
Abstract
Diabetes is associated with macrovascular and microvascular complications and is a major risk factor for neurological and psychiatric disorders, such as dementia and depression. Type 1 diabetes (T1DM) and type 2 diabetes (T2DM) have distinct etiologies and pathophysiological effects while sharing a common endpoint of persistent hyperglycemia. Neuroimaging studies in T1DM have revealed reductions in numerous regions, including the parahippocampal and occipital regions, while in T2DM there have been numerous reports of hippocampal atrophy. This meta-analysis aimed to identify consistent regional abnormalities in cerebral structures in T1DM and T2DM respectively, and also to examine the impact of potential confounds, including age, depression and vascular risk factors. Neuroimaging studies of both voxel-based morphometry (VBM) data and volumetric data were included. Ten T1DM studies (n = 613 patients) and 23 T2DM studies (n = 1364 patients) fulfilled inclusion criteria. The T1DM meta-analysis revealed reduced bilateral thalamus grey matter density in adults. The T2DM meta-analysis revealed reduced global brain volume and regional atrophy in the hippocampi, basal ganglia, and orbitofrontal and occipital lobes. Moreover, hippocampal atrophy in T2DM was not modified by hypertension, although there were more marked reductions in younger patients relative to healthy controls. In conclusion, T1DM and T2DM demonstrated distinct cerebral effects with generalised and specific target areas of grey matter reduction. Thalamic atrophy in T1DM may be a substrate of associated cognitive deficits. In T2DM, global cerebral atrophy may reflect atherosclerotic factors, while hippocampal atrophy was an independent effect providing a potential common neuropathological etiology for the comorbidity of T2DM with dementia and depression.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Diabetes is a multi-system disease characterized by persistent hyperglycemia associated with numerous macrovascular complications, such as cardiovascular and cerebrovascular disease, as well as microvascular complications, including retinopathy, nephropathy, and neuropathy. Diabetes has emerged as a major risk factor for numerous conditions affecting the central nervous system. Up to 20 % of adults with diabetes meet criteria for major depression (Ali et al. 2006), and diabetes has been found to double the risk of Alzheimer’s disease and all-cause dementia (Ott et al. 1999).
Diabetes is currently subdivided into two subtypes, which share a common end-point of persistent hyperglycemia. However, type 1 diabetes (T1DM) and type 2 diabetes (T2DM) have different etiologies. T1DM results from autoimmune destruction of islet beta-cells with a typical onset in childhood and young adulthood. T2DM usually results from increasing resistance to the end-organ actions of insulin and progressive impairment of insulin secretion, and its end effects often occur with other cardiovascular risk factors, such as hypertension, dyslipidemia, and obesity (Stamler et al. 1993).
Altered cerebral metabolism has been observed in T1DM in children (Sarac et al. 2007). T1DM in adults has been associated with neurocognitive deficits, such as impaired attention and information processing (Brands et al. 2005), and corresponding reductions in cortical and subcortical regions, including occipital and inferior frontal (Wessels et al. 2006), thalamus, and parahippocampal regions (Northam et al. 2009). In T2DM, decreases in whole brain volume (Espeland et al. 2013) as well as in regional volumes, namely in the hippocampus and amygdala (den Heijer et al. 2003), anterior cingulate (Kumar et al. 2008) and cerebellum (Manor et al. 2012) have been observed. Findings have been mixed in part due to variability in method of analysis, some studies using voxel-based morphometry (Musen et al. 2006) with others using volumetric analysis (Lobnig et al. 2005), and some studies have limited their analyses to specific regions of interest (Anan et al. 2010; Kamiyama et al. 2010).
In order to investigate the regional cerebral correlates of T1DM and T2DM, we performed a meta-analysis of studies of grey matter effects including studies of both VBM and volumetric measures. Our aim was to identify the consistent regional abnormalities in T1DM and T2DM and to examine the effects of the potential confounds of depression, hypertension, hyperlipidemia, glycemic control, age, and gender. We hypothesized that grey matter changes in T2DM would demonstrate strong parallels with the deficits observed in depression and dementia, namely hippocampal atrophy (Cole et al. 2011), whilst other regional abnormalities would be observed in T1DM.
Materials and methods
Literature search
The meta-analyses followed the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines. We systematically searched Web of Science and MEDLINE for original articles published before May 2013 using the following search strategy: (diabetes OR diabetic) AND (morphometry OR voxel-based OR voxelwise OR VBM OR volumetric OR ‘gray matter’ OR ‘grey matter’ OR ‘brain volume’ OR ‘brain volumes’ OR ‘cortical atrophy’ OR ‘brain atrophy’ OR ‘subcortical atrophy’ OR MR OR MRI OR ‘magnetic resonance’). Studies using animals, other imaging modalities, and patients with major psychiatric comorbidity (e.g. psychosis, alcoholism) or previous cerebrovascular disease were excluded. Titles were reviewed to exclude studies not meeting the inclusion criteria, and remaining abstracts were examined. From abstracts meeting inclusion criteria, full-text articles were reviewed, and relevant studies carried forward for data extraction. Reference lists of included papers were checked for additional publications, and both published and unpublished articles were included.
Criteria for inclusion in analysis
Inclusion criteria were: 1) volumetric and/or voxel-based morphometry data on grey matter structure in either T1DM or T2DM; 2) at least one direct structural or longitudinal comparison was made between patients and controls; 3) MRI imaging was used with voxel-based analysis or volumetric data produced using a validated manual- or automatic segmentation method; and 4) whole-brain and/or region of interest (ROI) analysis was produced. Studies were excluded if they met any of the following criteria: 1) patients had a diagnosis of insulin resistance; 2) study did not differentiate T1DM from T2DM; 3) only white matter data were presented; 4) fewer than ten individuals were included or fewer than five subjects in either group; 5) study included patients with dementia only; 6) study re-analyzed previously published data; 7) study was a review article without original data; or 8) not published in English (Fig. 1).

Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) flow diagram of search strategy
Data extraction
The following data were extracted for all relevant studies: authors; year of publication; sample size; age (mean + standard deviation (SD)); type of diabetes; MRI modality; and method of analysis used.
We identified the following possible confounders: duration of diabetes (mean + SD); HbA1c percentage at time of study (mean + SD), converted to International Federation of Clinical Chemistry-standardized concentrations (mmol/mol); hypertension; hyperlipidemia; previous stroke; dementia; insulin therapy; and correction for intracranial volume. Covariate analysis was performed when there were at least five independent studies with complete data for a particular brain region. If covariate analysis was not possible, stratified analysis was performed (e.g. analyzing studies with children and adult participants separately). For any data reported as mean +/- standard error, the latter was converted to its SD for inclusion in the meta-analysis. For prospective studies, baseline measures were also included in the meta-analysis when available.
Studies presenting overlapping or identical samples were identified, and only the study presenting the largest number of subjects retained. If there was possible overlap but different results were presented, for example hippocampus presented in one study and frontal lobe in the other, all data were included. The following 9 studies were assimilated into 4 studies because of potential sample overlap: i) Brundel et al. 2010; de Bresser et al. 2010 and Jongen et al. 2007; ii) Anan et al. 2010 and Anan et al. 2011; iii) Manor et al. 2012 and Novak et al. 2011; iv) Hershey et al. 2010 and Perantie et al. 2011.
Volumetric meta-analysis
Grey matter volumetric measurements of global (whole brain) or regional volumes were included that were reported by at least 2 studies. Authors were contacted for studies in which the reported measurements were unclear.
DerSimonian-Laird random-effects meta-analysis was used (DerSimonian and Laird 1986). In this method, the standard errors of the study-specific estimates are adjusted to incorporate a measure of heterogeneity observed in different studies. The amount of variation, and hence the adjustment carried out, is estimated from the intervention effects and standard errors of included studies. R statistical software was used to produce an effect size estimate (Hedge’s g) for each region analyzed (standardized mean difference for continuous variables), 95 % confidence interval, and z-value and p-value (Viechtbauer 2010). The heterogeneity quantifier I2 was produced for each area of interest (Higgins et al. 2003).
Voxel-based morphometry meta-analysis
Voxel-based morphometry (VBM) is a fully-automated and standardized method of determining tissue-type probabilities (Ashburner and Friston 2000), and we performed a meta-analysis to produce a whole-brain summary map based on VBM effect sizes. The following data were extracted: normalization template (such as MNI or Talairach); x/y/z coordinates; area examined (whether whole brain or region of interest); statistic used to report the findings (if p-values, whether uncorrected or corrected and the type of correction used); effect sizes; and the size of the smoothing kernel used. If the statistical threshold was not reported by a study, the smallest effect size of significant findings was used as a conservative estimate. For region of interest analyses or when the coordinates were not specified, automated anatomical labels (AAL) were used to record the relevant area (Tzourio-Mazoyer et al. 2002). Where appropriate, we duplicated the result to account for more than one AAL label.
Parametric coordinate-based meta-analysis (PCM) was used to analyze the VBM data (Costafreda 2012). PCM is a method that can incorporate positive, negative and sub-threshold statistical findings from VBM studies, themselves with different statistical thresholds, to generate a valid pooled effect size summary. A finding can be incorporated into PCM if sufficient information is reported to compute the effect size associated with the finding, and PCM employs a spatial kernel to account for the spatial uncertainty in location. If a study reported both whole-brain and ROI contrasts, the most precise measurement available (either an exact measurement for a significant finding or most conservative threshold for a negative result) is used in the summary map for that study. PCM corrects for multiple comparisons using the false discovery rate correction (q = 0.05).
Each significant triplet of spatial coordinates is assigned to the corresponding location in MNI space, and this location receives the effect size reported, either as exact value or significant interval (not crossing zero). Two maps are created, reflecting the lower and upper bounds of these intervals. After convolution with the kernel, the outcome is a brain map for study i, in which either a significant measurement (if the location is located within ρ of a significant finding) or a non-significant measurement (determined by the statistical threshold) is assigned to each location within the field of view. Outside of this, the measurement is not defined. The non-significant measurement will be left- or interval-censored containing zero, bounded by the threshold ti,. If the threshold is not stated in the paper and cannot be computed or approximated, the significance of the smallest significant finding is used to estimate ti, thereby maximizing the width of the non-significance interval and producing a conservative estimate. This method demonstrates high agreement between its estimates and those obtained from the meta-analysis of unthresholded manual volumetric measurements, and further details on PCM methods have been published (Costafreda 2012).
Results
In T1DM, 10 studies (n = 613 T1DM patients, mean age 26.6 years; n = 375 healthy controls, mean age 25.8 years) consisting of 6 VBM (Musen et al. 2006; Northam et al. 2009; Perantie et al. 2007; Perantie et al. 2011; Wessels et al. 2007; Van Elderen et al. 2011) (n = 453 patients, n = 286 controls) and 4 volumetric studies (n = 160 pts, n = 89 controls) were eligible for inclusion (Table 1). In T2DM, 23 studies (n = 1364 T2DM patients, mean age 63.2 years; n = 3433 healthy controls, mean age 60.5 years), consisting of 6 VBM (Anan et al. 2010; Anan et al. 2011; Chen et al. 2012; Gold et al. 2007; Hayashi et al. 2011; Kamiyama et al. 2010) (n = 216 pts, n = 202 controls) and 17 volumetric studies (Alosco et al. 2013; Bruehl et al. 2009; Brundel et al. 2010; de Bresser et al. 2010; den Heijer et al. 2003; Espeland et al. 2013; Hempel et al. 2012; Jongen et al. 2007; Korf et al. 2006; Kumar et al. 2008; Last et al. 2007; Manor et al. 2012; Musen et al. 2012; Novak et al. 2011; Tiehuis et al. 2008; Watari et al. 2008; Yau et al. 2010) (n = 1148 patients, n = 3231 controls) were included (Table 2).
Average duration of diabetes was 14.5 years for T1DM and 10.3 years for T2DM, and average HbA1c was 8.2 % (66 mmol/mol) for T1DM (Table 1) and 7.6 % (60 mmol/mol) for T2DM (Table 2), as a proxy measure of glycemic control. Additional potential confounders of depression, dementia, stroke, hypertension, hyperlipdemia, and correction for intracranial volume are presented in Supplementary Tables 1 and 2.
Type 1 diabetes
In the volumetric meta-analysis, there were no significant differences between in total brain volume, total grey matter volume, or hippocampal volume (all p > 0.1), and these results are published as supplementary material (Supplementary Figure 1, Supplementary Table 3).
In the VBM meta-analysis, significant reductions were observed in both the left thalamus (cluster size 5768 mm3, subregion volume 5112 mm3, MNI coordinates (−14, −22, 0), z = 4.46, p < 0.005 FDR corrected) and right thalamus (cluster size 5768 mm3, subregion volume 612 mm3, MNI coordinates (4, −16, 0), z = 4.46, p < 0.005 FDR corrected) as compared to controls (Fig. 2). Stratification by age revealed significant thalamic reductions in the aggregation of studies which included adults (Musen et al. 2006; Northam et al. 2009; Wessels et al. 2006), but no differences in children (Perantie et al. 2007; Perantie et al. 2011). No other regions showed any significant differences between T1DM and healthy controls.

VBM meta-analysis map of T1DM (axial, sagittal and transverse cerebral views are presented, overlaid on MNI template). Red areas are statistically significant results (P < 0.05 FDR corrected). Significant reductions were found in left thalamus (MNI coordinates (−14, −22, 0), z = 4.46) and right thalamus (MNI coordinates (4, −16, 0), z = 4.46) in T1DM compared to controls
Type 2 diabetes
The volumetric meta-analysis demonstrated significant global reductions in total brain volume (Z = −2.77, p = 0.006), brain parenchymal fraction (Z = −7.77, p < 0.001) and grey matter volume (Z = −4.75, p < 0.001) (Table 3, Supplementary Figures 2 and 3). Regional reductions were revealed in orbitofrontal cortex grey matter (Table 3, Z = −3.29, p = 0.001), the hippocampus (Fig. 3, Z = −3.67, p = 0.0002) and basal ganglia (Table 3, Z = −2.61, p = 0.009). Reductions in frontal and temporal volumes approached statistical significance (p = 0.057 and p = 0.066, respectively) (Supplementary Figure 4). There were no significant differences in anterior cingulate, superior temporal, and parietal regions (Table 3).

Forest plot of hippocampal volume in T2DM studies. Significant reduction in T2DM compared to controls
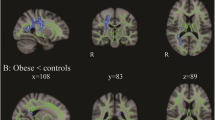
The VBM meta-analysis revealed significant reductions in both the left hippocampus (cluster size 7224 mm3, subregion volume 7224 mm3, MNI coordinates (−32, −6, −26), z = 2.67, p = 0.0076) and right hippocampus (cluster size 7296 mm3, subregion volume 7296 mm3, MNI coordinates (26, −6, −26), z = 2.67, p = 0.0076) compared to healthy controls (Fig. 4).

VBM meta-analysis map of T2DM (axial, sagittal and transverse cerebral views are presented, overlaid on MNI template). Red areas are statistically significant results (P < 0.05 FDR corrected). Significant reductions were found in left hippocampus (MNI coordinates (−32, −6, −26), z = 2.67) and right hippocampus (MNI coordinates (26, −6, −26), z = 2.67) in T2DM compared to controls
We also examined potential longitudinal changes, which was possible for total brain volume from 2 studies (n = 113 patients, mean age 65.9 years; n = 668 controls, mean age 78.1 years) (de Bresser et al. 2010; Espeland et al. 2013). Accelerated brain atrophy was evident in older T2DM patients relative to age-matched healthy controls (Hedges’ g = −0.25 95 % CI (−0.49; −0.02); z = 2.16, p = 0.03) with a follow-up period of 4.7 and 4.1 years respectively.
Modifier analysis
Modifier analysis was only performed if there were at least 5 studies with complete data for a particular region. Reductions in total brain volume and the hippocampus in T2DM were robust to adjustment by age (Table 3), and age was a significant modifier of the differences between patients and controls in hippocampal volume, with studies that included younger patients showing more marked reductions between patients and controls than those including older participants (modifier effect Z = 6.2, p < 0.001).
Hypertension was associated with more pronounced differences in frontal volumes between T2DM patients and controls: studies where a higher proportion of patients with T2DM had comorbid hypertension reported smaller frontal volume in T2DM relative to controls (modifier Z = −2.51, p = 0.012). There were no significant effects of gender or age on frontal reductions. For total grey matter volume, hypertension as a modifier of the differences between patients and controls approached statistical significance (modifier Z = −1.89, p = 0.058), whereas age, gender or HbA1c did not modify these differences. For hippocampal volume, neither hypertension nor glycemic control (using HbA1c as a proxy measure) modified the differences between patients and controls, though again data were limited and between-study differences in HbA1c relatively small.
Where possible, the effect of depression on results was analyzed by estimating differences within each study between both groups of patients. There was no evidence that depression was associated with differences in anterior cingulate or orbitofrontal volumes, although only 2 studies examined these areas, and no data were available for hippocampal volume. Due to the limited number of studies, modifier analysis was not performed in T1DM studies or to examine the effects of hyperlipidemia.
Discussion
Our meta-analysis of the structural brain changes associated with T1DM and T2DM demonstrated distinct effects within the conditions with generalized and specific target areas of grey matter reduction. T1DM was associated with significant bilateral reductions that were limited to the thalamus in adults with T1DM. In contrast, T2DM was associated with widespread global cerebral atrophy, which declined more rapidly over time compared to healthy controls, as well as regional atrophy, particularly in the hippocampus, basal ganglia, orbitofrontal, and occipital lobes. While hypertension modified the global cerebral effects of diabetes, indicating a potential atherosclerotic contribution, there was no significant modifier effect of hypertension on hippocampal atrophy, which may provide a common neural pathology for the high comorbidity of T2DM with dementia as well as depression.
Type 1 diabetes
The VBM meta-analysis revealed significantly lower grey matter density in the right thalamus in T1DM. The significant reductions in thalamus were not seen when analysing childhood studies only. In support, blunted thalamic blood flow has been reported in T1DM (Mangia et al. 2012). There is growing interest in the neuropsychological effects of T1DM in both children and adults, with decrements found in IQ, attention and higher-order executive function (Gaudieri et al. 2008; Naguib et al. 2009). The area of the right thalamus identified in our meta-analysis is thought to have prominent projections to the prefrontal cortex with no motor connections (Oxford Thalamic Connectivity Atlas 2003), so could potentially be a substrate of associated cognitive deficits. Insulin receptor expression is particularly rich in the anterior thalamic and hypothalamic nuclei of rat brains (Bondy et al. 1992), identifying the thalamus as an area of potential vulnerability to damage in T1DM.
The effects of T1DM on cognitive impairment have been linked both with prolonged exposure to hyperglycemia (Musen et al. 2006) and with repeated episodes of hypoglycemia, though this has been disputed (DCCT et al. 2007). In our meta-analysis, the effects of previous hypoglycemia and hyperglycemia were mixed (Musen et al. 2006; Perantie et al. 2011; Aye et al. 2011), but there were insufficient data to test either exposure quantitatively (Supplementary Table 1). .
Some authors have highlighted parallels between the cerebral correlates of T1DM and those of depression, advancing this as evidence for shared biological mechanisms between the two conditions (Korczak et al. 2011). Our meta-analysis, however, suggests that this conclusion is not supported by the available grey matter studies of T1DM and, indeed, that distinct cerebral effects may be observed.
Type 2 diabetes
T2DM patients have a 39 % increased risk of Alzheimer’s disease and a 47 % increased risk of all-cause dementia (Lu et al. 2009). Our meta-analysis revealed global cerebral atrophy and implicates the hippocampus as a target of T2DM pathology in the brain. Hippocampal atrophy was demonstrated at high significance and survived covarying for hypertension, making it unlikely that hippocampal atrophy is the result only of vascular risk factors and atherosclerosis. Furthermore, the difference in hippocampal volume was even more marked for younger patients. This probably reflects the increased contribution of factors other than T2DM leading to hippocampal atrophy in older patients, and thereby strengthens the association of hippocampal atrophy with T2DM. In rat models, it has been demonstrated that insulin provides significant neurotrophic support to the hippocampus, which may be impaired in the diabetic brain (Balakrishnan et al. 2010).
Our findings also showed lower volumes of subcortical structures in T2DM compared to healthy controls. In early Alzheimer’s disease, amyloid deposits have been found in the striatum (Klunk et al. 2007), while Alzheimer’s disease is associated with reduced putamen and thalamic volumes (de Jong et al. 2008). As the basal nuclei and thalamus are involved with emotional, motivational, and cognitive abilities (Herrero et al. 2002), the shared finding of reduced subcortical volume in Alzheimer’s disease and in T2DM could provide further insight into their relationship. In our meta-analysis, only five studies did not exclude participants with dementia and one of these studies found that covarying for dementia did not modify the significant difference between T2DM and controls (Korf et al. 2006).
As well as dementia, there is a well-established bidirectional link between T2DM and depression (Ali et al. 2006; Mezuk et al. 2008). Reduced hippocampal volume has been consistently observed in major depression (Cole et al. 2011; Videbech and Ravnkilde 2004), therefore our finding of reduced hippocampal volume in T2DM may implicate a common pathophysiology. Deficiency of brain derived neurotrophic factor (BDNF) has been linked both to depression and T2DM (Karege et al. 2002; Fujinami et al. 2008), potentially via decrements in hippocampal function (Egan et al. 2003). Unfortunately, in our meta-analysis there were insufficient data to examine whether the interaction between depression and T2DM had a greater effect on hippocampal volume than T2DM alone.
Both diabetes and depression are associated with activation of the innate inflammatory response (Dowlati et al. 2010; Pickup and Crook 1998), and patients with depression and T2DM have been shown to have higher levels of inflammatory markers than patients with T2DM alone (Laake et al. 2014). Depressed subjects have significantly higher concentrations of tumour necrosis factor-α and interleukin-6 (IL-6) (Dowlati et al. 2010), and high IL-6 levels in depression are associated with reduced hippocampal volumes (Frodl et al. 2012). In addition, both depression and diabetes are also associated with overactivation of the hypothalamic-pituitary-adrenal (HPA) axis (Stetler and Miller 2011). Dysregulation of the HPA axis has been observed in patients who experienced childhood stress/adversity, and such patients are at increased risk of developing both depression and incident diabetes in adult life (Colman et al. 2014; Scott et al. 2011). In the brain, structural consequences of early stress include attenuated development of the left neocortex and hippocampus, which have a high density of glucocorticoid receptors (Teicher et al. 2003), while excess cortisol has been shown to hinder hippocampal neurogenesis (Elder et al. 2006). It is possible, therefore, that overactivation of the innate inflammatory response and/or HPA axis may provide a common link between both diabetes and depression and hippocampal atrophy.
Another finding in our meta-analysis was reduced orbitofrontal cortex grey matter in T2DM. This area has been implicated in the regulation and modulation of cognition and mood (Ballmaier et al. 2004), and as a predictor of clinical outcome in depression (Fu et al. 2013), pointing to a potential link between T2DM and associated cognitive impairment as well as depressive symptomatology.
Although the relationship between diabetes and depression and dementia individually is well established, these associations should not be considered in isolation. In T2DM, depression doubles the risk of incident dementia over five years (Katon et al. 2012). The risk of dementia was even more marked for younger patients, while cohort studies of T2DM have found that patients with depression are more likely to be younger (Twist et al. 2013). It is possible, therefore, that younger patients with T2DM – who demonstrated more marked hippocampal atrophy relative to older patients in our meta-analysis – are more vulnerable to the development of psychiatric complications such as depression and dementia, and/or to their detrimental effects on grey matter structure. The temporal relationship of these associations will require longitudinal study, and our meta-analysis identifies the relationship between T2DM and depression as a major gap in the neuroimaging literature which requires further exploration.
Possible cellular mechanisms
Several molecular mechanisms have been proposed to explain the regional cerebral effects of diabetes, including cerebral capillary basement membrane thickening, amyloid angiopathy and increased rate of brain infarcts (Biessels et al. 2006). Hyperglycaemia decreases vasodilatation by decreased expression of endothelial nitric oxide synthase, which in turn decreases cerebral blood flow (Tesfomariam et al. 1991; Williams et al. 1998). Impaired insulin signalling and hyperglycemia cause increased expression and activation of NF-κB, which is a key player in the inflammatory cascade and a modulator of apoptosis (Sima et al. 2009). Chronic hyperglycemia is associated with increased advanced glycation end products, which may enhance inflammatory processes and oxidative stress in the cerebral vasculature (Brownlee 1995). The latter can also contribute to amyloid deposition and tau phosphorylation (Necula and Kuret 2004), reminiscent of the pathogenesis of Alzheimer’s disease. Some authors have highlighted diminished O-linked N-acetylglucosamine glycosylation (O-GlcNAcylation) as a molecular mechanism linking diabetes and Alzheimer’s disease. A rat model using stroptozotocin showed down-regulation of O-GlcNAcylation and impaired insulin signalling (Deng et al. 2009). As decreased O-GlcNAcylation is found in Alzheimer’s disease and is inversely related to the phosphorylation of tau (Liu et al. 2009), subsequent hyperphosphorylation of tau could lead to pathology associated with Alzheimer’s disease.
Limitations
Our study was limited by a lack of data for many brain areas, in particular for T1DM. Where study numbers were particularly limited, the findings should be treated with caution. Although the lack of a modifying effect of hypertension and more marked differences seen in younger patients lend robustness to the hippocampal findings in T2DM, the greater number of studies examining the hippocampus increases the likelihood of positive findings compared to other brain areas. Lack of data limited the amount of covariate analysis possible, meaning the effect of other confounders on our findings cannot be excluded. A very small number of studies examined patients with potentially confounding disease processes, such as heart failure and arterial disease, though studies examining previous stroke were excluded altogether. Although various hypotheses were examined by the included studies, the vast majority examined the general population with T1DM or T2DM.
Our analysis was limited to grey matter changes, thereby not including the large amount of research on white matter changes in T2DM, which would also benefit from consolidation in meta-analysis. As with any meta-analysis, there is heterogeneity in the study populations and methods, for example with different MRI scanners and different segmentation techniques. The largely cross-sectional nature of our analysis makes it difficult to draw firm conclusions about the temporal relationship between diabetes and associated psychiatric conditions. However, by performing two separate meta-analyses, we were able to include 33 suitable studies in the analysis and, through PCM meta-analysis, to include negative results, a feature not possible with some other VBM analysis techniques.
Conclusions
Diabetes is associated with global cerebral deficits as well as discrete regional cortical and subcortical atrophy. T1DM was associated with lower VBM signal in the thalamus bilaterally, which may provide insight into associated cognitive deficits. T2DM was associated with atrophy in numerous brain areas but most strikingly in hippocampus, a potential link for its high comorbidity and risk for both depression and dementia. The mechanisms behind this may include inflammatory processes, oxidative stress and apoptosis as a result of aberrant insulin signalling. Further research is needed to understand these changes, and longitudinal research into the factors leading to grey matter loss in diabetes and its relationship with other psychiatric conditions is required.
References
Ali, S., Stone, M. A., Peters, J. L., Davies, M. J., & Khunti, K. (2006). The prevalence of co-morbid depression in adults with type 2 diabetes: a systematic review and meta-analysis. Diabetic Medicine, 23, 1165–1173.
Alosco, M., Brickman, A. M., Spitznagel, M. B., Griffith, E. Y., Narkhede, A., Raz, N., et al. (2013). The adverse impact of type 2 diabetes on brain volume in heart failure. Journal of Clinical and Experimental Neuropsychology, 35, 309–318.
Anan, F., Masaki, T., Shimomura, T., Fujiki, M., Umeno, Y., Eshima, N., et al. (2010). Abdominal visceral fat accumulation is associated with hippocampus volume in non-dementia patients with type 2 diabetes mellitus. NeuroImage, 49, 57–62.
Anan, F., Masaki, T., Shimomura, T., Fujiki, M., Umeno, Y., Eshima, N., et al. (2011). High-sensitivity C-reactive protein is associated with hippocampus volume in nondementia patients with type 2 diabetes mellitus. Metabolism, Clinical and Experimental, 60, 460–66.
Ashburner, J., & Friston, K. J. (2000). Voxel-based morphometry–the methods. NeuroImage, 11(6 Pt 1), 805–21.
Aye, T., Reiss, A. L., Kesler, S., Hoang, S., Drobny, J., Park, Y., et al. (2011). The feasibility of detecting neuropsychological and neuroanatomic effects of type 1 diabetes in young children. Diabetes Care, 34, 1458–1462.
Balakrishnan, S., Mathew, J., & Paulose, C. S. (2010). Cholinergic and glutamergic receptor functional regulation in long-term, low dose somatotropin and insulin treatment to ageing rats: rejuvenation of brain function. Mollecular and Cellular Endocrinology, 314, 23–30.
Ballmaier, M., Toga, A. W., Blanton, R. E., Sowell, E. R., Lavretsky, H., et al. (2004). Anterior cingulate, gyrus rectus, and orbitofrontal abnormalities in elderly depressed patients: an MRI-based parcellation of the prefrontal cortex. American Journal of Psychiatry, 161, 99–108.
Biessels, G. J., Staekenborg, S., Brunner, E., Brayne, C., & Scheltens, P. (2006). Risk of dementia in diabetes mellitus: a systematic review. Lancet Neurology, 5, 64–74.
Bondy, C., Bach, M. A., & Lee, W.-H. (1992). Mapping of brain insulin and insulin-like growth factor receptor gene expression by in situ hybridization. Neuroprotocols, 1, 240–249.
Brands, A. M., Biessels, G. J., de Haan, E. H., Kappelle, L. J., & Kessels, R. P. (2005). The effects of type 1 diabetes on cognitive performance: a meta-analysis. Diabetes Care, 28, 726–735.
Brownlee, M. (1995). Advanced protein glycosylation in diabetes and aging. Annual Review of Medicine, 46, 223–234.
Bruehl, H., Wolf, O. T., & Convit, A. (2009). A blunted cortisol awakening response and hippocampal atrophy in type 2 diabetes mellitus. Psychoneuroendocrinology, 34, 815–821.
Brundel, M., van den Heuvel, M., de Bresser, J., Kappelle, L. J., & Biessels, G. J. (2010). Cerebral cortical thickness in patients with type 2 diabetes. Journal of the Neurological Sciences, 299, 126–130.
Chen, Z., Li, L., Sun, J., & Ma, L. (2012). Mapping the brain in type II diabetes: voxel-based morphometry using DARTEL. European Journal of Radiology, 81, 1870–1876.
Cole, J., Costafreda, S. G., McGuffin, P., & Fu, C. H. Y. (2011). Hippocampal atrophy in first episode depression: a meta-analysis of magnetic resonance imaging studies. Journal of Affective Disorders, 134, 483–487.
Colman, I., Jones, P. B., Kuh, D., Weeks, M., Naicker, K., Richards, M., & Croudace, T. J. (2014). Early development, stress and depression across the life course: pathways to depression in a national British birth cohort. Psychological Medicine, 44(13), 2845–54.
Costafreda, S. G. (2012). Parametric coordinate-based meta-analysis: valid effect size meta-analysis of studies with differing statistical thresholds. Journal of Neuroscience Methods, 210, 291–300.
de Bresser, J., Tiehuis, A. M., van den Berg, E., Reijmer, Y. D., Jongen, C., Kappelle, L. J., et al. (2010). Progression of cerebral atrophy and white matter hypersensitivites in patients with type 2 diabetes. Diabetes Care, 33, 1309–1314.
de Jong, L. W., van der Hiele, K., Veer, I. M., Houwing, J. J., Westendorp, R. G., Bollen, E. L., et al. (2008). Strongly reduced volumes of putamen and thalamus in Alzheimer’s disease: an MRI study. Brain, 131, 3277–3285.
den Heijer, T., Vermeer, S. E., van Dijk, E. J., Prins, N. D., Koudstaal, P. J., Hofman, A., et al. (2003). Type 2 diabetes and atrophy of medial temporal lobe structures on brain MRI. Diabetologia, 46, 1604–1610.
Deng, Y., Li, B., Liu, Y., Iqbal, K., Grundke-Iqbal, I., & Gong, C. X. (2009). Dysregulation of insulin signalling, glucose transporters, O-GlcNAcylation, and phosphorylation of tau and neurofilaments in the brain: implication for Alzheimer’s disease. American Journal of Pathology, 175, 2089–2098.
DerSimonian, R., & Laird, N. (1986). Meta-analysis in clinical trials. Controlled Clinical Trials, 7(3), 177–188.
Diabetes Control and Complications Trial/Epidemiology of Diabetes Interventions and Complications Study Research Group, Jacobson, A. M., Musen, G., Ryan, C. M., Silvers, N., Cleary, P., et al. (2007). Long-term effect of diabetes and its treatment on cognitive function. New England Journal of Medicine, 356, 1842–1852.
Dowlati, Y., Herrmann, N., Swardfager, W., Liu, H., Sham, L., Reim, E. K., et al. (2010). A meta-analysis of cytokines in major depression. Biological Psychiatry, 67, 446–457.
Egan, M. F., Kojima, M., Callicott, J. H., Goldberg, T. E., & Kolachana, B. S. (2003). The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell, 2003(112), 257–69.
Elder, G. A., De Gasperi, R., & Gama Sosa, M. A. (2006). Research update: neurogenesis in adult brain and neuropsychiatric disorders. Mount Sinai Journal of Medicine, 73, 931–40.
Espeland, M. A., Bryan, R. N., Goveas, J. S., Robinson, J. G., Siddiqui, M. S., Liu, S., et al. (2013). Influence of type 2 diabetes on brain volumes and changes in brain volumes. Diabetes Care, 36, 90–97.
Frodl, T., Carballedo, A., Hughes, M. M., Saleh, K., Fagan, A., Skokauskas, N., et al. (2012). Reduced expression of glucocorticoid-inducible genes GILZ and SGK-1: high IL-6 levels are associated with reduced hippocampal volumes in major depressive disorder. Translational Psychiatry, 2, e88.
Fu, C. H. Y., Steiner, H., & Costafreda, S. G. (2013). Predictive neural biomarkers of clinical response in depression: a meta-analysis of functional and structural neuroimaging studies of pharmacological and psychological therapies. Neurobiology of Disease, 52, 75–83.
Fujinami, A., Ohta, K., Obayashi, H., Fukui, M., Hasegawa, G., & Nakamura, N. (2008). Serum brain-derived neurotrophic factor in patients with type 2 diabetes mellitus: Relationship to glucose metabolism and biomarkers of insulin resistance. Clinical Biochemistry, 41(10–11), 812–7.
Gaudieri, P. A., Chen, R., Greer, T. F., & Holmes, C. S. (2008). Cognitive function in children with type 1 diabetes: a meta-analysis. Diabetes Care, 31, 1892–1897.
Gold, S. M., Dziobek, I., Sweat, V., Tirsi, A., Rogers, K., Bruehl, H., et al. (2007). Hippocampal damage and memory impairments as possible early brain complications of type 2 diabetes. Diabetologia, 50, 711–719.
Hayashi, K., Kurioka, S., Yamaguchi, T., Morita, M., Kanazawa, I., Takase, H., et al. (2011). Association of cognitive dysfunction with hippocampal atrophy in elderly Japanese people with type 2 diabetes. Diabetes Research and Clinical Practice, 94, 180–185.
Hempel, R., Onopa, R., & Convit, A. (2012). Type 2 diabetes affects hippocampus volume differentially in men and women. Diabetes/Metabolism Research and Reviews, 28, 76–83.
Herrero, M. T., Barcia, C., & Navarro, J. M. (2002). Functional anatomy of thalamus and basal ganglia. Child’s Nervous System, 18, 386–404.
Hershey, T., Perantie, D. C., Wu, J., Weaver, P. M., Black, K. J., et al. (2010). Hippocampal volumes in youth with type 1 diabetes. Diabetes, 59, 236–241.
Higgins, J., Thompson, S., Deeks, J., & Altman, D. (2003). Measuring inconsistency in meta-analysis. BMJ, 327, 557–560.
Jongen, C., van der Grond, J., Kappelle, L. J., Biessels, G. J., Viergever, M. A., & Pluim, J. P. (2007). Automated measurement of brain and white matter lesion volume in type 2 diabetes mellitus. Diabetologia, 50, 1509–1516.
Kamiyama, K., Wada, A., Sugihara, M., Kurioka, S., Hayashi, K., Hayashi, T., et al. (2010). Potential hippocampal region atrophy in diabetes mellitus type 2: a voxel-based morphometry VSRAD study. Japanese Journal of Radiology, 28, 266–272.
Karege, F., Perret, G., Bondolfi, G., Schwald, M., & Bertschy, G. (2002). Decreased serum brain-derived neurotrophic factor levels in major depressed patients. Psychiatry Research, 109, 143–8.
Katon, W., Lyles, C. R., Parker, M. M., Karter, A. J., Huang, E. S., & Whitmer, R. A. (2012). Association of depression with increased risk of dementia in patients with type 2 diabetes. Archives of General Psychiatry, 69, 410–417.
Klunk, W. E., Price, J. C., Mathis, C. A., Tsopelas, N. D., Lopresti, B. J., Ziolko, S. K., et al. (2007). Amyloid deposition begins in the striatum of presenilin-1 mutation carriers from two unrelated pedigrees. The Journal of Neuroscience, 27, 6174–84.
Korczak, D. J., Pereira, S., Koulajian, K., Matejcek, A., & Giacca, A. (2011). Type 1 diabetes mellitus and major depressive disorder: evidence for a biological link. Diabetologia, 54(10), 2483–93.
Korf, E. S., White, L. R., Scheltens, P., & Launer, L. J. (2006). Brain aging in very old men with type 2 diabetes. Diabetes Care, 29, 2268–2274.
Kumar, A., Haroon, E., Darwin, C., Pham, D., Ajilore, O., Rodriguez, G., & Mintz, J. (2008). Gray matter prefrontal changes in type 2 diabetes detected using MRI. Journal of Magnetic Resonance Imaging, 27, 14–19.
Laake, J.P., Stahl, D., Amiel, S.A., Petrak, F., Sherwood, R.A., Pickup, J.C., & Ismail, K. (2014). The association between depressive symptoms and systemic inflammation in people with type 2 diabetes: findings from the south london diabetes study. Diabetes Care, May 19. pii: DC_132522. (Epub ahead of print).
Last, D., Alsop, D. C., Abduljalil, A. M., Marquis, R. P., de Bazelaire, C., Hu, K., et al. (2007). Global and regional effects of type 2 diabetes on brain tissue volumes and cerebral vasoreactivity. Diabetes Care, 30(5), 1193–9.
Liu, F., Shi, J., Tanimukai, H., Gu, J., Gu, J., Grundke-Iqbal, I., et al. (2009). Reduced O-GlcNAcylation links lower brain glucose metabolism and tau phosphorylation in Alzheimer’s disease. Brain, 132, 1820–1832.
Lobnig, B. M., Krömeke, O., Optenhostert-Porst, C., & Wolf, O. T. (2005). Hippocampal volume and cognitive performance in long-standing type 1 diabetic patients without macrovascular complications. Diabetic Medicine, 23, 32–39.
Lu, F. P., Lin, K. P., & Kuo, H. K. (2009). Diabetes and the risk of multi-system aging phenotypes: a systematic review and meta-analysis. PLoS One, 4(1), e4144.
Mangia, S., Tesfaye, N., De Martino, F., Kumar, A. F., Kollasch, P., Moheet, A. A., et al. (2012). Hypoglycemia-induced increases in thalamic cerebral blood flow are blunted in subjects with type 1 diabetes and hypoglycemia unawareness. Journal of Cerebral Blood Flow & Metabolism, 32, 2084–2090.
Manor, B., Newton, E., Abduljalil, A., & Novak, V. (2012). The relationship between brain volume and walking outcomes in older adults with and without diabetic peripheral neuropathy. Diabetes Care, 35, 1907–1912.
Mezuk, B., Eaton, W. W., Albrecht, S., & Golden, S. H. (2008). Depression and type 2 diabetes over the lifespan: a meta-analysis. Diabetes Care, 31, 2383–2390.
Musen, G., Lyoo, I. K., Sparks, C. R., Weinger, K., Hwang, J., Ryan, C. M., et al. (2006). Effects of type 1 diabetes on gray matter density as measured by voxel-based morphometry. Diabetes, 55, 326–333.
Musen, G., Jacobson, A. M., Bolo, N. R., Simonson, D. C., Shenton, M. E., McCartney, R. L., et al. (2012). Resting-state brain functional connectivity is altered in type 2 diabetes. Diabetes, 62, 2375–2379.
Naguib, J. M., Kulinskaya, E., Lomax, C. L., & Garralda, M. E. (2009). Neuro-cognitive performance in children with type 1 diabetes: a meta analysis. Journal of Pediatric Psychology, 34, 271–282.
Necula, M., & Kuret, J. (2004). Pseudophosphorylation and glycation of tau protein enhance but do not trigger fibrillization in vitro. The Journal of Biological Chemistry, 279, 49694–49703.
Northam, E. A., Rankins, D., Lin, A., Wellard, R. M., Pell, G. S., Finch, S. J., et al. (2009). Central nervous system function in youth with type 1 diabetes 12 years after disease onset. Diabetes Care, 32, 445–450.
Novak, V., Zhao, P., Manor, B., Sejdic, E., Alsop, D., Abduljalil, A., et al. (2011). Adhesion molecules, altered vasoreactivity, and brain atrophy in type 2 diabetes. Diabetes Care, 34, 2438–2411.
Ott, A., Stolk, R. P., van Harskamp, F., Pols, H. A., Hofman, A., Breteler, M. M. (1999). Diabetes mellitus and the risk of dementia: The Rotterdam Study. Neurology, 53(9), 1937–42.
Oxford centre for functional magnetic resonance imaging of the brain. (2003). Oxford thalamic connectivity atlas: www2.fmrib.ox.ac.uk/connect.
Perantie, D. C., Wu, J., Koller, J. M., Lim, A., Warren, S. L., Black, K. J., et al. (2007). Regional brain volume differences associated with hyperglycemia and severe hypoglycemia in youth with type 1 diabetes. Diabetes Care, 30, 2331–2337.
Perantie, D. C., Koller, J. M., Weaver, P. M., Lugar, H. M., Black, K. J., White, N. H., et al. (2011). Prospectively determined impact of type 1 diabetes on brain volume during development. Diabetes, 60, 3006–3014.
Pickup, J. C., & Crook, M. A. (1998). Is type II diabetes a disease of the innate immune system? Diabetologia, 41, 1241–1248.
Sarac, K., Akinci, A., Alkan, A., Aslan, M., Baysal, T., & Ozcan, C. (2007). Brain metabolite changes on proton magnetic resonance spectroscopy in children with poorly controlled type 1 diabetes. Neuroradiology, 47, 562–565.
Scott, K. M., Von Korff, M., Angermeyer, M. C., Benjet, C., Bruffaerts, R., de Girolamo, G., et al. (2011). Association of childhood adversities and early-onset mental disorders with adult-onset chronic physical conditions. Archives of General Psychiatry, 68(8), 838–44.
Sima, A. A., Zhang, W., Kreipke, C. W., Rafols, J. A., & Hoffman, W. H. (2009). Inflammation in diabetic encephalopathy is prevented by C-peptide. The Review of Diabetic Studies, 6, 37–42.
Stamler, J., Vaccaro, O., Neaton, J. D., & Wentworth, D. (1993). Diabetes, other risk factors, and 12-yr cardiovascular mortality for men screened in the Multiple Risk Factor Intervention Trial. Diabetes Care, 16(2), 434–44.
Stetler, C., & Miller, G. E. (2011). Depression and hypothalamic-pituitary-adrenal activation: a quantitative summary of four decades of research. Psychosomatic Medicine, 73, 114–26.
Teicher, M. H., Andersen, S. L., Polcari, A., Anderson, C. M., Navalta, C. P., & Kim, D. M. (2003). The neurobiological consequences of early stress and childhood maltreatment. Neuroscience and Biobehavioral Reviews, 27(1–2), 33–44.
Tesfomariam, B., Brown, M. L., & Cohen, R. A. (1991). Elevated glucose impairs endothelium-dependent relaxation by activating protein kinase C. The Journal of Clinical Investigation, 87, 1643–1648.
Tiehuis, A. M., van der Graaf, Y., Visseren, F. L., Vincken, K. L., Biessels, G. J., Appelman, A. P., et al. (2008). Diabetes increases atrophy and vascular lesions on brain MRI in patients with symptomatic arterial disease. Stroke, 39, 1600–1603.
Twist, K., Stahl, D., Amiel, S. A., Thomas, S., Winkley, K., & Ismail, K. (2013). Comparison of depressive symptoms in type 2 diabetes using a two-stage survey design. Psychosomatic Medicine, 75(8), 791–7.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix, N., Mazoyer, B., & Joliot, M. (2002). Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. NeuroImage, 15(1), 273–89.
van Elderen, S. G., Brandts, A., van der Grond, J., Westenberg, J. J., Kroft, L. J., van Buchem, M. A., et al. (2011). Cerebral perfusion and aortic stiffness are independent predictors of white matter brain atrophy in type 1 diabetic patients assessed with magnetic resonance imaging. Diabetes Care, 34, 459–463.
Videbech, P., & Ravnkilde, B. (2004). Hippocampal volume and depression: a meta-analysis of MRI studies. American Journal of Psychiatry, 161(11), 1957–1966.
Viechtbauer, W. (2010). Conducting meta-analyses in R with the metafor package. Journal of Statistical Software, 36(3), 1–48.
Watari, K., Elderkin-Thompson, V., Ajilore, O., Haroon, E., Darwin, C., et al. (2008). Neuroanatomical correlates of executive functioning in depressed adults with type 2 diabetes. Journal of Clinical and Experimental Neuropsychology, 30(4), 389–397.
Wessels, A. M., Simsek, S., Remijnse, P. L., Veltman, D. J., Biessels, G. J., Barkhof, F., et al. (2006). Voxel-based morphometry demonstrates reduced grey matter density on brain MRI in patients with diabetic retinopathy. Diabetologia, 49, 2474–2480.
Wessels, A. M., Rombouts, S. A., Remijnse, P. L., Boom, Y., Scheltens, P., Barkhof, F., et al. (2007). Cognitive performance in type 1 diabetes patients is associated with cerebral white matter volume. Diabetologia, 50, 1763–1769.
Williams, S. B., Goldfine, A. B., Timimi, F. K., Ting, H. H., Roddy, M. A., Simonson, D. C., & Creager, M. A. (1998). Acute hyperglycemia attenuates endothelium-dependent vasodilation in humans in vivo. Circulation, 97, 1695–1701.
Yau, P. L., Javier, D. C., Ryan, C. M., Tsui, W. H., Ardekani, B. A., Ten, S., et al. (2010). Preliminary evidence for brain complications in obese adolescents with type 2 diabetes mellitus. Diabetologia, 58, 2298–2306.
Conflict of interest
Calum D. Moulton, Sergi G. Costafreda, Paul Horton, Khalida Ismail, and Cynthia H. Y. Fu declare that they have no conflicts of interest.
Informed Consent
All procedures followed were in accordance with the ethical standards of the responsible committee on human experimentation (institutional and national) and with the Helsinki Declaration of 1975, and the applicable revisions at the time of the investigation. Informed consent was obtained from all patients for being included in the study.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Fig. 1
Forest plot of hippocampal volume in T1DM studies. Non-significant reduction found in T1DM compared to controls (DOCX 54 kb)
Supplementary Fig. 2
Forest plots of total brain volume. Non-significant reduction found in T1DM compared to controls (left) and significant reduction in T2DM compared to controls (right) (DOCX 108 kb)
Supplementary Fig. 3
Forest plots of total grey matter volume. Non-significant reduction found in T1DM compared to controls (left) and significant reduction in T2DM compared to controls (right) (DOCX 114 kb)
Supplementary Fig. 4
Forest plot of total frontal volume. Non-significant reduction found in T2DM compared to controls (DOCX 65 kb)
Supplementary Table 1
Potential confounding variables for T1DM studies (DOCX 15 kb)
Supplementary Table 2
Potential confounding variables for T2DM studies (DOCX 17 kb)
Supplementary Table 3
Volumetric meta-analysis results in T1DM (DOCX 13 kb)
Rights and permissions
About this article

Cite this article
Moulton, C.D., Costafreda, S.G., Horton, P. et al. Meta-analyses of structural regional cerebral effects in type 1 and type 2 diabetes. Brain Imaging and Behavior 9, 651–662 (2015). https://doi.org/10.1007/s11682-014-9348-2
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11682-014-9348-2