
Abstract
Drought is a harmful abiotic factor that clearly affects the growth and production of plants. The negative impacts of drought can be reduced by ascorbic acid (AsA) which is an important non-enzymatic antioxidant in plants. The purpose of the current investigation was to study the effects of different levels of drought and the external application of AsA on sweet pepper plants. The experiment had two factors, the first factor was drought: irrigation within the field capacity, moderate stress (irrigation within the 60% field capacity) and severe stress (irrigation within the 30% field capacity). The second factor was AsA: 0 mM sprayed with distilled water, 0.5 and 1 mM. The experiment had three replications. The results illustrated that drought stress reduced the fresh and dry weight of roots and shoots. It also reduced the relative water content (RWC), diameter and length of fruit, chlorophyll content and leaf area. Drought stress increased the activity of polyphenol oxidase and ascorbate peroxidase enzymes. It caused improvements in antioxidant properties, total phenolic content, ascorbate and electrical conductivity (EC) in the leaves of sweet pepper. Higher concentrations of AsA by foliar application significantly increased the values of all measured parameters except the EC. The interaction between ascorbic acid and drought stress showed that ascorbic acid significantly increased the shoot fresh weight, root dry weight, antioxidant properties, ascorbate, polyphenol oxidase and ascorbate peroxidase. However, it reduced the electrical conductivity. It is claimed that sweet pepper plants acquire tolerance to drought because AsA contributes to an increase in antioxidant properties.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Drought stress is a widespread environmental factor that causes anatomical, physio-biochemical changes in plants (Kasim et al. 2017). Relative water content can be noted from different aspects of physiological changes when being affected by drought stress (Tuna et al. 2010). It can induce various morphological responses that result in reduced leaf area (Hu and Xiong 2014), as well as a loss of fresh and dry weight of the roots and shoots (Kasim et al. 2017). In fact, drought stress reduces biomass by retarding cell growth (Xu et al. 2015). As an abiotic stress, drought can hamper the mechanism of photosynthesis in plants by changing the inner and outer structure of plant tissues (Mahmoud 2014) and the concentration of different pigments and biological molecules such as enzymes involved in this mechanism (Ashraf and Harris 2013).
It is notable that drought stress can lead to the formation of active oxygen species, including superoxide (O−2) and hydrogen peroxide (H2O2) radicals (Xu et al. 2015). Through increased leakage of electrons by molecular oxygen, it follows oxidative damage to the photosynthetic device and distorts vital cellular macromolecules (Kolarovic et al. 2009). Plants synthesize several enzymatic antioxidants and non-enzymatic antioxidants that defenses to protect them from oxidative injuries caused by ROS (Ashraf 2009; Shafiq et al. 2014; Bayat and Moghadam 2019). The harmful effects of H2O2 can be reduced by ascorbate peroxidase (APX).
Non-enzymatic antioxidant such as AsA, have remarkable antioxidant properties that act as electron donors and decreased accumulation of ROS (Noctor and Foyer 1998). They also perform in substructure reactions in the cycle of enzyme production (Mittler 2002). Research on the use of plant growth regulators has partly been directed at overcoming the problems of drought stress (Farooq et al. 2009). The available literature cites the importance of compounds with low molecular weight. These compounds include AsA (vitamin C), glutathione (GSH), carotenoids and flavonoids (Seminario et al. 2017). AsA plays a key role in stress signals and in other physiological mechanisms (Wolucka et al. 2005) such as the regulation of cell division and photosynthesis (Smirnoff 1995). In addition, the free accumulation of AsA in plant cells is very important for ROS elimination (Xu et al. 2015) and possibly for making plants more tolerant to photosensitive oxidative stress (Foyer et al. 1993; Smirnoff 1995). Since AsA plays a role in both enzymatic and non-enzymatic Mechanisms as well as in other important functions in plants, studies have been implemented on the physiological functions of leaf tissue in some plants treated with AsA when facing drought stress (Athar et al. 2008; Dolatabadian et al. 2009). In a study, Pignocchi and Foyer (2003) showed that the foliar spray of AsA in plants under not stress and stress conditions can change the oxidative defense system of plants, improve plant growth and production, contribute to cell expansion, ion transport and other effective processes. The impact of AsA as foliar application on growth improvement has been studied in different organs of plants under environmental stress (Khan et al. 2011). Also, similar results were obtained from the investigation of the effect of ascorbic acid on sweet peppers (Shawky et al. 2003). Various studies have exhibited that plant tolerance to drought stress is related with an increase in antioxidant properties in plants (Alam et al. 2014; Hoque et al. 2008). The foliar application of AsA in plants showed an increase in the amount of endogenous AsA in the plant. In general, it can be said that maintaining the antioxidant defense system in plants is essentially necessary to counteract oxidative damage by abiotic stresses (Qian et al. 2014). In a study on canola, AsA reportedly increased plant tolerance to abiotic stress (Bybordi 2012).
Peppers are a group of popular vegetables which have distinct features like color, flavor and nutritional properties. Different types of peppers contain a broad range of phytochemicals. They are a rich source of vitamin C and carotenoids which are important nutritional antioxidants in food (Byers and Perry 1992). These phytochemicals include phenols and flavonoids which are important antioxidants in plants (Byers and Perry 1992). The amount of AsA in pepper is reportedly higher than the amounts in many other fruits and vegetables (Korkutata and Kavaz 2015). Further to this endogenous feature of sweet peppers, the current study was carried out to determine the influence of AsA application on the physio-biochemical parameters of sweet pepper plants growing under drought stress.
Materials and methods
Plant material and growth conditions
This research was carried out as a factorial experiment in a completely randomized design (RCD) with two factors. The main factors were different levels of drought and AsA concentrations. Each treatment had three replications. The research was situated in a greenhouse at the Faculty of Agriculture, University of Ilam. It lasted 4 months, from seed cultivation to sampling. Seeds of sweet pepper were provided by the Faculty of Agriculture. The peppers seeds were sterilized by 1% sodium hypochlorite (NaOCl) for 10 min before being washed with tap water for 1 min. Then, the seeds were sown in plastic pots that measured 20 cm in height and 23 cm in diameter. The pots were filled with fine sand, sand, leaf mold and garden soil with a ratio of 1:1:1. After filling, each pot weighed 7 kg. The soil samples were analyzed for different soil properties (Table 1).
The average temperature and relative humidity during plant growth were 18/25 °C (day/night) and 60–70%, respectively. A small amount of Tween-20 (polyoxyethylenesorbitan monolaurate) was added as a surfactant to a solution of AsA to increase adhesion between leaves. The AsA treatment started at the fourth leaf stage. AsA was used at 0, 0.5 and 1 mM. The AsA was sprayed onto the leaves so that both sides of each leaf became completely wet. The foliar spray was applied twice. The first instance of application was at 3 days before the drought stress. The second spray was applied 2 weeks after the drought stress began. In the early stages of plant growth, irrigation was complete. Accordingly, 72 h after the foliar spray until sampling (it lasted approximately 4 months), all plants were exposed to three levels of drought stress: stress-free conditions (full irrigation, i.e., the control group), moderate stress (60% of field capacity) and severe stress (30% of field capacity). We have not used any fertigation in this experiment. These conditions were maintained until the end of the experiment. All pots were weighed on a daily basis.
In this experiment, 27 sweet pepper plants were harvested at 80% maturity (in the green stage). Leaf samples consisted of three replicates. Each replicate was obtained from three pots, and therefore, a total of 9 plants existed in each treatment group. The samples were quickly placed in liquid nitrogen after harvest. They were stored in the freezer (− 80 °C) before measuring the biochemical properties, the physiological features and the activity of antioxidant enzymes.
Fruit physical properties
Pepper fruits were harvested and their morphological characteristics (diameter and length of fruit) were recorded. The diameter and length of fruits were measured at their maximum width and length respectively. All measurements were performed by means of a Caliber (Setiamihardja and Knavei 1990).
Growth parameters
To measure leaf area (LA), fully mature leaves that had developed from the main stem of the apical meristem were selected and the total leaf area was determined by an area meter (AM 300 Bio Scientific Ltd., UK) (Phimchan et al. 2012). Leaf chlorophyll index (CHLI) was measured from the last fully developed leaf of sweet pepper by SPAD502 (Wang et al. 2017). The fresh weight of the shoots of pepper seedlings was evaluated by a digital balance. The shoots dry weight was recorded after being dried at 60 °C (Ahmed et al. 2014). The roots of the sweet pepper plants were carefully separated from the pots and, after being washed with water several times, the fresh weight of roots was measured with a balance. Then, they were placed at 60 °C to calculate the dry weight of roots (Ahmed et al. 2014). A method by Ali and Ashraf (2011) was used for determining the relative water content (RWC) of leaves. In this method, fully developed leaves were selected and their fresh weight was recorded. Then, the leaves were placed in test tubes containing distilled water at room temperature for 3 h. Their turgid weights were recorded. The leaves dry weight was recorded after drying the leaves at 70 °C for 48 h. The leaf RWC was obtained using the equation below:
Determining the relative electrical conductivity
The electrical conductivity of pepper leaves was evaluated according to Barranco et al. (2005). The fresh leaves were cut into 0.1-g disks. Contaminants and residues on the samples were removed by distilled water. Then, the leaf samples were added to the test tubes with 10-ml distilled water. The tubes were incubated for 30 min at a temperature of 30 °C and then the initial electric conductivity of the solution (EC1) was measured. The tubes were placed in a bath of water at 100 °C for 15 min and the secondary electric conductivity (EC2) was calculated. Ultimately, the total electrical conductivity was determined by the following equation:
Determination of non-enzymatic antioxidants
Determination of AsA content
AsA content of pepper leaves was estimated by a modified procedure following a method by Luwe et al. (1993). Initially, pepper leaf samples (0.5 g) were poured in liquid nitrogen by mortar and pestle. They were homogenized into ice-cold trichloroacetic acid (TCA, 1% w/v). Then, the solution was centrifuged at 12,000 rpm for 20 min at a temperature of 4 °C. This was followed by adding 50 µl potassium phosphate buffer mixture (0.95 ml, 100 mm, pH 7.0) along with ascorbate oxidase (1 μl of 1 μl−1 unit) to the supernatant. Eventually, the absorbance was read at 265 nm.
Total phenolic content measurement
In sweet pepper plants, total phenol content was measured according to a method by Kahkonen et al. (1999). The fresh pepper leaves (0.4) were homogenized in 4 ml of methanol and 0.1 Mol l−1 HCl mixtures. Then the homogenized material was centrifuged at 15,000 g for 20 min at 4 °C. This was followed by adding distilled water (1:10) and by mixing 2 ml diluted solution with 400 μl supernatant. Subsequently, 1.6 ml of sodium carbonate (7.5%) was added to the above compounds. The mixture was saved kept in the dark for 30 min and was centrifuged at 5000×g for 5 min. Finally, the absorbance was read at 765 nm. The total phenolic content was determined by Gallic acid mg ml−1 fresh material.
Antioxidant capacity
Antioxidant activity of pepper leaves was determined by the method described by Abe et al. (1998). The fresh leaves (0.2 g) were mashed by a pestle in a mortar containing 2 ml of ethanol at 4 °C. Half of the solution was mixed with a solution containing 0.25 ml of 0.5 mM DPPH in ethanol and 0.5 ml of 100 mm acetate buffer (pH 5.5). The absorbance was read at 517 nm after 30 min. The antioxidant activity was determined using the following equation:
Determination of antioxidant enzyme activities
Preparation of extracts
The leaf tissue was added to mortar and pestle containing 50-ml sodium phosphate buffer (pH 7.0) and then crushed. After being centrifuged at 16,000g at 10 °C, the above supernatant was used for measuring the activity of APX and PPO enzymes (Zhao et al. 2007).
Assaying polyphenol oxidase activity
The PPO activity was evaluated according to a method by Jiang et al. (2002). For this purpose, 100 μl of enzyme solution was added to a mixture containing [900 μl of catechol solution, which comprised 40 mM in a sodium phosphate buffer of 0.01 M (pH 6.8)]. Then, the absorbance was read at 400 nm in 25 °C for 3 min.
Determination of ascorbate peroxidase activity
APX activity was evaluated by the Asada (1992) method. The contents of the reaction mixture contained 0.1-ml ascorbate (0.5 mM), 0.1-ml enzyme extract, 0.2-ml H2O2 1% and 2.5 ml of 50-mM potassium phosphate buffer. This solution was supplemented with 0.1-mM EDTA (pH 7.0). Finally, the absorbance was read at 290 nm at 25 °C.
Statistical analysis
The data obtained in this study were evaluated by analysis of variance (ANOVA) using SPSS statistical 10 software and Duncan’s multiple range tests. The analyses were carried out to determine significant variation between the means at a significance level of P < 0.05. Pearson’s correlation between different concentrations of AsA and various levels of drought stress were calculated by SPSS software.
Results
Correlation coefficient
The correlation coefficient was used for determining the natural and extensive relationships between the studied traits in this research. According to the results, some of the studied traits were significant (Table 2). A positive significant correlation was reported between fresh weight of shoot and dry weight of shoot, fresh weight of root, dry weight of root, RWC, fruit diameter, fruit length, chlorophyll index and leaf area index. Also, the fresh weight of shoots had a significant, negative correlation with total phenol, polyphenol oxidase and ascorbic peroxidase. Meanwhile, the dry weight of shoots had a positive significant correlation, with root fresh and dry weight, fruit length, chlorophyll index and leaf area index. However, these traits correlated negatively with the total phenol content and ascorbate peroxidase. In the present study, the fresh weight of roots showed significant positive correlations with the dry weight of roots, fruit diameter, fruit length, relative water content and chlorophyll index. Meanwhile, a high significant correlation was found between root dry weight and relative water content, fruit diameter, fruit length, chlorophyll index and leaf area index. The root dry weight had a significant negative correlation with total phenol content. In addition, a significant and positive correlation existed between RWC and fruit diameter, fruit length, chlorophyll index and leaf area index. However, RWC had a significant and negative correlation with total phenol content.
Also, a significant positive correlation was apparent between fruit diameter, fruit length and chlorophyll index, but these correlated negatively with total phenol content and ascorbate peroxidase. A positive and significant correlation was observed among fruit length, chlorophyll index, leaf area index, whereas these parameters correlated negatively with the total phenol content. A significant but negative correlation was observed between electrical conductivity, ascorbic acid and polyphenol oxidase. Antioxidant capacity showed a significant positive correlation with total phenol, polyphenol oxidase, ascorbate peroxidase and ascorbic acid. In studies on leafy vegetables (Wangcharoen and Morasuk 2007) and chili (Sahitya et al. 2018), it was observed that there was a significant correlation between the DPPH radical scavenging activity and the amount of total phenols. In fact, the amount of total phenols correlated positively and significantly with the amounts of polyphenol oxidase, ascorbate peroxidase and AsA. Ultimately, a correlation analysis showed that polyphenol oxidase has a positive and significant correlation with ascorbate peroxidase and AsA. The correlation analysis showed that ascorbate peroxidase has a positive correlation with AsA. In a study on apples, Pearson’s correlation analysis revealed that antioxidant activity can be seen as a function of the total phenolic measurement because the relationship between total phenolic content and activity of antioxidant was reportedly significant (Candrawinata et al. 2014).
Growth parameters
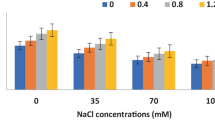
The analyses of data obtained from the factors of this section are shown in Tables 3 and 4. Statistical results showed that severe levels of drought stress led to the decrease in fruit diameter and length, leaf area index, chlorophyll index, and the fresh and dry weight of roots and shoots. As shown in Tables 3 and 4, the application of AsA markedly enhanced the parameters mentioned above at all levels of drought stress. Investigating the interaction between ascorbic acid and drought stress showed that ascorbic acid treatment significantly increased the fresh weight of shoots (Fig. 1a) and the dry weight of roots (Fig. 1b).

Effects of foliar application of AsA and drought stress on some factors of sweet pepper plants. FWSH fresh weight shoot, DWR dry weight root, AC antioxidant capacity, AsA ascorbic acid. Variations between different factors were determined by univariate analyses followed by post hoc analysis. Means with the similar letters are not significantly different using Duncan’s test
Relative water contents (RWC)
Drought stress reduced relative water contents in sweet pepper plants (Table 4). The maximum decrease in this parameter was observed at the most severe level of drought stress; whereas, the application of AsA enhanced RWC at all levels of drought.
Electrical conductivity
Parameters such as electrical conductivity were analyzed so as to determine the severity of oxidative stress in sweet pepper plants under different levels of drought (Table 4). Electrical conductivity was enhanced under drought stress. The highest value of this parameter was observed in response to the most severe drought stress as compared with the control. The foliar application of AsA markedly reduced the electrical conductivity in stressed and non-stressed sweet pepper plants (Table 4). A study of the interaction between drought stress and foliar application level of ascorbic acid showed that as drought stress becomes more severe, the electric conductivity increases. On the other hand, using ascorbic acid reduces the electric conductivity (Fig. 2g).

Effects of foliar application of AsA and drought stress on some factors sweet pepper plants. PPO polyphenol oxidase, APX ascorbate peroxidase, EC electrical conductivity. Variations between different factors were determined by univariate analyses followed by post hoc analysis. Means with the similar letters are not significantly different using Duncan’s test
Ascorbic acid
Drought stress significantly increased AsA content in sweet pepper plants when compared with control plants (Table 5). Accordingly, the amount of AsA increased by higher levels of drought stress. The exogenous application of AsA increased the AsA content inside the stressed and non-stressed sweet pepper plants (Table 5). The interaction between drought stress and ascorbic acid showed that ascorbic acid content in sweet pepper plants increases in response to higher levels of drought stress and ascorbic acid (Fig. 1d).
Total phenolic compounds
Total phenolic compounds increased considerably in sweet pepper plants under drought stress conditions (Table 5). The analysis showed that using different levels of AsA as foliar application increased total phenolic contents in sweet pepper plants under both stress and unstressed conditions (Table 5).
Antioxidant capacity
The endogenous content of antioxidant activity markedly increased in sweet pepper plants parallel to increasing levels of drought compared with control conditions (Table 5). It markedly enhanced the content of antioxidant activity in sweet pepper plants when foliar AsA was applied on pepper seedlings (Table 5). The interaction between drought stress levels and concentrations of ascorbic acid showed that exacerbating the drought stress and increasing the amount of ascorbic acid led to higher levels of antioxidant capacity (Fig. 1c).
Antioxidant enzyme activities
To identify the antioxidant response to drought stress in sweet pepper plants, changes of antioxidant enzymes action were assayed. The activities of antioxidant enzymes measured in this study include those of PPO and APX. The activities of these enzymes in sweet pepper plants were lower under normal conditions, compared to the considerably higher levels of enzymes activities under stress conditions (Table 5). The application of AsA significantly increased the PPO activities and APX in all levels of drought (Table 5). Investigating the interaction between drought stress and ascorbic acid showed that with increasing the drought stress and foliar utilization of ascorbic acid, the content of polyphenol oxidase and ascorbic peroxidase increased in sweet pepper plants (Fig. 2e, f).
Discussion
It was obvious that plant growth limitation under drought stress conditions due to a number of alterations in the physio-biochemical processes (Akram et al. 2009; Kosar et al. 2015). In numerous economically important agricultural products, a substantial decrease in growth and development can be caused by drought stress (Tesfamariam et al. 2010). In most plants, ROS is usually caused by cellular activities, such as respiration and photosynthesis, especially under conditions of various environmental stresses (Tripathy and Oelmuller 2012). The free radicals and reactive oxygen species prevent cell division and proliferation, thereby causing cell death (Dolatabadian et al. 2009).
Dolatabadian et al. (2010) stated that under stress conditions in corn plants, AsA could have a positive effect on protecting cells against active species of oxygen and free radicals in the metabolism of plants. Using AsA on plants as an antioxidant has reportedly reduced the adverse effects of drought stress. Also, a previous work demonstrated that AsA is one of the most important growth regulators that have potential use in the development of resistance to various stresses (Shafiq et al. 2014). In the present study, the protective role of ascorbic acid as an antioxidant system in sweet pepper was investigated under drought stress conditions.
Some reviews have shown the adverse impacts of drought stress on some parameters include fresh and dry weight of plants like sunflower (Hossain et al. 2010) and canola (Tesfamariam et al. 2010), which can be explained by changes that occur at the biochemical and physiological levels of the plant because of drought (Ashraf et al. 2011) (Shafiq et al. 2014). Water in any plant is one of the most important compounds that can be used as a product, reactant, solvent and carrier. It plays a major role in the transfer of nutrients in plants (Furlan et al. 2013). Also, weight reduction in sweet pepper plants can be due to the reduction of the turgor pressure (Ahmed et al. 2014) which is in line with the results of our experiment.
As our study showed, drought stress has a remarkable effect on the growth parameters of pepper plants and fruits. By increasing the drought stress, there were reductions in growth parameters such as fresh weight of shoots, root dry weight (Table 3), fruit length and diameter (Table 3) leaf area (Table 4) and chlorophyll index (Table 4). In the current study, the effects of AsA on pepper have been effective in both the control group and in drought stress conditions. Therefore, according to the obtained data, the exogenous application of ascorbic acid increased all growth factors (Tables 3 and 4). The results of the present study are consistent with the results obtained by (Khan et al. 2011) which showed that the use of ascorbic acid reduced the adverse effects of drought stress on growth parameters of wheat.
Leaf area index (LAI) is the principal mean biogeochemical cycles for light interception and is an important photosynthesis parameter for the plant. Furthermore, it is considered as the best measure for photosynthesis capacity of plants regarding the area occupied by plant biomass (Breda 2003). Reduction of leaf area index in drought stress conditions is attributed to reduced cell growth and cell division rates in the plant. In such a condition, the absorption of sunlight and the amount of photosynthesis of the plant are reduced. Ultimately, the dry matter of the plant decreases (Ahmed et al., 2014). The results of the present experiment were consistent with previous findings (Kasim et al. 2017) on Vigna unguiculata where it was stated that plant growth parameters such as leaf area in each plant decreased due to soil moisture reduction. In another study by Hamama and Murniati (2010) on maize plants under drought stress and ascorbic acid treatments, it was shown that the leaf area of the plant increased significantly parallel to the increase in ascorbic acid.
Drought stress reduced the content of chlorophyll and photosynthesis in wheat (Anjum et al. 2011), which is consistent with the current results (Table 4). In this study, the external application of ascorbic acid improved growth parameters such as chlorophyll index and fruit length and diameter in drought conditions (Tables 3 and 4). The results of Singh et al. (2001) showed that using ascorbic acid on cassia leads to an increment in the content of chlorophyll, which is consistent with our results on pepper.
Relative water content (RWC) is a parameter by which the metabolic activity in tissues of plants can be estimated (Soltys-Kalina et al. 2016). In this study, RWC was remarkably reduced by drought stress conditions (Table 4). However, the use of AsA led to an increment in RWC (Table 4). The results of the analysis of AsA effect in cucumber seedlings under drought stress showed that the positive effects of AsA in the studied traits can be attributed to improving the AsA efficiency in water absorption, which can ultimately improve the turgor pressure and maintain a water balance in the tissues of cucumber (Tuna et al. 2010).
Electrolyte leakage is one of the parameters that is used for measuring the damaged membrane index. The amount of electrolyte leakage increases during stress which can be explained by the peroxidation of saturated fatty acids in the phospholipid membrane. So, electrolyte leakage can be calculated as a norm for estimating the damage of the bio-membrane (Lukatkin and Anjum 2014). Data presented in Table 4 shows that electrolyte leakage increases in response to higher levels of drought stress. Hemmati et al. (2018) reported that damage to cell membranes in pot marigold plants can be due to drought stress condition. The results of the current experiment showed that AsA reduced the negative effects of ROS and increased plant tolerance to drought stress. When canola plants are exposed to water deficit, the molecules with antioxidant properties such as AsA can increase the antioxidant capability of the plant, and thus ascorbic acid reduces the generation of active oxygen species and ameliorates their effect on the cell membrane, which is alignment with the results of a study by Shafiq et al. (2014) and is also consistent with our results.
In this study, the amounts of two antioxidants of AsA and polyphenol in sweet pepper plants increased significantly parallel to the increase in drought stress (Table 5). One of the highlights non-enzymatic antioxidants in plants are phenols that are a part of the antioxidant system of plants (Sahitya et al. 2018). Phenolic compounds protect oxidative damage to cells and increment the stability of cell membranes (Heim et al. 2002). The AsA and polyphenol content increased in sweet pepper plants with foliar application of AsA when compared with control plants (Table 5). AsA improved these biomolecules in drought stress conditions. In a research by Hasanuzzaman and Fujita (2011) on canola plants, the prolific synthesis of ascorbic acid resulted in an increase in the total amount of phenols in drought tolerant plants, which is consistent with our results in pepper. Increasing the AsA in the plant may occur due to the breakdown of proteins, as well as the response to osmotic changes in cellular content. AsA accumulation is very important for osmotic regulation in the cells and osmotic modification is one of the highlights mechanisms in plants that can reduce the harmful effects of water deficit (Jaleel et al. 2007). Results obtained by Ghorbanli et al. (2013) in tomato plants showed that AsA treatment has a considerable effect on the amount of endogenous AsA at times of drought stress. These results are alignment with the results of this study.
To assay the antioxidant activity of sweet pepper, the DPPH test is widely used (Batra et al. 2017). In the current study, we saw that increasing the AsA concentration and the levels of drought led to an increment in the antioxidant activity (Table 5). In a study on the effects of AsA on tea plants (Majchrzak et al. 2004) and in another study on pepper (Sahitya et al. 2018), it was observed that drought stress and ascorbic acid together caused an increase in antioxidant activity in plants. In addition to phenolic compounds, flavonoids and AsA play a role in total antioxidant activity in plants.
In addition to non-enzymatic compounds, enzymatic compounds play a role in eliminating ROS. The removal of ROS in plants is important in their tolerance to oxidative stress, and it is also possible that an enhanced antioxidant activity can increase the resistance to drought stress (Jaleel et al. 2007; Mittler 2002). Accordingly, in this investigation, the activity of the PPO and APX enzymes were measured, along with the activity of both enzymes which increased under both normal and drought situation. The combination of drought stress and AsA can cause changes in sweet pepper plants (Table 5), which is consistent with a report published by Tsaniklidis et al. (2014) about Chrysotile Tomato. The increase in PPO activity in stressed plants could decrease the harmful effects of drought stress. PPO activity reduces the amount of H2O2 and maintains membrane integrity. In this study, the activity of APX enzyme was seen as similar to the findings observed by Wang et al. (2014) on Oryza sativa L. under salt stress. Furthermore, Khanna-Chopra and Selote (2007) reported similar results in studying wheat under drought stress condition. In a work by Wang et al. (2014) showed that APX is an important enzyme in the elimination of H2O2. Therefore, adding ascorbic acid as an electron donor resulted in an increase in APX activity, which eventually results in the conversion of H2O2 into water.
Conclusion
The results of the current investigation on pepper showed that ascorbic acid content in plants enhances in reply to higher levels of drought. The AsA causes changes in the level of antioxidant enzymes activity such as PPO and APX. We concluded that AsA directly reduced the harmful effects of drought stress and, by exacerbating the activity of antioxidant enzymes, ultimately increased drought tolerance in sweet pepper plants.
Author contribution statement
Miss Khazaei carried out the experiment and wrote the manuscript with support from Dr. Estaji. Dr. verified the analytical methods. Miss Khazaei worked out almost all of the technical details, and performed the numerical calculations.
References
Abe N, Murata T, Hirota A (1998) Novel DPPH radical scavengers, bisorbicillinol and demethyltrichodimerol, from a fungus. Biosci Biotechnol Biochem 62(4):661–666. https://doi.org/10.1271/bbb.62.661
Ahmed AF, Yu H, Yang X, Jiang W (2014) Deficit irrigation affects growth, yield, vitamin C content, and irrigation water use efficiency of hot pepper grown in soilless culture. Hortic Sci 49(6):722–728. https://doi.org/10.21273/HORTSCI.49.6.722
Akram MS, Ashraf M, Akram NA (2009) Effectiveness of potassium sulfate in mitigation salt-induced adverse effects on different physiobiochemical attributes in sunflower (Helianthus annuus L.). Flora 204:471–483. https://doi.org/10.1016/j.flora.2008.05.008
Alam MM, Nahar K, Hasanuzzaman M, Fujita M (2014) Alleviation of osmotic stress in Brassica napus, B. campestris, and B. juncea by ascorbic acid application. Biol Plant 58(4):697–708. https://doi.org/10.1007/s10535-014-0447-0
Ali Q, Ashraf M (2011) Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of trehalose: growth, photosynthesis, water relations and oxidative defence mechanism. J Agron Crop Sci 197(4):258–271. https://doi.org/10.1111/j.1439-037X.2010.00463.x
Anjum S, Xie XY, Wang LC, Saleem MF, Man C, Wang L (2011) Morphological, physiological and biochemical responses of plants to drought stress. Afr J Agric Res 6:2026–2032. https://doi.org/10.5897/AJAR10.027
Asada RD (1992) Ascorbate peroxidase—a hydrogen peroxide scavenging enzyme in plants. Plant Physiol 85:235–241. https://doi.org/10.1111/j.1399-3054.1992.tb04728.x
Ashraf M (2009) Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol Adv 27(1):84–93. https://doi.org/10.1016/j.biotechadv.2008.09.003
Ashraf MH, Harris PJ (2013) Photosynthesis under stressful environments: an overview. Photosynthetica 51(2):163–190. https://doi.org/10.1007/s11099-013-0021-6
Ashraf M, Akram NA, Al-Qurainy F, Foolad MR (2011) Drought tolerance: roles of organic osmolytes, growth regulators, and mineral nutrients. Adv Agron 111:249–296. https://doi.org/10.1016/B978-0-12-387689-8.00002-3
Athar H, Khan A, Ashraf M (2008) Exogenously applied ascorbic acid alleviates salt-induced oxidative stress in wheat. Environ Exp Bot 63(1–3):224–231. https://doi.org/10.1016/j.envexpbot.2007.10.018
Barranco D, Ruiz N, Gomes M (2005) Frost tolerance of eight olive cultivars. HortScience 40:558–560. https://doi.org/10.21273/HORTSCI.40.3.558
Batra G, Gortzi O, Lalas SI, Galidi A, Alibade A, Nanos GD (2017) Enhanced antioxidant activity of Capsicum annuum L. and Moringa oleifera L. extracts after encapsulation in microemulsions. Chem Eng 1(2):15. https://doi.org/10.3390/chemengineering1020015
Bayat H, Moghadam AN (2019) Drought effects on growth, water status, proline content and antioxidant system in three Salvia nemorosa L. cultivars. Acta Physiol Plant 41(9):149
Breda NJ (2003) Ground-based measurements of leaf area index: a review of methods, instruments and current controversies. J Exp Bot 54(392):2403–2417. https://doi.org/10.1093/jxb/erg263
Bybordi A (2012) Effect of ascorbic acid and silicium on photosynthesis, antioxidant enzyme activity, and fatty acid contents in canola exposure to salt stress. J Integr Agric 11(10):1610–1620. https://doi.org/10.1016/S2095-3119(12)60164-6
Byers T, Perry G (1992) Dietary carotenes, vitamin C, and vitamin E as protective antioxidants in human cancers. Annu Rev Nutr 12(1):139–159. https://doi.org/10.1146/annurev.nu.12.070192.001035
Candrawinata VI, Golding JB, Roach PD, Stathopoulos CE (2014) Total phenolic content and antioxidant activity of apple pomace aqueous extract: effect of time, temperature and water to pomace rati. Int Food Res J 21(6):2337–2344
Dolatabadian A, Modarres Sanavy SA, Sharifi M (2009) Alleviation of water deficit stress effects by foliar application of ascorbic acid on Zea mays L. J Agron Crop Sci 195(5):347–355. https://doi.org/10.1111/j.1439-037X.2009.00382.x
Dolatabadian A, Sanavy SA, Asilan KS (2010) Effect of ascorbic acid foliar application on yield, yield component and several morphological traits of grain corn under water deficit stress conditions. Not Sci Biol 2(3):45–50. https://doi.org/10.15835/nsb.2.3.4717
Farooq M, Wahid A, Kobayashi N, Fujita D, Basra SM (2009) Plant drought stress: effects, mechanisms and management. J Sustain Agric. https://doi.org/10.1007/978-90-481-2666-8_12
Foyer CH, Alscher RG, Hess JL (1993) Antioxidants in higher plants. CRC Press, Inc., Florida, pp 31–58
Furlan A, Llanes A, Luna V, Castro S (2013) Abscisic acid mediates hydrogen peroxide production in peanut induced by water stress. Biol Plant 57(3):555–558. https://doi.org/10.1007/s10535-012-0296-7
Ghorbanli M, Gafarabad M, Amirkian T, Allahverdi Mamaghani B (2013) Investigation of proline, total protein, chlorophyll, ascorbate and dehydroascorbate changes under drought stress in Akria and Mobil tomato cultivars. J Plant Physiol 3(2):651–658
Hamama H, Murniati E (2010) The effect of ascorbic acid treatment on viability and vigor maize (Zea mays L.) seedling under drought stress. Hayati J Biosci 17(3):105–109. https://doi.org/10.4308/hjb.17.3.105
Hasanuzzaman M, Fujita M (2011) Selenium pretreatment upregulates the antioxidant defense methylglyoxal detoxification system and confers enhanced tolerance to drought stress in rapeseed seedlings. Biol Trace Elem Res 143:1758–1776. https://doi.org/10.1007/s12011-011-8998-9
Heim KE, Tagliaferro AR, Bobilya DJ (2002) Flavonoid antioxidants: chemistry, metabolism and structure-activity relationships. J Nutr Biochem 13(10):572–584. https://doi.org/10.1016/S0955-2863(02)00208-5
Hemmati K, Ebadi A, Khomari S, Sedghi M (2018) Influence of ascorbic acid and 24-epibrassinolide on physiological characteristics of pot marigold under water-stress condition. J Plant Interact 13(1):364–372. https://doi.org/10.1080/17429145.2018.1483033
Hoque MA, Banu MN, Nakamura Y, Shimoishi Y, Murata Y (2008) Proline and glycinebetaine enhance antioxidant defense and methylglyoxal detoxification systems and reduce NaCl-induced damage in cultured tobacco cells. J Plant Physiol 165:813–824. https://doi.org/10.1016/j.jplph.2007.07.013
Hossain MI, Khatunm A, Talukder MS, Dewan MM, Uddin MS (2010) Effect of drought on physiology and yield contributing characters of sunflower. Bangladesh J Agric Res 35(1):113–124. https://doi.org/10.3329/bjar.v35i1.5872
Hu H, Xiong L (2014) Genetic engineering and breeding of drought-resistant crops. Annu Rev Plant Biol 65:715–741. https://doi.org/10.1146/annurev-arplant-050213-040000
Jaleel CA, Manivannan P, Kishorekumar A, Sankar B, Gopi R, Somasundaram R, Panneerselvam R (2007) Alterations in osmoregulation, antioxidant enzymes and indole alkaloid levels in Catharanthus roseus exposed to water deficit. Colloid Surf B 59(2):150–157. https://doi.org/10.1016/j.colsurfb.2007.05.001
Jiang Y, Zhang Z, Joyce DC, Ketsa S (2002) Postharvest biology and handling of longan fruit (Dimocarpus longan Lour.). Postharvest Biol Technol 26(3):241–252. https://doi.org/10.1016/S0925-5214(02)00047-9
Kahkonen MP, Hopia AI, Vuorela HJ, Rauha JP, Pihlaja K, Kujala TS, Heinonen M (1999) Antioxidant activity of plant extracts containing phenolic compounds. J Agric Food Chem 47(10):3954–3962. https://doi.org/10.1021/jf990146l
Kasim WA, Nessem AA, Gaber A (2017) Alleviation of drought stress in Vicia faba by seed priming with ascorbic acid or extracts of garlic and carrot. In: The 7th international conference plant and microbial biotechnology and their role in the development of the society, pp 45–59. https://doi.org/10.21608/ejbo.2017.831.1057
Khan T, Mazid M, Mohammad F (2011) A review of ascorbic acid potentialities against oxidative stress induced in plants. J Agrobiol 28(2):97–111. https://doi.org/10.2478/v10146-011-0011-x
Khanna-Chopra R, Selote DS (2007) Acclimation to drought stress generates oxidative stress tolerance in drought-resistant than susceptible wheat cultivar under field conditions. Environ Exp Bot 60(2):276–283. https://doi.org/10.1016/j.envexpbot.2006.11.004
Kolarovic L, Valentovic P, Luxova M, Gasparíkova O (2009) Changes in antioxidants and cell damage in heterotrophic maize seedlings differing in drought sensitivity after exposure to short-term osmotic stress. Plant Growth Regul 59(1):21–26
Korkutata NF, Kavaz A (2015) A comparative study of ascorbic acid and capsaicinoid contents in red hot peppers (Capsicum annum L.) grown in southeastern Anatolia region. Int J Food Prop 18(4):725–734. https://doi.org/10.1080/10942912.2013.850507
Kosar F, Akram NA, Ashraf M (2015) Exogenously-applied 5-aminolevulinic acid modulates some key physiological characteristics and antioxidative defense system in spring wheat (Triticum aestivum L.) seedlings under water stress. S Afr J Bot 96:71–77. https://doi.org/10.1016/j.sajb.2014.10.015
Lukatkin AS, Anjum NA (2014) Control of cucumber (Cucumis sativus L.) tolerance to chilling stress—evaluating the role of ascorbic acid and glutathione. Front Environ Sci 2(62):1–6. https://doi.org/10.3389/fenvs.2014.00062
Luwe MW, Takahama U, Heber U (1993) Role of ascorbate in detoxifying ozone in the apoplast of spinach (Spinacia oleracea L.) leaves. Plant Physiol 101(3):969–976
Mahmoud AAS (2014) Suppression of weeds growing with Vicia faba by glucosinolates of some species of Brassicaceae as compared with the herbicide fusilade. Ph.D. Thesis, Faculty of Science, Tanta University, Egypt
Majchrzak D, Mitter S, Elmadfa I (2004) The effect of ascorbic acid on total antioxidant activity of black and green teas. Food Chem 88(3):447–451. https://doi.org/10.1016/j.foodchem.2004.01.058
Mittler R (2002) Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 7(9):405–410. https://doi.org/10.1016/S1360-1385(02)02312-9
Noctor G, Foyer CH (1998) Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Biol 49(1):249–279. https://doi.org/10.1146/annurev.arplant.49.1.249
Phimchan P, Techawongstien S, Chanthai S, Bosland PW (2012) Impact of drought stress on the accumulation of capsaicinoids in Capsicum cultivars with different initial capsaicinoid levels. Hortic Sci 47(9):1204–1209. https://doi.org/10.21273/HORTSCI.47.9.1204
Pignocchi C, Foyer CH (2003) Apoplastic ascorbate metabolism and its role in the regulation of cell signalling. Curr Opin Plant Biol 6(4):379–389. https://doi.org/10.1016/S1369-5266(03)00069-4
Qian HF, Peng XF, Han X, Ren J, Zhan KY, Zhu M (2014) The stress factor, exogenous ascorbic acid, affects plant growth and the antioxidant system in Arabidopsis thaliana. Russ J Plant Physiol 61(4):467–475. https://doi.org/10.1134/S1021443714040141
Sahitya UL, Krishna MS, Deepthi R, Prasad GS, Kasim D (2018) Seed Antioxidants Interplay with drought stress tolerance indices in chilli (Capsicum annuum L) seedlings. Biomed Res Int 208:1–14. https://doi.org/10.1155/2018/1605096
Seminario A, Song L, Zulet A, Nguyen HT, González EM, Larrainzar E (2017) Drought stress causes a reduction in the biosynthesis of ascorbic acid in soybean plants. Front Plant Sci 8:1042. https://doi.org/10.3389/fpls.2017.01042
Setiamihardja R, Knavei DE (1990) Association of pedicel length and diameter with fruit length and diameter and ease of fruit detachment in pepper. J Am Soc Hortic Sci 115(4):677–681. https://doi.org/10.21273/JASHS.115.4.677
Shafiq S, Akram NA, Ashraf M, Arshad A (2014) Synergistic effects of drought and ascorbic acid on growth, mineral nutrients and oxidative defense system in canola (Brassica napus L.) plants. Acta Physiol Plant 36(6):1539–1553. https://doi.org/10.1007/s11738-014-1530-z
Shawky NB, Higazy MA, Ahmed AH, Elbahr MK (2003) Physiological studies on the effect of salinity, ascorbic acid and putrescine on sweet pepper plant. Doctoral dissertation, Ph.D. Thesis, Department of Agricultural Botany, Faculty of Agriculture, University of Cairo
Singh DV, Srivastava GC, Abdin MZ (2001) Amelioration of negative effect of water stress in Cassia angustifolia by benzyladenine and/or ascorbic acid. Biol Plant 44(1):141–143. https://doi.org/10.1023/A:1017955328875
Smirnoff N (1995) Antioxidant systems and plant response to the environment. Environment and plant metabolism: flexibility and acclimation. Bios Scientific Publishers, Abingdon, pp 217–243
Soltys-Kalina D, Plich J, Strzelczyk-Zyta D, Sliwka J, Marczewski W (2016) The effect of drought stress on the leaf relative water content and tuber yield of a half-sib family of ‘Katahdin’-derived potato cultivars. Breed Sci 66(2):328–331. https://doi.org/10.1270/jsbbs.66.328
Tesfamariam EH, Annandale JG, Steyn JM (2010) Water stress effects on winter canola growth and yield. Agron J 102(2):658–666. https://doi.org/10.2134/agronj2008.0043
Tripathy BC, Oelmuller R (2012) Reactive oxygen species generation and signaling in plants. Plant Signal Behav 7(12):1621–1633. https://doi.org/10.4161/psb.22455
Tsaniklidis G, Delis C, Nikoloudakis N, Katinakis P, Passam HC, Aivalakis G (2014) l-Ascorbic acid metabolism in parthenocarpic and seeded cherry tomatoes. Plant Growth Regul 72(2):141–153. https://doi.org/10.1007/s10725-013-9845-0
Tuna AL, Kaya C, Ashraf M (2010) Potassium sulfate improves water deficit tolerance in melon plants grown under glasshouse conditions. J Plant Nutr 33(9):1276–1286. https://doi.org/10.1080/01904167.2010.484089
Wang R, Liu S, Zhou F, Ding C (2014) Exogenous ascorbic acid and glutathione alleviate oxidative stress induced by salt stress in the chloroplasts of Oryza sativa L. Z Naturforsch C 69(5–6):226–236. https://doi.org/10.5560/znc.2013-0117
Wangcharoen W, Morasuk W (2007) Antioxidant capacity and phenolic content of some Thai culinary plants. Mj Int J Sci Tech 01(02):100–106
Wang X, Wu L, Xie J, Li T, Cai J, Zhou Q, Dai T, Jiang D (2017) Herbicide isoproturon aggravates the damage of low temperature stress and exogenous ascorbic acid alleviates the combined stress in wheat seedlings. Plant Growth Regul 84(2):293–301. https://doi.org/10.1007/s10725-017-0340-x
Wolucka BA, Goossens A, Inze D (2005) Methyl jasmonate stimulates the de novo biosynthesis of vitamin C in plant cell suspensions. J Exp Bot 56(419):2527–2538. https://doi.org/10.1093/jxb/eri246
Xu Y, Xu Q, Huang B (2015) Ascorbic acid mitigation of water stress-inhibition of root growth in association with oxidative defense in tall fescue (Festuca arundinacea Schreb.). Front Plant Sci 6:1–14. https://doi.org/10.3389/fpls.2015.00807
Zhao H, Tan J, Qi C (2007) Photosynthesis of (Rehmannia glutinosa) subjected to drought stress is enhanced by choline chloride through alleviating lipid peroxidation and increasing proline accumulation. Plant Growth Regul 51(3):255–262. https://doi.org/10.1007/s10725-007-9167-1
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R. Aroca.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Khazaei, Z., Estaji, A. Effect of foliar application of ascorbic acid on sweet pepper (Capsicum annuum) plants under drought stress. Acta Physiol Plant 42, 118 (2020). https://doi.org/10.1007/s11738-020-03106-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-020-03106-z