 2013; Gensberger et al. 2015). Because the bacterial count on R2A plates at 20 and 37°C was too low to count (TFTC), only the 4°C counts were considered. Furthermore, Reasoner and Geldreich (1985) developed R2A, which was categorized as low nutrient media since it contained lower carbon concentration and ionic strength (Reasoner and Geldreich 1985; Allen et al. 2004) than high nutritional medium (e.g., NA). This could also explain why some isolates were unable to grow at higher temperatures on R2A.
Besides, Gram-positive bacteria are more dominant than Gram-negative bacteria (Gangwar et al. 2009; Zhang et al. 2009; Pradhan et al. 2010; Shivaji et al. 2011) in Himalayan glacial soils. Since the survival of psychrotrophic bacteria in extremely stressful circumstances is attributable to the development of spores in Gram-positive bacteria (Miteva and Brenchley 2005; Rafiq et al. 2017). These results support our research as psychrotrophic and spore formers were the majority of the Gram-positive bacteria in our study. The development of spores may help resolve low-temperature stress conditions (Nicholson et al. 2000; Rafiq et al. 2017).
Although diversity was lower at 37°C (incubation temperature) in our samples, as shown by the Shannon diversity index and evenness (Table 2), it could therefore be inferred that the lower diversity observed at 37°C is explained by lower richness, i.e., lesser number of microorganism functional classes. Since higher temperatures limit the growth of psychrophilic (cold-adapted) microorganisms that are unable to grow above 20°C, only a small portion of the psychrotrophic (cold-tolerant) population was active at 37°C, resulting in lower diversity (Pessi et al. 2012). Besides, because of the maximum decrease in diversity observed at 37°C, it can be inferred from these results that the populations examined here are predominantly composed of psychrotrophic microorganisms since higher diversity was observed at 20°C than 4°C (Table 2), which tends to be the important criteria of classification for bacterial communities of the Himalayan and other polar glaciers (Yergeau and Kowalchuk 2008; Shivaji et al. 2011; Srinivas et al. 2011; Pessi et al. 2012). In cold environments, the prevalence of psychrotrophic microorganisms may be associated with an increase in soil temperature through solar radiation, especially during the summer season, when soil temperatures may rise above 9-15°C (Möller and Dreyfuss 1996; Liu et al. 2009).
The results also indicated low species richness and decreased bacterial count with increasing altitude, and this might be due to low nitrogen and organic carbon in the glacial soil. Similar observations were also made previously in several studies that showed that nutrient availability (nitrogen and carbon content) greatly influences microbial diversity and composition (Fierer et al. 2003; LaMontagne et al. 2003; Fierer et al. 2007; Pradhan et al. 2010; Pessi et al. 2012). The decreased bacterial diversity with increased altitude can be related to the fact that the glacial soil at higher altitudes lacks traceable amounts of carbon and nitrogen (Gangwar et al. 2009), which could otherwise provide the suitable media constituents for their growth.
Himalayan glaciers offer a vast resource of unexplored cold-habitats that are rich in psychrophilic and psychrotrophic bacterial species and are exposed to one of the world’s harshest cold environments. Under such high-stress conditions, these psychrophilic and psychrotrophic bacteria produce extracellular cold-active enzymes to get acclimatized to such environment (Shivaji et al. 2011; Srinivas et al. 2011; Rafiq et al. 2017). These enzymes can thus be explored for their industrial application since they are active even at low temperatures and can decrease industrial energy consumption thereby reducing the environmental pollution. Among these enzymes, the cold-active proteases, lipases, amylases, urease and cellulase are vital for industrial uses because they possess high catalytic activity at lower temperatures (Gangwar et al. 2011; Kuddus and Ramteke 2012; Singh et al. 2014; Farooq et al. 2021) and have gained much attention in the effort to decrease the industrial energy costs and pollution. Cold-active enzymes are emerging as the first choice for several industrial applications particularly in the detergent, dehairing, food and pharmaceutical industry as these enzymes can be exploited to replace the ones derived from mesophilic organisms since the former have more economic benefits by decreasing the industrial energy expenditures, pollution and costs than the later (Al-Ghanayem and Joseph 2020). Previous studies have also used skim milk (Farooq et al. 2021; Gangwar et al. 2011; Alam and Singh 2002), tributyrin (Maharana and Ray 2015; Salwoom et al. 2019; Gangwar et al. 2011), starch (Gangwar et al. 2011; Kuddus et al. 2012; Singh et al. 2014), CMC (Gangwar et al. 2011; Singh et al. 2014) and urea (Singh et al. 2014) as substrates and methods for screening for protease, lipase, amylase, cellulase and urease activity respectively, in psychrophilic and psychrotrophic bacterial species.
Conclusion
Firmicutes were the common bacterial phyla besides organic carbon and nitrogen were probably the key parameters that influenced the bacterial diversity. Comparing the Thajwas glacial soil’s bacterial diversity with the diversity of other cold ecosystems in the Himalayas showed that the taxa distribution and abundance differ. However, the similarity was noticed for Gram-positive psychrotrophic dominance. The discovery of these psychrotrophic/psychrophilic and their genetic engineering will offer novel opportunities for biocatalysis and biotransformation in biotechnology. Cold-adapted enzymes screened from these bacterial isolates can be exploited to replace the enzymes derived from mesophilic organisms as the former has more economic benefits by decreasing the industrial energy expenditures and costs than the latter.
Data availability
Data about the 16S rRNA gene sequences have been deposited at NCBI GenBank under the accession numbers: MN102377-91, MT478140, MT478142-50, MT477765, MT478875, and MT495407-10.
Code availability
Not applicable.
Abbreviations
- BLAST:
-
Basic Local Alignment Search Tool
- CFU:
-
Colony forming units
- CMC:
-
Carboxymethyl cellulose sodium salt
- DNA:
-
Deoxyribonucleic acid
- MEGA:
-
Molecular Evolutionary Genetic Analysis
- NA:
-
Nutrient agar
- NCBI:
-
National Center for Biotechnology Information
- OGT:
-
Optimum growth temperature
- PAST:
-
PAleontological Statistics
- PCR:
-
Polymerase chain reaction
- R2A:
-
Reasoner’s 2 agar
- rRNA:
-
Ribosomal Ribonucleic acid
- SPSS:
-
Statistical Package for the Social Sciences
- USA:
-
United States of America
- WCA:
-
Whole colony appearance
References
Al-Ghanayem AA, Joseph B (2020) Current prospective in using cold-active enzymes as eco-friendly detergent additive. Appl Microbiol Biotechnol 104:2871–2882. https://doi.org/10.1007/s00253-020-10429-x
Alam SI, Singh L (2002) Proteolytic heterotrophic bacteria of cyanobacterial assemblage from Schirmacher oasis, Antarctica, capable of growing under extreme conditions. Curr Sci 83:1000–1004
Allen MJ, Edberg SC, Reasoner DJ (2004) Heterotrophic plate count bacteria–what is their significance in drinking water? Int J Food Microbiol 92:265–274. https://doi.org/10.1016/j.ijfoodmicro.2003.08.017
Anesio AM, Laybourn-Parry J (2012) Glaciers and ice sheets as a biome. Trends Ecol Evol 27:219–225. https://doi.org/10.1016/j.tree.2011.09.012
Bhatia RK, Ullah S, Hoque MZ et al (2020) Psychrophiles: a source of cold-adapted enzymes for energy efficient biotechnological industrial processes. J Environ Chem Eng 9(1):104607. https://doi.org/10.1016/j.jece.2020.104607
Brankatschk R, Towe S, Kleineidam K et al (2011) Abundances and potential activities of nitrogen cycling microbial communities along a chronosequence of a glacier forefield. ISME J 5(6):1025–37. https://doi.org/10.1038/ismej.2010.184
Bremner JM (1965) Total nitrogen. In: Methods of soil analysis, part 2: chemical and microbiological properties. Wiley, pp 1149–1178
Donhauser J, Frey B (2018) Alpine soil microbial ecology in a changing world. FEMS Microbiol Ecol 94(9):9. https://doi.org/10.1093/femsec/fiy099
Duc L, Noll M, Meier BE et al (2009) High diversity of diazotrophs in the forefield of a receding alpine glacier. Microbial Ecol 57:179–90. https://doi.org/10.1007/s00248-008-9408-5
Farooq S, Nazir R, Ganai SA, Ganai BA (2021) Isolation and characterization of a new cold-active protease from psychrotrophic bacteria of Western Himalayan glacial soil. Sci Reports 11:1–14. https://doi.org/10.1038/s41598-021-92197-w
Fierer N, Allen AS, Schimel JP, Holden PA (2003) Controls on microbial CO2 production: A comparison of surface and subsurface soil horizons. Glob Chang Biol 9:1322–1332. https://doi.org/10.1046/j.1365-2486.2003.00663.x
Fierer N, Bradford MA, Jackson RB (2007) Toward an ecological classification of soil bacteria. Ecology 88:1354–1364. https://doi.org/10.1890/05-1839
Frey B, Rieder SR, Brunner I et al (2010) Weathering-associated bacteria from the Damma glacier forefield: physiological capabilities and impact on granite dissolution. Appl Environ Microbiol 76(14):88–96. https://doi.org/10.1128/AEM.00657-10
Frey B, Buhler L, Schmutz S et al (2013) Molecular characterization of phototrophic microorganisms in the forefield of a receding glacier in the Swiss Alps. Environ Res Lett 8:1–11. https://doi.org/10.1088/1748-9326/8/1/015033
Gangwar P, Alam SI, Bansod S, Singh L (2009) Bacterial diversity of soil samples from the western Himalayas, India. Can J Microbiol 55:564–577. https://doi.org/10.1139/W09-011
Gangwar P, Alam SI, Singh L (2011) Metabolic Characterization of cold active Pseudomonas, Arthrobacter, Bacillus, and Flavobacterium spp. from Western Himalayas. Indian J Microbiol 51:70–75. https://doi.org/10.1007/s12088-011-0092-7
Gardner WH (1986) Water content. pp 493–594. In: A Klute (ed) Methods of soil analysis. Part 1. Physical and mineralogical methods. Agron. Monogr., 9. 2nd edn. ASA, CSSA, and SSSA, Madison
Gensberger ET, Gössl E-M, Antonielli L et al (2015) Effect of different heterotrophic plate count methods on the estimation of the composition of the culturable microbial community. PeerJ 3:e862. https://doi.org/10.7717/PEERJ.862
Gilichinsky D, Vishnivetskaya T, Petrova M et al (2008) Bacteria in permafrost Psychrophiles: from Biodiversity to Biotechnology Margesin R, Schinner F, Marx J-C, Gerday C (eds). Springer-Verlag: Berlin Heidelberg, pp 83–102
Hammer DAT, Ryan PD, Hammer Ø, Harper DAT (2001) Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 4(1):9
Hassan N, Anesio AM, Rafiq M et al (2020) Temperature driven membrane lipid adaptation in glacial psychrophilic bacteria. Front Microbiol 11:824. https://doi.org/10.3389/FMICB.2020.00824
Johnson SS, Hebsgaard MB, Christensen TR et al (2007) Ancient bacteria show evidence of DNA repair. Proc Natl Acad Sci USA 104:14401–14405. https://doi.org/10.1073/pnas.0706787104
Kuddus M, Ramteke PW (2012) Recent developments in production and biotechnological applications of cold-active microbial proteases. Crit Rev Microbiol 38:330–338. https://doi.org/10.3109/1040841X.2012.678477
Kuddus M, Roohi Saima, Ahmad IZ (2012) Cold-active extracellular α-amylase production from novel bacteria Microbacterium foliorum GA2 and Bacillus cereus GA6 isolated from Gangotri glacier, Western Himalaya. J Genet Eng Biotechnol 10:151–159. https://doi.org/10.1016/j.jgeb.2012.03.002
Kumar S, Stecher G, Tamura K et al (2016) MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol 33:1870–1874. https://doi.org/10.1093/molbev/msw054
Kumar S, Suyal DC, Yadav A et al (2019) Microbial diversity and soil physiochemical characteristic of higher altitude. PLoS One 14:1–15. https://doi.org/10.1371/journal.pone.0213844
Lauro FM, Allen MA, Wilkins D et al (2011) Psychrophiles: Genetics, genomics, evolution. In: Extremophiles Handbook. Japan: Springer, pp 865–890.
LaMontagne MG, Schimel JP, Holden PA (2003) Comparison of subsurface and surface soil bacterial communities in California grassland as assessed by terminal restriction fragment length polymorphisms of PCR-amplified 16S rRNA genes. Microb Ecol 46:216–227. https://doi.org/10.1007/s00248-003-1006-y
Liu Y, Yao T, Jiao N et al (2009) Bacterial diversity in the snow over Tibetan Plateau Glaciers. Extremophiles 13:411–423. https://doi.org/10.1007/s00792-009-0227-5
Maharana A, Ray P (2015) A novel cold-active lipase from psychrotolerant Pseudomonas sp. AKM-L5 showed organic solvent resistant and suitable for detergent formulation. J Mol Catal B Enzym 120:173–178. https://doi.org/10.1016/J.MOLCATB.2015.07.005
Margesin R, Collins T (2019) Microbial ecology of the cryosphere (glacial and permafrost habitats): current knowledge. Appl. Microbiol. Biotechnol. 103:2537–2549. https://doi.org/10.1007/s00253-019-09631-3
Miteva VI, Brenchley JE (2005) Detection and isolation of ultrasmall microorganisms from a 120,000-year-old Greenland glacier ice core. Appl Environ Microbiol 71:7806–7818. https://doi.org/10.1128/AEM.71.12.7806-7818.2005
Möller C, Dreyfuss MM (1996) Microfungi from Antarctic lichens, mosses and vascular plants. Mycologia 88:922–933. https://doi.org/10.1080/00275514.1996.12026732
Morita RY (1975) Psychrophilic bacteria. Bacteriological Reviews 39:144–167
Nemergut DR, Anderson SP, Cleveland CC et al (2007) Microbial community succession in an unvegetated, recently deglaciated soil. Microbiol Ecol 53:110–122. https://doi.org/10.1007/s00248-006-9144-7
Nicholson WL, Munakata N, Horneck G et al (2000) Resistance of bacillus endospores to extreme terrestrial and extraterrestrial environments. Microbiol Mol Biol Rev 64:548–572. https://doi.org/10.1128/mmbr.64.3.548-572.2000
Pessi IS, Elias SdeO, Simões FL et al (2012) Functional diversity of microbial communities in soils in the vicinity of Wanda Glacier, Antarctic Peninsula. Microbes Environ 27:200–203. https://doi.org/10.1264/jsme2.ME11311
Pradhan S, Srinivas TNR, Pindi PK et al (2010) Bacterial biodiversity from Roopkund Glacier, Himalayan mountain ranges, India. Extremophiles 14:377–395. https://doi.org/10.1007/s00792-010-0318-3
Priscu JC, Christner BC (2004) Earth’s icy biosphere. In: Bull AT (ed) Microbial diversity and bioprospecting. ASM Press, Washington, DC, pp 130–145
Priscu JC, Christner BC, Foreman CM et al (2007) Biological material in ice cores. In: Encyclopedia of quaternary sciences. Elsevier
Rafiq M, Hayat M, Anesio AM et al (2017) Recovery of metallo-tolerant and antibiotic resistant psychrophilic bacteria from Siachen glacier, Pakistan. PLoS One 12(7):e0178180. https://doi.org/10.1371/journal.pone.0178180
Reasoner DJ (2004) Heterotrophic plate count methodology in the United States. Int J Food Microbiol 92:307–315. https://doi.org/10.1016/j.ijfoodmicro.2003.08.008
Reasoner DJ, Geldreich EE (1985) A new medium for the enumeration and subculture of bacteria from potable water. Applied Environ Microbiol 49(1):1–7
Rime T, Hartmann M, Frey B (2016) Potential sources of microbial colonizers in an initial soil ecosystem after retreat of an alpine glacier. ISME J 10:25–41. https://doi.org/10.1038/ismej.2015.238
Salwoom L, Rahman RNZRA, Salleh AB et al (2019) Isolation, characterisation, and lipase production of a cold-adapted bacterial strain Pseudomonas sp. LSK25 isolated from Signy Island, Antarctica. Molecules 24:1–14. https://doi.org/10.3390/molecules24040715
Schmidt SK, Reed SC, Nemergut DR et al (2008) The earliest stages of ecosystem succession in high-elevation (5000 metres above sea level), recently deglaciated soils. Proc R Soc B 275:2793–2802. https://doi.org/10.1098/rspb.2008.0808
Sherpa MT, Najar IN, Das S, Thakur N (2018) Bacterial diversity in an alpine debris-free and debris-cover accumulation zone Glacier Ice, North Sikkim, India. Indian J Microbiol 58:470–478. https://doi.org/10.1007/s12088-018-0747-8
Shivaji S, Reddy GSN, Aduri RP et al (2004) Bacterial diversity of a soil sample from Schirmacher Oasis, Antarctica. Cell Mol Biol 50:525–536.
Shivaji S, Pratibha MS, Sailaja B et al (2011) Bacterial diversity of soil in the vicinity of Pindari glacier, Himalayan mountain ranges, India, using culturable bacteria and soil 16S rRNA gene clones. Extremophiles 15:1–22. https://doi.org/10.1007/s00792-010-0333-4
Singh P, Singh SM, Dhakephalkar P (2014) Diversity, cold active enzymes and adaptation strategies of bacteria inhabiting glacier cryoconite holes of High Arctic. Extremophiles 18:229–242. https://doi.org/10.1007/s00792-013-0609-6
Srinivas TNR, Singh SM, Pradhan S et al (2011) Comparison of bacterial diversity in proglacial soil from Kafni Glacier, Himalayan Mountain ranges, India, with the bacterial diversity of other glaciers in the world. Extremophiles 15:673–690. https://doi.org/10.1007/s00792-011-0398-8
Steven B, Briggs G, McKay CP et al (2007) Characterization of the microbial diversity in a permafrost sample from the Canadian high Arctic using culture-dependent and culture-independent methods. FEMS Microbiol Ecol 59:513–523. https://doi.org/10.1111/j.1574-6941.2006.00247.x
Sułowicz S, Bondarczuk K, Ignatiuk D et al (2020) Microbial communities from subglacial water of naled ice bodies in the forefield of Werenskioldbreen, Svalbard. Sci Total Environ 723:138025. https://doi.org/10.1016/j.scitotenv.2020.138025
Varin T, Lovejoy C, Jungblut AD et al (2012) Metagenomic analysis of stress genes in microbial mat communities from Antarctica and the High Arctic. Appl Environ Microbiol 78(2):549–559. https://doi.org/10.1128/AEM.06354-11
Walkley A, Black TA (1934) An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci 37:29–38
Watson ME, Brown JR (1998) pH and Lime Requirement. In: Brown JR (ed) Recommended Chemical Soil Test Procedures for the North Central Region, NCR Publication No. 221, Missouri Agricultural Experiment Station, Columbia, MO, 13-16
Wernicke F, Kampfer P, Dott W (1990) Influence of media and conditions on the isolation of bacterial species from drinking water. Zentralblatt fur Hygiene 190(1–2):26–38
Willerslev E, Hansen AJ, Ronn R et al (2004) Long-term persistence of bacterial DNA. Curr Biol 14(1):9–10. https://doi.org/10.1016/j.cub.2003.12.012
Yergeau E, Kowalchuk GA (2008) Responses of Antarctic soil microbial communities and associated functions to temperature and freeze-thaw cycle frequency. Environ Microbiol 10:2223–2235. https://doi.org/10.1111/j.1462-2920.2008.01644.x
Zhang LM, Wang M, Prosser JI et al (2009) Altitude ammonia-oxidizing bacteria and archaea in soils of Mount Everest. FEMS Microbiol Ecol 70:208–217. https://doi.org/10.1111/j.1574-6941.2009.00775.x
Zhang DC, Brouchkov A, Griva G et al (2013) Isolation and characterization of bacteria from Ancient Siberian Permafrost Sediment. Biology (Basel) 2:85–106. https://doi.org/10.3390/biology2010085
Zumsteg A, Luster J, Göransson H et al (2012) Bacterial, archaeal and fungal succession in the forefield of a receding glacier. Microb Ecol 63:552–64. https://doi.org/10.1007/s00248-011-9991-8
Author information
Authors and Affiliations
Contributions
SF framed the work design, collected, compiled and interpreted the data. Further, conducted experiments, wrote the manuscript and designed the figures. RN designed the research work, evaluated the data and did the revision of the manuscript. BAG and HM did the critical revision of the manuscript. GJD helped in statistical analysis. Both RN and BAG helped in designing the experiments and supervised the overall work. All the authors helped in writing the manuscript and provided critical feedback.
Corresponding author
Ethics declarations
Ethical statement
The authors in the current study carried out no animal or human studies. Authors further confirm that this work is original and has not been published elsewhere, nor is it currently under consideration for publication elsewhere.
Consent to participate
Consent was obtained from all the authors for the submission of the manuscript to the journal.
Consent for publication
The publication of this manuscript has been approved by all the authors.
Conflict of interest
The authors report no conflict of interest.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Farooq, S., Nazir, R., Ganai, B.A. et al. Psychrophilic and psychrotrophic bacterial diversity of Himalayan Thajwas glacial soil, India. Biologia 77, 203–213 (2022). https://doi.org/10.1007/s11756-021-00915-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11756-021-00915-6