
Abstract
Global warming is a critical issue and has great impact on all living organisms, including algae. Generally, algae play significant roles in aquatic ecosystems and employ diverse strategies to survive under abiotic stress. For example, heat stress affects membrane fluidity, and algae, in response, can modify their membrane fatty acid composition to maintain homoeostasis. Moreover, they protect their proteins and enzymes using molecular chaperones or degrade denatured proteins in processes involving ubiquitin. In addition, algae regulate their carbohydrate concentrations and structures to utilise the energy of endogenous carbon sources efficiently and protect other molecules via accumulation of compatible solutes. Algae regulate the photosynthetic machinery to acclimatise to stress conditions. In fact, algae have a range of acclimation and repair strategies; and in the case where these strategies fail, programmed cell death (PCD) will be activated. Among algae, green algae have been massively studied due to their broad-range applications such as pharmaceutical, biofuel production and wastewater management, and being a suitable model to study plant and photosynthesis. Enhanced knowledge about the genes and proteins involved in the acclimation of green algae would enlighten our understanding of their acclimation pathways, and enable the genetic improvement of stress-tolerant strains. Thus, the mechanisms and pathways associated with green algal acclimation and repair strategies with an emphasis on temperature-related stress are highlighted in this review.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Massive consumption of fossil fuel and the release of greenhouse gases into the air contribute to global warming (Judkins et al. 1993; Solomon et al. 2009; McGlade and Ekins 2015). Global warming is a deleterious factor affecting aquatic ecosystems, and sequentially leads to significant shifts in aquatic biogeochemical cycles, dynamics, biodiversity and aquatic food web structure (Harley et al. 2006; Wrona et al. 2006). In aquatic ecosystems, algae play essential roles in carbon sequestration and contribute to food webs as primary producers (Sayre 2010; Tsai et al. 2015). Their roles and distribution are influenced by the shift in environmental variables due to global warming (Beardall and Raven 2004; Bopp et al. 2005; Paul 2008; Olsenz 2011). Acute changes in temperature, irradiance, salinity, or pH will influence their cellular integrity and bio-molecular composition, thereby disrupting cellular homoeostasis (Jauzein and Erdner 2013; Van Creveld et al. 2015). Environmental stress could lead to severe damage and initiate responses that result in either acclimation or programmed cell death, depending on the sensitivity of the algae (Zuppini et al. 2007; Sulmon et al. 2015). Commonly, algae utilise several stress management and repair strategies (Fig. 1), such as adjusting membrane fluidity via changing the fatty acid saturation level (Murata and Los 1997; Los et al. 2013; Maksimov et al. 2017), reducing protein synthesis to avoid an upsurge of mis-folded proteins (Schroda et al. 2015), accumulating compatible solutes to maintain cell osmolality (Klahn and Hagemann 2011), and regulating photosynthesis to balance energy production and consumption (Ras et al. 2013). In this review, green algal acclimation mechanisms and pathways responding to temperature stress are emphasised.

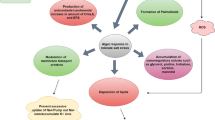
Molecular events involved in DNA damage repair and stress adaptation. Algae have developed a range of adaptation and repair strategies. However, these strategies might fail leading to the activation of programmed cell death
Membrane homeostasis and lipid content
The capability of green algae to survive against various ecological conditions is strongly reflected in the unusual pattern of their cellular lipids and their capacities for changing lipid metabolism (Thompson 1996; Liu et al. 2016). The physical properties of the membrane rely on its fatty acid profile and the degree of saturation which regulates the fluidity of membranes (Singh et al. 2002). The fluidity of membranes is affected by temperature fluctuations. Low temperature reduces membrane fluidity, which is ameliorated via membrane lipid desaturation by means of fatty acid desaturases (FADs) (Sakamoto and Murata 2002). Indeed, at low temperature, algal cells increase the content of membrane unsaturated fatty acids (UFAs) to achieve a looser packing of lipids, thus, decrease the solidification of membrane lipids (Lyon and Mock 2014). Furthermore, it was noted that by increasing the percentage of unsaturated fatty acids in the membrane, the photosynthetic machinery of algae could also be stabilised under chilling (Wada et al. 1994) and salinity stress conditions (Allakhverdiev et al. 2001). Conversely, heat increases membrane fluidity, which is ameliorated by the integration of de novo-synthesised saturated fatty acids (SFAs) into membrane lipids, and the presence of membrane-stabilizing proteins (Los et al. 2013). Fundamentally, two main family groups of enzymes control fatty acid structures; desaturases and elongases (Khozin-Goldberg and Cohen 2011). During stress conditions, cells show an increase in fatty acid synthesis. The mechanisms involved in lipid homoeostasis can be monitored by analysing the expression of the central regulatory genes, such as genes encoding subunits of acetyl-CoA carboxylase (ACCase), ketoacyl-ACP synthase (KAS), desaturase, and elongases. ACCase is known as one of the enzymes that control fatty acid biosynthesis rate and present in almost all algae (Podkowinski and Tworak 2011; Singh et al. 2016). It comprises four subunits, namely accA, accD, accC and accB. In certain stress conditions, such as metal stress (e.g. iron), up-regulation of ACCase subunits was observed along with elevated ACCase activity and fatty acid synthesis in Chlorella sorokiniana (Wan et al. 2014). ACCase activity yields malonyl-ACP, where the malonyl entity participates in a sequence of condensation reactions, resulting in the lengthening of the precursor fatty acid in which KAS is the rate-limiting enzyme (Rismani-Yazdi et al. 2011; Lei et al. 2012). In Dunaliella salina, high levels of KAS were found when cells were exposed to increased salinity levels (Azachi et al. 2002).
It is essential to consider that the SFAs formed might undergo elongation and desaturation steps to make longer chains of unsaturated fatty acids, or polyunsaturated fatty acids (PUFAs) (Pereira et al. 2003). PUFAs are fatty acids containing 18 carbons or beyond with the presence of at least two double bonds (Huang et al. 2004). They affect membrane fluidity owing to their very low melting points. The solidification of the membrane lipids can, therefore, be reduced by increasing the ratio of PUFAs in the membrane (Brett and Müller-Navarra 1997). Amidst the enzymes participating in lipid metabolism, ∆9 fatty acid desaturases (FADs) catalyse the first committed step in desaturation and initiate the conversion of SFAs to mono-unsaturated fatty acids (MUFAs), which are vital for the generation of PUFAs (Xue et al. 2016). These desaturases introduce the first double bond to palmitic acid (C16:0) and stearic acid (C18:0) and convert them to palmitoleic acid (C16:1) and oleic acid (C18:1), individually (Sakuradani 1999; Xue et al. 2016). Up-regulation of several fatty acid desaturases (Δ9ACPCiFAD, Δ12CiFAD, ω3CiFAD2 and Δ6CiFAD) was observed in the Antarctic ice alga Chlamydomonas sp. ICE-L when it was exposed to salinity stress (An et al. 2013a). Under low temperature (0 °C), Δ9CiFAD, ω3CiFAD2 and ω3CiFAD1 were up-regulated in this microalga while Δ6CiFAD amplified with rising temperature. However, the temperature did not cause deregulation of Δ12CiFAD except at a specific temperature (15 °C). These observations suggested that ω3CiFADs might have important roles in cold adaptation while Δ6CiFAD enhanced survival under high temperature (An et al. 2013b). On the other hand, in response to cold and salt stress, Antarctic C. vulgaris up-regulated the Δ12FAD enzyme which catalysed desaturation at the Δ12 position (Lu et al. 2010a, b). Stearoyl-acyl carrier protein desaturase (SAD) is another enzyme involved in oleic acid (18:1) synthesis via the insertion of the first double bond into stearic acid (18:0) (Shanklin and Somerville 1991; Shanklin and Cahoon 1998). Stress conditions such as high light and nitrogen deficiency (N-) drastically up-regulated the transcripts of SAD in C. zofingiensis and resulted in the accumulation of whole fatty acids, comprising oleic acid (Liu et al. 2012).
In eukaryotes, energy can be stored as triacylglycerols (TAGs), which act as an essential carbon source (Merchant et al. 2012), contributing to a vital part of survival in environmentally harsh conditions (Singh et al. 2002; Fan et al. 2014). Most microalgae accumulate TAG under nitrogen- or phosphorus-limiting conditions (Fan et al. 2014), high salinity (Bartley et al. 2013) and temperature stress (Fakhry and El Maghraby 2015; Elsayed et al. 2017). Two pathways to form TAG are emphasised, namely the acyl-CoA-dependent (Kennedy pathway) and the acyl-CoA-independent pathways (Lenka et al. 2016). In algae, the Kennedy is the main pathway where TAG is generated via consecutive acylation of glycerol-3-phosphate (G3P) using acyl-CoA. Briefly, glycerol-3-phosphate acyltransferase (GPAT) begins TAG biosynthesis via catalysing the transfer of FA from the acyl-CoA pool on to G3P forming lysophosphatidic acid (Wendel et al. 2009), followed by the production of phosphotidic acid (PA) catalysed by lysophosphatidic acid acyltransferase (LPAAT). Overexpression of GPAT and LPAAT led to an increase of lipid percentage in Chlamydomonas reinhardtii (Iskandarov et al. 2016; Yamaoka et al. 2016). Meanwhile, diacylglycerol acyltransferase (DGAT) catalyses the last stage of TAG assembly, and is proposed as a key regulator of TAG biosynthesis (Lenka et al. 2016). Indeed, the transformation of the gene encoding DGAT was used as a strategy to obtain higher lipid content in C. reinhardtii (Ahmad et al. 2015). Alternatively, TAG can also be produced by three enzymatic reactions in the acyl-CoA-independent pathway, involving phospholipid diacylglycerol acyltransferase (PDAT), DGAT, and diacylglycerol transacylase (DGTA) (Chen and Smith 2012). PDAT was reported as the main enzyme for TAG biosynthesis in C. reinhardtii (Yoon et al. 2012) and increased levels of PDAT were observed when the microalga was subjected to nutrient limitations, such as iron (Urzica et al. 2013) and nitrogen (Boyle et al. 2012).
Protein homoeostasis
Several stress conditions, particularly heat, disrupt weak interactions in protein structures and affect the stability of protein conformation (Daniel et al. 1996). Heat stress could induce the aggregation and denaturation of proteins, resulting in membrane injury and alterations to metabolic fluxes (Fu et al. 2008). Consequently, accumulation of unfolded proteins under stress conditions triggers the expression of molecular chaperones, namely heat-shock proteins (HSPs), that act to re-establish protein homoeostasis (Fulda et al. 2010). Expression of HSPs induced by high-temperature is commonly observed in various living organisms (Parsell and Lindquist 1993; Gupta et al. 2010). According to their molecular masses, several HSPs have been identified including HSP90 and HSP70 (Waters et al. 1996). HSP90s are known to have a noticeable function in conserving protein homoeostasis by stabilising and maintaining the conformation of unstable proteins close to their native forms. In addition, HSP90s are also involved in various signalling and cellular pathways (Richter and Buchner 2001; Young et al. 2001). Meanwhile, HSP70 chaperones are involved in other aspects of protein processing, specifically protein translocation and protein folding (Bukau and Horwich 1998). Chloroplast stromal HSP70B in C. reinhardtii and Dunaliella were reported to contribute to the molecular protection and repair of photosystem II (PSII) (Schroda et al. 1999; Yokthongwattana et al. 2001). An increase of 5–10 °C above normal temperature triggered the expression of the HSP70B gene in the Antarctic algae (Vayda and Yuan 1994). Chankova et al. (2013) recommended that HSP70B could be a marker for temperature stress in Chlorella from different environments and with different temperature tolerance. They observed that under similar temperature stress, about 30% higher levels of HSP70B were recorded for Antarctic C. vulgaris when compared with thermophilic C. vulgaris and mesophilic C. kesslery. Interestingly, no reduction of HSP70B levels was observed in Antarctic C. vulgaris at low temperature indicating the role of HSP70B for cell survival under Antarctic extreme conditions. Similarly, levels of HSPs increased in C. acidophila under temperature stress and HSPs might also contribute to the survival of acidophilic algae in harsh conditions (Gerloff-Elias et al. 2006). Moreover, the expression of several small HSPs was up-regulated at species-specific threshold temperature indicating the association of temperature sensing systems in algae (Kobayashi et al. 2014).
In addition to HSPs, several other proteins are involved in protecting cells against stress-induced damages. Late-embryogenesis abundant (LEA) proteins reported in green algae (Honjoh et al. 1995, 2000) increased freezing tolerance in Chlorella (Liu et al. 2011). They are thought to stabilise membranes and other proteins during cellular dehydration via a mechanism involving hydrogen-bond stabilisation (Shih et al. 2008). During stress, proteins that are irreversibly denatured or mis-folded are labelled by ubiquitins for degradation (Hershko and Ciechanover 1998). The ubiquitinated proteins can then be acted upon by proteolytic enzymes known as proteasomes. Ubiquitination can be induced by different stresses and it regulates various biological processes in eukaryotes, namely receptor endocytosis, cell-cycle control, intracellular signalling, transcription, gene silencing, DNA repair, and stress responses (Kwapisz et al. 2005; Kaliszewski and Zoladek 2008). Enhanced ubiquitin levels are associated with high levels of protein degradation and turnover (Hawkins 1991; Shahsavarani et al. 2012). Up-regulation of ubiquitin has been observed in C. reinhardtii in response to heat and light stress (Wettern et al. 1990; Von Kampen et al. 1995). It was described that the ubiquitin–proteasome pathway (UPP) in C. reinhardtii could be impaired by severe or prolonged stress exposure due to the accumulation of reactive oxygen species (ROS) (Vallentine et al. 2014). In addition to protein degradation, under unfavourable conditions such as heat stress, protein synthesis might be reduced as a strategy to limit the risk of generating mis-folded proteins (Allakhverdiev and Murata 2004).
Carbohydrate homoeostasis
According to energy allocation models, there is an increase in cell maintenance costs under harsh conditions due to the higher energy demand to maintain homoeostasis (Calow 1991). Hence, carbohydrate metabolism is affected by abiotic stress leading to a decrease of carbon storage and an upsurge in levels of dissolved sugars (Arbona et al. 2013). González-Fernandez and Ballesteros (2012) reviewed the impacts of various environmental parameters on the carbohydrate content of algae. Cultivation parameters such as limiting nutrients or starvation, inorganic carbon supply, sodium chloride concentration, irradiance and temperature can all affect the activity of enzymes associated with carbohydrate accumulation (Vinocur and Altman 2005). In contrast, in a study on tropical microalgae from Australia, no consistent pattern was found in carbohydrate content in response to temperature treatment experiments (Renaud et al. 2002). Among carbohydrates, sucrose is the major outcome of photosynthesis and has crucial parts in plant growth and development, storage reserve, cell signalling and stress acclimation (Jiang et al. 2015). Sucrose formation is heavily temperature dependent and is enhanced by an increase in temperature, which may be due to the involvement of a thermophilic enzyme system in sucrose synthesis (Müller and Wegmann 1978). However, algae respond differently in altering sucrose levels when subjected to temperature changes; for example, in Dunaliella, sucrose was accumulated at elevated temperatures (Müller and Wegmann 1978), while in Chlorella (Guy 1990) and Klebsormidium flaccidum (Nagao and Uemura 2012) sucrose accumulation was instead reported at low temperatures. In C. vulgaris at temperatures above its ambient, the sucrose level greatly increased due to degradation of starch (Nakamura and Miyachi 1982).
The enzyme sucrose phosphate phosphatase (SPP), an essential component of the sucrose synthesis pathway, is involved in hydrolysing sucrose 6-phosphate (S6P) into sucrose and inorganic phosphate (Nagao and Uemura 2012). The SPP family exists ubiquitously in algae (Jiang et al. 2015). In C. reinhardtii, the enhanced expression of the SPP gene in response to cold stress resulted in a consistent rise in sucrose levels (Valledor et al. 2013). Algae also respond to environmental stress, especially nutrient deficiency, by accumulating starch as the primary carbon and energy storage (Geider and La Roche 2002; Zhu et al. 2014b). The biosynthesis of starch is regulated by the rate-limiting enzyme ADP-glucose pyrophosphorylase (AGPase) whose activity is affected by temperature and redox mechanisms (Ball and Morell 2003; Thitisaksakul et al. 2012). For example, in C. zofingiensis starch abundance increased significantly at an early stage of nitrogen starvation, with a significant reduction of starch synthesis rate at a later stage, followed by an increase in starch degradation and lipid production (Zhu et al. 2014a). Similarly, AGPase was highly expressed in C. vulgaris in the first few hours of response to N-starved conditions, while its expression was reduced after 120 h of incubation (Ikaran et al. 2015). Vitova et al. (2015) conducted a review of the approaches to enhance the bioaccumulation of starch and lipids, namely the use of inhibitors to prevent DNA synthesis or cell division as well as by nutrient starvation involving the macro-elements such as nitrogen, phosphorus and sulphur. Starch is known as the most common storage form of carbohydrate and as a compact and osmotically inert form of energy storage (Pfister and Zeeman 2016). In phototrophically grown algae, starch which is accumulated prior to cellular division, is utilised as a carbon and energy reserve to ensure completion of cell division processes is independent of external supplies of carbon and energy either during photosynthesis or in the dark (Vitova et al. 2015). Hence, starch degradation is an essential process in the cell, especially in photosynthetic organisms during the dark (Smith et al. 2005). Starch phosphorylase plays an essential role in starch degradation (Rathore et al. 2009) and its expression is significantly induced in green algae by nitrogen starvation (Li et al. 2012).
Compatible solutes
Under several stress conditions, typically osmotic stress, green algae accumulate organic solutes, such as the amino acid proline and sugar alcohols. These ‘compatible solutes’ do not hinder enzymatic reactions even at high concentration, but protect the membranes and enzymes against the lethal effects of destabilising ions (Yancey et al. 1982; Rathinasabapathi 2000). These compatible solutes comprised quaternary amino acid derivatives (Ahmad and Hellebust 1988; Parvaiz and Satyawati 2008); monosaccharides, disaccharides, and polysaccharides (Peters et al. 2007; Nagao et al. 2008); sugar alcohols (Borowitzka and Brown 1974; Wegmann 1986; Bohnert et al. 1995; Hagemann 2016) and sulphonium compounds (Rhodes and Hanson 1993; Scholz and Liebezeit 2012). Proline accumulation is known to occur under various extreme conditions such as UV radiation and low temperature (Hayat et al. 2012). Amplified levels of proline also protect enzymes against inactivation by heat (Liang et al. 2013). The biosynthesis of proline through the glutamate pathway is catalysed via ∆1-pyrroline-5-carboxylate synthetase (P5CS) and ∆1-pyrroline-5-carboxylate reductase (P5CR), where the latter limits the rate of flux through the pathway (Hu et al. 1992). In various plant studies, the expression pattern of P5CS was examined as an indicator of proline production (Zhang et al. 2008; Amini et al. 2015). In green algae, an increased level of P5CS gene expression was found in C. reinhardtii exposed to copper (Zhang et al. 2008) and mercury (Elbaz et al. 2010). Proline exhibited protection against metal-induced oxidative stress via quenching oxygen singlets and scavenging radicals such as hydroxyl radicals (Siripornadulsil et al. 2002). However, report about the effect of temperature on proline level in green microalgae was not found in the literature.
Another compatible solute, trehalose or a non-reducing disaccharide stabilizes biological structures under abiotic stress, possibly by its capacity to absorb water, thus, protecting biomolecules from dehydration (Garg et al. 2002; Vinocur and Altman 2005; Lunn et al. 2014). It was proposed that trehalose works as a ‘chemical chaperone’ due to its ability to stabilise proteins and other biological structures against damage (Crowe 2007). In addition, it is very stable and can tolerate high temperature and acidic conditions (Richards et al. 2002). It was stated that increase levels of trehalose resulted in enhanced photosynthetic rates and reduced photo-oxidative damage induced by stress. The most common trehalose biosynthesis pathway involves two enzymes: trehalose-6-phosphate synthase (TPS) and T6P-phosphatase (TPP) (Avonce et al. 2010). The expression of these genes (TPS and TPP) improved plant resistance to abiotic stresses (Jang et al. 2003; Miranda et al. 2007). Accumulation of trehalose was reported in Chlorella sp. during osmotic stress (Bremauntz et al. 2011). However, unexpected repression of the gene involved in the synthesis of trehalose was observed in Chlorella strain subjected to UV stress (Poong et al. 2018).
Oxylipins and polyamines
In addition, two other groups of molecules known as oxylipins and polyamines are involved in defence systems. Oxylipins are lipid metabolites generated from unsaturated fatty acids either through chemical auto-oxidation or enzymatically. Oxylipins resulting from the oxidative metabolism of PUFAs are recognized as performing in response to environmental changes and developmentally regulated processes in both terrestrial animals and plants (Andreou et al. 2009). Among eukaryotic algae, oxylipins have been found to be abundant as a defence mechanism against both biotic and abiotic stresses (Bouarab et al. 2004). Algae undergo octadecanoid and/or eicosanoid oxylipin biosynthetic pathways where several enzymes such as lipoxygenases (LOXs) and cyclooxygenase (COX) are involved in the production of a variety of oxylipins. LOXs are the initial and fundamental enzymes in the oxylipin pathway and are omnipresent in plants, prokaryotes, and animals (Andreou and Feussner 2009). The presence of LOXs was reported in Chlorella pyrenoidosa (Zimmerman and Vick 1973; Nuñez et al. 2002). Much of the research about oxylipins focused on the jasmonate family of molecules, which comprises jasmonic acid (JA) and methyl jasmonate (MeJA) (Bouarab et al. 2004; Cosse et al. 2007; Potin et al. 2002). Jasmonic acid (JA) plays diverse functions including defence and stress response activity (Wasternack and Hause 2013). In several microalgae, JA affects growth, levels of the pigments (chlorophyll, carotenoids and astaxanthin) and protein profiles (Czerpak et al. 2006; Lu et al. 2010a, b; Raman and Ravi 2011; Piotrowska-Niczyporuk et al. 2012). In addition, Fujii et al. (1997) suggested the existence of JA in axenic cultures of Dunaliella. However, further research about the role of this molecule in microalgae is needed.
On the other hand, polyamines (PAs) are aliphatic amines which can appear in free form or as conjugated molecules. Their roles as stress alleviators have only recently been recognized (Kumar et al. 2010, 2012; Li et al. 2013). It was proposed that PAs could exert protection to cells through the formation of complexes with anions which led to reduced reactive oxygen species (ROS) production (García-Jiménez and Robaina 2012). Numerous studies have stated the involvement of PAs in response to environmental stress (Kumar et al. 2011, 2012, 2014).
Antioxidant defence
Under optimal growth conditions, ROS or free radicals such as singlet oxygen (1O2) and superoxide (O2·−) are primarily produced as a part of the cellular growth at low levels in various organelles including chloroplasts and mitochondria (Ahmad et al. 2014). However, the ROS homeostasis could be affected by stress, leading to high ROS levels and eventually oxidative stress (Malan et al. 1990; Apel and Hirt 2004; Murik et al. 2014). In chloroplasts, the main reason for ROS production is the suppression of CO2 fixation, linked to the over-reduction of the electron transport chain in thylakoid (Davidson and Schiestl 2001). High ROS levels in chloroplasts may occur via the Mehler reaction where the formation of superoxide occurs owing to the transfer of electrons from photosystem I (PSI) to O2 (Dietz et al. 2016). During oxidative stress, superoxide radicals act as oxidants and facilitate the production of short-lived hydroxyl radicals (Fridovich 1995) which are highly reactive and can induce DNA damage via denaturation (Lesser 2006). Furthermore, ROS hinder the mending of PSII (Nishiyama et al. 2004). It is suggested that both the singlet oxygen and superoxide radical are strong oxidants and both oxidise elongation factor G (EF-G), which participates in the production of the D1 protein (Nishiyama and Murata 2014). Initially, ROS were thought to be toxic by-products generated from aerobic metabolism; however, studies revealed that ROS played important role in signal transduction (Mittler et al. 2004). Thus, a basal level of ROS is vital for the functions of living cells (Mittler 2017). Principally, ROS elevation induces the synthesis of ROS scavenging enzymes, namely ascorbate peroxidase (APX), catalase (CAT), glutathione peroxidase (GPX), superoxide dismutase (SOD) and thioredoxins (TRXs) as well as enzymes involved in the biosynthesis of glutathione (Lemaire et al. 2007; Gill and Tuteja 2010). As the first line of defence, superoxide dismutases (SODs) rapidly convert O2·− to O2 and H2O2 and the generated H2O2 can be converted into H2O by peroxidases, such as APX and GPX found in vacuoles, cell walls and the cytosol (Alscher et al. 2002; Elbaz et al. 2010; Caverzan et al. 2012) (Fig. 2). Induced activities of SODs, APX and GPX under diverse abiotic stress conditions have been reported in microalgae (Fischer et al. 2006; Park et al. 2006; Yildiztugay et al. 2014; Chen et al. 2015; Yanguez et al. 2015). High levels of SOD were reported in Chlorella ellipsoidea in response to temperature stress (Clare et al. 1984). The high APX activity seen in Chlamydomonas under stress conditions indicated that APX potentially contributed to this alga’s tolerance to oxidative stress (Tanaka et al. 2011). In addition, catalase (CAT) is recognised as a dominant enzyme that catalyses the exchange of H2O2 to H2O or other non-toxic molecules (Mittler et al. 2004; Luis et al. 2006). In C. reinhardtii, catalase (CAT) scavenges H2O2 from photorespiration and oxidation of fatty acids (Kato et al. 1997; Vega et al. 2005). Interestingly, CAT activity was correlated with the chlorophyll content during photoacclimation, and exhibited protection against photodynamic damage induced by ROS (Dykens and Shick 1984). Manganese superoxide dismutase (MnSOD) and CAT are the predominant enzymes responsible for mitochondrion protection in C. reinhardtii under metal stress, while SOD and CAT are associated with the recovery of photosynthesis (Aksmann et al. 2014). In addition, CAT activity also contributed to the antioxidant defence and acclimation of the green alga Scenedesmus vacuolatus when exposed to copper (Sabatini et al. 2009). The other reported antioxidant compounds in relation to photosynthesis are carotenoids and α-tocopherols. Carotenoids are essential components found in photosynthetic algae. They are mainly involved in the light-harvesting process during photosynthesis while protecting the photosynthetic system against photo-induced oxidative stress (Vidhyavathi et al. 2008). As antioxidants, carotenoids are capable of quenching 1O2 and excitation energy from chlorophyll, hence reducing the production and accumulation of 1O2 (Demmig-Adams 1990; Young 1991). It was also noted that ROS triggers the parallel accumulation of carotenoids in Dunaliella (Ye et al. 2008) and amplified expression of carotenoid biosynthesis-related genes was observed in Haematococcus pluvialis under several stress conditions (Vidhyavathi et al. 2008). Furthermore, tocopherols (Toc) also have vital roles in maintaining redox homoeostasis and their biosynthesis is found to increase during stress. Toc functions by deactivating ROS (mainly 1O2 and OH·) generated in thylakoid membranes during photosynthesis and by inhibiting lipid peroxidation via removal of lipid peroxyl radicals (LOO·) (Maeda et al. 2005). Although ROS are produced in both normal and stress conditions, cell defence mechanisms counter radical damage by reducing ROS production and facilitating ROS removal.

ROS and antioxidant defence mechanism. The generated O2·− due to abiotic stress is converted to H2O2 by SOD activity, then three primary enzymes, catalase (CAT), glutathione peroxidase (GPX) and ascorbate peroxidase (APX) catalyse, the conversion of H2O2 to H2O
Photosynthetic efficiency
Alterations in the environmental factors could cause an inequity between energy consumption and supply in photosynthetic algae, leading to alterations to the photosynthetic apparatus. In algae and higher plants, the reaction centre of PSII is bounded by the central antenna (CP47 and CP43) and the outer antenna, which comprises the less-abundant minor light-harvesting complex I (LHC) and the major light-harvesting complex II (LHCII) (Elrad et al. 2002). These LHCs comprise proteins that bind chlorophylls (Chl a and Chl b). The LHCb gene family encodes both the minor LHC proteins and the LHCII. In the thylakoid membrane, LHCII is the most abundant pigment–protein complex and about half of all chlorophylls are associated with it. The main role of LHCII is to receive and pass on the energy to the reaction centre of PSII and participate in controlling the supply of excitation energy among PSII and PSI. Transcriptional control of the genes encoding LHCII plays an essential role in antenna size adjustment (Elrad et al. 2002; Teramoto et al. 2001). Algae regulate the expression of nuclear-encoded chlorophyll a–b light-harvesting complex genes, (LHCa, LHCb), thereby controlling the antenna size. For example, when C. reinhardtii was grown under light-limiting conditions, the up-regulation of LHC gene expression resulted in a larger antenna size (Elrad et al. 2002). Moreover, environmental changes can influence the subunit arrangement of reaction centres (PSI and PSII). For example, the psbC gene encodes a PSII chlorophyll-binding protein, functioning as an oxygen-evolving enzyme and participate in water-splitting of photosynthesis (Qian et al. 2009a), and is influenced by severe circumstances like temperature stress (Chong et al. 2011). The gene psaB, encoding one of the subunits of PSI (Qian et al. 2009a), was down-regulated in Chlorella vulgaris when this alga was in contact to toxic chemicals (Qian et al. 2011). Although other stresses, such as salinity, might have an influence on the expression of these genes, high light illumination was more commonly associated with their regulation (Shapira et al. 1997; Nama et al. 2015). It is noteworthy to consider that gene expression is controlled at several steps starting with transcription of a gene, splicing, editing steps, and finally mature mRNA. The expression of chloroplast genes (psaB, psbC and rbcL) was initially thought to be regulated at the post-transcriptional level, associated with mRNA stability (Pfannschmidt 2003). While it is likely that photosynthetic gene expression is controlled at all levels of expression (Qian et al. 2008), studying their expression at the transcriptomic level can provide a suitable resolution of their synthesis.
Photosynthetic fixation of CO2 is vital for the development and growth of algae, supplying the carbohydrates needed for metabolism, as well as for structural constituents and supplying cellular structural units. Also, photosynthesis acts as a global sensor of environmental stress that causes an imbalance in cellular energy which leads to the distinct changes in redox chemistry linked to thylakoid membranes and adjustment of cellular sugar grade (Biswal et al. 2011). Photosystems and key enzymes associated with carbon dioxide fixation, such as ribulose-1,5-bisphosphate carboxylase/oxygenase (RubisCO) play critical roles in photosynthesis. The expression levels of genes involved in photosystem structure and RubisCO subunits were significantly influenced by various stresses (Qian et al. 2009a, b, 2012; Luo et al. 2015). For example, rbcL, which encodes the large subunit of RubisCO (Spreitzer and Salvucci 2002) was up-regulated upon a temperature increase to 20 °C, but with a further temperature rise, its expression was down-regulated (Deng et al. 2014b). Also, expression of rbcL was inhibited under desiccation, low and high salinity plus at temperatures under and beyond the normal ambient temperature (Xu et al. 2013).
Photoinhibition associated with impairment to the D1 protein of PSII can occur whenever algal cells are receiving light, and the impairment is constantly fixed by de novo synthesis of D1, after the activation of the reaction centre to achieve a balance among the photodamage of PSII and its re-production (Takahashi and Murata 2008). Three key steps in PSII repair are conspicuously sensitive to abiotic stress, namely the breakdown of the D1 protein in photodamaged PSII, generation of pre-D1 and its conversion to the mature D1 protein (Nishiyama and Murata 2014). It was suggested that adverse stresses might not increase photodamage directly but rather affect the PSII repair process by hindering the (re)synthesis of D1 (Takahashi and Murata 2008). In chloroplasts of C. reinhardtii, the expression of psbA was highly controlled by mRNA processing, especially during the initiation of translation (Mulo et al. 2012). Besides the regulation of D1 protein synthesis during photoinhibition, amino acid changes in the D1 protein were reported in response to temperature changes. Together with the physicochemical properties of the thylakoid membrane, these changes might contribute to the structural flexibility required for electron transfer in the PSII system (Giardi et al. 1997; Lukes et al. 2014). Table 1 shows a list of genes related to acclimation and repair strategies of algae under different stress conditions.
DNA damage and repair mechanisms
Among cellular biomolecules only DNA is regularly repaired and preserved, while the others such as proteins and lipids are broken down when damaged, followed by de novo biosynthesis. Cells have developed several checkpoints during DNA replication, where the cell cycle is arrested to avoid error by providing additional time for DNA repair prior to mitosis (Ševčovičová et al. 2008). In the process, DNA damage is detected and repaired to ensure the integrity and proper functioning of the genome and the organism (Holzinger and Karsten 2013). Discontinuities that occur in one strand of the DNA double helix can also be referred to as single-strand breaks (SSBs). SSBs can happen directly via the disintegration of the oxidised sugar moiety or indirectly throughout the DNA base-excision repair (BER). In addition, functional errors of enzymes such as DNA topoisomerase 1 (TOP1) may result in SSBs (Caldecott 2008). Both ROS and ultraviolet radiation (UVR) are known as the primary causes of SSBs in DNA (Bray and West 2005). Alternatively, if discontinuities take place in double-stranded DNA, these are called DNA double-strand breaks (DSBs) and these can be made by endogenous and exogenous factors including mistakes throughout the DNA replication process, ionising radiation, or particular chemicals (Dudas and Chovanec 2004). A DSB is considered to be the most relevant signal-activating recombination process in the cell (Barlow et al. 2008). Living cells have developed a variety of mechanisms to mitigate the effects of DNA damage. BER is the first type of repair and is used to repair SSB damage by the processes in which DNA glycosylases, namely methyladenine glycosylase and uracil glycosylase, play key roles (Seeberg et al. 1995). BER is also used for dealing with lesions resulting from strong alkylating agents and ionising radiation (Seeberg et al. 1995). Though little knowledge about BER in algae, its presence has been stated in C. reinhardtii (Vlcek et al. 2008). On the other hand, DSBs are usually fixed by non-homologous end joining (NHEJ) or homologous recombination (HR). HR is the favourite manner of repair in numerous unicellular organisms, whereas NHEJ is the common pathway in plants and animals (Nagaria et al. 2013; Plecenikova et al. 2014). It was suggested that C. reinhardtii principally carried out DNA repair by involving NHR, rather than HR (Vlcek et al. 2008; Plecenikova et al. 2013). The contribution of NHEJ in the repair of DSBs in C. reinhardtii was observed under heat shock and gamma radiation exposure (Boreham and Mitchel 1993). The NHEJ pathway is facilitated by the DNA-dependent protein kinase catalytic subunit (DNA-PKcs), the Ku70–Ku80 heterodimer (Ku complex), and the DNA ligase IV–Xrcc4 complex (Ninomiya et al. 2004). Formation of cyclobutane pyrimidine dimers (CPDs) is another type of DNA damage and its occurrence as a result of UVR exposure was reported in green algae (Lud et al. 2001). CPDs can be fixed by a light-dependent procedure named photoreactivation or by dark repair mechanisms, namely nucleotide excision repair (NER) (Hercegova et al. 2008). During photoreactivation, the enzyme photolyase is triggered by blue light and UVA emission. It splits the covalent bond of the pyrimidine dimer (Sancar 1990; Britt 2004). The enzyme photolyase is perhaps one of the simplest and oldest DNA repair systems found ubiquitously in all organisms. Photolyase from Ostreococcus tauri (a green alga) has been characterised (Usman et al. 2009). A gene called PHR2 showing DNA photolyase activity in chloroplast and the nucleus was identified in C. reinhardtii (Petersen et al. 1999) and Dunaliella salina (Cheng et al. 2007). On the other hand, NER recognises lesions that distort the DNA helix. It is the most flexible and adaptable DNA repair pathway of organisms, as it works with a wide variety of structurally distinct DNA lesions (Costa et al. 2003). NER is well characterised in yeast and mammalian cells, and has been presented to be composed of approximately 30 gene products (Sinha and Hader 2002). NER has been observed to also function in the unicellular green alga C. reinhardtii (Hsu et al. 2000). Five proteins associated with NER were identified in C. reinhardtii, including the excision repair protein ERCC1. The DNA damage and associated repair mechanisms are summarised in Fig. 3.

DNA damage and repair mechanisms. Single-strand breaks (SSB), double-strand breaks (DSB) and cyclobutane pyrimidine dimers (CPD) are the common DNA lesions which trigger a range of specific repair mechanisms in algae to protect genomic integrity
Programmed cell death (PCD)
Organisms’ tolerance or sensitivity to abiotic stress may also be linked to the stimulation or suppression of death in the cells (Darehshouri et al. 2008). Breakdowns of DNA, chromatin condensation, the release of cytochrome c, the contribution of caspase-like enzymes, a reduction in photosynthetic activity, vacuolization of cytoplasm, ATP depletion, and disruption of membrane integrity have all been observed to convoy cell death following various kinds of inductions (Franklin and Berges 2004; Geitmann et al. 2004; Andronis and Roubelakis-Angelakis 2010). The two terms apoptosis and PCD are often confused. PCD results from changes in the gene expression of dying cells and is mediated by the activity of a series of enzymes resulting in a distinctive morphology of the dead cells. On the other hand, apoptosis refers to a series of morphological features found in dead/dying metazoan cells throughout/after the initiation of PCD (Franklin et al. 2006). PCD is a cellular suicide process controlled genetically that removes unwanted or damaged cells (Vaux and Korsmeyer 1999). The presence of a PCD process in single-cell organisms has also been recognised (Zuo et al. 2012). It is hypothesised that the essential factor in the cell death pathway was available in the genome of primary eukaryotes via ancient viral infection, prior to the development of multicellular organisms (Segovia et al. 2003). Stress might cause severe damage to DNA via the introduction of alterations to its structure and conformation and, in spite of DNA repair mechanisms, cells might then initiate mechanisms leading to programmed cell death. Table 2 shows some well-studied examples of PCD in microalgae. The hypotheses concerning the advantages of PCD in unicellular organisms include the fact that PCD can eliminate impaired and/or ageing cells and result in the release of nutrients to supplement the nutritional necessities of other non-PCD algae as well as to maximise the biological fitness of the community (Huang et al. 2016). Caspase or metacaspase gene activity can be used to determine PCD occurrence in the cell. In animals, caspase proteins are cysteine proteases that are vital regulators of PCD. However, structural homologs of caspase genes are not found in plants. As an alternative, they contain metacaspases, which are also found in organisms such as protozoa, fungi, bacteria and algae (Saheb et al. 2014; Fagundes et al. 2015). Increased level of caspase-like protein or metacaspase was reported in green algal cells (Dunaliella viridis) experiencing PCD under different stressors (Jiménez et al. 2009). Moreover, it was suggested that metacaspases might contribute in other metabolic procedures including signal transduction cascades, stress acclimation and regulation of protein aggregations, rather than entirely being accountable for PCD initiation (Bidle and Falkowski 2004; Bidle et al. 2007; Richie et al. 2007; Lee et al. 2010; Huang et al. 2016). Generally, a greater understanding of processes involved in phytoplankton cell death will possibly deliver knowledge about intercellular signalling, population regulation and bloom development.
Omics applications and future directions
Knowledge about algae biological basics and regulatory networks can assist in enlightening their successful distribution, proliferation, and survival. Furthermore, understanding their responses to stress conditions is required if successful outcomes are needed for the potential role of algae in dealing with increased demand for biomass, feed and food (Moreno-Risueno et al. 2010). A large body of literature has been devoted to genomic studies to define the functions and roles of genes related to stress. This knowledge could be translated into practical industrial applications, for example, the production of lipids and antioxidants induced by stress. Lipid content is extremely important to biofuel production, whereas antioxidants such as astaxanthin are recognised to be the main biomolecules in the pharmaceutical industry (Hu et al. 2008; Ambati et al. 2014). Although the desired product can be obtained by introducing the associated stress, if the growth is compromised by the stress, the overall productivity is also reduced. Lately, advances in ‘omics’ sciences including genomics, proteomics, transcriptomics, and metabolomics have provided deeper understanding about algae (Nouri et al. 2015). For instance, genomics studies contributed to the complete sequence of C. reinhardtii (Merchant et al. 2007). Grossman et al. (2007) have compiled genomic information of four algae including C. reinhardtii for a better understanding of many metabolic pathways. In the area of transcriptomics, the development of a microarray for C. reinhardtii containing 10,000 oligonucleotide sequences which cover nearly its full genome (http://www.chlamy.org) may further contribute to algal research. Similarly, C. reinhardtii proteomics would provide valuable information about the functional groups of proteins and cellular subunits (Stauber and Hippler 2004). Furthermore, metabolomics, a comprehensive and quantitative analysis of metabolites in a living cell at a particular moment could reveal the factual combination of protein expression and gene regulation as well as the influence of the environment (Fiehn 2002). Current advances in bioinformatics and analytical tools have improved our ability to analyse a large number of metabolites, assess metabolic alterations in response to exterior stimuli and explain metabolic pathways (Kumar et al. 2016). A series of analytical systems have been established such as nuclear magnetic resonance (NMR) and mass spectrometry (MS). Combination of chromatographic separation with different mass spectra is presented in systems such as gas chromatography (GC–MS) and liquid chromatography (LC–MS). Owing to a high level of structural variety, polarity and molecular weight differences among metabolites, no single analytical tool is able to measure and detect all conceivable metabolites in one run (Kueger et al. 2012). Bölling and Fiehn (2005) demonstrated metabolite profiles generated using GC–MS system could distinguish C. reinhartii cell growth under different nutrient deficiency conditions. Finally, ‘omics’ analyses are crucial in understanding the complete processes of molecular networks in response to abiotic stress. It is essential to define the roles of newly identified stress-responsive molecules to explain the complex abiotic stress responses of algae.
Interestingly, the genomic information not directly encoded in the DNA sequence can also be transferred among generations. For example, non-genetic information can be transferred when DNA or its related proteins are altered, such as through DNA methylation and histone modifications (Chinnusamy and Zhu 2009; Cortijo et al. 2014). Together, these alterations are called epigenetic modifications. It has now been established that epigenetic alterations can be passed on not only via mitotic cell division but also from parent to offspring (Johannes et al. 2008; Cortijo et al. 2014; Audergon et al. 2015). Mutation accumulation experiments have suggested that spontaneous epigenetic modifications happen much like genetic mutations. Nevertheless, epigenetic mutations happen at a faster rate, although they are less stable than genetic mutations (Kronholm et al. 2017). Kornholm et al. (2017) summarized that epigenetic variation can contribute to adaptation, although environmental selection plays an important role in controlling its extent.
Furthermore, various gene manipulation or gene editing approaches such as CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats)/Cas9 (CRISPR-associated gene 9) can be applied to generate stress-tolerant algae (Gan and Mags 2017). CRISPR/Cas9 technology has been adapted to the model microalga C. reinhardtii to efficiently produce targeted gene editing while reducing Cas9-associated toxicity (Jiang et al. 2014; Shin et al. 2016; Greiner et al. 2017). CRISPR/Cas9 can directly edit the genome precisely at the region of interest by either NHEJ or HDR mutations, which leads to a defined DNA replacement, deletion and insertion (Xu et al. 2014). With this technology, further accomplishments can also be achieved in the functional study of algal proteins and genes associated with abiotic stress, understanding of their biology, and description of their metabolic pathways (Lander et al. 2016; Gan and Mags 2017). There is an extensive relation among constituents of several regulatory, signalling and metabolic pathways, which results in adaptation or responses (Nakashima et al. 2009; Garg et al. 2014; Mickelbart et al. 2015). In many instances, it is challenging to find the role of the genes using knockout of a single gene since this may not produce any desired phenotype. However, using the CRISPR–Cas9 system several genes can be knocked out at the same time, which can beat the issue due to genetic redundancy (Jain 2015). Although CRISPR technology is still in its infancy in algal studies, it is hoped to be of more use in the near future where it can contribute to the improvement of our understanding on algal metabolism and responses to environment, and also be used in producing more stress-tolerant algae to be used in industry.
Author contribution statement
All authors contributed to this work and commented on the manuscript at all stages and revised it accordingly. BB, SYG, P-EL and SMP contributed in the conception of the content of the manuscript. BB wrote the manuscript, S-YG substantially helped in improvement of the manuscript by her comments and revisions, P-EL supervised the work and revised the manuscript, figures and tables. JB and S-MP substantially helped in both English and scientific revision.
References
Ahmad I, Hellebust JA (1988) The relationship between inorganic nitrogen metabolism and proline accumulation in osmoregulatory responses of two euryhaline microalgae. Plant Physiol 88(2):348–354
Ahmad P, Jamsheed S, Hameed A, Rasool S, Sharma I, Azooz MM, Hasanuzzaman M (2014) Oxidative damage and antioxidants in plants. In: Ahmad P (ed) Oxidative damage to plants. Elsevier, New York, pp 345–367
Ahmad I, Sharma AK, Daniell H, Kumar S (2015) Altered lipid composition and enhanced lipid production in green microalga by introduction of brassica diacylglycerol acyltransferase 2. Plant Biotechnol J 13(4):540–550
Aksmann A, Pokora W, Bascik-Remisiewicz A, Dettlaff-Pokora A, Wielgomas B, Dziadziuszko M, Tukaj Z (2014) Time-dependent changes in antioxidative enzyme expression and photosynthetic activity of Chlamydomonas reinhardtii cells under acute exposure to cadmium and anthracene. Ecotoxicol Environ Saf 110:31–40
Allakhverdiev SI, Murata N (2004) Environmental stress inhibits the synthesis de novo of proteins involved in the photodamage-repair cycle of Photosystem II in Synechocystis sp. PCC 6803. Biochim Biophys Acta 1657:23–32
Allakhverdiev SI, Kinoshita M, Inaba M, Suzuki I, Murata N (2001) Unsaturated fatty acids in membrane lipids protect the photosynthetic machinery against salt-induced damage in Synechococcus. Plant Physiol 125(4):1842–1853
Alscher RG, Erturk N, Heath LS (2002) Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J Exp Bot 53(372):1331–1341
Ambati RR, Phang SM, Ravi S, Aswathanarayana RG (2014) Astaxanthin: sources, extraction, stability, biological activities and its commercial applications-a review. Mar Drugs 12:128–152
Amini S, Ghobadi C, Yamchi A (2015) Proline accumulation and osmotic stress: an overview of P5CS gene in plants. J Plant Mol Breed 3(2):44–55
An M, Mou S, Zhang X, Zheng Z, Ye N, Wang D, Zhang W, Miao J (2013a) Expression of fatty acid desaturase genes and fatty acid accumulation in Chlamydomonas sp. ICE-L under salt stress. Bioresour Technol 149:77–83
An M, Mou S, Zhang X, Ye N, Zheng Z, Cao S, Xu D, Fan X, Wang Y, Miao J (2013b) Temperature regulates fatty acid desaturases at a transcriptional level and modulates the fatty acid profile in the Antarctic microalga Chlamydomonas sp ICE-L. Bioresour Technol 134:151–157
Andreou A, Feussner I (2009) Lipoxygenases-structure and reaction mechanism. Phytochemistry 70(13–14):1504–1510
Andreou A, Brodhun F, Feussner I (2009) Biosynthesis of oxylipins in non-mammals. Prog Lipid Res 48(3–4):148–170
Andronis EA, Roubelakis-Angelakis KA (2010) Short-term salinity stress in tobacco plants leads to the onset of animal-like PCD hallmarks in planta in contrast to long-term stress. Planta 231(2):437
Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 55:373–399
Arbona V, Manzi M, de Ollas C, Gomez-Cadenas A (2013) Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int J Mol Sci 14:4885–4911
Audergon PN, Catania S, Kagansky A, Tong P, Shukla M, Pidoux AL, Allshire RC (2015) Epigenetics. Restricted epigenetic inheritance of H3K9 methylation. Science 348(6230):132–135
Avonce N, Wuyts J, Verschooten K, Vandesteene L, Van Dijck P (2010) The Cytophaga hutchinsonii ChTPSP: first characterized bifunctional TPS-TPP protein as putative ancestor of all eukaryotic trehalose biosynthesis proteins. Mol Biol Evol 27:359–369
Azachi M, Sadka A, Fisher M, Goldshlag P, Gokhman I, Zamir A (2002) Salt induction of fatty acid elongase and membrane lipid modifications in the extreme halotolerant alga Dunaliella salina. Plant Physiol 129:1320–1329
Ball SG, Morell MK (2003) From bacterial glycogen to starch: understanding the biogenesis of the plant starch granule. Annu Rev Plant Biol 54:207–233
Barlow JH, Lisby M, Rothstein R (2008) Differential regulation of the cellular response to DNA double-strand breaks in G1. Mol Cell 30:73–85
Bartley ML, Boeing WJ, Corcoran AA, Holguin FO, Schaub T (2013) Effects of salinity on growth and lipid accumulation of biofuel microalga Nannochloropsis salina and invading organisms. Biomass Bioenerg 54:83–88
Beardall J, Raven JA (2004) The potential effects of global climate change on microalgal photosynthesis, growth and ecology. Phycologia 43(1):26–40
Bidle KD, Falkowski PG (2004) Cell death in planktonic, photosynthetic microorganisms. Nat Rev Microbiol 2:643–655
Bidle KD, Haramaty L, Barcelos ERJ, Falkowski P (2007) Viral activation and recruitment of metacaspases in the unicellular coccolithophore, Emiliania huxleyi. Proc Natl Acad Sci USA 104:6049–6054
Biswal B, Joshi PN, Raval MK, Biswal UC (2011) Photosynthesis, a global sensor of environmental stress in green plants: stress signalling and adaptation. Curr Sci India 101(1):47–56
Bohnert HJ, Nelson DE, Jensen RG (1995) Adaptations to environmental stresses. Plant Cell 7(7):1099–1111
Bölling C, Fiehn O (2005) Metabolite profiling of Chlamydomonas reinhardtii under nutrient deprivation. Plant Physiol 139(4):1995–2005
Bopp L, Aumont O, Cadule P, Alvain S, Gehlen M (2005) Response of diatoms distribution to global warming and potential implications: a global model study. Geophys Res Lett 32:L19606
Boreham DR, Mitchel RE (1993) DNA repair in Chlamydomonas reinhardtii induced by heat shock and gamma radiation. Radiat Res 135(3):365–371
Borowitzka LJ, Brown AD (1974) The salt relations of marine and halophilic species of the unicellular green alga. Dunaliella Arch Mikrobiol 96(1):37–52
Bouarab K, Adas F, Gaquerel E, Kloareg B, Salaun JP, Potin P (2004) The innate immunity of a marine red alga involves oxylipins from both the eicosanoid and octadecanoid pathways. Plant Physiol 135(3):1838–1848
Boyle NR, Page MD, Liu B, Blaby IK, Casero D, Kropat J, Cokus SJ, Hong-Hermesdorf A et al (2012) Three acyltransferases and nitrogen-responsive regulator are implicated in nitrogen starvation-induced triacylglycerol accumulation in Chlamydomonas. J Biol Chem 287:15811–15825
Bray CM, West CE (2005) DNA repair mechanisms in plants: crucial sensors and effectors for the maintenance of genome integrity. New Phytol 168(3):511–528
Bremauntz MP, Torres-Bustillos LG, Cañizares-Villanueva R-O, Duran-Paramo E, Fernández-Linares L (2011) Trehalose and sucrose osmolytes accumulated by algae as potential raw material for bioethanol. Nat Resour 2:173–179
Brett M, Müller-Navarra D (1997) The role of highly unsaturated fatty acids in aquatic foodweb processes. Freshwater Biol 38(3):483–499
Britt AB (2004) Repair of DNA damage induced by solar UV. Photosynth Res 81:105–112
Bukau B, Horwich AL (1998) The Hsp70 and Hsp60 chaperone machines. Cell 92(3):351–366
Caldecott KW (2008) Single-strand break repair and genetic disease. Nat Rev 9(8):619–631
Calow P (1991) Physiological costs of combating chemical toxicants: ecological implications. Comp Biochem Physiol C 100(1–2):3–6
Caverzan A, Passaia G, Rosa SB, Ribeiro CW, Lazzarotto F, Margis-Pinheiro M (2012) Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet Mol Biol 35(4):1011–1019
Chankova S, Mitrovska Z, Miteva D, Oleskina YP, Yurina NP (2013) Heat shock protein HSP70B as a marker for genotype resistance to environmental stress in Chlorella species from contrasting habitats. Gene 516:184–189
Chen JE, Smith AG (2012) A look at diacylglycerol acyltransferases (DGATs) in algae. J Biotechnol 162:28–39
Chen L, Mao F, Kirumba GC, Jiang C, Manefield M, He Y (2015) Changes in metabolites, antioxidant system, and gene expression in Microcystis aeruginosa under sodium chloride stress. Ecotoxicol Environ Saf 122:126–135
Cheng L, Qiao DR, Lu XY, Xiong Y, Bai LH, Xu H, Yang Y, Cao Y (2007) Identification and expression of the gene product encoding a CPD photolyase from Dunaliella salina. J Photochem Photobiol B 87(2):137–143
Chinnusamy V, Zhu JK (2009) Epigenetic regulation of stress responses in plants. Curr Opin Plant Biol 12(2):133–139
Chong GL, Chu WL, Othman RY, Phang SM (2011) Differential gene expression of an Antarctic Chlorella in response to temperature stress. Polar Biol 34(5):637–645
Clare DA, Rabinowitch HD, Fridovich I (1984) Superoxide dismutase and chilling injury in Chlorella ellipsoidea. Arch Biochem Biophys 231(1):158–163
Cortijo S, Wardenaar R, Colomé-Tatché M, Gilly A, Etcheverry M, Labadie K, Caillieux E, Hospital F et al (2014) Mapping the epigenetic basis of complex traits. Science 343:1145–1148
Cosse A, Leblanc C, Potin P (2007) Dynamic defense of marine macroalgae against pathogens: from early activated to gene-regulated responses. Adv Bot Res 46:221–266
Costa RM, Chigancas V, Galhardo Rda S, Carvalho H, Menck CF (2003) The eukaryotic nucleotide excision repair pathway. Biochimie 85(11):1083–1099
Crowe JH (2007) Trehalose as a “chemical chaperone”. Molecular aspects of the stress response: Chaperones, membranes and networks. Springer, Berlin, pp 143–158
Czerpak R, Piotrowska A, Szulecka K (2006) Jasmonic acid affects changes in the growth and some components content in alga Chlorella vulgaris. Acta Physiol Plant 28:195–203
Daniel RM, Dines M, Petach HH (1996) The denaturation and degradation of stable enzymes at high temperatures. Biochem J 317(1):1–11
Darehshouri A, Affenzeller M, Lütz-Meindl U (2008) Cell death upon H2O2 induction in the unicellular green alga Micrasterias. Plant Biol 10(6):732–745
Davidson JF, Schiestl RH (2001) Mitochondrial respiratory electron carriers are involved in oxidative stress during heat stress in Saccharomyces cerevisiae. Mol cell biol 21(24):8483–8489
Demmig-Adams B (1990) Carotenoids and photoprotection in plants: a role for the xanthophyll zeaxanthin. BBA-Bioenerg 1020(1):1–24
Deng Y, Wang X, Guo H, Duan D (2014a) A trehalose-6-phosphate synthase gene from Saccharina japonica (Laminariales, Phaeophyceae). Mol Biol Rep 41(1):529–536
Deng Y, Zhan Z, Tang X, Ding L, Duan D (2014b) Molecular cloning and expression analysis of RbcL cDNA from the bloom-forming green alga Chaetomorpha valida (Cladophorales, Chlorophyta). J Appl Phycol 26(4):1853–1861
Dietz KJ, Turkan I, Krieger-Liszkay A (2016) Redox and Reactive oxygen species-dependent signaling into and out of the photosynthesizing chloroplast. Plant Physiol 171:1541–1550
Dittami SM, Gravot A, Renault D, Goulitquer S, Eggert A, Bouchereau A, Boyen C, Tonon T (2011) Integrative analysis of metabolite and transcript abundance during the short-term response to saline and oxidative stress in the brown alga Ectocarpus siliculosus. Plant Cell Environ 34(4):629–642
Dong MT, Zhang XW, Zhuang ZM, Zou J, Ye NH, Xu D, Mou SL, Liang CW, Wang WQ (2012) Characterization of the LhcSR gene under light and temperature stress in the green alga ulva linza. Plant Mol Biol Rep 30(1):10–16
Dudas A, Chovanec M (2004) DNA double-strand break repair by homologous recombination. Mutat Res 566:131–167
Dykens JA, Shick JM (1984) Photobiology of the symbiotic sea-anemone, Anthopleura-Elegantissima—defenses against photodynamic effects, and seasonal photoacclimatization. Biol Bull 167(3):683–697
Elbaz A, Wei YY, Meng Q, Zheng Q, Yang ZM (2010) Mercury-induced oxidative stress and impact on antioxidant enzymes in Chlamydomonas reinhardtii. Ecotoxicology 19:1285–1293
Elrad D, Niyogi KK, Grossman AR (2002) A major light-harvesting polypeptide of photosystem II functions in thermal dissipation. Plant Cell 14(8):1801–1816
Elsayed KNM, Kolesnikova TA, Noke A, Klock G (2017) Imaging the accumulated intracellular microalgal lipids as a response to temperature stress. 3 Biotech 7(1):41
Eom HS, Park S, Lee C-G, Jin ES (2005) Gene expression profiling of an eukaryotic microalga, Haematococcus pluvialis. J Microbiol Biotech 15:1060–1066
Fagundes D, Bohn B, Cabreira C, Leipelt F, Dias N, Bodanese-Zanettini MH, Cagliari A (2015) Caspases in plants: metacaspase gene family in plant stress responses. Funct Integr Genom 15(6):639–649
Fakhry EM, El Maghraby DM (2015) Lipid accumulation in response to nitrogen limitation and variation of temperature in Nannochloropsis salina. Bot Stud 56(1):6
Fan J, Cui Y, Wan M, Wang W, Li Y (2014) Lipid accumulation and biosynthesis genes response of the oleaginous Chlorella pyrenoidosa under three nutrition stressors. Biotechnol Biofuels 7:17
Fiehn O (2002) Metabolomics-the link between genotypes and phenotypes. Plant Mol Biol 48:155–171
Fischer BB, Eggen RIL, Trebst A, Krieger-Liszkay A (2006) The glutathione peroxidase homologous gene GPXH in Chlamydomonas reinhardtii is upregulated by singlet oxygen produced in photosystem II. Planta 223:583–590
Franklin DJ, Berges JA (2004) Mortality in cultures of the dinoflagellate Amphidinium carterae during culture senescence and darkness. Proc Biol Sci 271(1553):2099–2107
Franklin DJ, Brussaard CP, Berges JA (2006) What is the role and nature of programmed cell death in phytoplankton ecology? Eur J Phycol 41:1–14
Fridovich I (1995) Superoxide radical and superoxide dismutases. Annu Rev Biochem 64(1):97–112
Fu J, Momcilovic I, Clemente TE, Nersesian N, Trick HN, Ristic Z (2008) Heterologous expression of a plastid EF-Tu reduces protein thermal aggregation and enhances CO2 fixation in wheat (Triticum aestivum) following heat stress. Plant Mol Biol 68(3):277–288
Fujii S, Yamamoto R, Miyamoto K, Ueda J (1997) Occurrence of jasmonic acid in Dunaliella (Dunaliellales, Chlorophyta). Phycol Res 45(4):223–226
Fulda S, Gorman AM, Hori O, Samali A (2010) Cellular stress responses: cell survival and cell death. Int J Cell Biol 6:214074
Gan SY, Maggs CA (2017) Random mutagenesis and precise gene editing technologies: applications in algal crop improvement and functional genomics. European J Phycol 52(4):466–481
Garcia-Gomez C, Mata MT, Van Breusegem F, Segovia M (2016) Low-steady-state metabolism induced by elevated CO2 increases resilience to UV radiation in the unicellular green-algae Dunaliella tertiolecta. Enviro Exp Bot 132:163–174
García-Jiménez P, Robaina RR (2012) Effects of ethylene on tetrasporogenesis in Pterocladiella Capillacea (Rhodophyta) 1. J Phycol 48:710–715
Garg AK, Kim JK, Owens TG, Ranwala AP, Choi YD, Kochian LV, Wu RJ (2002) Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses. Proc Natl Acad Sci USA 99:15898–15903
Garg R, Verma M, Agrawal S, Shankar R, Majee M, Jain M (2014) Deep transcriptome sequencing of wild halophyte rice, Porteresia coarctata, provides novel insights into the salinity and submergence tolerance factors. DNA research 21(1):69–84
Geider RJ, La Roche J (2002) Redfield revisited: variability of C: N: P in marine microalgae and its biochemical basis. Eur J Phycol 37(1):1–7
Geitmann A, Franklin-Tong VE, Emons AC (2004) The self-incompatibility response in Papaver rhoeas pollen causes early and striking alterations to organelles. Cell Death Differ 11:812–822
Gerloff-Elias A, Barua D, Molich A, Spijkerman E (2006) Temperature- and pH-dependent accumulation of heat-shock proteins in the acidophilic green alga Chlamydomonas acidophila. FEMS Microbiol Ecol 1 56(3):345–354
Giardi MT, Masojídek J, Godde D (1997) Effects of abiotic stresses on the turnover of the D1 reaction centre II protein. Physiol Plant 101(3):635–642
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48(12):909–930
Gonzalez-Fernandez C, Ballesteros M (2012) Linking microalgae and cyanobacteria culture conditions and key-enzymes for carbohydrate accumulation. Biotechnol Adv 30(6):1655–1661
Greiner A, Kelterborn S, Evers H, Kreimer G, Sizova I, Hegemann P (2017) Targeting of photoreceptor genes in Chlamydomonas reinhardtii via zinc-finger nucleases and CRISPR/Cas9. Plant Cell 29(10):2498–2518
Grossman AR, Croft M, Gladyshev VN, Merchant SS, Posewitz MC, Prochnik S, Spalding MH (2007) Novel metabolism in Chlamydomonas through the lens of genomics. Curr Opin Plant Biol 10(2):190–198
Gupta SC, Sharma A, Mishra M, Mishra RK, Chowdhuri DK (2010) Heat shock proteins in toxicology: how close and how far? Life Sci 86(11):377–384
Guy CL (1990) Cold acclimation and freezing stress tolerance: role of protein metabolism. Annu Rev Plant Physiol 41:187–223
Hagemann M (2016) Coping with high and variable salinity: molecular aspects of compatible solute accumulation. In: Borowitzka M, Beardall J, Raven J (eds) The physiology of microalgae. Developments in applied phycology, vol 6. Springer, Cham, pp 359–372
Harley CD, Randall Hughes A, Hultgren KM, Miner BG, Sorte CJ, Thornber CS, Rodriguez LF, Tomanek L, Williams SL (2006) The impacts of climate change in coastal marine systems. Ecol Lett 9(2):228–241
Hawkins AJS (1991) Protein turnover: a functional appraisal. Funct Ecol 5(2):222–233
Hayat S, Hayat Q, Alyemeni MN, Wani AS, Pichtel J, Ahmad A (2012) Role of proline under changing environments: a review. Plant Signal Behav 7(11):1456–1466
Hema R, Senthil-Kumar M, Shivakumar S, Chandrasekhara Reddy P, Udayakumar M (2007) Chlamydomonas reinhardtii, a model system for functional validation of abiotic stress responsive genes. Planta 226(3):655–670
Hercegova A, Sevcovicova A, Galova E (2008) UV light-induced DNA damage detection in the unicellular green alga Chlamydomonas reinhardtii. Biologia 63(6):958–961
Hershko A, Ciechanover A (1998) The ubiquitin system. Annu Rev Biochem 67:425–479
Holzinger A, Karsten U (2013) Desiccation stress and tolerance in green algae: consequences for ultrastructure, physiological and molecular mechanisms. Front Plant Sci 4:327
Honjoh K, Yoshimoto M, Joh T, Kajiwara T, Miyamoto T, Hatano S (1995) Isolation and characterization of hardening-induced proteins in Chlorella vulgaris C-27: identification of late embryogenesis abundant proteins. Plant Cell Physiol 36(8):1421–1430
Honjoh KI, Matsumoto H, Shimizu H, Ooyama K, Tanaka K, Oda Y, Takata R, Joh T, Suga K, Miyamoto T, Iio M, Hatano S (2000) Cryoprotective activities of group 3 late embryogenesis abundant proteins from Chlorella vulgaris C-27. Biosci Biotechnol Biochem 64(8):1656–1663
Hsu T, Sheu R, Lai Y (2000) Possible involvement of a 72-kDa polypeptide in nucleotide excision repair of Chlorella pyrenoidosa identified by affinity adsorption and repair synthesis assay. Plant Sci 156(1):95–102
Hu CA, Delauney AJ, Verma DP (1992) A bifunctional enzyme (delta 1-pyrroline-5-carboxylate synthetase) catalyzes the first two steps in proline biosynthesis in plants. Proc Natl Acad Sci USA 89(19):9354–9358
Hu Q, Sommerfeld M, Jarvis E, Ghirardi M, Posewitz M, Seibert M, Darzins A (2008) Microalgal triacylglycerols as feedstocks for biofuel production: perspectives and advances. Plant J 54(4):621–639
Huang YS, Pereira SL, Leonard AE (2004) Enzymes for transgenic biosynthesis of long-chain polyunsaturated fatty acids. Biochimie 86(11):793–798
Huang X, Huang B, Chen J, Liu X (2016) Cellular responses of the dinoflagellate Prorocentrum donghaiense Lu to phosphate limitation and chronological ageing. J Plankton Res 38(1):83–93
Ikaran Z, Suarez-Alvarez S, Urreta I, Castanon S (2015) The effect of nitrogen limitation on the physiology and metabolism of Chlorella vulgaris var L3. Algal Res 10:134–144
Iskandarov U, Sitnik S, Shtaida N, Didi-Cohen S, Leu S, Khozin-Goldberg I, Cohen Z, Boussiba S (2016) Cloning and characterization of a GPAT-like gene from the microalga Lobosphaera incisa (Trebouxiophyceae): overexpression in Chlamydomonas reinhardtii enhances TAG production. J Appl Phycol 28:907–919
Jain M (2015) Function genomics of abiotic stress tolerance in plants: a CRISPR approach. Front Plant Sci 6:375
Jamers A, Van der Ven K, Moens L, Robbens J, Potters G, Guisez Y, Blust R, De Coen W (2006) Effect of copper exposure on gene expression profiles in Chlamydomonas reinhardtii based on microarray analysis. Aquat Toxicol 80(3):249–260
Jang IC, Oh SJ, Seo JS, Choi WB, Song SI, Kim CH, Kim YS, Seo HS, Do Choi Y, Nahm BH, Kim JK (2003) Expression of a bifunctional fusion of the Escherichia coli genes for trehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase in transgenic rice plants increases trehalose accumulation and abiotic stress tolerance without stunting growth. Plant Physiol 131(2):516–524
Jauzein C, Erdner DL (2013) Stress-related responses in Alexandrium tamarense cells exposed to environmental changes. J Eukaryot Microbiol 60(5):526–538
Jiang T, Zhai H, Wang FB, Zhou HN, Si ZZ, He SZ, Liu QC (2014) Cloning and characterization of a salt tolerance-associated gene encoding trehalose-6-phosphate synthase in sweetpotato. J Integr Agric 13:1651–1661
Jiang SY, Chi YH, Wang JZ, Zhou JX, Cheng YS, Zhang BL, Ma A, Vanitha J, Ramachandran S (2015) Sucrose metabolism gene families and their biological functions. Sci Rep 5:17583
Jiménez C, Capasso JM, Edelstein CL, Rivard CJ, Lucia S, Breusegem S, Berl T, Segovia M (2009) Different ways to die: cell death modes of the unicellular chlorophyte Dunaliella viridis exposed to various environmental stresses are mediated by the caspase-like activity DEVDase. J Exp Bot 60(3):815–828
Johannes F, Colot V, Jansen RC (2008) Epigenome dynamics: a quantitative genetics perspective. Nature Rev Genet 9:883–890
Judkins RR, Fulkerson W, Sanghvi MK (1993) The dilemma of fossil-fuel use and global climate change. Energy Fuels 7(1):14–22
Kaliszewski P, Zoladek T (2008) The role of Rsp5 ubiquitin ligase in regulation of diverse processes in yeast cells. Acta Biochim Pol 55(4):649–662
Kato J, Yamahara T, Tanaka K, Takio S, Satoh T (1997) Characterization of catalase from green algae Chlamydomonas reinhardtii. J Plant Physiol 151(3):262–268
Kebeish R, El-Ayouty Y, Hussein A (2014) Effect of salinity on biochemical traits and photosynthesis-related gene transcription in Chlorella vulgaris Egypt. J Bot 54(2):281–294
Khozin-Goldberg I, Cohen Z (2011) Unraveling algal lipid metabolism: recent advances in gene identification. Biochimie 93(1):91–100
Klahn S, Hagemann M (2011) Compatible solute biosynthesis in cyanobacteria. Environ Microbiol 13(3):551–562
Kobayashi Y, Harada N, Nishimura Y, Saito T, Nakamura M, Fujiwara T, Kuroiwa T, Misumi O (2014) Algae sense exact temperatures: small heat shock proteins are expressed at the survival threshold temperature in Cyanidioschyzon merolae and Chlamydomonas reinhardtii. Genome Biol Evol 6(10):273127–273140
Kronholm I, Bassett A, Baulcombe D, Collins S (2017) Epigenetic and genetic contributions to adaptation in Chlamydomonas. Mol Biol Evol 34(9):2285–2306
Kueger S, Steinhauser D, Willmitzer L, Giavalisco P (2012) High-resolution plant metabolomics: from mass spectral features to metabolites and from whole-cell analysis to subcellular metabolite distributions. Plant J 70(1):39–50
Kumar M, Kumari P, Gupta V, Anisha PA, Reddy CR, Jha B (2010) Differential responses to cadmium induced oxidative stress in marine macroalga Ulva lactuca (Ulvales, Chlorophyta). Biometals 23(2):315–325
Kumar M, Gupta V, Trivedi N, Kumari P, Bijo AJ, Reddy CRK, Jha B (2011) Desiccation induced oxidative stress and its biochemical responses in intertidal red alga Gracilaria corticata (Gracilariales, Rhodophyta). Enviro Exp Bot 72(2):194–201
Kumar M, Bijo AJ, Baghel RS, Reddy CR, Jha B (2012) Selenium and spermine alleviate cadmium induced toxicity in the red seaweed Gracilaria dura by regulating antioxidants and DNA methylation. Plant Physiol Biochem 51:129–138
Kumar M, Kumari P, Reddy C, Jha B (2014) Salinity and desiccation induced oxidative stress acclimation in seaweeds. Adv Bot Res 71:91–123
Kumar M, Kuzhiumparambil U, Pernice M, Jiang ZJ, Ralph PJ (2016) Metabolomics: an emerging frontier of systems biology in marine macrophytes. Algal Res Biomass Biofuels Bioprod 16:76–92
Kwapisz M, Cholbinski P, Hopper AK, Rousset JP, Zoladek T (2005) Rsp5 ubiquitin ligase modulates translation accuracy in yeast Saccharomyces cerevisiae. RNA 11(11):1710–1718
Lander N, Chiurillo MA, Docampo R (2016) Genome Editing by CRISPR/Cas9: a game change in the genetic manipulation of protists. J Eukaryot Microbiol 63(5):679–690
Lee REC, Brunette S, Puente LG, Megeney LA (2010) Metacaspase Yca1 is required for clearance of insoluble protein aggregates. Proc Natl Acad Sci USA 107(30):13348–13353
Lei A, Chen H, Shen G, Hu Z, Chen L, Wang J (2012) Expression of fatty acid synthesis genes and fatty acid accumulation in Haematococcus pluvialis under different stressors. Biotechnol Biofuels 5(1):18
Lemaire SD, Michelet L, Zaffagnini M, Massot V, Issakidis-Bourguet E (2007) Thioredoxins in chloroplasts. Curr Genet 51(6):343–365
Lenka SK, Carbonaro N, Park R, Miller SM, Thorpe I, Li Y (2016) Current advances in molecular, biochemical, and computational modeling analysis of microalgal triacylglycerol biosynthesis. Biotechnol Adv 34(5):1046–1063
Lesser MP (2006) Oxidative stress in marine environments: biochemistry and physiological ecology. Annu Rev Physiol 68:253–278
Li YJ, Fei XW, Deng XD (2012) Novel molecular insights into nitrogen starvation-induced triacylglycerols accumulation revealed by differential gene expression analysis in green algae Micractinium pusillum. Biomass Bioenerg 42:199–211
Li LC, Hsu YT, Chang HL, Wu TM, Sung MS, Cho CL, Lee TM (2013) Polyamine effects on protein disulfide isomerase expression and implications for hypersalinity stress in the marine alga Ulva lactuca Linnaeus. J Phycol 49:1181–1191
Liang X, Zhang L, Natarajan SK, Becker DF (2013) Proline mechanisms of stress survival. Antioxid Redox Signal 19(9):998–1011
Liu X, Wang Y, Gao H, Xu X (2011) Identification and characterization of genes encoding two novel LEA proteins in Antarctic and temperate strains of Chlorella vulgaris. Gene 482(1–2):51–58
Liu J, Sun Z, Zhong Y, Huang J, Hu Q, Chen F (2012) Stearoyl-acyl carrier protein desaturase gene from the oleaginous microalga Chlorella zofingiensis: cloning, characterization and transcriptional analysis. Planta 236(6):1665–1676
Liu Z, Zhang L, Pu Y, Liu Z, Zhao Y, Qin S (2014) Cloning and expression of a cytosolic HSP90 gene in Chlorella vulgaris. BioMed Res Int 14:487050
Liu C, Wang X, Wang X, Sun C (2016) Acclimation of Antarctic Chlamydomonas to the sea-ice environment: a transcriptomic analysis. Extremophiles 20(4):437–450
Los DA, Mironov KS, Allakhverdiev SI (2013) Regulatory role of membrane fluidity in gene expression and physiological functions. Photosynth Res 116(2–3):489–509
Lu Y, Jiang P, Liu S, Gan Q, Cui H, Qin S (2010a) Methyl jasmonate- or gibberellins A3-induced astaxanthin accumulation is associated with up-regulation of transcription of β-carotene ketolase genes (bkts) in microalga Haematococcus pluvialis. Bioresour Technol 101:6468–6474
Lu Y, Chi X, Li Z, Yang Q, Li F, Liu S, Gan Q, Qin S (2010b) Isolation and characterization of a stress-dependent plastidial ∆12 fatty acid desaturase from the Antarctic microalga Chlorella vulgaris NJ-7. Lipids 45(2):179–187
Lud D, Buma AG, Van De Poll W, Moerdijk TC, Huiskes AH (2001) DNA damage and photosynthetic performance in the Antarctic terrestrial alga Prasiola Crispa ssp. Antarctica (chlorophyta) under manipulated UV-B radiation. J Phycol 37(4):459–467
Luis A, Sandalio LM, Corpas FJ, Palma JM, Barroso JB (2006) Reactive oxygen species and reactive nitrogen species in peroxisomes. Production, scavenging, and role in cell signaling. Plant Physiol 141(2):330–335
Lukes M, Prochazkova L, Shmidt V, Nedbalova L, Kaftan D (2014) Temperature dependence of photosynthesis and thylakoid lipid composition in the red snow alga Chlamydomonas cf. nivalis (Chlorophyceae). FEMS Microbiol Ecol 89(2):303–315
Lunn JE, Delorge I, Figueroa CM, Van Dijck P, Stitt M (2014) Treholose metabolism in plants. Plant J 79(4):544–567
Luo QJ, Zhu ZJ, Yang R, Qian FJ, Yan XJ, Chen HM (2015) Characterization of a respiratory burst oxidase homologue from Pyropia haitanensis with unique molecular phylogeny and rapid stress response. J Appl Phycol 27(2):945–955
Lyon BR, Mock T (2014) Polar microalgae: new approaches towards understanding adaptations to an extreme and changing environment. Biology (Basel) 3(1):56–80
Machida T, Murase H, Kato E, Honjoh K, Matsumoto K, Miyamoto T, Iio M (2008) Isolation of cDNAs for hardening-induced genes from Chlorella vulgaris by suppression subtractive hybridization. Plant Sci 175(3):238–246
Maeda H, Sakuragi Y, Bryant DA, Dellapenna D (2005) Tocopherols protect Synechocystis sp. strain PCC 6803 from lipid peroxidation. Plant Physiol 138(3):1422–1435
Maksimov EG, Mironov KS, Trofimova MS, Nechaeva NL, Todorenko DA, Klementiev KE, Tsoraev GV, Tyutyaev EV, Zorina AA, Feduraev PV, Allakhverdiev SI, Paschenko VZ, Los DA (2017) Membrane fluidity controls redox-regulated cold stress responses in cyanobacteria. Photosynth Res 133(1–3):215–223
Malan C, Greyling MM, Gressel J (1990) Correlation between cu-zn superoxide-dismutase and glutathione-reductase, and environmental and xenobiotic stress tolerance in maize inbreds. Plant Sci 69(2):157–166
McGlade C, Ekins P (2015) The geographical distribution of fossil fuels unused when limiting global warming to 2 °C. Nature 517(7533):187–190
Merchant SS et al (2007) The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 318:245–250
Merchant SS, Kropat J, Liu BS, Shaw J, Warakanont J (2012) TAG, You’re it! Chlamydomonas as a reference organism for understanding algal triacylglycerol accumulation. Curr Opin Biotech 23(3):352–363
Mickelbart MV, Hasegawa PM, Bailey-Serres J (2015) Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat Rev Gen 16(4):237
Miranda JA, Avonce N, Suarez R, Thevelein JM, Van Dijck P, Iturriaga G (2007) A bifunctional TPS-TPP enzyme from yeast confers tolerance to multiple and extreme abiotic-stress conditions in transgenic Arabidopsis. Planta 226(6):1411–1421
Mittler R (2017) ROS are good. Trends Plant Sci 22(1):11–19
Mittler R, Vanderauwera S, Gollery M, Van Breusegem F (2004) Reactive oxygen gene network of plants. Trends Plant Sci 9(10):490–498
Moharikar S, D’Souza JS, Kulkarni AB, Rao BJ (2006) Apoptotic-like cell death pathway is induced in unicellular chlorophyte Chlamydomonas reinhardtii (chlorophyceae) cells following UV irradiation: Detection and functional analyses1. J Phycol 42(2):423–433
Moreno-Risueno MA, Busch W, Benfey PN (2010) Omics meet networks - using systems approaches to infer regulatory networks in plants. Curr Opin Plant Biol 13(2):126–131
Müller W, Wegmann K (1978) Sucrose biosynthesis in Dunaliella. Planta 141(2):159–163
Mulo P, Sakurai I, Aro EM (2012) Strategies for psbA gene expression in cyanobacteria, green algae and higher plants: from transcription to PSII repair. Biochim Biophys Acta 1817(1):247–257
Murata N, Los DA (1997) Membrane fluidity and temperature perception. Plant Physiol 115(3):875–879
Murik O, Elboher A, Kaplan A (2014) Dehydroascorbate: a possible surveillance molecule of oxidative stress and programmed cell death in the green alga Chlamydomonas reinhardtii. New Phytol 202(2):471–484
Nagao M, Uemura M (2012) Sucrose phosphate phosphatase in the green alga Klebsormidium flaccidum (Streptophyta) lacks an extensive C-terminal domain and differs from that of land plants. Planta 235(4):851–861
Nagao M, Matsui K, Uemura M (2008) Klebsormidium flaccidum, a charophycean green alga, exhibits cold acclimation that is closely associated with compatible solute accumulation and ultrastructural changes. Plant Cell Environ 31:872–885
Nagaria P, Robert C, Rassool FV (2013) DNA double-strand break response in stem cells: mechanisms to maintain genomic integrity. Biochim Biophys Acta 1830(2):2345–2353
Nakamura Y, Miyachi S (1982) Effect of temperature on starch degradation in Chlorella vulgaris 11 h cells. Plant Cell Physiol 23(2):333–341
Nakashima K, Ito Y, Yamaguchi-Shinozaki K (2009) Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiol 149(1):88–95
Nama S, Madireddi SK, Devadasu ER, Subramanyam R (2015) High light induced changes in organization, protein profile and function of photosynthetic machinery in Chlamydomonas reinhardtii. J Photochem Photobiol B 152(PtB):367–376
Ninomiya Y, Suzuki K, Ishii C, Inoue H (2004) Highly efficient gene replacements in Neurospora strains deficient for nonhomologous end-joining. Proc Natl Acad Sci USA 101(33):12248–12253
Nishiyama Y, Murata N (2014) Revised scheme for the mechanism of photoinhibition and its application to enhance the abiotic stress tolerance of the photosynthetic machinery. Appl Microbiol Biotechnol 98(21):8777–8796
Nishiyama Y, Allakhverdiev SI, Yamamoto H, Hayashi H, Murata N (2004) Singlet oxygen inhibits the repair of photosystem II by suppressing the translation elongation of the D1 protein in Synechocystis sp. PCC 6803. Biochemistry 43(35):11321–11330
Nouri MZ, Moumeni A, Komatsu S (2015) Abiotic stresses: insight into gene regulation and protein expression in photosynthetic pathways of plants. Int J Mol Sci 16(9):20392–20416
Nuñez A, Savary BJ, Foglia TA, Piazza GJ (2000) Anaerobic lipoxygenase activity from Chlorella pyrenoidosa responsible for the cleavage of the 13-hydroperoxides of linoleic and linolenic acids. Eur J Lipid Sci Technol 102:181–188
Nuñez A, Savary BJ, Foglia TA, Piazza GJ (2002) Purification of lipoxygenase from Chlorella: production of 9- and 13-hydroperoxide derivatives of linoleic acid. Lipids 37(11):1027–1032
Okamoto OK, Robertson DL, Fagan TF, Hastings JW, Colepicolo P (2001) Different regulatory mechanisms modulate the expression of a dinoflagellate iron-superoxide dismutase. J Biol Chem 276(23):19989–19993
Oliveira MD, Monteiro M, Robbs P, Leite S (1999) Growth and chemical composition of Spirulina maxima and Spirulina platensis biomass at different temperatures. Aqu Int 7(4):261–275
Olsenz JL (2011) Stress ecology in fucus: abiotic, biotic and genetic interactions. Adv Mar Biol 59(57):37
Park S, Polle JE, Melis A, Lee TK, Jin E (2006) Up-regulation of photoprotection and PSII-repair gene expression by irradiance in the unicellular green alga Dunaliella salina. Mar Biotechnol (NY) 8(2):120–128
Parsell DA, Lindquist S (1993) The function of heat-shock proteins in stress tolerance: degradation and reactivation of damaged proteins. Annu Rev Genet 27(1):437–496
Parvaiz A, Satyawati S (2008) Salt stress and phyto-biochemical responses of plants-a review. Plant Soil Env 54(3):89
Paul VJ (2008) Global warming and cyanobacterial harmful algal blooms. In: Hudnell HK (ed.) Cyanobacterial harmful algal blooms: state of the science and research needs. Springer, Berlin, pp 239–257
Pereira SL, Leonard AE, Mukerji P (2003) Recent advances in the study of fatty acid desaturases from animals and lower eukaryotes. Prostag Leukotr Ess 68(2):97–106
Peters S, Mundree SG, Thomson JA, Farrant JM, Keller F (2007) Protection mechanisms in the resurrection plant Xerophyta viscosa (Baker): both sucrose and raffinose family oligosaccharides (RFOs) accumulate in leaves in response to water deficit. J Exp Bot 58:1947–1956
Petersen JL, Lang DW, Small GD (1999) Cloning and characterization of a class II DNA photolyase from Chlamydomonas. Plant Mol Biol 40(6):1063–1071
Pfannschmidt T (2003) Chloroplast redox signals: how photosynthesis controls its own genes. Trends Plant Sci 8(1):33–41
Pfister B, Zeeman SC (2016) Formation of starch in plant cells. Cell Mol Life Sci 73(14):2781–2807
Piotrowska-Niczyporuk A, Bajguz A, Zambrzycka E, Godlewska-Zylkiewicz B (2012) Phytohormones as regulators of heavy metal biosorption and toxicity in green alga Chlorella vulgaris (Chlorophyceae). Plant Physiol Biochem 52:52–65
Plecenikova A, Mages W, Andresson OS, Hrossova D, Valuchova S, Vlcek D, Slaninova M (2013) Studies on recombination processes in two Chlamydomonas reinhardtii. endogenous genes, NIT1 and ARG7. Protist 164(4):570–582
Plecenikova A, Slaninova M, Riha K (2014) Characterization of DNA repair deficient strains of Chlamydomonas reinhardtii generated by insertional mutagenesis. PLoS One 9(8):e105482
Podkowinski J, Tworak A (2011) Acetyl-coenzyme A carboxylase–an attractive enzyme for biotechnology. BioTechnol 92(4):321–335
Poong SW, Lim PE, Phang SM, Wong CY, Pai TW, Chen CM, Yang CH, Liu CC (2018) Transcriptome sequencing of an Antarctic microalga, Chlorella sp. (Trebouxiophyceae, Chlorophyta) subjected to short-term ultraviolet radiation stress. J Appl Phycol 30(1):87–99
Potin P, Bouarab K, Salaun JP, Pohnert G, Kloareg B (2002) Biotic interactions of marine algae. Curr Opin Plant Biol 5(4):308–317
Qian H, Chen W, Sheng GD, Xu X, Liu W, Fu Z (2008) Effects of glufosinate on antioxidant enzymes, subcellular structure, and gene expression in the unicellular green alga Chlorella vulgaris. Aquat Toxicol 88(4):301–307
Qian H, Chen W, Li J, Wang J, Zhou Z, Liu W, Fu Z (2009a) The effect of exogenous nitric oxide on alleviating herbicide damage in Chlorella vulgaris. Aquat Toxicol 92(4):250–257
Qian H, Chen W, Sun L, Jin Y, Liu W, Fu Z (2009b) Inhibitory effects of paraquat on photosynthesis and the response to oxidative stress in Chlorella vulgaris. Ecotoxicology 18(5):537–543
Qian H, Yu S, Sun Z, Xie X, Liu W, Fu Z (2010) Effects of copper sulfate, hydrogen peroxide and N-phenyl-2-naphthylamine on oxidative stress and the expression of genes involved photosynthesis and microcystin disposition in Microcystis aeruginosa. Aquat Toxicol 99(3):405–412
Qian H, Pan X, Shi S, Yu S, Jiang H, Lin Z, Fu Z (2011) Effect of nonylphenol on response of physiology and photosynthesis-related gene transcription of Chlorella vulgaris. Environ Monit Assess 182(1–4):61–69
Qian H, Pan X, Chen J, Zhou D, Chen Z, Zhang L, Fu Z (2012) Analyses of gene expression and physiological changes in Microcystis aeruginosa reveal the phytotoxicities of three environmental pollutants. Ecotoxicology 21(3):847–859
Raman V, Ravi S (2011) Effect of salicylic acid and methyl jasmonate on antioxidant systems of Haematococcus pluvialis. Acta Physiol Plant 33(3):1043–1049
Ras M, Steyer JP, Bernard O (2013) Temperature effect on microalgae: a crucial factor for outdoor production. Rev Environ Sci Bio 12(2):153–164
Rathinasabapathi B (2000) Metabolic engineering for stress tolerance: Installing osmoprotectant synthesis pathways. Ann Bot-London 86(4):709–716
Rathore RS, Garg N, Garg S, Kumar A (2009) Starch phosphorylase: role in starch metabolism and biotechnological applications. Crit Rev Biotechnol 29(3):214–224
Renaud SM, Thinh LV, Lambrinidis G, Parry DL (2002) Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures. Aquaculture 211(1–4):195–214
Rhodes D, Hanson AD (1993) Quaternary ammonium and tertiary sulfonium compounds in higher plants. Annu Rev Plant Biol 44(1):357–384
Richards AB, Krakowka S, Dexter LB, Schmid H, Wolterbeek APM, Waalkens-Berendsen DH, Shigoyuki A, Kurimoto M (2002) Trehalose: a review of properties, history of use and human tolerance, and results of multiple safety studies. Food Chem Toxicol 40(7):871–898
Richie DL, Miley MD, Bhabhra R, Robson GD, Rhodes JC, Askew DS (2007) The Aspergillus fumigatus metacaspases CasA and CasB facilitate growth under conditions of endoplasmic reticulum stress. Mol Microbiol 63(2):591–604
Richter K, Buchner J (2001) Hsp90: chaperoning signal transduction. J Cell Physiol 188(3):281–289
Rismani-Yazdi H, Haznedaroglu BZ, Bibby K, Peccia J (2011) Transcriptome sequencing and annotation of the microalgae Dunaliella tertiolecta: pathway description and gene discovery for production of next-generation biofuels. BMC Genom 12(1):148
Sabatini SE, Juarez AB, Eppis MR, Bianchi L, Luquet CM, Rios de Molina Mdel C (2009) Oxidative stress and antioxidant defenses in two green microalgae exposed to copper. Ecotoxicol Environ Saf 72(4):1200–1206
Saheb E, Trzyna W, Bush J (2014) Caspase-like proteins: Acanthamoeba castellanii metacaspase and Dictyostelium discoideum paracaspase, what are their functions? J Biosci 39(5):909–916
Sakamoto T, Murata N (2002) Regulation of the desaturation of fatty acids and its role in tolerance to cold and salt stress. Curr Opin Microbiol 5(2):208–210
Sakuradani E, Kobayashi M, Shimizu S (1999) ∆9-Fatty acid desaturase from arachidonic acid-producing fungus. Eur J Biochem 260(1):208–216
Sancar GB (1990) DNA photolyases: physical properties, action mechanism, and roles in dark repair. Mutat Res 236(2–3):147–160
Sayre R (2010) Microalgae: The potential for carbon capture. Bioscience 60(9):722–727
Scholz B, Liebezeit G (2012) Compatible solutes in three marine intertidal microphytobenthic Wadden Sea diatoms exposed to different salinities. Eur J Phycol 47(4):393–407
Schroda M, Vallon O, Wollman FA, Beck CF (1999) A chloroplast-targeted heat shock protein 70 (HSP70) contributes to the photoprotection and repair of photosystem II during and after photoinhibition. Plant Cell 11(6):1165–1178
Schroda M, Hemme D, Muhlhaus T (2015) The Chlamydomonas heat stress response. Plant J 82(3):466–480
Seeberg E, Eide L, Bjoras M (1995) The base excision repair pathway. Trends Biochem Sci 20(10):391–397
Segovia M, Haramaty L, Berges JA, Falkowski PG (2003) Cell death in the unicellular chlorophyte Dunaliella tertiolecta. A hypothesis on the evolution of apoptosis in higher plants and metazoans. Plant Physiol 132(1):99–105
Ševčovičová A, Hamzová A, Gálová E, Vlček D (2008) Use of algae in the study of essential cell processes. Biologia 63(6):952–957
Shahsavarani H, Sugiyama M, Kaneko Y, Chuenchit B, Harashima S (2012) Superior thermotolerance of Saccharomyces cerevisiae for efficient bioethanol fermentation can be achieved by overexpression of RSP5 ubiquitin ligase. Biotech Adv 30(6):1289–1300
Shanklin J, Cahoon EB (1998) Desaturation and related modifications of fatty acids 1. Annu Rev Plant Biol 49(1):611–641
Shanklin J, Somerville C (1991) Stearoyl-Acyl-Carrier-protein desaturase from higher-plants is structurally unrelated to the animal and fungal homologs. Proc Natl Acad Sci USA 88(6):2510–2514
Shapira M, Lers A, Heifetz PB, Irihimovitz V, Osmond CB, Gillham NW, Boynton JE (1997) Differential regulation of chloroplast gene expression in Chlamydomonas reinhardtii during photoacclimation: light stress transiently suppresses synthesis of the Rubisco LSU protein while enhancing synthesis of the PS II D1 protein. Plant Mol Biol 33(6):1001
Shih MD, Hoekstra FA, Hsing YIC (2008) Late embryogenesis abundant proteins. Adv Bot Res 48:211–255
Shin SE, Lim JM, Koh HG, Kim EK, Kang NK, Jeon S, Kwon S, Shin WS, Lee B, Hwangbo K et al (2016) CRISPR/Cas9-induced knockout and knock-in mutations in Chlamydomonas reinhardtii. Sci Rep 6:27810
Singh SC, Sinha RP, Hader DP (2002) Role of lipids and fatty acids in stress tolerance in cyanobacteria. Acta Protozool 41(4):297–308
Singh P, Kumari S, Guldhe A, Misra R, Rawat I, Bux F (2016) Trends and novel strategies for enhancing lipid accumulation and quality in microalgae. Renew Sust Energ Rev 55:1–6
Sinha RP, Hader DP (2002) UV-induced DNA damage and repair: a review. Photochem Photobiol Sci 1(4):225–236
Siripornadulsil S, Traina S, Verma DPS, Sayre RT (2002) Molecular mechanisms of proline-mediated tolerance to toxic heavy metals in transgenic microalgae. Plant Cell 14(11):2837–2847
Smith AM, Zeeman SC, Smith SM (2005) Starch degradation. Annu Rev Plant Biol 56:73–98
Solomon S, Plattner GK, Knutti R, Friedlingstein P (2009) Irreversible climate change due to carbon dioxide emissions. Proc Natl Acad Sci USA 106(6):1704–1709
Spreitzer RJ, Salvucci ME (2002) Rubisco: structure, regulatory interactions, and possibilities for a better enzyme. Annu Rev Plant Biol 53:449–475
Stauber EJ, Hippler M (2004) Chlamydomonas reinhardtii proteomics. Plant Physiol Bioch 42(12):989–1001
Sulmon C, Van Baaren J, Cabello-Hurtado F, Gouesbet G, Hennion F, Mony C, Renault D, Bormans M, El Amrani A, Wiegand C (2015) Abiotic stressors and stress responses: What commonalities appear between species across biological organization levels? Environ Pollut 202:66–77
Takahashi S, Murata N (2008) How do environmental stresses accelerate photoinhibition? Trends in Plant Sci 13(4):178–182
Tanaka S, Ikeda K, Miyasaka H, Shioi Y, Suzuki Y, Tamoi M, Takeda T, Shigeoka S, Harada K, Hirata K (2011) Comparison of three Chlamydomonas strains which show distinctive oxidative stress tolerance. J Biosci Bioeng 112(5):462–468
Teramoto H, Ono T, Minagawa J (2001) Identification of Lhcb gene family encoding the light-harvesting chlorophyll-a/b proteins of photosystem II in Chlamydomonas reinhardtii. Plant Cell Physiol 42(8):849–856
Thitisaksakul M, Jimenez RC, Arias MC, Beckles DM (2012) Effects of environmental factors on cereal starch biosynthesis and composition. J Cereal Sci 56(1):67–80
Thompson GA (1996) Lipids and membrane function in green algae. Biochim Biophys Acta 1302(1):17–45
Tsai DDW, Chen PH, Chou CMJ, Hsu CF, Ramaraj R (2015) Carbon sequestration by alga ecosystems. Ecol Eng 84:386–389
Urzica EI, Vieler A, Hong-Hermesdorf A, Page MD, Casero D, Gallaher SD, Kropat J, Pellegrini M, Benning C, Merchant SS (2013) Remodeling of membrane lipids in iron-starved Chlamydomonas. J Biol Chem 288:30246–30258
Usman A, Brazard J, Martin MM, Plaza P, Heijde M, Zabulon G, Bowler C (2009) Spectroscopic characterization of a (6–4) photolyase from the green alga Ostreococcus tauri. J Photochem Photobiol B 96(1):38–48
Valledor L, Furuhashi T, Hanak AM, Weckwerth W (2013) Systemic cold stress adaptation of Chlamydomonas reinhardtii. Mol Cell Proteom 12(8):2032–2047
Vallentine P, Hung CY, Xie J, Van Hoewyk D (2014) The ubiquitin–proteasome pathway protects Chlamydomonas reinhardtii against selenite toxicity, but is impaired as reactive oxygen species accumulate. AoB Plants 6:plu062
Van Creveld SG, Rosenwasser S, Schatz D, Koren I, Vardi A (2015) Early perturbation in mitochondria redox homeostasis in response to environmental stress predicts cell fate in diatoms. ISME J 9(2):385
Vaux DL, Korsmeyer SJ (1999) Cell death in development. Cell 96(2):245–254
Vavilala SL, Sinha M, Gawde KK, Shirolikar SM, D’Souza JS (2016) KCl induces a caspase-independent programmed cell death in the unicellular green chlorophyte Chlamydomonas reinhardtii (Chlorophyceae). Phycol 55(4):378–392
Vayda ME, Yuan ML (1994) The Heat-Shock response of an Antarctic alga is evident at 5° C. Plant Mol Biol 24(1):229–233
Vega JM, Rubiales MA, Vilchez C, Vigara J (2005) Effect of abiotic stress on photosynthesis, respiration and antioxidant system in Chlamydomonas reinhardtii. Phyton 45(3):97–106
Verbruggen N, Hermans C (2008) Proline accumulation in plants: a review. Amino Acids 35(4):753–759
Vidhyavathi R, Venkatachalam L, Sarada R, Ravishankar GA (2008) Regulation of carotenoid biosynthetic genes expression and carotenoid accumulation in the green alga Haematococcus pluvialis under nutrient stress conditions. J Exp Bot 59(6):1409–1418
Vinocur B, Altman A (2005) Recent advances in engineering plant tolerance to abiotic stress: achievements and limitations. Curr Opin Biotechnol 16(2):123–132
Vitova M, Bisova K, Kawano S, Zachleder V (2015) Accumulation of energy reserves in algae: from cell cycles to biotechnological applications. Biotechnol Adv 33(6):1204–1218
Vlcek D, Sevcovicova A, Sviezena B, Galova E, Miadokova E (2008) Chlamydomonas reinhardtii: a convenient model system for the study of DNA repair in photoautotrophic eukaryotes. Curr Gen 53(1):1–22
Von Kampen J, Nieländer U, Wettern M (1995) Expression of ubiquitin genes in Chlamydomonas reinhardtii: involvement in stress response and cell cycle. Planta 197(3):528–534
Wada H, Gombos Z, Murata N (1994) Contribution of membrane lipids to the ability of the photosynthetic machinery to tolerate temperature stress. Proc Natl Acad Sci USA 91(10):4273–4277
Wan M, Jin X, Xia J, Rosenberg JN, Yu G, Nie Z, Oyler GA, Betenbaugh MJ (2014) The effect of iron on growth, lipid accumulation, and gene expression profile of the freshwater microalga Chlorella sorokiniana. Appl Microbiol Biotechnol 98(22):9473–9481
Wang JX, Zhang XZ, Chen YS, Sommerfeld M, Hu Q (2008) Toxicity assessment of manufactured nanomaterials using the unicellular green alga Chlamydomonas reinhardtii. Chemosphere 73(7):1121–1128
Wasternack C, Hause B (2013) Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in annals of botany. Ann Bot 111(6):1021–1058
Waters ER, Lee GJ, Vierling E (1996) Evolution, structure and function of the small heat shock proteins in plants. J Exp Bot 47(3):325–338
Wegmann K (1986) Osmoregulation in eukaryotic algae. FEMS Microbiol Lett 39(1–2):37–43
Wendel AA, Lewin TM, Coleman RA (2008) Glycerol-3-phosphate acyltransferases: rate limiting enzymes of triacylglycerol biosynthesis. Biochim Biophys Acta 1791(6):501–506
Wettern M, Parag HA, Pollmann L, Ohad I, Kulka RG (1990) Ubiquitin in Chlamydomonas reinhardii. Distribution in the cell and effect of heat shock and photoinhibition on its conjugate pattern. Eur J Biochem 191(3):571–576
Wrona FJ, Prowse TD, Reist JD, Hobbie JE, Levesque LM, Vincent WF (2006) Climate change effects on aquatic biota, ecosystem structure and function. Ambio 35(7):359–369
Xu J, Zhang X, Ye N, Zheng Z, Mou S, Dong M, Xu D, Miao J (2013) Activities of principal photosynthetic enzymes in green macroalga Ulva linza: functional implication of C4 pathway in CO2 assimilation. Sci China Life Sci 56(6):571–580
Xu T, Li Y, Van Nostrand JD, He Z, Zhou J (2014) Cas9-based tools for targeted genome editing and transcriptional control. Appl Environ Microbiol 80(5):1544–1552
Xue WB, Liu F, Sun Z, Zhou ZG (2016) A delta-9 fatty acid desaturase gene in the microalga Myrmecia incisa reisigl: cloning and functional analysis. Int J Mol Sci 17(7):1143
Yamaoka Y, Achard D, Jang S, Legéret B, Kamisuki S, Ko D, Schulz-Raffelt M, Kim Y, Song WY, Nishida I, Li-Beisson Y, Lee Y (2016) Identification of a Chlamydomonas plastidial 2-lysophosphatidic acid acyltransferase and its use to engineer microalgae with increased oil content. Plant Biotech J 14(11):2158–2167
Yancey PH, Clark ME, Hand SC, Bowlus RD, Somero GN (1982) Living with water stress: evolution of osmolyte systems. Science 217(4566):1214–1222
Yanguez K, Lovazzano C, Contreras-Porcia L, Ehrenfeld N (2015) Response to oxidative stress induced by high light and carbon dioxide (CO2) in the biodiesel producer model Nannochloropsis salina (Ochrophyta, Eustigmatales). Rev Biol Mar Oceanog 50:163–175
Ye ZW, Jiang JG, Wu GH (2008) Biosynthesis and regulation of carotenoids in Dunaliella: progresses and prospects. Biotechnol Adv 26(4):352–360
Yildiztugay E, Ozfidan-Konakci C, Kucukoduk M (2014) The role of antioxidant responses on the tolerance range of extreme halophyte Salsola crassa grown under toxic salt concentrations. Ecotoxicol Environ Saf 110:21–30
Yokthongwattana K, Chrost B, Behrman S, Casper-Lindley C, Melis A (2001) Photosystem II damage and repair cycle in the green alga Dunaliella salina: involvement of a chloroplast-localized HSP70. Plant Cell Physiol 42(12):1389–1397
Yoon K, Han D, Li Y, Sommerfeld M, Hu Q (2012) Phospholipid:diacylglycerol acyltransferase is a multifunctional enzyme involved in membrane lipid turnover and degradation while synthesizing triacylglycerol in the unicellular green microalga Chlamydomonas reinhardtii. Plant Cell 24:3708–3724
Young AJ (1991) The photoprotective role of carotenoids in higher-plants. Physiol Plant 83(4):702–708
Young JC, Moarefi I, Hartl FU (2001) Hsp90: a specialized but essential protein-folding tool. J Cell Biol 154(2):267–273
Zhang LP, Mehta SK, Liu ZP, Yang ZM (2008) Copper-induced proline synthesis is associated with nitric oxide generation in Chlamydomonas reinhardtii. Plant Cell Physiol 49(3):411–419
Zhang N, Wang F, Meng X, Luo S, Li Q, Dong H, Xu Z, Song R (2011) Molecular cloning and characterization of a trehalose-6-phosphate synthase/phosphatase from Dunaliella viridis. Mol Biol Rep 38(4):2241–2248
Zhang H, Li W, Li J, Fu W, Yao J, Duan D (2012) Characterization and expression analysis of hsp70 gene from Ulva prolifera J. Agardh (Chlorophycophyta, Chlorophyceae). Mar Genom 5:53–58
Zhu S, Huang W, Xu J, Wang Z, Xu J, Yuan Z (2014a) Metabolic changes of starch and lipid triggered by nitrogen starvation in the microalga Chlorella zofingiensis. Bioresour Technol 152:292–298
Zhu S, Wang Y, Huang W, Xu J, Wang Z, Xu J, Yuan Z (2014b) Enhanced accumulation of carbohydrate and starch in Chlorella zofingiensis induced by nitrogen starvation. Appl Biochem Biotechnol 174(7):2435–2445
Zimmerman D, Vick B (1973) Lipoxygenase in Chlorella pyrenoidosa. Lipids 8(5):264–266
Zuo Z, Zhu Y, Bai Y, Wang Y (2012) Acetic acid-induced programmed cell death and release of volatile organic compounds in Chlamydomonas reinhardtii. Plant Physiol Biochem 51:175–184
Zuppini A, Andreoli C, Baldan B (2007) Heat stress: an inducer of programmed cell death in Chlorella saccharophila. Plant Cell Physiol 48(7):1000–1009
Acknowledgements
The study was sponsored by the research Grants from the Ministry of Higher Education, Malaysia, HiCOE research Grant (IOES-2014H), University of Malaya Postgraduate Research Fund (PG146-2015A) and University of Malaya Research Grant (RP002C-13SUS).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M. Horbowicz.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Barati, B., Gan, SY., Lim, PE. et al. Green algal molecular responses to temperature stress. Acta Physiol Plant 41, 26 (2019). https://doi.org/10.1007/s11738-019-2813-1
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-019-2813-1