
Abstract
Purpose
The identification of the external attaching fish parasitic cymothoid, Nerocila phaiopleura Bleeker 1857, is still based on the brief description of Australian specimens provided by Bruce (1987). The present study aimed to provide a redescription and molecular characterisation of Indian specimens of N. phaiopleura.
Materials and Methods
Morphological identification was carried out based on microscopic examinations and taxonomic drawings. mitochondrial DNA cox1 was selected as the target gene for sequencing and molecular identification. Nucleotide genetic divergence (p-distance) and base-pair differences among the different species were determined using MEGA11.
Results
Nerocila phaiopleura can be well separated from its congeners by the following combination of characteristics: Body about 2.4 times as long as wide, cephalon broadly rounded anteriorly; coxae posteriorly directed, acute and extending beyond their corresponding pereonite; pereonite 7 posterior angle produced, extending to the pleonite 1; pleonites 1 and 2 ventrolateral process posteriorly directed; uropod exopod straight and elongate, 1.7–2.0 times longer than endopod; uropod endopod lateral margin not serrate, no notch on medial margin; pereopods with short ischium; pleotelson triangular. The p-distance among N. phaiopleura and other available Nerocila spp. ranged from 21 to 19%.
Conclusion
This study represents the first detailed taxonomic redescription of Indian specimens of N. phaiopleura. Key taxonomic features of the life stages and molecular data are provided here to identify the species properly. Interspecific genetic divergence between N. phaiopleura and other Nerocila spp. is assessed for the first time. Studies in cymothoid life histories, genetics, and morphology are necessary to understand one of the least understood parasite families.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The external attaching fish parasitic cymothoid genus Nerocila Leach, 1818 comprises 57 valid species [1] and is associated with many commercially important fish species worldwide [2]. Several species of Nerocila are morphologically highly variable, and their identification is often difficult [3, 4]. Notwithstanding the few recent reports, many species of Nerocila still require revision, including Nerocila phaiopleura Bleeker, 1857. Twelve valid species of Nerocila (N. loveni Bovallius, 1887, N. exocoeti Pillai, 1954, N. trichiura Miers, 1878, N. serra Schioedte and Meinert, 1881, N. depressa H. Milne Edwards, 1840, N. poruvae Rameshkumar, Ravichandran and Trilles, 2011, N. recurvispina Schioedte and Meinert, 1881, N. longispina Miers, 1880, N. arres Bowman and Tareen, 1983, N. phaiopleura Bleeker, 1857, N. sigani Bowman and Tareen, 1983 and N. sundaica Bleeker, 1857) have been known from Indian waters [5,6,7].
Nerocila phaiopleura is a widely distributed and variable species found as an external parasite attaching to the skin of nearly 50 species of fishes belonging to 18 families, including Clupidae, Engraulidae, Carangidae, Scombridae, Dussumieriidae, Chirocentridae, Pristigasteridae, Mugilidae, Sphyraenidae, Leiognathidae, Plotosidae, Polynemidae, Ariidae and Istiophoridae from the Indian and Pacific oceans [8,9,10,11,12]. The species has been recorded from about 20 different fish species from India [10, 13].
Bleeker [14] provided the original description of N. phaiopleura. Pillai [8] gave brief descriptions of the species with illustrations of only a dorsal view of the species. Subsequently, Bruce [3] provided a short description of Australian specimens of N. phaiopleura, and after this, no detailed taxonomic information on the species was available. The previous reports of this species were brief and required further clarification on the identity. There is no previous taxonomic description for the moulting stage of N. phaiopleura.
The taxonomic status of many parasitic isopod species is still confusing, emphasizing the significance of detailed descriptions with accurate illustrations. The identification of N. phaiopleura is still based on the descriptions mentioned above. A detailed description based on Indian specimens is highly needed; since the Indian specimen is not adequately described yet. The present study provides molecular characterization and a detailed morphological description of the female and moulting stages of the parasitic isopod, N. phaiopleura, collected from the two host fishes Dussumieria acuta Valenciennes, 1847 and Rastrelliger kanagurta Cuvier, 1816 from the Kerala coast, India.
Materials and Methods
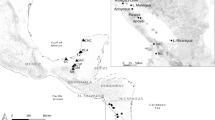
The specimens of Nerocila phaiopleura Bleeker 1857 were collected from the host fish (Dussumieria acuta Valenciennes, 1847 and Rastrelliger kanagurta Cuvier, 1816) in Neendakara fishing harbour (Lat. 8°56′19" N, Long. 76°32′25" E), Southwest coast of India. The mode of attachment of isopods to the host skin and the gross changes they made were observed. The parasites isolated were cleaned (removing mucus and other debris adhering to their bodies) and preserved in 70% alcohol/10% Neutral Buffered formalin for taxonomic studies. A few samples were preserved in 100% ethanol for genetic analysis.
Morphological Identification
For morphological identification, the parasite's general morphological features were studied using a hand lens and under a Stereo Dissection Microscope (SDM) (Carl Zeiss Microscopy; GmbH Stemi 508). Later, the parasites were dissected in 50% lactic acid; taxonomically important body parts such as pereopods, pleopods, uropods and mouthparts were carefully dissected according to the techniques described by Aneesh et al. [6, 15,16,17]. The parasites' total length and maximum width was measured to the nearest millimetre using a Vernier calliper. The species identification followed Bruce [3, 4]. Sources for the fish taxonomy and host nomenclature were based on Fish Base [18] and Catalogue of Fishes [19].
The mode of attachment of isopods to the host skin and the gross changes made by the parasite were photographed using a Canon EOS 800D with a 35 mm macro lens. Microphotographs of the parasite and its body parts were taken using SDM. The drawings of the body parts of the parasite were made with a drawing tube attached to the Transmission Light Microscope (TLM) (Optika Microscope; Optikam B5 Digital Camera). Taxonomic drawings were made using CorelDraw software (CorelDRAW X7). The voucher specimens are deposited in the Western Ghat Field Research Centre of Zoological Survey of India, Kozhikode (ZSI/WGRC).
Molecular Analysis
Genomic DNA was isolated from the tissues following the protocol for animal tissue extraction using NucleoSpin® Tissue Kit (Macherey–Nagel, Düren, Germany) following the manufacturer's instructions. A targeted part of the mitochondrial cytochrome c oxidase subunit I (COI) gene (approximately 680 bp) of these specimens was subjected to PCR amplification with the aid of a PCR thermal cycler (GeneAmp PCR System 9700, Applied Biosystems) and universal invertebrate primers LCO1490 (5′-GGTCAACAAATCATAAAGATATTGG-3′) and HC02198 (5′-TAAACTTCAGGGTGACCAAAAAATCA-3′).
PCR reactions were performed with volumes of 10.5 μl, using 5 μl 2 × Phire Master Mix, 0.25 μl of each primer, 4 μl of PCR–grade nuclease-free water and 1 μl of DNA. Conditions for the PCR were as follows: initial denaturation at 98 °C for 30 s, followed by ten cycles of 98°C for 5 s, annealing at 45 °C for 10 s with an end extension at 72 °C for 15 s, followed by 30 cycles of 98°C for 5 s, annealing at 50 °C for 10 s with an end extension at 72 °C for 15 s, and ending with a final extension of 72 °C for 60 s.
The sequencing reaction was done in a PCR thermal cycler (GeneAmp PCR System 9700, Applied Biosystems) using the BigDye Terminator v3.1 Cycle sequencing Kit (Applied Biosystems, USA) following the manufacturer's protocol. The cleaned-up air-dried product was sequenced in ABI 3500 DNA Analyzer (Applied Biosystems). The sequence quality was checked using Sequence Scanner Software v1 (Applied Biosystems). Sequence alignment and required editing of the obtained sequences were carried out using Geneious Pro v5.1 [20].
Comparative sequences of Nerocila species from GenBank were downloaded and aligned to one sequence. These sequences included: OP890359 (N. phaiopleura), KY933655 (N. loveni), ON661340 (N. exocoeti), OK001962 (N. longispina), LC160331 (N. japonica) and MZ644982 (N. orbignyi). Nucleotide genetic divergence (p-distance) and base-pair differences among the different species were determined using MEGA11 [21]. Maximum Composite Likelihood method is used for p-distance calculation.
Results
Taxonomy
Suborder Cymothoida Wägele, 1989
Superfamily Cymothooidea Leach, 1814
Family Cymothoidae Leach, 1814
Genus Nerocila Leach, 1818
Nerocila phaiopleura Bleeker, 1857
(Figs. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10).

Nerocila phaiopleura Bleeker 1857 from Rastrelliger kanagurta Cuvier (a-c) Ovigerous female dorsal, ventral, and lateral view, respectively (d, e) non-ovigerous female dorsal and ventral view, respectively

Nerocila phaiopleura Bleeker 1857 from Rastrelliger kanagurta Cuvier (a–c) Ovigerous female dorsal, ventral, and lateral view, respectively (d and e) Non-ovigerous female, dorsal and ventral view respectively (f and g) Ovigerous female cephalon, dorsal and ventral view, respectively

Nerocila phaiopleura Bleeker 1857 from Rastrelliger kanagurta Cuvier, female (a–m) Ovigerous female mouth parts (a) Antennula (b) Antenna (c) Antenna apex (d) Antennule apex (e) Mandible (f and g) Mandibular palp variations (h) Maxillule (i) Maxillule apex (j) Maxilla (k) Maxilla apex (l) Maxilliped apex (m) Maxilliped (n) Non-ovigerous female, maxilliped

Nerocila phaiopleura Bleeker 1857 from Rastrelliger kanagurta Cuvier, ovigerous female (a–g) Pereopod 1–7, respectively (h) Oostegite

Nerocila phaiopleura Bleeker 1857 from Rastrelliger kanagurta Cuvier, ovigerous female (a–e) Pleopod 1–5, respectively

Nerocila phaiopleura Bleeker 1857 from Rastrelliger kanagurta Cuvier, partially moulted male, (a and b) Dorsal and ventral view, respectively

Nerocila phaiopleura Bleeker 1857 from Rastrelliger kanagurta Cuvier, male moulting stage, (a and b) Dorsal and ventral view respectively (c) Cephalon (d) Antennula (e) Antenna (f) Mandible (g) Mandibular palp variation (h) Maxillule (i) Maxilla (j) Maxilliped apex (k) Maxilliped

Nerocila phaiopleura Bleeker 1857 from Rastrelliger kanagurta Cuvier, male moulting stage, (a–g) Pereopods 1–7, respectively (h) Pleotelson (i) Uropod

Nerocila phaiopleura Bleeker 1857 from Rastrelliger kanagurta Cuvier, male moulting stage (a–e) Pleopod 1–5 respectively

Nerocila phaiopleura Bleeker 1857 infestation on host, Rastrelliger kanagurta Cuvier (a) Heavy infestation of the parasite on the host (b) Parasite attachment and feeding (c and d) Small pin holes with the formation of epidermal plaques at the attachment site of parasite (red arrows) (e) Large haemorrhagic wound made by the parasite due to the attachment and feeding activities
Nerocila phaiopleura Bleeker, 1857: 25, pl. 1 (Fig. 3)—Monod, 1976: 857—Trilles, 1979: 253, pl. 1 (Fig. 3).
Nerocila phaeopleura—Miers, 1880: 467;—Schiöedte & Meinert, 1881: 13, pl. 1 (Figs. 6, 7)—Gerstaecker, 1882: 260.—Nierstrasz, 1915: 75, pl. 3 (Figs. 1, 2); 1918: 113, pl. 9 (Figs. 6, 7); 1931: 124—Barnard, 1925: 392.—Chilton, 1926: 180, Fig. 3a–b.—Monod, 1934: 12.—Serene, 1937: 69—Morton, 1974: 143, pl. 1—Kensley, 1978: 82, Fig. 33d, g
Nerocila (Nerocila) phaeopleura—Bruce, 1982: 316, Figs. 1, 4a–c
Nerocila (Nerocila) phaiopleura Bowman & Tareen, 1983: 5, Fig. 5
Nerocila phaeopleura [sic]—Barnard, 1936: 164, Fig. 6a–c
Nerocila spp. Monod, 1976: 857, Figs. 14, 15.—Meenakshisundaram, 1965: 202–204, Figs 1, 2.—Seshagiri Rao, 1974: 428— Ranjitsingh & Padmalatha, 1997: 171, Figs. 1, 2.—Radhakrishnan & Nair, 1983: 93–115, Fig. 10d
Nerocila phaiopleura—Bruce, 1987b: 384, Figs 18, 19.—Bruce & Harrison-Nelson, 1988: 596.—Ravichandran, Ranjith Singh & Veerappan, 2001: 622–623, Figs. 1, 2.—Kazmi, Schotte & Yousuf, 2002: 103, Fig. 84.—Trilles, Ravichandran & Rameshkumar, 2011: 452.—Trilles, Rameshkumar & Ravichandran, 2013: 1273–1286, Fig. 2f.—Bharadhirajan, Murugan, Sakthivel & Selvakumar, 2014: 268–272, Fig. 2a–d.—Rameshkumar, Ramesh, Ravichandran & Trilles, 2014c: 940–944, Fig. 1g.—Rameshkumar, Ravichandran & Ramesh, 2014: 124–128, Fig. 5.—Ravichandran, Sivasubramanian, Parasuraman, Karthick Rajan & Rameshkumar, 2016a: 1–5, Figs 1, 3.—Ravichandran, Vigneshwaran & Rameshkumar, 2019: 58, Fig. 8j–l.
Material examined: Voucher specimens: All materials from Rastrelliger kanagurta (Scombridae), off Neendakara coast, 08°30.0’N, 76°53.30’E, coll. Amrutha SS on December, 2021. 1 ovigerous female (22.5 mm L; 9.5 mm W) (Reg. No. ZSI/WGRC/I.R-INV 27087); 1 ovigerous female (22 mm L; 9.5 mm W) (Reg. No. ZSI/WGRC/I.R-INV. 27088); 1 ovigerous female (20 mm L; 8.5 mm W) (Reg. No. ZSI/WGRC/I.R-INV. 27089); 1 ovigerous female (20 mm L; 9 mm W) (Reg. No ZSI/WGRC/I.R-INV. 27090); 1 ovigerous female (21 mm L; 9.5 mm W) (Reg. No. ZSI/WGRC/I.R-INV. 27091); 1 non-ovigerous female (19 mm L; 8 mm W) (Reg. No. ZSI/WGRC/I.R-INV. 27092); 1 non-ovigerous female (20.5 mm L; 8 mm W) (Reg. No.); 1 non-ovigerous female (17 mm L; 6.5 mm W) (Reg. No. ZSI/WGRC/I.R-INV. 27093); 1 transitional stage (male moulting stage) (15.5 mm L; 5.5 mm W) (Reg. No. ZSI/WGRC/I.R-INV. 27094); 1 transitional stage (male moulting stage) (16.5 mm L; 7 mm W) (Reg. No ZSI/WGRC/I.R-INV. 27096).
Redescription of ovigerous female (Figs. 1, 2, 3, 4, 5): Body ovate, about 2.4 times as long as wide, widest at pereonite 5–6. Cephalon broadly rounded anteriorly, 1.6 times wider than long, posterior margin prominently trisinuate. Eyes large, with brownish-black colour, not distinct, 0.1 times as wide as cephalon. Coxae 1–7 visible in dorsal view, extending beyond their corresponding pereonites, gradually increasing the length and width, produced posteriorly, apex acute. Pereonite 1 shortest, anterolateral corners slightly produced; pereonites 1–5 gradually increase in width; pereonites 5 widest, 5 and 6 subequal; pereonite 7 shorter than 6, postero-lateral corners strongly produced, acute. Pleon narrower than pereon, all pleonites visible in dorsal view, lateral margins darkly pigmented, not overlapped by pereonite 7; pleonites 1 and 2 with lateral processes; pleonite 1 longest and widest, lateral margin strongly produced posteriorly; pleonites 2–5 sub equal, lateral margins posteriorly directed. Pleotelson triangular, converging smoothly to a caudomedial point, 1.2 times wider than long.
Antennula distinctly stouter than the antenna, 8-articled, articles 4–8 setose, articles 2 and 3 long, articles 2–8 decreasing in width, extending beyond the posterior of eye. Antenna slender, decreasing gradually in width, 11-articled, article 5 longest, 5–11 with distal setae, antenna extending to the posterior of the pereonite 1. Mandible palp slender, proximal segment longest and widest, lateral margin with few robust setae (RS); mandibular incisor blunt, well developed. Maxillula with 2 small and 2 large recurved apical RS. Maxilla medial lobe with 1, lateral lobe with 2 hooked robust setae in ovigerous female. In non-ovigerous females, 2 hooked robust setae on both medial and lateral lobes. Maxilliped oostegial lobe with many plumose setae; palp with 2 apical and one lateral small recurved robust seta on article 3. In non-ovigerous females, maxilliped palp with 2 apical and 2 lateral recurved robust setae on article 3.
Pereopods 1–7 with weak lobes on antereo-proximal margin of dactylus, gradually increase in length, ischium short and stout; pereopod 1 short; pereopods 1–6 without marginal robust setae, dactylus falcate; pereopod 7 long, dactylus extend to posterior of the carpus, carpus and merus stouter than the other pereopods, 4 robust setae on the margin of propodus, 2 robust setae on the carpus and merus, a single robust seta on distal inferior margins of the ischium.
Pleopods not distinctly visible in dorsal view; pleopods 1–2, endopod without lobes, pleopods 3–5 with proximo-medial lobe; Pleopods 3 and 4 endopod with a small single fold; pleopod 5 endopod with large multiple folds. Uropod slender; exopod straight, tapering, darkly pigmented, elongate about 8 times longer than wide, 1.7–2.0 times longer than endopod; endopod apex narrowly rounded or obliquely truncate.
Description of partially moulted male (Figs. 6, 7, 8, 9): Body 2.2 times as long as wide, widest at pereonite 6. Cephalon broadly rounded anteriorly, 1.3 times as wide as long, posterior margin trisinuate. Eyes large, distinctly visible, 0.2 times as wide as cephalon. Coxae 2–4 not visible in dorsal view; coxae 5–7 visible dorsally, long, produced posteriorly slightly beyond their pereonites, apex acute. Pereonites 1–4 narrower and slightly increase in width from 1–4 pereonites, pereonite 1 shortest; pereonites 5–7 abruptly broader, pereonite 6 widest and 2.5 times wider than pereonite 1, pereonite 5 slightly smaller than pereonite 6, pereonite 7 slightly smaller than pereonite 5. Pleon 1.2–1.5 times broader than first 4 pereonites and 1.0–1.3 times narrower than posterior 3 pereonites. All pleonites visible in dorsal view, not immersed in pereonite 7; pleonites 1 widest; pleonites 2–5 subequal; the ventral process of pleonites 1 and 2 broad. Pleotelson as long as wide, triangular, converging smoothly to a caudomedial point.
Antennula distinctly stouter than the antenna, 8–articled; a cluster of setae on 4–8 articles; article 3 longest; article decreasing in width. Antenna slender, decreasing gradually in width; 11–articled, 5th article longest, 3, 4, 7 and 11 articles with distal setae. Mandibular palp article 3 lateral margin with about 11–14 stout setae, article 2 with 2 setae. Maxillula with 3 apical robust setae. Maxilla, medial lobe with 2 robust setae, lateral lobe with 1 robust seta. Maxilliped article 3, with 2 apical and 1 lateral small robust seta.
Pereopods 1–7 prehensile with weak swelling on antereo-proximal margin of dactylus, gradually increase in length, ischium short and stout, carpus immersed in merus; pereopod 1 short, dactylus reaching up to posterior of the merus; pereopods 1–6 without marginal robust setae, dactylus falcate; pereopod 7 long, dactylus extend to posterior of the carpus, carpus and merus stouter than the other pereopods, 4 robust setae on the margin of propodus, 2 robust setae on the carpus and merus, a single robust seta on distal inferior margins of the ischium.
Pleopods not distinctly visible in dorsal view; pleopods 1 and 2, endopod without lobes, pleopods 3–5 with proximo–medial lobe; pleopod 2 with appendix masculina about half the length of endopod; pleopods 3 and 4 endopod with a single small fold; pleopod 5 endopod with large multiple folds. Uropodal rami slender, long; exopod 1.3–1.4 times longer than endopod, overreaching telson.
Colour. Pale tan with chromatophores along posterior of pereon and pleon segments and lateral margin of uropod peduncle.
Size. Ovig. females: 17–26 mm, non ovig. females: 19–25 mm, moulting stage: 15–18 mm.
Host and distribution: list of all the recorded host fishes of Nerocila phaiopleura and its global distribution is provided in Table 1. The present materials are collected from Dussumieria acuta Valenciennes, 1847 and Rastrelliger kanagurta Cuvier, 1816, off south-west coast of India.
Ecological remarks: The cymothoid, N. phaiopleura, was found attached firmly to the posterior third of the body, overlying the lateral line and facing the head of the host fish (Fig. 10a, b). Discrete alterations such as haemorrhages, loss of scales, and extensive skin erosions/skin ulceration were gross pathological symptoms of the attachment site of the parasite. On the body surface, small pinholes with the formation of epidermal plaques (Fig. 10c, d) and large, round-shaped haemorrhagic wounds/ulcers (no fish skin was present on the wounds where the muscle was exposed) (Fig. 10e) were noticed.
Molecular analysis: 100% similar COI sequences for N. phaiopleura were generated when submitted to GenBank with GenBank accession number OP890359. The COI sequence was compared to other known Nerocila spp. sequences available on GenBank (Table 2). The alignment was 647 bp, codon positions included were 1st + 2nd + 3rd, all positions containing gaps and missing data were eliminated. Nucleotide genetic divergence (p-distance) among N. phaiopleura and other available Nerocila spp. ranged from 21 to 29%. The P genetic distance was high (29%) between N. phaiopleura and N. longispina. The genetic distance was less (21%) between N. phaiopleura and N. poruvae.
Discussion
Nerocila phaiopleura Bleeker, 1857, is redescribed here in detail based on the specimens collected from the Southwest coast of India. The results revealed that the cymothoid, N. phaiopleura, could be identified by the following combinations of characters: body about 2.4 times as long as wide; cephalon broadly rounded anteriorly; coxae posteriorly directed, acute and extending beyond their corresponding pereonites; pereonites 7 posterior angle produced, extending to the pleonite 1; pleonites 1 and 2 ventrolateral process posteriorly directed; uropod exopod straight and elongate about 8–9 times longer than proximal width, 1.7–2.0 times longer than endopod; uropod endopod lateral margin not serrate; no notch on medial margin; pleotelson triangular and the distinctive pereopod morphology (e.g. pereopods with a short ischium). Nerocila phaiopleura differs from the closely related species N. depressa by the coxae, and posterolateral comers of the pleonites are posteriorly directed and are not bent dorsally in N. phaiopleura (vs the coxae and posterolateral comers of the pleonites are slightly bent and directed dorsally in N. depressa); the uropod endopod with an obliquely truncate apex in N. phaiopleura (refer Figs. 1, 2, 3, 4, 5).
The marked difference between the male moulting stage and the female is that the body shape of the male moulting stage is not oval; instead, the body is abruptly narrower anteriorly (anterior-most pereonites 1–4 are abruptly narrower than the remaining pereonites). The variations shown by the male moulting stage from the female are: pereonite 5, 2 times wider than pereonite 4 (in females, pereonite 5, 1.1 times wider than pereonite 4); pleon about 1.4 times broader than first 4 pereonites and about 1.2 times narrower than posterior 3 pereonites (in females, pleon narrower than pereon); mandibular palp article 3 lateral margin with more (about 11–14) stout setae; robust setae on pereopods 1–4 and 7 (in females, robust setae present on pereopod 7 only) (refer Figs. 1, 2, 3, 4, 5, 6, 7, 8, 9).
The previous description of N. phaiopleura are short and do not consider most of the taxonomic details. The male-moulting stage of N. phaiopleura is not characterized yet. The general morphology and appendages of life cycle stages (transitional (partially moulted) and ovigerous female) of Indian specimens are described and illustrated here. Key taxonomic features provided for the proper identification of N. phaiopleura.
Variations in the morphology of the body parts were observed in the present Indian specimens of N. phaiopleura compared to Australian samples described by Bruce [3]. The mandible palp article lateral margin has about 22 stout setae in the descriptions provided by Bruce [3], whereas the present specimens have mandible palp article lateral margin without setae or with a few setae. Pereopod 7 has 2 spines on the posterior margin of the propodus, according to Bruce [3], whereas in the present study, pereopod 7 has 4 spines on the margin of the propodus, 2 spines on the carpus and a single spine on the distal inferior margins of the ischium. Illustrations and descriptions of the other morphological characters such as maxilla, maxillula, 1st and 2nd antenna, pereopod 2–6, and pleopods of ovigerous females were not included in the descriptions provided by Bruce [3].
The comparison of genetic distances among N. phaiopleura and the available Nerocila spp. on GenBank was provided based on COI sequences (refer to Table 2). The genes (COI sequences) of Nerocila spp. can provide more information on genetic distances among species, which can be linked with their morphological aspects. Integrative taxonomy for Nerocila can aid a better understanding of the species, as in the case of the present study, drawing more conclusions on the actual diversity and distribution of the genus.
The isopod parasite, N. phaiopleura, causes discrete alterations such as haemorrhages, loss of scales, and extensive skin erosions/ ulceration at the attachment site. On the body surface, small pinholes with the formation of epidermal plaques and large, round-shaped haemorrhagic wounds/ulcers were observed as gross pathological symptoms (refer Fig. 10). Amrutha et al. [22] observed that, at the insertion site of the parasite’s pereopods, there was deep skin depression along with massive lymphocyte infiltrations. The epidermis around the pereopod attachment site was hyperplasic, clearly manifesting epidermal spongiosis. The host response includes infiltration of inflammatory cells, primarily macrophages, lymphocytes and eosinophils. Muscle fibrosis and degeneration, along with oedema and tissues undergoing necrosis, were evident in the muscle tissues. The significant histopathological changes in the fish tissues caused by N. phaiopleura are probably due to attachment, movement and feeding. Furthermore, localized loss of osmoregulatory skin function may occur at the lesions. Finally, the isopods may mechanically impede swimming performance, making infested fish more susceptible to predation and indirectly causing increased mortality in wild fish populations.
Nerocila phaiopleura has a wide geographical distribution and host range. It is reported from 50 species of fish belonging to 14 families, including Clupidae, Engraulidae, Carangidae, Scombridae, Dussumieriidae, Chirocentridae, Pristigasteridae, Mugilidae, Sphyraenidae, Leiognathidae, Plotosidae, Polynemidae, Ariidae and Istiophoridae from the Indian Ocean and Pacific Ocean [8, 9, 12, 23]. This clearly shows that this species is the least host-specific and can potentially threaten the wild and farming of fish. Almost all host fish are important and form a significant catch in commercial ladings in many countries, including India.
Data Availability
No datasets were generated or analysed during the current study.
Abbreviations
- RS:
-
Robust seta/e
- TL:
-
Total length
- W:
-
Width
References
Boyko CB, Bruce NL, Hadfield KA, Merrin KL, Ota Y, Poore GCB, Taiti S (2008) Nerocila Leach, 1818. World Register of Marine Species: World marine, freshwater and terrestrial isopod Crustaceans database. https://www.marinespecies.org/aphia.php?p=taxdetails&id=118417. Accessed 1 March 2023
Trilles JP, Rameshkumar G, Ravichandran S (2013) Nerocila species (Crustacea, Isopoda, Cymothoidae) from Indian marine fishes. Parasitol Res 112:1273–1286. https://doi.org/10.1007/s00436-012-3263-5
Bruce NL (1987a) Australian species of Nerocila Leach, 1818, and Creniola n. gen. (Isopoda: Cymothoidae), crustacean parasites of marine fishes. Rec Aust Mus 39(6):355–412.
Bruce NL (1987) Australian Pleopodias Richardson, 1910 and Anilocra Leach, 1818 (Isopoda: Cymothoidae), crustacean parasites of marine fishes. Rec Aust Mus 39(2):85–130.
Aneesh PT, Helna AK, Valarmathi K, Chandra K, Santanu M (2017) Redescription of Nerocila exocoeti Pillai, 1954 (Crustacea: Isopoda: Cymothoidae) parasitic on beloniform (Exocoetidae and Hemiramphidae) hosts with Nerocila madrasensis Ramakrishna & Ramaniah, 1978 placed into synonymy. Zootaxa 4356(3):385–394. https://doi.org/10.11646/zootaxa.4365.3.7
Aneesh PT, Valarmathi K, Santanu M (2019) Redescription of Nerocila recurvispina Schiödte and Meinert 1881: (Crustacea: isopoda) from the Hooghly River, Kolkata, India. Mar Biodivers 49:301–313. https://doi.org/10.1007/s12526-017-0799-8
Ravichandran S, Vigneshwaran P, Rameshkumar G (2019) A taxonomic review of the fish parasitic isopod family Cymothoidae Leach, 1818 (Crustacea: Isopoda: Cymothooidea) of India. Zootaxa 4622(1):001–099. https://doi.org/10.11646/zootaxa.4622.1.1
Pillai NK (1958) Tanaidacea and isopoda of Travancore. Dissertation, University of Kerala.
Rameshkumar G, Ravichandran S, Sivasubramanian K, Trilles JP (2013) New occurrence of parasitic isopods from Indian fishes. J Parasit Dis 37(1):42–46. https://doi.org/10.1007/s12639-012-0128-x
Aneesh PT, Sudha K, Arshad K, Anilkumar G, Trilles JP (2013) Seasonal fluctuation of the prevalence of cymothoids representing the genus Nerocila (Crustacea, Isopoda), parasitizing commercially exploited marine fishes from the Malabar Coast. India Acta parasitol 58(1):80–90. https://doi.org/10.2478/s11686-013-0112-3
Kulanthaivel R, Vijayakumar R, Velmurugan K, Saravanakumar A (2014) Occurrence of isopod Nerocila phaiopleura infestation on Whitefin wolf-herring (Chirocentrus nudus) from Southeast coast of India. J Parasit Dis 38(2):205–207. https://doi.org/10.1007/s12639-012-0224-y
Nagasawa K, Isozaki S (2017) Three new host records for the marine fish ectoparasite, Nerocila phaiopleura (Isopoda: Cymothoidae), with a list of its known hosts. J Crustac Biol 46:153–159. https://doi.org/10.18353/CRUSTACEA.46.0_153
Trilles JP, Ravichandran S, Rameshkumar G (2011) A checklist of the Cymothoidae (Crustacea, Isopoda) recorded from Indian fishes. Acta Parasitol 56(4):446–459. https://doi.org/10.2478/s11686-011-0077-z
Bleeker P (1857) Recherches sur les Crustacés de l’Inde archipélagique. II. Sur les Isopodes Cymothoadiens de l’archipel Indien. Natuurkundige Vereeniging in Nederlandsch-Indië, Batavia, Verhandelingen. https://doi.org/10.5962/bhl.title.9908
Aneesh PT, Helna AK, Kumar AB, Trilles JP (2020) A taxonomic review of the branchial fish parasitic genus Elthusa Schioedte & Meinert, 1884 (Crustacea: Isopoda: Cymothoidae) from Indian waters with the description of three new species. Mar Biodivers 50:65. https://doi.org/10.1007/s12526-020-01084-6
Aneesh PT, Hadfield KA, Smith NJ, Biju Kumar A (2021) Morphological description and molecular characterization of a new species of Anilocra Leach, 1818 (Crustacea: Isopoda: Cymothoidae) from India. Int J Parasitol Parasites Wildl 14:321–328. https://doi.org/10.1016/j.ijppaw.2021.03.007
Aneesh PT, Nashad M, Kumar AB, Bineesh KK, Hatha AAM (2022) Review on the global distribution and hosts of the external fish parasitic isopod genus Renocila Miers, 1880, (Crustacea: Cymothoidae), with the description of a new species from Andaman Islands. India J Nat Hist 55(43–44):2761–2785. https://doi.org/10.1080/00222933.2021.2019341
Froese R, Pauly D (2013) FishBase. World Wide Web electronic publication. https://www.fishbase.se/search.php. Accessed 03 March 2021
Fricke R, Eschmeyer WN, Fong JD (2021) Eschmeyer's Catalogue of Fishes. http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp#Sphyraenidae. Accessed 03 March 2021
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, Thierer T, Ashton B, Meintjes P, Drummond A (2012) Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28(12):1647–1649. https://doi.org/10.1093/bioinformatics/bts199
Tamura K, Stecher G, Kumar S (2021) MEGA 11: molecular evolutionary genetics analysis version 11. MBE 38(7):3022–3027. https://doi.org/10.1093/molbev/msab120
Amrutha SS, Balamurali RS, Arya U, Binumon TM (2021) First report of histopathological alterations by an isopod parasite in Indian mackerel, Rastrelliger kanagurta (Cuvier, 1817). J Mar Biol Ass India 63(2):88–92. https://doi.org/10.6024/JMBAI.2021.63.2.2203-12
Raja K, Vijayakumar R, Karthikeyan V, Saravanakumar A, Sindhuja K, Gopalakrishnan A (2014) Occurrence of isopod Nerocila phaiopleura infestation on Whitefin wolf-herring (Chirocentrus nudus) from Southeast coast of India. J Parasit Dis 38:205–207. https://doi.org/10.1007/s12639-012-0224-y
Barnard KH (1925) Contributions to the crustacean fauna of South Africa. No. 9. Further addition to the list of Isopoda. Ann S Afr Mus 20:381–412. https://biostor.org/reference/235227
Bowman TE, Tareen IU (1983) Cymothoidae from fishes of Kuwait (Arabian Gulf) (Crustacea: Isopoda). Smithson Contrib Zool 382:1–30. https://doi.org/10.5479/si.00810282.382
Ravichandran S, Ranjit Singh AJA, Veerappan B (2001) Parasite-induced vibriosis in Chirocentrus dorab off Parangipettai coastal waters. Curr Sci 80(5):622–623
Ghani N (2003) Isopod parasites of marine fishes of Pakistan. Proc Pakistan Congress Zool 23:217–221
Bruce NL, Harrison-Nelson EB (1988) New records of fish parasitic marine isopod crustaceans (Cymothoidae, subfamily Anilocrinae) from the Indo-West Pacific. Proc Biol Soc Wash 101:585–602. https://biostor.org/reference/74657
Mitani I (1982) Variations of condition factor of sardine as host by Nerocila phaiopleura Bleeker as parasites. Bull Japan Soci Sci Fish 48:611–615. https://doi.org/10.2331/suisan.48.611
Bleeker P (1857) Recherches sur les Crustacés de l’Inde Archipelagique. II. Sur les Isopodes Cymothoadiens de l’Archipel Indien. Verhandelingen der Natuurkundige Vereeniging in Nederlandsch-Indië. d.2. Batavia, Lange, pp 20–40. https://doi.org/10.5962/bhl.title.9908
Morton B (1974) Host specificity and position on the host in Nerocila phaeopleura Bleeker (Isopoda, Cymothoidae). Crustaceana 26:143–148
Bharadhirajan P, Murugan S, Sakthivel A, Selvakumar P (2014) Isopods parasites infection on commercial fishes of Parangipettai waters, southeast coast of India. Asian Pac J Trop Med 4:S268–S272. https://doi.org/10.1016/S2222-1808(14)60453-9
Rameshkumar G, Ramesh M, Ravichandran S, Trilles JP (2016) Parasitic isopods from marine fishes off Nagapattinam coast. India J Parasit Dis 40(3):940–944. https://doi.org/10.1007/s12639-014-0610-8
Bruce NL (1982) On a small collection of marine Isopoda from Hong Kong. In: Morton BS, Cheng CK (eds) Proceedings of the first international marine biological workshop: the marine flora and fauna of Hong Kong and Southern China, Hong Kong. Hong Kong University Press, Hong Kong, pp 315–324.
Williams EH, Bunkley-Williams L (1986) The first Anilocra and Pleopodias isopods (Crustacea: Cymothoidae) parasitic on Japanese fishes, with three new species. Proc Biol Soc 99:647–657
Hiramoto K (1996) Natural history of Japanese sardine–survival strategy of ‘rice of the sea’. Chuokoron-sha, Tokyo, pp 183.
Saito N, Hayase Y (2000) Note on an aegathoid stage of cymothoid isopod, Nerocila phaiopleura Bleeker, 1857 (Crustacea: Isopoda: Cymothoidae) stranded at Miho beach, Suruga Bay, middle of Japan. I. O. P. Diving News 11(10): 2–6.
Nunomura N (2011) Crustaceans No. 2 (Isopoda). Special Publication of the Toyama Science Museum 24:1–133
Hata H, Sogabe A, Tada S, Nishimoto R, Nakano R, Kohya N, Takeshima H, Kawanishi R (2017) Molecular phylogeny of obligate fish parasites of the family Cymothoidae (Isopoda, Crustacea): evolution of the attachment mode to host fish and the habitat shift from saline water to freshwater. Mar Biol 164:105. https://doi.org/10.1007/s00227-017-3138-5
Nagasawa K, Nitta M, Otawa T, Ishikawa T (2020) Nerocila phaiopleura (Isopoda: Cymothoidae): a new record from Ibaraki Prefecture, central Japan, with a discussion of its distribution in Japanese waters. Crustac Res 49:41–47. https://doi.org/10.18353/crustacea.49.0_41
Monod T (1934) Isopodes marins des campagnes du ‘de Lanessan’. Institut Océanographique de l’Indochine. Station Maritime de Cauda, Nha Trang, Viet Nam, Note. Gouvernement Général de l’Indochine, Saigon 23:1–22
Ravichandran S, Rameshkumar G (2014) Effect of parasitic isopods in commercial marine fishes. J Aquat Biol Fish 2:574–579
Rajkumar M, Perumal P, Trilles JP (2006) On the occurrence of a double parasitism (copepod and isopod) on the anchovy fish in India. J Environ Biol 27(3):613–614
Rajkumar M, Thavasi R, Perumal P, Trilles JP (2007) Parasite induced vibriosis in Stolephorus commersonii. Res J Microbiol 2:972–977. https://scialert.net/abstract/?doi=jm.2007.972.977
Barnard KH (1936) Isopods collected by the R.I.M.S. “Investigator.” Records of the Indian Museum, Calcutta, vol 38, pp 147–191
Trilles JP (1979) Les Cymothoidae (Isopoda, Flabellifera; parasites de poissons) du Rijksmuseum van Natuurlijke Historie de Leiden. 2. Afrique, Amerique et regions Indo-ouest-Pacifiques. Zool Med Leiden 5417:245–275
Nagasawa K, Tensha K (2016) Nerocila phaiopleura (Isopoda: Cymothoidae) parasitic on Japanese Spanish mackerel Scomberomorus niphonius in the Seto Inland Sea, Japan. Biogeography 18:71–75. https://doi.org/10.11358/biogeo.18.71
Nagasawa K, Isozaki S (2020) The cymothoid isopod Nerocila phaiopleura parasitic on gnomefish, Scombrops boops, in coastal Pacific waters of central Japan, with an updated list of the hosts reported from Japan. Nature Kagoshima 46:525–530. http://journal.kagoshima-nature.org/archives/NK_046/046-093
Masakazu K, Takehiro K, Hiroyuki Y, Shinya Y (2021) New host records for Nerocila japonica and N. phaiopleura (Isopoda: Cymothoidae). J Natl Fish Univ 69(4):103–107.
Rameshkumar G, Ravichandran S (2010) New host record, Rastrelliger kanagurta, for Nerocila phaeopleura parasites (Crustacea, Isopoda, Cymothoidae). Mid East J Sci Res 5:54–56.
Seth JK, Sahoo S, Mitra S (2014) First record of isopod parasite, Nerocila phaeopleura on the host fish Rastrelliger kanagurta collected from Bay of Bengal, Odisha coast. India Int J Curr Res 6(4):6092–6093
Ramudu KR, Rathod JL (2023) A temporal analysis of parasitic infections in Indian Mackerel, Rastrelliger kanagurta (Cuvier, 1817) along the Western Coast of Karwar Bay, Karnataka: employing a monthly assessment with histopathology. J Adv Zool 44(3):465–472. https://doi.org/10.17762/jaz.v44i3.497
Ramudu KR, Rathod JL (2024) Seasonal parasitic study on Indian mackerel, Rastrelliger kanagurta (Cuvier, 1817) associated with a secondary bacterial infection in Karwar, Uttara Kannada, Karnataka. J Exp Zool India 27(1):667–672. https://doi.org/10.51470/jez.2024.27.1.667
Nagasawa K, Nakao H (2017) Chub mackerel, Scomber japonicus (Perciformes: Scombridae), a new host record for Nerocila phaiopleura (Isopoda: Cymothoidae). Biosphere Sci 56:7–11. https://doi.org/10.15027/44653
Nagasawa K, Shirakashi S (2017) Nerocila phaiopleura, a cymothoid isopod parasitic on Pacific bluefin tuna, Thunnus orientalis, cultured in Japan. Crustac Res 46:95–101. https://doi.org/10.18353/crustacea.46.0_95
Jalaja Kumari C, Rao KH, Shyamasundari K (1987) A new cymothoid isopod of the genus Nerocila leach Crustacea Isopoda Flabellifera parasitic on marine fishes of the Waltair coast India. Rev Iberica Parasitol 47(4):413–418. https://eurekamag.com/research/004/593/004593518.php
Acknowledgements
We thankfully acknowledge the University of Kerala for providing the financial support through Research Grant (AcE VI/117/ZOO/15059/2017).
Funding
Open Access funding provided by Hiroshima University.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study's conception and design. ASS wrote the first draft of the manuscript and conducted experiments. PTA and BRPSN analysed the results, reviewed the draft of the paper. BTM sample collection and prepared the drawings of the specimens. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Conflict of interests
The authors declare that they have no known competing financial interests or personal relationships that could have influenced the work reported in this paper. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Suresh, A.S., Nair, B.R.P.S., Mangalathettu, B.T. et al. Redescription and Molecular Characterization of the External Attaching Fish Parasitic Cymothoid, Nerocila phaiopleura Bleeker, 1857 (Crustacea: Isopoda) off the Southwest Coast of India. Acta Parasit. (2024). https://doi.org/10.1007/s11686-024-00870-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11686-024-00870-7