
Abstract
Zawita natural forest has recently has been subject to mass recreational activities during spring that have denuded large areas of the forest. It was thus essential to assess regeneration before designing optimizing strategies. To this end, we studied the overstory canopy and microhabitat conditions for recruitment of Pinus brutia Ten in 10 plots (20 × 25 m) on the southern aspects where the Zawita natural forest is still present. In total, 1540 regenerating P. brutia were recorded, 854 seedlings, 597 saplings, and 89 trees. Seedlings and saplings were more frequent beyond the canopy than under the canopy of the parent trees. Regeneration requirements differed between seedlings and saplings. The probability of the occurrence of seedlings was negatively correlated with increasing litter depth and increasing soil compaction. The density of saplings only showed a positive significant correlation with increasing slope. The nearest neighbor index showed a trend toward a positive spatial association between understory shrubs with their neighboring seedlings at a mean distance of 1.6 m. Overall, the study highlighted the requirements for seedling regeneration as a relatively open canopy cover, a light understory litter layer, and non-compacted soils. These results are a step towards designing effective management and restoration programs.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Natural forest regeneration is a highly complex process that can be affected by several factors. Recent studies suggest that regeneration niches change over the life cycle of individuals (Delerue et al. 2015; Leverkus et al. 2016; Ghosh et al. 2016), and these changes are known as ontogenetic niche development, i.e., niche requirements become more restrictive with age from seedlings to saplings (Quero et al. 2008). The forest understory provides regeneration habitats composed of mineral soil, decaying logs, rocks, and leaf litter that might be considered an establishment niche depending on tree species (Titus and Moral 1998). However, microenvironments suitable for seed germination are not necessarily appropriate for seedling establishment or sapling survival (Quero et al. 2008). Bulky litter layers under a pine forest canopy can inhibit seedling emergence by preventing contact between the seminal root and soil (Caccia and Ballaré 1998; Ibáñez and Schupp 2002) or through mechanical restriction to hypocotyl elongation and the emergence of cotyledons (Facelli and Pickett 1991). García et al. (2006) reported strong correlations between the litter cover and the overstory canopy. Marchi and Paletto (2010) reported that canopy cover can facilitate early seedling survival in summer through the positive effect of shade that reduces soil temperature, but that accumulated litter on the ground might lessen early-stage seedling survival. Caccia and Ballaré (1998) concluded that in old-growth coniferous forests with relatively thick litter layers and dense vegetation cover, the removal of litter combined with gap creation significantly promoted the initial establishment of Douglas fir beneath the stands. Some conifers grow better under low shade during their first year, whereas older seedlings and saplings require full sunlight (Gaudio et al. 2011). Pinus brutia, a light-demanding species, suffers from reduced regeneration at light intensity levels below 65–70% (Özdemir 1977). Besides thick litter and closed canopy cover, early growth of planted conifers is negatively affected by soil compaction (Froehlich et al. 1986; Kozlowski 1999). Bulmer and Simpson (2005) reported that survival of Pinus contorta seedlings after 2 years was reduced by 40% and seedling shoot weight was reduced by approximately 50% in response to increasing soil compaction. However, the detrimental effects of compaction are not general and mainly depend on the soil type (Gomez et al. 2002), as well as the level of compaction (Kabzems and Haeussler 2005; Sanchez et al. 2006). In northern Iraq, natural regeneration should be considered a valuable process for moving towards forest types that provide non-woody forest products and fulfil multipurpose forest functions such as recreation, conservation, and preservation of habitats of endangered species of plants and wildlife. P. brutia forms a unique natural pine forest that stretches from Zawita to Atrush town in the Kurdistan Region of northern Iraq. It represents a relict boundary of the southern limits of the species distribution in the eastern Mediterranean (Shahbaz 2007). For two decades, Zawita natural forest has been subject to human exploitation, including shifting agriculture, fires, and recently, mass recreational activities during spring, denuding of large areas of the natural forest. Due to the negative impacts of these anthropogenic factors on pine forests and the lack of resources to fund large afforestation campaigns, it is essential to assess the regeneration niche requirements of P. brutia before designing effective management and restoration programs. Based on studies demonstrating that regeneration niches change over the course of the lifespan of a tree or a stand (Gómez-Aparicio 2008; Granda et al. 2012; Delerue et al. 2015), we hypothesized that seedlings and saplings of P. brutia would respond differently to the environmental factors affecting its regeneration. We expected more seedlings to establish themselves outside the closed overstory canopy in the plots with shallow litter layers, light soil compaction, and with decreasing slope gradients. We expected saplings to establish themselves outside the overstory canopy, even with increasing slope gradients and we predicted that saplings would show greater tolerance of soil compaction than seedlings.
Materials and methods
Study site
Our study site was located on the northern slopes of the Zawita foothills, Duhok district, in northern Iraq (36°53′48″N−43°08′48″E). The natural forest occupied about 100 km2 on a limestone foothill, 16 km from Dohuk city center. The climate is continental Mediterranean with mild, wet winters and warm dry summers. Mean annual rainfall is about 550 mm, the average minimum temperature is 9.0 °C, and the average maximum temperature is 22.3 °C. The monthly average relative humidity ranges from 18.5 to 68%, with an average of 44% (Directorate of Meteorology of Duhok 2014). The soil is acidic (mean pH = 5.0) and low in nutrients compared to the natural oak forest (Ali et al. 2016). Zawita is famous for its pine forest with a ridge that is 70% covered by P. brutia. This natural forest is of great ecological and economic interest. Sites shaded by P. brutia in Zawita make it particularly attractive in the summer. The study site has a long history of grazing and land use, particularly shifting cultivation and more recently urbanization and mass recreation.
Species
In Iraq, P. brutia (Zawita pine in Iraqi) occurs as a relict along the southern limits of the species distribution in the eastern Mediterranean. It is a unique natural pine forest in the Kurdistan region of Iraq on rocky limestone substrate at elevations of 800–1200 m, on red marl valley deposits but not on wet and compacted clay soils. The species is distributed naturally in Zawita and Atrush-Duhok districts where is covers an area of 10,000 hectares. Several physiological and morphological characteristics of P. brutia make it well adapted to the Mediterranean type climate (Thanos and Scordilis 1987). P. brutia is an obligate seeder (non-resprouter), a light-demanding species that is tolerant of drought, poor soils, and frost (Fernandes et al. 2008). Zawita pine has been widely planted in the Kurdistan region and even outside its natural distribution area all over Iraq for the purposes of re-establishing vegetation on bare watersheds and roadsides, and establishing urban gardens.
Sampling and data analyses
Sampling methods
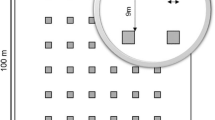
Our study of P. brutia regeneration was conducted between mid September to the end of November 2015 on the northern slopes of Zawiat where natural forest is still present. Ten plots were sampled at approximately 200 m intervals along a transect. Regeneration was assessed by counting all pine stems in 10 rectangular plots measuring 20 × 25 m (0.5 ha). Each plot was divided into 20 quadrats (25 m2) using rubber tapes (Fig. 1). Pine stems were classified in height and diameter classes as: (1) seedlings (height ≤ 50 cm); (2) saplings (height > 50 cm and diameter at breast height (DBH) < 4 cm); (3) adult trees (DBH ≥ 4 cm). We also recorded shrubs > 50 cm in height whose crown diameter was greater than 30 cm. The variables measured in each plot were classified as stand structure data including: (1) tree DBH measured using a tree caliper; (2) the projected area covered by the canopies of parent trees and shrubs estimated from two measurements of crown diameter (approximated to an ellipse) regardless of whether the stems originated inside or outside the plot, and the positions of the regenerating stems were recorded as either under or outside the canopy; (3) observed shrub-seedling spatial association was tested by measuring the distance to the shrub’s nearest three seedlings using a measuring tape; (4) litter thickness was measured by inserting a metal ruler down to the soil surface at 5 points (4 in the corners and 1 in the center of the quadrat); (5) soil compaction per quadrat (penetration resistance—psi) was measured on 1 and 2 October 2015, using Digital Soil Compaction Meter Specification Model: REIGN-450 and (6) slope gradient was measured using a Suunto PM5/360PC Clinometer at the center of each quadrat.

Each of the 10 (500 m2) studied plots were divided into 20 quadrates (25 m2) using rubber tapes
Data analyses
Pinus brutia recruit data were analyzed to assess the correlations between seedlings and saplings with litter depth, slope, and soil compaction. We used linear regression in Microsoft® Office Excel 2007 to quantify regeneration patterns. A Chi square test (χ2) based on Monte Carlo simulation (2000 replicates) was used to compare observed and expected numbers of pine seedlings and saplings under the canopy in each plot. Expected frequencies were derived by multiplying the percent canopy cover of trees and shrubs by the total frequency of pine seedlings and saplings. The null hypothesis was that the presence of seedlings and saplings beneath the tree and shrub canopies was independent of the size of the canopy (Muhamed et al. 2015). A Chi square test (χ2) was performed using R software for statistical computing (version 2.10.1, 2010). The spatial distribution of seedlings with respect to the shrubs was quantified using the nearest neighbor index (NNI) measure. This index is calculated from the relation of the mean distances between each shrub and its three nearest neighbor seedlings and the mean distances in a completely spatially random (CSR) pattern. The CSR pattern was generated on the assumption that seedlings were equally likely to be regenerated anywhere in the plot and unrelated to the spatial distribution of shrubs (Clark and Evans 1954). This test was only performed for seedlings because we expected that during establishment, the distribution of seedlings, more than saplings, would be influenced by the presence of shrubs:
where Dobs is the observed mean distance between each shrub and its three nearest neighbors. Dexp is the expected mean distance between shrubs given a random pattern.
In the above equations, \( di \) is the distance between shrub \( i \) and its nearest three seedlings. The nearest neighbor index (NNI) produces a result between 0 and 2.15. If the seedlings are randomly scattered with respect to the shrubs, Dexp = Dobs, NNI = 1, if the seedlings are skewed toward the shrubs, Dobs < Dexp, NNI = 0, but if the seedlings are skewed away from the shrubs Dobs > Dexp, NNI = 2.149.
The \( {\mathcal{Z}} \) ANN-score for the statistic is calculated as:
The value of the ANN statistic is transformed into a standard normal deviate z that is amenable to a z-test. This test measures the difference between an observed statistic and its hypothesized population parameter in units of the standard deviation (Eq. 4). Z statistics were used to assess the difference between the pattern determined by the NNI index and a random pattern (Eq. 5).
Results
A total of 1451 recruits were counted in the 10 plots, including 854 and 597 pine seedlings and saplings, respectively and 89 trees, in addition to 180 shrubs (Table 1). Shrub families were Fagaceae (40 shrubs of Quercus aegilops and 73 shrubs of Q. infectoria), Rosaceae (29 Prunus microcarpa, 9 Crataegus azarolus L. and 2 Prunus argentia (Lam.) Rehder), Fabaceae (15 Anagyris foetida), Rhamnaceae (4 Paliurus spina-christi), Cupressaceae (1 Juniperus oxycedrus) and Anacardacea (5 Pistacia khinjuk stocks).
Probability of regeneration
Seedling density ranged from 29 to 238 seedlings and 0–274 saplings in the 0.5 ha study plots (Table 1). Seedlings and saplings differed in their responses to the studied factors. There was a strong negative correlation between seedling abundance and litter depth (R2 = 0.67, p = 0.001, Fig. 2). Most seedlings were growing in locations where the litter was between 0.5 and 2.5 cm in depth. The number of saplings was not related to litter depth (Fig. 3, R2 = 0.056, p = 0.062). As litter depth increased from 2.5 to 4.5 cm or at depths < 0.5 cm, the number of empty quadrats increased. The number of regenerated seedlings was negatively correlated with soil compaction (R2 = 0.38, p = 0.001, Fig. 3), but the number of saplings was not (R2 = 0.005, p = 0.620). Sapling regeneration was only correlated with slope gradient (Fig. 5, R2 = 0.53, p = 0.001). As the slope gradient increased from 10 to 35%, the number of saplings increased from 1 to 45. The number of regenerated seedlings was not correlated with slope gradient (Fig. 4, R2 = 0.002, p = 0.71).

The correlation of P. bruita regeneration (seedlings and saplings) with litter depth in 200 quadrates of 10 (500 m2) plots

The correlation of the P. bruita regeneration (seedlings and saplings) with the soil compaction degree in 200 quadrates of 10 (500 m2) plots

The correlation of the P. bruita regeneration (seedlings and saplings) with the slope gradient in 200 quadrates of 10 (500 m2) plots
Both seedlings and saplings regenerated in greater numbers when not shaded by a tree or shrub canopy (Table 2, Fig. 5). The number of pine seedlings was significantly disassociated with the canopy cover in 7 out of 10 cases (70%), with positive and independent associations in 20 and 10% of the cases, respectively (Table 2). The same results were found for saplings (70%), (10%) and (20%) for negative, positive, and independent associations, respectively.

The spatial repulsion pattern between parent trees and P. bruita regeneration
Nearest neighbor analysis
The mean distances of three nearest seedlings to shrubs were used as a measure of the spatial association of seedlings and shrubs. Nine out of the ten plots showed significant positive spatial association between seedlings and their neighboring shrubs (Table 3). The average shrub canopy area was 1.04 m2 and the average scale of association was 1.6 m; the spatial association mainly occurred at the edges of the shrub canopies.
Discussion
Canopy cover and litter effect
The greater probability was associated with finding significantly fewer seedlings and saplings growing under the overstory canopy compared to the number growing beyond the canopy. This was due to the heliophilous nature of the species, meaning it mainly germinates and establishes itself in gaps and other well-illuminated places (Thanos and Doussi 2000). The low density of saplings in the quadrats under the tree canopy was probably due to the lack of sunlight because saplings are not affected by litter thickness. The low density of seedlings under canopy cover was not due to light deficiency, but most likely resulted from the physical and/or chemical influence of the litter. This is in agreement with the results obtained for other conifer species by Fady et al. (2004) and by Adili et al. (2013), who reported that Pinus pinea L. does not require abundant sunlight for seed germination and seedling emergence, but that sapling survival and growth are mainly determined by light availability. Ganatsas and Tsakaldimi (2007) also reported that percent germination of P. pinea seed was high and did not depend on light conditions, while Gaudio et al. (2011) reported that P. sylvestris L. needed more sunlight with increasing size of the trees. Although, P. brutia is considered to be a light demanding species and shows considerable drought tolerance (Thanos and Doussi 2000; Boydak 2004) the seedlings can also tolerate and to some extent prefer shade, particularly in drier areas with severely dry summers (Gómez-Aparicio et al. 2008; Granda et al. 2012). With an average shrub canopy cover of 1.04 m2 and 1.6 m average shrub-seedling association, most of the regeneration in the study area did not take place directly beneath shrub canopies (Table 3 and see Chi square tests in the Appendix). Seedlings that are concentrated at a certain distance beyond the shrub canopy can be affected differently, e.g., due to improvement in the soil caused by the root system or due to clonal growth of seedlings in inter-shrub spaces (García et al. 2000; Muhamed et al. 2013). We documented a stronger negative effect of litter on seedling density than on sapling density. P. brutia is an epigeal germination species that does not require a thick cover layer for germination. When the thickness of the ground litter increased from 0.5 to 4.5 cm, the number of seeds that germinated decreased because the litter prevented the radicals from reaching the soil. When the thickness of the litter exceeded 2.5 cm, the number of empty quadrats began to increase (Fig. 3). These results are consistent with previous reports that the accumulation of litter reduced seedling emergence of small-seeded tree species (Dzwonko and Gawronski 2002). According to Caccia and Ballaré (1998), the main inhibitory effect of litter on seedling establishment of Douglas fir (Pseudotsuga menziesii (Mirb.) Franco) was seeds landing in the upper litter layer, i.e. at distances of 2–4 cm above the mineral soil. This increased the risk of mortality of the germinated seeds. Kozlowski (2002) reported that Pinus menziesii Douglas ex D.Don seeds germinated and seedlings survived at higher rates when the roots could immediately penetrate the mineral soil and where seedlings were exposed to direct sunlight. Wangchuk (2007) and Fernandez et al. (2008) suggested that litter can mechanically limit seedling emergence and that the accumulated thick litter of pine species commonly induces autotoxicity, particularly during early successional stages. The physical barrier represented by litter probably accounted for the low rate of regeneration of P. brutia in our study plots. The specific and relative roles of canopy and litter cover in pine regeneration were hard to disentangle because litter accumulation increases with increasing canopy cover and declines with increasing gap area (light availability). However, the significant correlation showed that litter was the main factor affecting seedling emergence, whereas light was the main factor affecting sapling growth, as also reported by Zhu et al. (2003) and Adili et al. (2013) for Pinus thunbergii Parl. and P. pinea, respectively.
Soil compaction and slope
The main sources of soil compaction at Zawita forest are recreational activities in spring and summer combined with the residual impact of trampling by grazing sheep a decade ago. In the present study, seedling density decreased linearly with increasing soil compaction. At soil compaction > 500 psi, we began to observe fewer established seedlings, even though compaction in Zawita forest is shallow since it is caused by trampling by humans or livestock. Nevertheless, the fact that seedling density was affected suggests that the emergence of P. brutia seedlings is sensitive to soil compaction. This supports results reported for other conifer species (Sands and Bowen 1978 for Pinus radiate Don., Corns 1988 for P. contorta and Picea glauca (Moench) Voss). However, when the compaction was shallow, saplings with well-developed root systems showed more tolerance to soil compaction, in agreement with observations by Ampoorter et al. (2011) who reported that soil compaction had no effect on the average growth and survival of saplings. One can thus conclude that soil compaction is not always detrimental for tree regeneration, but depends on the species ontogeny (Miller et al. 1996) and on the degree of compaction (Ehlers et al. 1983). The increase in sapling density with increasing slope gradient (5–35%) may be explained by the distance of the saplings from any direct human disturbance as most anthropogenic activities are concentrated in flat areas and on very gentle slopes thus limiting the opportunity for the seeds dispersed to these sites to develop into saplings.
Conclusions
Our study showed that the factors affecting P. brutia regeneration differed between seedling and saplings. Overstory canopy and understory microhabitat conditions played important roles in the regeneration of P. brutia. Seedlings tended to be more abundant in habitats located at a distance from parent trees, but in close proximity to understory shrubs where litter cover was shallower, adequate light was available, and soils were not compacted. Open overstory canopy and steeper slope gradients proved favorable for the growth of P. brutia saplings. Despite these findings, we cannot recommend a reduction in the canopy cover to enhance P. brutia regeneration because our study did not enable us to clearly determine whether the low density of seedlings under the canopy was due to canopy shading or to the thickness of the litter layer. A study of seedling survival including soil scarification with different litter treatments directly beneath the tree canopy and at a distance from the tree canopy is now necessary. Based on our findings concerning the relationship between soil compaction and regeneration, a study on the impact of human recreation on the natural regeneration of P. brutia in Zawiat forest is also recommended.
References
Adili B, El Aouni MH, Balandier P (2013) Unraveling the influence of light, litter and understorey vegetation on Pinus pinea natural regeneration. Forestry 86:297–304
Ali NS, Ali OO, Esmail AO (2016) Comparison between some soil chemical properties and some nutrients in trees at zawita and hijran forest. Agric Vet Sci 19(1):701–712
Ampoorter E, Frenne PD, Hermy M, Verheyen K (2011) Effects of soil compaction on growth and survival of tree saplings: a meta-analysis. Basic Appl Ecol 12(5):394–402
Boydak M (2004) Silvicultural characteristics and natural regeneration of Pinus brutia Ten.: a review. Plant Ecol 171(1–2):153–163
Bulmer CE, Simpson DG (2005) Soil compaction and water content as factors affecting the growth of lodgepole pine seedlings on sandy clay loam soil. Can J Soil Sci 85:667–679
Caccia FD, Ballaré CL (1998) Effects of tree cover, understory vegetation, and litter on regeneration of Douglas-fir (Pseudotsug amanziessii) in south-western Argentina. Can J Soil Sci 28:683–692
Clark PJ, Evans FC (1954) Distance to nearest neighbor as a measure of spatial relationships in populations. Ecology 35(4):445–453
Corns IGW (1988) Compaction by forestry equipment and effects on coniferous seedling growth on four soils in the Alberta foothills. Can J For Res 18(1):75–84
Delerue F, Gonzalez M, Michalet R, Pellerin S, Augusto L (2015) Weak evidence of regeneration habitat but strong evidence of regeneration niche for a leguminous shrub. PLoS ONE. https://doi.org/10.1371/journal.pone.0130886
Dzwonko Z, Gawronski S (2002) Effect of litter removal on species richness and acidification of a mixed oak-pine woodland. Biol Conserv 106:389–398
Ehlers W, Popke V, Hesse F, Bohm W (1983) Penetration resistance and root growth of oats in tilled and untilled loam soil. Soil Tillage Res 3(3):261–275
Facelli JM, Pickett STA (1991) Plant litter: its dynamics and effects on plant community structure. Bot Rev 57(1):1–32
Fady B, Fineschi S, Vendramin GG (2004) EUFORGEN Technical Guidelines for genetic conservation and use for Italian stone pine (Pinus pinea). International Plant Genetic Resources Institute, Rome
Fernandes PM, Vega JA, Jimenez E, Rigolot E (2008) Fire resistance of European pines. For Ecol Manag 256:246–255
Fernandez C, Voiriot S, Mévy JP, Bruno V, Ormeno E, Dupouyet S, Bousquet-Mélou A (2008) Regeneration failure of Pinus halepensis Mill.: the role of autotoxicity and some abiotic environmental parameters. For Ecol Manag 255:2928–2936
Froehlich HA, Miles MD, Robbins RW (1986) Growth of young Pinus ponderosa and Pinus contorta on compacted soils in central Washington. For Ecol Manag 15:285–294
Ganatsas P, Tsakaldimi M (2007) Effect of light conditions and salinity on germination behaviour and early growth of umbrella pine (Pinus pinea L.) seed. J Hort Sci Biotechnol 82:605–610
García D, Zamora R, Hódar JA, Gómez JM, Castro J (2000) Yew (Taxus baccata L.) regeneration is facilitated by fleshy-fruited shrubs in Mediterranean environments. Biol Conserv 95:31–38
García LV, Maltez-Mouro S, Pérez-Ramos IM, Marañón T, Freitas H (2006) Counteracting gradients of light and soil nutrients in theunderstorey of Mediterranean oak forests. Web Ecol 6:67–74
Gaudio N, Balandier P, Philippe G, Dumas Y, Jean F, Ginisty C (2011) Light-mediated influence of three understorey species (Calluna vulgaris, Pteridiumaquilinum, Moliniacaerulea) on the growth of Pinus sylvestris seedlings. Eur J For Res 130(1):77–89
Ghosh S, Zhu K, Gelfand AE, Clark JS (2016) Joint modeling of climate niches for adult and juvenile trees. J Agric Biol Environ Stat 21(1):111–130
Gomez A, Powers RF, Singer MJ, Howarth WR (2002) Soil compaction effects on growth of young ponderosa pine following litter removal in California’s Sierra Nevada. Soil Sci Soc Am J 66:1334–1343
Gómez-Aparicio L (2008) Spatial patterns of recruitment in Mediterranean plant species: linking the fate of seeds, seedlings and saplings in heterogeneous landscapes at different scales. J Ecol 96:1128–1140
Gómez-Aparicio L, Zamora R, Castro J, Hódar JA (2008) Facilitation of tree saplings by nurse plants: microhabitat amelioration or protection against herbivores? J Veg Sci 19:161–172
Granda E, Escudero A, de la Cruz M, Valladares F (2012) Juvenile-adult tree associations in a continental Mediterranean ecosystem: no evidence for sustained and general facilitation at increased aridity. J Veg Sci 23:164–175
Ibáñez I, Schupp EW (2002) Effects of litter, soil surface conditions, and microhabitat on Cerocarpus ledifolius Nutt. seedling emergence and establishment. J Arid Env 52:209–221
Kabzems R, Haeussler S (2005) Soil properties, aspen and white spruce responses 5 years after organic mater removal and compaction treatments. Can J For Res 35:2045–2055
Kozlowski TT (1999) Soil compaction and growth of woody plants. Scand J For Res 14:596–619
Kozlowski TT (2002) Physiological ecology of natural regeneration of harvested and disturbed forest stands: implications for forest management. For Ecol Manag 158:195–221
Leverkus AB, Rey Benayas JM, Castro J (2016) Shifting demographic conflicts across recruitment cohorts in a dynamic post-disturbance landscape. Ecol 97:2628–2639
Marchi A, Paletto A (2010) Relationship between forest canopy and natural regeneration in the subalpine spruce-larch forest (north-east Italy). Folia Forestalia Polonica. Series A. Forestry 52(1):3–12
Miller RE, Scott W, Hazard JW (1996) Soil compaction and conifer growth after tractor yarding at three coastal Washington locations. Can J For Res 26(2):225–236
Muhamed H, Maalouf J-P, Michalet R (2013) Summer drought and canopy opening increase the strength of the oak seedlings–shrub spatial association. Ann For Sci 70:345–355
Muhamed H, Lingua E, Maalouf J-P, Michalet R (2015) Shrub-oak seedling spatial associations change in response to the functional composition of neighbouring shrubs in coastal dune forest communities. Ann For Sci 72:231–241
Özdemir Ô (1977) Studies on the possibilities of natural regeneration of Pinus brutia Ten. in Antalya region (in Turkish). Review of the Faculty of Forestry, University of Istanbul Á/2:(Abstract)
Quero JL, Gómez-Aparicio L, Zamora R, Maestre FT (2008) Shifts in the regeneration niche of an endangered tree (Acer opalus ssp. granatense) during ontogeny: using an ecological concept for application. Basic Appl Ecol 9:635–644
Sanchez FG, Scott DA, Ludovici KH (2006) Negligible effects of severe organic matter removal and soil compaction on loblolly pine growth over 10 years. For Ecol Manag 227:145–154
Sands R, Bowen GD (1978) Compaction of sandy soils in radiata pine forests. II. Effects of compaction on root configuration and growth of radiata pine seedlings. Aust For Res 8:163–170
Shahbaz S (2007) Pinales: Spirez Press and Publication, colored. Duhok, Kurdistan Region of Iraq, 188 pp
Thanos CA, Doussi M (2000) Post-fire regeneration of Pinus brutia forests. In: Ne’eman G, Trabaud L (eds) Ecology, biogeography and management of Pinus halepensis and P. brutia forest ecosystems in the Meditenanean basin. Backhuys Publisher, Leiden, pp 291–301
Thanos CA, Skordilis A (1987) The effect of light, temperature and osmotic stress on the germination of Pinus halepensis and Pinus brutia seeds. Seed Sci Technol 15:163–174
Titus JH, del Moral R (1998) Seedling establishment in different microsites on Mount St. Helens, Washington, USA. Plant Ecol 134:13–26
Wangchuk K (2007) Natural regeneration ecology of mixed conifer forests in Western Bhutan. M.Sc.thesis, University of Applied Life Sciences, Vienna, Austria, 79 pp
Zhu JJ, Takeshi M, Lee FQ, Yutaka G (2003) Effect of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest. For Ecol Manag 182:339–354
Acknowledgements
We thank the Directorate of the Forestry and Rangelands of Duhok for permission to work in Zawita forest and for their assistance in the field.
Author information
Authors and Affiliations
Corresponding author
Additional information
Project funding: The work was supported by the Agriculture College of Duhok University.
The online version is available at http://www.springerlink.com
Corresponding editor: Tao Xu.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Muhamed, H., Youssef, S., Mustafa, A. et al. Natural regeneration of Pinus brutia Ten. in a recreational public forest in Zawita-Kurdistan region, Iraq. J. For. Res. 30, 1849–1857 (2019). https://doi.org/10.1007/s11676-018-0739-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-018-0739-x