
Abstract
Although forests play important roles in the hydrological cycle, there is little information that relates the water retention capacity of litter in areas under passive restoration, especially in Cerrado savannas. This study relates litter levels to water holding capacity and effective water retention among forest fragments under different passive restoration stages: 46, 11, and 8 years to better understand litter hydrological functions in the Cerrado. Water retention capacity and effective water retention capacity of litters (unstructured materials, branches and leaves) in the field were monitored on a monthly basis. Total litter accumulation at 46 years was significantly higher than that of the other succession stages. Unstructured litter mass was significantly higher than that of leaves and branches. The 46-year stage had the highest water holding capacity in the leaf fraction, followed by unstructured material and branches. Although the water holding capacity was lower in the oldest restoration, this site showed the highest efficiency under field conditions. The process was quickly reestablished, as the 11-year restoration showed results closer to that for the 46-year stage in comparison to the area at 8 years. Thus, passive restoration plays a key role in soil water maintenance due to the influence of litter in Cerrado savannas. Deforestation and the imminent need of restoring degraded sites, highlight the need for further studies focused on better understanding of the process of forest restoration and its temporal effect on soil water recovery dynamics.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Rainfall is partitioned into different paths when it reaches a forest canopy and, to contribute to water balance in ecosystems, these paths are interception, throughfall and stemflow (Barbier et al. 2009; Liu et al. 2014; Carvalho et al. 2016; Bessi et al. 2018a, b; Friesen 2020). Throughfall and stemflow are the rainfall partition reaching the soil and interact with the litter, which plays a key role in rainfall interception and functions in the nutrient cycling and energy transmission within ecosystems (Acharya et al. 2017; Pang and Bao 2020). Litter is the organic biomass layer on the forest floor, formed by materials such as leaves, branches, flowers, fruits, seeds and animal waste. The composition of litter changes, varying on the ecosystem, age and density, as well as on abiotic factors and site degradation (Piovesan et al. 2012; Santos et al. 2017).
The litter layer is acknowledged as a forest ecosystem bioindicator, used to assess the quality of the environment, and it explains why it is often an object of water resource management projects (Dunkerley 2015; Ilek et al. 2015). Litter plays a key role in soil and water conservation (Castillo et al. 2003; Wang et al. 2013) and in other hydrological processes at a watershed management scale.
The interaction among rainfall, forest canopy, and litter is an interest of forest hydrologists who investigate rainfall partitioning and disposition (Dunkerley 2015). Litter water holding is an important hydrological indicator (Dickow et al. 2012; Acharya et al. 2017; Pinto and Negreiros 2018; Pang and Bao 2020). Litter acts as a spongeto intercept rainwater and protect the soil from direct impact, and delays or eliminates surface water flow (Giácomo et al. 2017), and mitigates soil water loss due to evaporation.
Litter structure, composition, amount, and properties change with agricultural systems and with forest management, be it for preservation or for timber that lead to changes in forest hydrological functions. Litter changes directional flows, humidity and soil water evaporation (Matthews 2005), so the amount of litter and its hydrological properties can change infiltration rates and affect watersheds. Finally, litter can affect both water quality and volume (Zhang and Shangguan 2016; Chen et al. 2018; Zagyvai-Kiss et al. 2019).
A lack or shortage of litter can affect water balance on watersheds and can lead to water loss two-four times higher than the mass of the ecosystem (Alberts and Neibling 1994). Litter removal results in increased flow on watersheds, mainly during large flood events (Gomyo and Kuraji 2016). Recent studies have added evidence about the effect of litter on water budgets in Eucalyptus crops, semi-arid scrublands and on forests having a significant litter layer (Acharya et al. 2017; Chen et al. 2018). These studies are essential to managing forest production systems and soil, water and nutrition resource sustainability.
Studies on natural forest restoration and on knowledge of soil restoration dynamics and the conservaton of hydrological processes remain scarce. Litter yield dynamics, its structure and association with the hydrological properties of forest ecosystems, and its relevance for water conservation management have not been widely evaluated (Acharya et al. 2017; Zagyvai-Kiss et al. 2019). Although there are studies of litter yields in different environments, there is not sufficient understanding of water holding capacity of litter (the maximum amount retained by litter) in ecosystems under field conditions. Research on litter hydrological functions in the water balance of forest ecosystems, either under stable conditions or restoration process, is extremely important (Sun et al. 2018; Zhou et al. 2018).
The aim of this study was to relate litter quantity to its water holding capacity in forest fragments representing different passive restoration stages in the Cerrado savanna. The data addressed the following questions: How does litter’s water holding capacity affect Cerrado passive restoration? How is litter’s effective water maintained under field conditions? We hypothesized that: litter quantity, its water holding capacity and its effective water holding capacity increase due to passive restoration time. Testing this hypothesis will improve understanding of litter hydrological functions in Cerrado savanna ecosystems subjected to passive restoration processes.
Materials and methods
2. 1 Study area
The study was carried out in the Águas Perenes Forest, a Private Reserve of the Natural Patrimony (PRNP) located in Lagoa Seca microbasin, Brotas County–São Paulo State (22°11.754′ S and 48°6.523′ W). This forest is the water recharge area of the Guarani Aquifer. In 2011, it was recognized by the Forest Stewardship Council as a High Conservation Value Forest, since it provides essential environmental services such as watershed protection. The PRNP covers approximately 809.8 ha, and is considered as secondary vegetation of Cerrado (trees cover more than 30% of the ground, with grass, forming an open savanna), and Cerradão (closed woodland savanna without grass cover) (Ratter et al. 1997; Oliveira-Filho and Ratter 2002; Durigan et al. 2012). The climate is of the Cwa type (Köppen/Geiger’s classification)―mean annual precipitation reaches 1337 mm and mean annual temperature is 20 °C (Pereira et al. 2021). Quartzarenic neosol soils prevail (Santos et al. 2018).
2.2 Sampling design
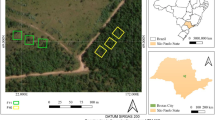
The study was carried out in three areas undergoing different degrees of passive restoration: fragment A (FA)–46-year restoration (tree species in Table S1), B (FB)–11-year restoration (tree species in Table S2), and C (FC)–8-year restoration mostly grass (Brachiaria spp.) (Table S3). Fragments were approximately 80 m apart (Fig. 1). The area was used to grow Eucalyptus spp. and after harvesting, the Eucalyptus was not renewed and the site was left for passive restoration. Table 1 shows features of the assessed fragments.

Plot locations in each fragment: Fragments undergoing passive restoration for 46 years (FA), 11 years (FB) and 8 years (FC). Águas Perenes Forest, Brotas County, Brazil
The study was carried out in three 400 m2 plots 10 m apart in each fragment with inventory based on the fixed area method (Bessi et al. 2018a). A total of 1200 m2 were sampled in each forest fragment. Total precipitation, throughfall, stemflow, and litter water holding capacity were measured on a monthly basis from May/2018 to April/2019. Samples were simultaneously collected in the three different plots on the same watershed hillside. The fragments were subjected to the same land use history to make the comparisons as accurate as possible.
2.3 Data collection
2.3 1 Open precipitation, throughfall and stemflow
Polyethylene gauges were used to monitor precipitation and throughfall, (78.5 cm catchment and 20 cm high) for a capacity of 1.57 L. Precipitation was measured with a rain gauge installed near the plot (maximum distance 30 m). A funnel on the top of the rain gauges prevented leaves, insects and small animals to enter to avoid interrupting water flow and to minimize evaporation. Precipitation was calculated by the following equation:
where, P is precipitation (mm), V rain gauge volume (mL) and A rain gauge catchment area (cm2).
Throughfall collectors were placed 1.2 m above the ground for 20 rain gauges distributed throughout the plots (Eq. 2):
where, Tf is throughfall (mm), V volume of each rain gauge (mL), A catchment area of each gauge (cm2), and ni the number of gauges.
Stemflow was measured on trees with DBH (diameter breast height at 1.3 m) > 5 cm. The stemflow was measurement using a polyurethane gutter system based on the methodology (Likens and Eaton 1970) and fixed at 1.3 m from the ground among the trees. The water that drained through the stem surface was directed by a 5/8″ hose to 20 L collectors. The stemflow yield (SF, mm) was calculated considering the ratio between the volume accumulated in the collectors (V, L) and canopy area (CA, m).
Precipitation, throughfall and stemflow samples were measured in the field with 1-L measuring cylinders (0.5-L graduation) or in 20-L graduated buckets (0.5-L graduation). Net precipitation per plot was found by summing throughfall and stemflow.
2.3 2 Forest litter samples
The litter layer on the soil surface was collected on a monthly basis using a 50 cm × 50 cm gauge partitioned into four quadrants. Collection was random and only material in one quadrant was collected at a time. This resulted in 10 collections in each plot, for a total of three composed samples per fragment/month.
The soil was sieved and removed from the litter samples, and the fresh mass of the litter was measured in the field. The samples were stored in plastic bags, and the litter screened for the following fractions: branches, leaves, reproductive material and unstructured material (pieces of bark, fruit, reproductive parts, seeds). The various fractions were subsequently dried in a forced air-circulation oven at 70 °C until constant weight. Monthly and annual litter accumulations were estimated by summing the fractions. Fragment C (FC) was from a young restoration stage and had only two trees; its litter was essentially composed of herbaceous Brachiaria spp., which made litter fractioning impossible.
2.3 3 Water holding capacity (WHC)
The WHC was determined under laboratory conditions. The methodology for WHC used in the current study was developed by Blow (1985). Each litter fraction was rehydrated by immersion in water for 90 min. The fractions were deposited on sieves, drained for 30 min for humid mass (HM) determination on a precision scale, dried again at 70 °C, until constant weight. They were weighed to get their dry mass (DM). WHC was found through:
where WHC is water holding capacity (%), HM is the humid mass (g), and DM is the dry mass (g).
The relationship between fresh mass and litter water holding capacity was determined to investigate the effective water holding capacity (EHC, %) for each fragment under field conditions. This was determined by Eq. 4:
where, FM is fresh mass (g), and HM is humid mass (g).
2.4 Data analyses
Litter accumulation (total and component parts), water holding capacity (total and component parts) and effective litter water holding capacity of all plots per fragment, were subjected to homoscedasticity tests of variance through the Bartlett test (“bartlett.test” function, “stats” package), and to normality analysis through Lilliefors (Kolmogorov–Smirnov) (“lillie.test” function, “nortest” package) in R software (R Development Core Team 2018) for statistical analysis. Analysis of variance (“aov” function, “stats” package) was applied to normal data through Tukey test (“TukeyHSD” function, “stats” package) at 5% probability level. Data that did not meet ANOVA assumptions were subjected to non-parametric Kruskal–Wallis test (“kruskal.test” function, “stats” package). The Spearman test (“pspearman.test” function, “pspearman” package) was adopted to determine whether there were significant correlations between litter accumulation and precipitation or net precipitation, as also between effective water holding capacity and net precipitation.
Results
3.1 Total and net precipitation
Accumulated precipitation throughout the research period was 936.7 mm, i.e., 6.3% higher than the precipitation recorded in the same period (877.8 mm) by the official meteorological station of the National Meteorology Institute (INMET 2020). The comparison between precipitation in the study area and the use of averaged data over 1981 − 2010 (climatological normal for the latest global standard normal period) showed an atypical year. Mean temperature was 21.6 °C, 5% higher than that recorded for normal (20.6 °C). The highest mean temperature was recorded in December (24.7% above normal) and the lowest in July (18.3%) (Fig. S1a in the Supplemental Materials). Net precipitation (NP) in FA (46 years) and FB (11 years) was 894.6 and 824.1 mm a−1, respectively, 4.5% and 12% lower than the precipitation without tree cover. FC (8 years) was characterized as an open field; its NP was similar to the accumulated precipitation (Figs. S1b, S1c and S1d in the Supplemental Materials).
3.2 Litter accumulation and composition
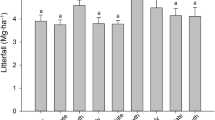
Total litter ranged from 5.5 t ha−1 a−1 in FA, 3.5 t ha−1 a−1 in FB and 3.4 t ha−1 a−1 in FC. The fractions showed significant differences between fragments (Fig. 2a) and had smaller variations in FA (54.1% unstructured, 25.9% leaves, 21.9% branches). The unstructured fraction in FB alone was 75.2% of the total litter, whereas it was 14.0% of leaves and 12.8% of branches. The ANOVA test at 95% confidence level showed that the unstructured fractions did not differ between fragments unlike the leaves and branch fractions (Fig. 2b, c, d).

Monthly mean litter accumulation (t ha−1) for a FA, FB and FC; b mean litter partition in unstructured material, c branches and d leaves from FA and FB. Different lower-case letters mean significant differences (p < 0.05)
Mean monthly litter accumulation on the FA site (Fig. S2 in the Supplemental Materials) is statistically different between months and between fractions. May had the highest litter accumulation in FA (0.77 t ha−1) when the lowest temperature was also recorded (18.3 °C). FB and FC showed statistical homogeneity between months. The highest litter accumulation in these ecological fragments was in March (0.53 t ha−1) and February (0.55 t ha−1) (Figs. S2b and S2c in the Supplemental Materials), after the largest net precipitation recorded at this time of the year. The highest temperatures were between December and March (23.7 °C on average).
Litter production was influenced by precipitation rates. Pearson correlation showed that this was more correlated to net precipitation in all fragments than to open precipitation (Table 2), although there was negative correlation in FA. None of the fragments showed statistical significance (p > 0.05).
3.3 Litter water holding capacity
Total litter water holding capacity was different between passive restoration sites. The highest average r water holding capacity was for the FC fragment (394%), followed by FA (224.6%) and FB (221.4%) (Fig. 3). WHC was statistically different between averages for the fragments.

Median litter water holding capacity (WHC, %) in FA, FB, and FC
The highest WHC values were in different months in each area. FA had the highest WHC in August (436.4%), FB the highest in June (345.9%) and FC, in November and December (847.7%). FC values were 3 − 4 times higher than those for FA and FB in the same period (Fig. S3 in the Supplemental Materials). These values were not records of highest litter accumulation but ones for net precipitation. The highest WHC for FA was after two consecutive months without any precipitation. The outcome for FB was related to the first month without precipitation. The highest WHC for FC was after a precipitation increase although it was followed by an 80% decrease.
The FA fraction assessment was possible by the following WHC order: unstructured material (344.1%) > leaves (307.0%) > branches (221.7%) (Fig. 4a). FB showed a different behavior, both in order and magnitude (values lower than those recorded for FA): leaves (290.4%) > unstructured material (241.6%) > branches (131.9%) (Fig. 4b). There was overall progress in restoration between FA and FB; litter had higher water holding capacity although its components recorded different WHC order. FA and FC were statistically different among months (p < 0.05). WHC in FA and FB was statistically significant different among averages for the fractions.

Average litter water holding capacity (WHC, %) branches, leaves and unstructured material in a FA and b FB
3.4 Effective water holding capacity of litter under field conditions
The analysis of the mean annual litter WHC which is correlated fresh to humid mass, showed that EHC increased as passive restoration evolved. FA and FB were similar but differed from FC. This means that FA and FB held more water in their litter, although their WHC was lower compared to FC, which, in turn, recorded 45% capacity (Table 3).
The correlation of fresh and humid mass ratio to net precipitation showed that net precipitation became more significant and higher as the successional stage evolved: FA (83%) > FB (44%) > FC (22%) (Table 4).
Discussion
4.1 Litter accumulation and restoration stage
Litter accumulation in FA was close to that observed by other researchers in tropical environments having similar features: 5.15 t ha−1 a−1 in a forest fragment under restoration for 30 years (Mateus et al. 2013); 5.6 t ha−1 a−1 in Cerradão (Cianciaruso et al. 2006), and 4.99 t ha−1 a−1 in Cerrado (Ribeiro et al. 2017).
Litterfall was higher in FA than in FB or FC, and is explained by high tree density (Table 1) since this was linked to forest structure (Almeida et al. 2015) and to organic matter as shown by other researchers (Borém and Ramos 2002; Costa et al. 2010; Mateus et al. 2013). Paiva et al. (2015) attributed the significantly higher annual litter to forest phytophysiognomy (6.3 t ha−1 a−1) in comparison to the savanna (3.5 t ha−1 a−1), which the last had values closer to those for FB and FC, although Brachiaria species were predominant in FC.
Litter accumulation for FB and FC were statistically different from each other but similar to results for Capoeira (10 years – dominated by Clidemia urceolata and shrub-tree species) and Capoeirão (15 years – dominated by Clidemia urceolata and Cecropia pachystachya) (Mateus et al. 2013). However, they were different from records of disturbed pasture (6 years) (Mateus et al. 2013). This indicates the evolution in litter accumulation in FC and its restoration stage, which was the initial restoration stage, in comparison to FB.
The current study showed that litter accumulation varied within fragments and months. Variations due to temperature and precipitation were associated with plant strategy to control water loss due to transpiration during warm periods, e.g., leaf and branch abscission (Pereira et al. 2017). In the Amazon, the largest litter accumulation occurred in the warmest periods of the year due to leaf renewal by trees (Kim et al. 2012) due to better radiation use for photosynthesis, even with low soil water availability.
In contrast to the current study, several reports indicated that the leaf fraction corresponded to 60% or more of the total litter in different forest ecosystems. In the Cerradão and mesophilic forests, the litter is 71.6% leaves, + 11.1% branches, and 68.8% of leaves + 10.3% of branches, respectively (Giácomo et al. 2012), and 75.0% leaf fraction in a forest-savanna transition (Paiva et al. 2015).
The material classified as unstructured consisted of leaves at the decomposition stage, and fruit and seeds. This makes it possible to suggest that the high percentage of unstructured material was from a more active litter decomposition in FA, although it was more evident in FB. The litter fractioning analysis indicated that litter decomposition in FA and FB was evolving and providing subsidies for the maintenance of management practices in these ecosystems.
4.2 Water holding capacity, effective water holding capacity and passive restoration
Litter water holding capacity (WHC) depends on its composition of leaves, branches, unstructured material, fruit and seeds) and on the size distribution of its fractions, mainly in the decomposition stage since small particles have reduced contact surface and absorb more water (Melos et al. 2010). With Eucalyptus, the litter water holding capacity was reported as 249.6% (Santos et al. 2017) and 235.0% (Melos et al. 2010), 230.0% in disturbed and abandoned pastures, and 206.0% in natural forest fragments (Mateus et al. 2013). Their findings are similar to what was observed in this study: FC accounted for higher water holding capacities than FA and FB.
Less decomposed materials had lower WHC, especially evident in the branches, as observed by Mateus et al. (2013) and Santos et al. (2017). Branches have lower water absorption due to their woody nature and to the presence of lignin, attributes that favored surface flow which was also enhanced by their shape (Santos et al. 2017). These features were also addressed by Voigt and Wash (1976) who reported that absorption depends on more than one aspect, such as variation in precipitation and temperature.
The highest water holding capacity in FB was observed in the leaves rather than in the unstructured litter. This is because of the high presence of young leaves which had lower lignin content – this is also linked to grass species. In the capoeira, the values are close to 342.0% (Mateus et al. 2013). This was attributed to the amount of leaves belonging to the species Clidemia arceolata DC., which made forest restoration easier (Matsumoto 1999) and, in the initial restoration stages, provided developing species with leaves on site hold humidity.
It is important to stress that litter WHC only reflected the water holding condition in the ideal state. Water holding capacity was higher than the actual litter interception capacity and did not reflect litter interception under field conditions (Li et al. 2018). The correlations between fresh and saturated biomass reinforced litter efficiency in holding water due to passive restoration maturation under field conditions. So, although the water holding capacity was lower in the oldest restoration, this site showed the highest efficiency under field.
This difference between the laboratory and field water holding is due to litter interaction with environmental factors such as radiation, temperature, humidity and wind. As passive restoration matured, there was more canopy area available and less radiation inside the forest. Bessi et al. (2018b) found that light intensity inside the forest fragment decreased (FC > FB > FA), and when the basal area and forest density increased (FC < FB < FA).
Wind, humidity and temperature also play key roles in ecosystems and water cycles as they strongly affect evapotranspiration rates. Atmospheric evaporation demand increased as temperature increased, and increased evapotranspiration rates. There are several studies showing that evapotranspiration is affected by these factors (Tonello and Teixeira Filho 2011; Liuzzo et al. 2016; Anderson et al. 2017; Gaertner et al. 2019).
Although the FC site had the highest water holding capacity, evapotranspiration was lower. FC was more characterized as open field, which allowed higher radiation, higher temperatures and lower relative humidity. These factors promoted higher atmospheric pressure deficits and consequently, higher evapotranspiration rates. Wind was a constant in the FC since it was an open field. These factors resulted in lower water permanence and lower effective water holding capacity in litter at the initial restoration stage.
The current study focused on forest fragments under passive restoration that are of international interest for water resource conservation. Litter hydrology studies, such as this one, are important to support discussions and financially support programs on forest hydrology. However, there is an immediate need for more research on native forests and to relate the effect of restoring degraded areas on soil water recovery.
Conclusion
Litter accumulated as the passive restoration process matured, and its decomposition rate varied with each environment. Litter water holding capacity was three to four times higher in the forest fragments at the initial restoration stage but did not reflect water holding capacity under field conditions. The litter effective water holding capacity increased from the youngest to the oldest forest fragment under passive restoration. The most mature fragment was the most efficient in this process, as well as showing better correlation to net precipitation. The hydrological functions of litter were gradually reestablished depending on the restoration time, as the 11-year restoration area showed results closer to the ones for the 46-year restoration period, compared to the 8-year restoration area. Thus, passive restoration plays a key role in soil water maintenance due to the influence of litter on the Cerrado savanna.
References
Acharya BS, Stebler E, Zou CB (2017) Monitoring litter interception of rainfall using leaf wetness sensor under controlled and field conditions. Hydrol Process 31:240–249. https://doi.org/10.1002/hyp.11047
Alberts EE, Neibling WH (1994) Influence of crop residues on water erosion. In: Unger PW (ed) Managing agricultural residues. Lewis Publ, Ann Arbor, MI, pp 19–39
Almeida EJ, Luizão F, Rodrigues DJ (2015) Litterfall production in intact and selectively logged forests in southern of Amazônia as a function of basal area of vegetation and plant density. Acta Amaz 45:157–166. https://doi.org/10.1590/1809-4392201402543
Anderson RG, Ferreira JFS, Jenkins DL, Dias NS, Suarez DL (2017) Incorporating field wind data to improve crop evapotranspiration parameterization in heterogeneous regions. Irrig Sci 35:533–547. https://doi.org/10.1007/s00271-017-0560-x
Barbier S, Balandier P, Gosselin F (2009) Influence of several tree traits on rainfall partitioning in temperate and boreal forests: a review. Ann Sci 66:602–602. https://doi.org/10.1051/forest/2009041
Bessi D, Dias HCT, Tonello KC (2018a) Rainfall partitioning in fragments of cerrado vegetation at different stages of conduction of natural regeneration. Rev Árvore 42:1–11. https://doi.org/10.1590/1806-90882018000200015
Bessi D, Tanaka MO, da Costa LA, Tonello KC (2018b) Forest restoration and hydrological parameters effects on soil water conditions: A structural equation modelling approach. Rev Bras Recur Hidricos. https://doi.org/10.1590/2318-0331.231820180043
Blow FE (1985) Quantity and hydrologic characteristics of litter under upland oak forests in eastern Tennessee. J For 53:190–195
Borém RAT, Ramos DP (2002) Variação estacional e topográfica de nutrientes na serapilheira de um fragmento de mata Atlántica. Cerne 8:42–59
Carvalho APV, Dias HCT, Tonello KC, Paiva HN (2016) Net precipitation and recharge of groundwater in riacho fundo watershed. Felixlândia-MG Rev Arvore. https://doi.org/10.1590/0100-67622016000600001
Castillo VM, Gómez-Plaza A, Martínez-Mena M (2003) The role of antecedent soil water content in the runoff response of semiarid catchments: a simulation approach. J Hydrol 284:114–130. https://doi.org/10.1016/S0022-1694(03)00264-6
Chen S, Cao T, Tanaka N, Gao T, Zhu L, Zou CB (2018) Hydrological properties of litter layers in mixed forests in Mt. Qinling. China Iforest 11:243–250. https://doi.org/10.3832/ifor2535-011
Cianciaruso MV, Pires JSR, Delitti WBC, Silva EFLP (2006) Produção de serapilheira e decomposição do material foliar em um cerradão na Estação Ecológica de Jataí, município de Luiz Antônio, SP, Brasil. Acta Bot Brasilica 20:49–59. https://doi.org/10.1590/s0102-33062006000100006
Costa CCA, Camacho RGV, Macedo ID, Silva PCM (2010) Análise comparativa da produção de serapilheira em fragmentos arbóreos e arbustivos em área de caatinga na Flona de açu-rn. Rev Arvore 34:259–265. https://doi.org/10.1590/s0100-67622010000200008
Dickow KMC, Marques R, Pinto CB, Höfer H (2012) Litter production in different successional stages of a subtropical secondary rain forest, in Antonina, PR. Cerne 18:75–86. https://doi.org/10.1590/S0104-77602012000100010
Dunkerley D (2015) Percolation through leaf litter: what happens during rainfall events of varying intensity? J Hydrol 525:737–746. https://doi.org/10.1016/j.jhydrol.2015.04.039
Durigan G, Melo ACG, Brewer JS (2012) The root to shoot ratio of trees from open—and closed-canopy cerrado in south-eastern Brazil. Plant Ecol Divers 5:333–343. https://doi.org/10.1080/17550874.2012.691564
Friesen J (2020) Flow pathways of throughfall and stemflow through the subsurface. In: Van JT Stan, II, E Gutmann, J Friesen (eds) Precipitation partitioning by vegetation: a global synthesis. Springer, Cham, pp 215–228. https://doi.org/10.1007/978-3-030-29702-2_13
Gaertner BA, Zegre N, Warner T, Fernandez R, He Y, Merriam ER (2019) Climate, forest growing season, and evapotranspiration changes in the central Appalachian mountains, USA. Sci Total Environ 650:1371–1381. https://doi.org/10.1016/j.scitotenv.2018.09.129
Giácomo RG, Pereira MG, Machado DL (2012) Aporte e decomposição de serapilheira em áreas de cerradão e mata mesofítica na estação ecológica de Pirapitinga – MG. Ciência Florest 22:669–680
Giácomo RG, Alves MC, Camara R, Pereira MG, Arruda OG, Souto SN, Moraes MLT (2017) Litterfall and nutrient input in a degraded area. Floresta e Ambiente. https://doi.org/10.1590/2179-8087.002816
Gomyo M, Kuraji K (2016) Effect of the litter layer on runoff and evapotranspiration using the paired watershed method. J For Res 21:306–313. https://doi.org/10.1007/s10310-016-0542-5
Ilek A, Kucza J, Szostek M (2015) The effect of stand species composition on water storage capacity of the organic layers of forest soils. Eur J For Res 134:187–197. https://doi.org/10.1007/s10342-014-0842-2
INMET (2020) Estações e Dados: BDMAP—Dados Históricos. Estação: 83726 – São Carlos. https://www.inmet.gov.br. Accessed 20 Mar 2021
Kim Y, Knox RG, Longo M, Medvigy D, Hutyra LR, Pyle EH, Wofsy SC, Bras RL, Moorcroft PR (2012) Seasonal carbon dynamics and water fluxes in an amazon rainforest. Glob Chang Biol 18:1322–1334. https://doi.org/10.1111/j.1365-2486.2011.02629.x
Li W, Li Y, Long M, Li X (2018) Study on water-holding properties of litters in different types of forests of Yuntaishan Mountain area in Shibing County, Guizhou Province. IOP Conf Ser Mater Sci Eng. https://doi.org/10.1088/1757-899X/394/2/022057
Likens GE, Eaton JE (1970) A polyurethane stemflow collector for trees and shrubs. Ecology 51:938–939. https://doi.org/10.2307/1933996
Liu G, Du S, Peng S, Wang G (2014) Rainfall interception in two contrasting forest types in the mount Gongga area of Eastern Tibet, China. Hydrol Curr Res 4:1–6. https://doi.org/10.4172/2157-7587.1000161
Liuzzo L, Viola F, Noto LV (2016) Wind speed and temperature trends impacts on reference evapotranspiration in Southern Italy. Theor Appl Climatol 123:43–62. https://doi.org/10.1007/s00704-014-1342-5
Mateus FA, Miranda CC, Valcarcel R, Figueiredo PHA (2013) Estoque e capacidade de retenção hídrica da serrapilheira acumulada na restauração florestal de áreas perturbadas na mata Atlântica. Floresta e Ambient 20:336–343. https://doi.org/10.4322/floram.2013.024
Matsumoto K (1999) A família Melastomataceae Juss. nas formações campestres do município de Carrancas, Minas Gerais. Universidade Estadual de Campinas, 91. Dissertação (mestrado) - Universidade Estadual de Campinas, Instituto de Biologia, Campinas-SP. http://www.repositorio.unicamp.br/handle/REPOSIP/314840
Matthews S (2005) The water vapour conductance of Eucalyptus litter layers. Agric for Meteorol 135:73–81. https://doi.org/10.1016/j.agrformet.2005.10.004
Melos AR, Sato AM, Netto ALC (2010) Produção, estoque e retenção hídrica da serrapilheira em encosta sob plantio de híbridos de Eucalyptus urophylla e Eucalyptus grandis: Médio vale do rio paraíba do sul. Anu Do Inst Geocien 33:66–73
Oliveira-Filho A, Ratter J (2002) Vegetation physiognomies and woody flora of the Cerrado biome. In: Oliveira PSMR (ed) The cerrados of Brazil: ecology and natural history of a neotropical savanna, 1st edn. Columbia University, New York, pp 91–120
Paiva AO, Silva LCR, Haridasan M (2015) Productivity-efficiency tradeoffs in tropical gallery forest-savanna transitions: linking plant and soil processes through litter input and composition. Plant Ecol 216:775–787. https://doi.org/10.1007/s11258-015-0466-8
Pang X, Bao W (2020) Effect of substituting plantation species for native shrubs on the water-holding characteristics of the forest floor on the Eastern Tibetan Plateau. J Resour Ecol 2:217–224. https://doi.org/10.3969/j.issn.1674-764x.2011.03.004
Pereira DN, Martins WBR, Andrade VMS, Oliveira FA (2017) Influence of litter removal in the content of phosphorus and potassium in Eastern Amazon. Rev Bras Ciencias Agrar 12:380–385. https://doi.org/10.5039/agraria.v12i3a5458
Pereira LC, Balbinot L, Matus GN, Dias HCT, Tonello KC (2021) Aspects of forest restoration and hydrology: linking passive restoration and soil–water recovery in Brazilian Cerrado. J for Res. https://doi.org/10.1007/s11676-021-01301-3
Pinto WJ, Negreiros AB (2018) A serrapilheira como bioindicador de qualidade ambiental em fragmentos de Eucalyptus. Rev Cont 7:175–202
Piovesan G, Schumacher MV, Viera M, Lopes VG, Welter C (2012) Deposição de serapilheira em povoamento de Pinus. Pesquisa Agropecuária Tropical 42:206–211. https://doi.org/10.1590/s1983-40632012000200012
R Development Core Team (2018) R version 4.0.4. The foundation for statistical computing. http://www.r-project.org/. Accessed 23 Feb 2021
Ratter JA, Ribeiro JF, Bridgewater S (1997) The Brazilian cerrado vegetation and threats to its biodiversity. Ann Bot 80:223–230. https://doi.org/10.1006/anbo.1997.0469
Ribeiro FP, Bussinguer AP, Hodecker BER, Gatto A (2017) Conteúdo de nutrientes na serapilheira em três fisionomias do Cerrado do Distrito Federal. Pesqui Florest Bras 37:465–473. https://doi.org/10.4336/2017.pfb.37.92.1312
Santos AFA, Carneiro ACP, Martinez DT, Caldeira SF (2017) Capacidade de retenção hídrica do estoque de serapilheira de eucalipto. Floresta e Ambient 24:1–9. https://doi.org/10.1590/2179-8087.030315
Santos HG, Jacomine PKT, Anjos LHC, Oliveira VA, Lumbreras JF, Coelho MR, Almeida JA, Araujo Filho JC, Oliveira JB, Cunha TJF (2018) Sistema Brasileiro de Classificação de Solos. Brasília-DF: Embrapa. 356p. https://www.infoteca.cnptia.embrapa.br/handle/doc/1094003
Sun J, Yu X, Wang H, Jia G, Zhao Y, Tu Z, Deng W, Jia J, Chen J (2018) Effects of forest structure on hydrological processes in China. J Hydrol 561:187–199. https://doi.org/10.1016/j.jhydrol.2018.04.003
Tonello KC, Teixeira Filho J (2011) Environmental variables effects in ecophysiological behavior of two clones of Eucalyptus grandis × Eucalyptus urophylla: Field conditions. Sci for 39:419–431
Voigt VPJ, Walsh RPD (1976) Hidrologische prozesse in bodenstreu. Schriften Des Naturwissenschaftlichen Vereins Fur Schleswig-Holstei 46:35–54
Wang C, Zhao CY, Xu ZL, Wang Y, Peng HH (2013) Effect of vegetation on soil water retention and storage in a semi-arid alpine forest catchment. J Arid Land 5:207–219. https://doi.org/10.1007/s40333-013-0151-5
Zagyvai-Kiss KA, Kalicz P, Szilágyi J, Gribovszki Z (2019) On the specific water holding capacity of litter for three forest ecosystems in the eastern foothills of the Alps. Agric for Meteorol 278:107656. https://doi.org/10.1016/j.agrformet.2019.107656
Zhang Y, Shangguan Z (2016) The change of soil water storage in three land use types after 10 years on the Loess Plateau. CATENA 147:87–95. https://doi.org/10.1016/j.catena.2016.06.036
Zhou Q, Keith DM, Zhou X, Cui X, Wei X, Luo Y (2018) Comparing the water-holding characteristics of broadleaved, coniferous, and mixed forest litter layers in a Karst Region. Mt Res Dev 38:220–229. https://doi.org/10.1659/mrd-journal-d-17-00002.1
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: This work is supported by the International Paper Co. from Brazil (Process 23112.000670/2015-59), by Brazilian National Council for Scientific and Technological Development (CNPq) and in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001.
The online version is available athttp://www.springerlink.com.
Corresponding editor: Yu Lei.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Pereira, L.C., Balbinot, L., Lima, M.T. et al. Aspects of forest restoration and hydrology: the hydrological function of litter. J. For. Res. 33, 543–552 (2022). https://doi.org/10.1007/s11676-021-01365-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-021-01365-1