
Abstract
In forest ecosystems, gap formation changes the allocation of abiotic resources and thus affects the survival and growth of understory plants. However, how tree seedling survival and growth respond to low-temperature events and the influencing mechanisms remain unclear. To clarify how low-temperature event limits the survival and growth of tree seedlings in the montane regions of eastern Liaoning Province, northeast China, we investigated temperature and light intensity within secondary forest gaps, and the survival and growth of Juglans mandshurica seedlings after a low-temperature event in the spring of 2014. Damage to seedlings due to low temperature significantly varied in different aspects. Seedlings in gaps on southeast-facing slopes were the most seriously damaged, followed by those in gaps on northeast-facing slopes. In contrast, seedlings in west-facing gaps and in control plots without slope aspect were not damaged. The freezing injury index for seedlings was negatively correlated with minimum temperature (r = − 0.608, P < 0.01), but it was positively correlated with light intensity (r = 0.818, P < 0.01). In addition, height and root collar diameter of damaged seedlings were significantly lower than those of the undamaged seedlings (P < 0.01) during the early growing season (April–July), but no significant difference were observed during the late growing season (July–October) (P > 0.05). The extent of seedling damage was directly related to slope aspect. Low temperature and high light intensity were found to be the dominant factors affecting extent of damage to seedlings on southeast- and northeast-facing slopes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Low-temperatures during the growing season fatally damage plant tissues, seedlings and even whole saplings (Blennow et al. 1998; Dittmar et al. 2006; Tan 2013). However, studies on the effects of low-temperature have focused on crops, while relatively little work has explored the effects on forests (Li et al. 2013). Low-temperature events affect survival and growth of seedlings in natural and plantation forests (Lindkvist and Chen 1999; Chen et al. 2000; Wu et al. 2002), and ultimately affect forest development and the ecological services provided by forests (Jönsson et al. 2004; Awaya et al. 2009; Ramming et al. 2010). Therefore, studying the effect of low-temperature event on regeneration in forest ecosystems will provide a scientific basis for protecting existing forest resources and promoting sustainable forest management.
The impact of a low-temperature event depends on both the duration and intensity of the low temperature. Topography, slope and stand characteristics (e.g., degree of canopy closure) are also important factors, particularly in complex, hilly and mountainous terrain (Zhang 1992). Forest gaps (induced by the death of trees) play an important role in regulating composition and structural diversity (Hubbell et al. 1999; De Grandpré et al. 2011; Zhu et al. 2015). Gaps exist widely in forest ecosystems, providing potential space for the growth and regeneration of forests (Zhu et al. 2007). The canopy structure changes significantly following gap formation, including the redistribution of light, temperature and water within the gaps, creating a specific micro-climate in the gaps (Zhang et al. 2002). Thus, it is widely recognized that the environmental heterogeneity induced by gap formation influences the regeneration and distribution of species, population dynamics, species diversity and succession (Gray et al. 2012; Forrester et al. 2014; Liu et al. 2014; Stan and Daniels 2014; Jing et al. 2015; Zenner et al. 2015; Guan et al. 2016). The features of micro-climate and seedling regeneration after the formation of gaps is well understood, including differences in micro-climate between the inside and outside of gaps, the influence of light on the regeneration of forests, and heterogeneities in the soil moisture and air humidity (Duan et al. 2008; Romell et al. 2009; Zhu et al. 2009; Li et al. 2012; Zenner et al. 2015). However, few researches have been reported on the influence of adverse meteorological conditions on seedlings within forest gaps. In fact, the influences induced by adverse meteorological conditions are expected to differ both within an individual gap and among gaps. This in turn will cause variations in the regeneration and growth of seedlings within gaps because spatial differences exist among gaps (Duan et al. 2007; Wang et al. 2015; Zhu et al. 2016). Therefore, research on adverse meteorological conditions within gaps would be of great importance for furthering our understanding of early regeneration processes of forest ecosystems and reasonable management under natural disturbances.
The montane region of eastern Liaoning Province has suffered from the most serious and frequent low-temperature events. Low-temperature events commonly threaten both agriculture and forestry in this region and are known to be one of the main factors limiting early growth and regeneration of seedlings and saplings. Low-temperature events have commonly occurred in this area, with most of these events occurring in early May (Zhang 1992) when the leaves of plants are initially formed (Yan et al. 2016) and seedlings are unable to withstand low-temperatures. In fact, low-temperature events occur more easily when temperature falls below 0 °C in the early spring.
In the present study, we investigate the survival and growth of Juglans mandshurica seedlings in secondary forest gaps in the montane region of eastern Liaoning Province, Northeast China, after a low-temperature event occurred in the early May of 2014. The purposes of this research are to clarify how low temperature limits the early survival and growth of tree seedlings within secondary forest gaps with different slope aspects, and hence to clarify the impact of low temperature on early regeneration of tree seedlings within gaps. In addition, some practical tending felling measures will be proposed to alleviating the seedling damage. These research results will helpful for further understanding how adverse meteorology conditions affect the regeneration of forest within gaps.
Materials and methods
Study sites
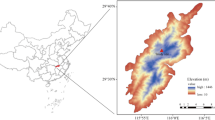
The study was conducted at the Qingyuan Forest CERN, Chinese Academy of Sciences, located in a mountainous area of Liaoning Province, Northeast China (41°51′N, 124°54′E, 500–1100 m above sea level). This region has a continental monsoon climate, with humid and rainy summers and cold and dry winters. The mean annual temperature is 4.5 °C, ranging from − 37.6 °C in January to 36.5 °C in July. The annual precipitation fluctuates between 700 and 850 mm, with up to 80% falling from June to August. On average, the frost-free period lasts for about 130 days, with an early and late frost in October and April, respectively (Zhu et al. 2007).
The study site was dominated primarily by mixed broadleaf Korean pine forests prior to the 1930s but it was subjected to unregulated timber removal for decades thereafter. In the early 1950s, a large-scale fire almost decimated the original forest, resulting in that the study site naturally regenerated to secondary forest. The natural secondary forests are mainly composed of Fraxinus mandschurica, Juglans mandshurica, Phellodendron amurense, Quercus mongolica and Acer mono in the tree layer; Acer mandshricum, Acer triflorum, Acer tegmentosum and Syringa amurensis in the understory layer, and Cardamine leucantha, Allium monanthum, Arisaema amurense and Polygonatum involucratum in the herbage layer. In the early 1960s, patches of the natural secondary forests were partially cleared and replaced by larch (Larix gmelinii) plantations. The larch plantations mainly consisted of Acer tegmentosum, Acer pseudo-sieboldianum, Schisandra chinensis, Syringa wolfi and Acanthopanax senticosus in the shrub layer and Cardamine leucantha, Rubia sylvatica and Spuriopimpinella brachycarpa in the herbage layer (Yang et al. 2013).
Experimental design and seedling transplantation
The selected gaps located in the secondary forests were formed by damage during a severe windstorm in 2003. The six gaps studied has similar area (260–300 m2) with different aspects (west, northeast and southeast). Each aspect was represented by two replicates and two control stands located in nearby secondary forests. These plots were situated in the middle of each aspect replicate, which were all located on slopes between 20 and 25° and at elevations ranging from 580 to 660 m asl. The soil in all plots was typical brown forest soil. The height of the secondary forests was 15–20 m, respectively. The shape of the studied gaps was an irregular ellipse (Fig. 1). We laid out five 1 m × 1 m plots (A, B, C, D and E) in each gap (Fig. 1) and three randomly placed plots (1 m × 1 m) in each control stand. Twenty one-year-old seedlings of J. mandshurica with similar heights and root collar diameters were planted in each plot (with a row space of 0.2 m).

Schematic of the experimental design for each replicate gap
Environmental factors and seedling growth
The temperature and photosynthetic photon flux density (PPFD) (μmol m−2 s−1) at a height of 0.5 m above the forest floor were measured automatically in each plot during the growing season (April–October). The temperature was continuously monitored at 10-min intervals using a HOBO U23 Pro v2 external temperature data logger (U23-004, MacArthur, USA), and the PPFD was recorded by using a LI-191SA (LI-COR, Lincoln, NE, USA) at 1-h intervals during the daytime on May 6–9, 2014. The measurements were taken between 8:00 and 17:00. The height and root collar diameter of each seedling were measured in the middle of each month during the growing season.
Investigation of seedling damage
After the low-temperature event, the extent of damage that occurred to each seedling in the plots was recorded using the following classification: 0, no leaf damage; 1 (light damage), 1/3 of the leaf area was damaged; 2 (moderate damage), 1/2 of the leaf area was damaged; 3 (moderate-heavy damage), 2/3 of the leaf area was damaged, 4 (heavy damage), the entire leaf area was damaged (Li et al. 2005). The freezing injury index (FI) was used to quantify the level of leaf damage for each plot, and the calculation of FI (at A, B, C, D and E) was as follows:
where 1, 2, 3 and 4 represent the damage levels (1–4) and S1, S2, S3 and S4 represent the number of leaves damaged relative to each level (1–4).
The damage ratio (DR) refers to the number of damaged seedlings relative to the total number of seedlings in each plot.
Data analysis
A t test was used to determine the differences in damage among seedlings of J. mandshurica for different gaps and the differences in the increases in the height and root collar diameter between the damaged and undamaged seedlings. Correlation analysis and multivariate analysis of variance (MANOVA) were used to determine the relationships between the damage of seedlings and the light and temperature in the gaps. All the statistical analyses were conducted using SPSS 19.0 for windows. The significance level was set at of P < 0.05.
Results
Overview of the low-temperature events
Controlled by cold vortex weather, most regions of Liaoning Province suffered from three times of apparent temperature decreases during the period of May 2–9, 2014, which involved a long period of rainy, cold weather. The minimum temperature during the period dropped to − 2.0 °C. This stage was the critical period for early plant growth. As a result, crops and seedlings in the region were severely damaged by the low-temperature event.
The study plots were also subjected to the low-temperature event. Night time and early morning temperatures were below zero on May 3, May 5–6 and May 8–9, and the longest period of low temperature (more than 10 h) occurred on May 5–6, with a minimum temperature of − 1.4 °C (Fig. 2). The leaves of seedlings were initially formed during this period because the temperature of late April was commonly higher than 10 °C. However, seedlings at this site (including the J. mandshurica seedlings in gaps) were damaged by the early spring freezing resulting from the sudden and long period of rain and low temperatures.

The variations of 24-h minimum temperature from April 20 to May 9 of 2014
The damage of seedlings in different gaps
The damage ratio (DR) of the J. mandshurica seedlings in the southeast-, northeast-, west-gaps and the control plots were 71.2, 54.6, 0 and 0%, respectively. The corresponding seedling mortality of above four plots were 23.1, 18.9, 0 and 0%, respectively. Significant differences were observed for FI between the southeast and northeast gaps (t = − 2.75, P < 0.05, Table 1).
The relationships between FI and light and temperature
Across all plots, FI was significantly and negatively correlated with minimum temperature (r = − 0.608, P < 0.01). By contrast, FI was significantly and positively correlated with PPFD for the 8:00 a.m.–12:00 p.m. period after low temperature events (r = 0.818, P < 0.01). No significant relationships were observed between FI and mean temperature, diurnal temperature range or PPFD for the 1:00–5:00 p.m. period (Table 2). This result suggests that the minimum temperature, as well as PPFD after the minimum temperature, closely link to the FI. Therefore, we performed multivariate variance analysis to examine the relationship between the low temperature and PPFD (8:00 a.m.–12:00 p.m.) for different gaps.
Multivariate variance analysis showed that there were significant differences relative to the minimum temperature (P = 0.012) and the PPFD for the 8:00 a.m.–12:00 p.m. period (P = 0.001) among the gaps. The minimum temperature in the southeast-facing gap (highest FI) was not the lowest in all the studied gaps, but the PPFD was the highest. In contrast, the west-facing gap (with the minimum temperature and a relatively low PPFD) and the control plots (with relatively higher temperature and the lowest PPFD) did not suffer from low-temperature (Fig. 3). These results indicate that freezing damage to J. mandshurica seedlings was caused by a combination of low temperature and high PPFD after low temperature.

The minimum temperature and PPFD in different gaps
The influence of low-temperature on the seedling growth
The increases in the heights and root collar diameters of damaged seedlings in the early growth season (from April to July) were significantly lower than those of undamaged seedlings (P = 0.001 and P = 0.000, respectively). There were no significant differences in the increases in the heights and root collar diameters of the damaged seedlings in the later growth season (from July to October) (P > 0.05) (Fig. 4). This indicates that the seedling growth in early growth season is susceptible to low temperature.

The increase in the height and root collar diameter in early (from April to July) and later (from July to October) stages between the damaged and undamaged seedlings of J. mandshurica
Discussion
When seedlings are stressed by short-term periods of low temperatures, they can adapt to the low temperature by self-regulating and establish a new equilibrium to reduce the damage caused by the low temperature and to resist cold to a certain extent (Luo et al. 2007; Jie et al. 2008). However, the physiological functions of the seedlings are completely destroyed under sustained periods of low temperature, e.g., cell membranes are severely damaged and their rate of electrolyte leakage increases, causing severe damage to plants (Zhou et al. 2010; Wang et al. 2011). Although the minimum temperature during this low-temperature process was not much below freezing (− 1.4 °C), the damage occurred due to the sustained stress of low temperatures.
In general, damage caused by low temperature is greater under high light conditions; i.e., low-temperature damage occurs more easily in the light than in the dark (Jie et al. 2008). The damage to photosynthetic tissue caused by low temperatures is exacerbated when plants are co-stressed by low temperature and strong light (Egerton et al. 2000; Guo et al. 2015). Dirks et al. (2002) reported that photoinhibition occurs under the stress of low temperatures even when plants are exposed to moderate-intensity light. Ultimately, light-induced oxidative stress occurs in the leaves of plants (Sane et al. 2003; Wang and Feng 2005). Therefore, light, especially strong light, intensifies plant damage when temperature is low (Jie et al. 2008). The degree of canopy openness and light intensity markedly increase in gaps, which potentially aggravates the stress caused by low temperatures. Light and temperature conditions in gaps are affected by topography (e.g., depending on slope aspect), which in turn exerts different degrees of physiological stress on the seedlings growing in gaps (Table 2, Fig. 3).
Although the temperature in the west-facing gaps is the lowest, seedlings did not experience damage due to the relatively smaller PPFD. By contrast, seedlings in the southeast-facing and northeast-facing gaps with relatively lower temperature and higher PPFD were seriously damaged, especially in the southeast-facing gaps having the highest PPFD. This indicates that the combined stress of low temperature and strong light is the dominant factor for determining the extent of damage in J. mandshurica seedlings, which is consistent with previous studies (Hu et al. 2011; Song et al. 2013; Tian et al. 2013).
Variations in light and temperature are relatively complex in montane regions due to variations in topography (e.g., slope aspects). Many researchers have simulated the spatial variations of low temperature and light in montane regions in order to assess damage or to predict the damage caused by low temperature and strong light (Laughlin and Kalm 1990; Blennow 1998; Blennow and Lindkvist 2000). The aim of these studies was to reveal how the distribution of low temperature and strong light, in combination with the physiological characteristics of trees, can be used to guide forestation and damage alleviation. For example, Blennow et al. (1998) demonstrated that under the simulated temperature and light conditions, areas with low temperature and high light coincided with the areas containing damaged seedlings. More specifically, the combination of increased interception of direct radiation energy (IDRE) and reduced temperature would result in strong photoinhibition. Further, Song et al. (2013) reported that shading treatment for Populus xiaohei should be performed after the onset of low temperatures (i.e., after the seedlings are transplanted in the spring), with the aim of reducing photoinhibition and thus maximizing the survival rate. Cai et al. (2003) noted that economic crops and precious plants should be planted in a low-light understory or under artificial shading to reduce the damage caused by low temperatures and strong light. Our results also show that defensive measures (e.g., shading treatment) should be taken into consideration with seedling transplant in southeast-facing gaps to reduce the low temperature and strong light induced damages. In addition, our results show that the seedlings in the control plots are undamaged after the low temperature event, which are attributed to decrease of long-wave radiation from ground and incoming solar radiation in the control plots because the seedlings are sheltered by the bigger branches and canopies in the control plots compared to that plots within gaps (Zhu et al. 2015). This suggests that certain measures will help to mitigate damage extent of tree seedlings suffering from low temperature, such as keeping some trees as protection tree, thinning instead of clear cutting during tending felling processes (Nunez and Bowman 1986; Blennow 1998; Rambo and North 2009; Li et al. 2013).
The seedling stage is an important period in the history of a plant’s life, and the growth rate of seedlings influences the quantity and development of the population (Clark 1998; Jin et al. 2007; Nordlander et al. 2011), which is of great importance for population stability and community composition (Muhamed et al. 2013; Verduin et al. 2013). The seedling stage is also the most vulnerable and sensitive period in the life cycles of forest plants (Han and Wang 2002; Tang and Zhai 2006), and seedlings of this stage are very sensitive to changes in habitat conditions (James et al. 2005; Diaci et al. 2012). Therefore, changes in weather can exert a great influence on the survival and growth of seedlings, which are directly related to the survival and development of populations and the regeneration, development and succession of forests. Gap formation is considered to be a crucial process in driving “the forest cycle and succession” and provides an opportunity for the regeneration of seedlings and saplings (Nagel et al. 2010). Consequently, variations in meteorological conditions in the gaps directly affect the growth and development of seedlings. Our results show that some seedlings are killed, some seedlings are damaged in some extent, and some seedlings are not damaged after the low temperature event in the gaps, indicating that the seedlings are sensitive to low temperature. Our results also show that the increases in the height and root collar diameter of the damaged seedlings are significantly lower than those of undamaged seedlings. Additionally, low temperature only affects the early growth of seedlings but has no effect on the later growth, suggesting that the seedling growth in early growth season was more susceptible to low temperature. Based on these results, it can be speculated that low temperature can lead to death of part of seedlings in gaps and damage of part of seedlings, while damaged seedlings will suffer from growth deficiency during early growth season, hence affecting the regeneration of seedlings in the gaps. Further study should be conducted to clarify ecological and physiological mechanism behind the effect of adverse metrological conditions on the regeneration of seedling in secondary forest gaps.
Conclusions
The seedlings of J. mandshurica in different gaps suffered from variable degrees of damage induced by low temperatures. Our results show that leaf damage to J. mandshurica in southeast-facing gaps is significantly higher than that of plants in northeast-facing gaps. By contrast, the seedlings in west-facing gaps and control plots are not damaged. Additionally, the increases in the height and root collar diameter (from April to July) of the damaged seedlings are significantly lower than those of undamaged seedlings, while no significant differences are observed in the later growing season (from July to October). The minimum temperature and light intensity in gaps are the main factors responding for leaf damage, therefore seedlings in southeast- and northeast-facing gaps with the highest light intensity are the most seriously damaged. This suggests that strong light is the key factor affecting damage after a low-temperature event. In addition, strong light after low-temperature only affects the growth of seedlings in the early part of the growing season without affecting growth in the later growing season.
References
Awaya Y, Tanaka K, Kodani E, Nishizono T (2009) Responses of a beech (Fagus crenata Blume) stand to late spring frost damage in Morioka, Japan. For Ecol Manag 257(12):2359–2369
Blennow K (1998) Modelling minimum air temperature in partially and clear felled forests. Agric For Meteorol 91(3):223–235
Blennow K, Lindkvist L (2000) Models of low temperature and high irradiance and their application to explaining the risk of seedling mortality. For Ecol Manag 135(1–3):289–301
Blennow K, Lang ARG, Dunne P, Ball MC (1998) Cold-induced photoinhibition and growth of seedling snow gum (Eucalyptus pauciflora) under differing temperature and radiation regimes in fragmented forests. Plant Cell Environ 21(4):407–416
Cai ZQ, Cao KF, Feng YL, Feng ZL (2003) Acclimation of foliar photosynthetic apparatus of three tropical woody species to growth irradiance. Chin J Appl Ecol 14(4):493–496
Chen XW, Zhang Y, Ma HB (2000) Occurrence and control of frost in Tilia amurensis and Fraxinus mand shurica young plantations. Chin J Appl Ecol 11(6):809–813
Clark JS (1998) Why trees migrate so fast: confronting theory with dispersal biology and the paleorecord. Am Nat 152(2):204–224
De Grandpré L, Boucher D, Bergeron Y, Gagnon D (2011) Effects of small canopy gaps on boreal mixedwood understory vegetation dynamics. Commun Ecol 12(1):67–77
Diaci J, Adamic T, Rozman A (2012) Gap recruitment and partitioning in an old-growth beech forest of the Dinaric Mountains: influences of light regime, herb competition and browsing. For Ecol Manag 285(6):20–28
Dirks BOM, Van-Oijen M, Schapendonk AHCM, Goudriaan J, Wolf J (2002) Temperature sensitivity of photosynthesis in Lolium perenne swards: a comparison of two methods for deriving photosynthetic parameters from in vivo measurements. Photosynthetica 40(3):405–413
Dittmar C, Fricke W, Elling W (2006) Impact of late frost events on radial growth of common beech (Fagus sylvatica L.) in southern Germany. Eur J For Res 125(3):249–259
Duan WB, Li Y, Chen LX (2007) Microenvironmental heterogeneity of near surface layer illumination in the broad-leaved Pinus koraiensis forest gap. Sci Soil Water Conserv 5(6):82–86
Duan WB, Wang J, Li Y (2008) Microclimate characteristics of different size gaps in Pinus koraiensis-dominated broad leaved mixed forests. Chin J Appl Ecol 19(12):2561–2566
Egerton JJG, Banks JCG, Gibson A, Cunningham RB, Ball MC (2000) Facilitation of seedling establishment: reduction in irradiance enhances winter growth of Eucalyptus pauciflora. Ecology 81(5):1437–1449
Forrester JA, Lorimer CG, Dyer JH, Gower ST, Mladenoff DJ (2014) Response of tree regeneration to experimental gap creation and deer herbivory in north temperate forests. For Ecol Manag 329:137–147
Gray AN, Spies TA, Pabst RJ (2012) Canopy gaps affect long-term patterns of tree growth and mortality in mature and old-growth forests in the Pacific Northwest. For Ecol Manag 281(1):111–120
Guan YY, Fei F, Guan QW, Chen B (2016) Advances in studies of forest gap ecology. Sci Silvae Sin 52(4):91–99
Guo R, Zeng R, Zhang JX, Zhang XL, Sun GY (2015) Effects of different light intensities on reactive oxygen species metabolism and chlorophyll fluorescence parameters in leaves of flue-curing tobacco seedlings after chilling stress. Acta Agric Boreali-sin 30:225–230
Han YZ, Wang ZQ (2002) Spatial heterogeneity and forest regeneration. Chin J Appl Ecol 13(5):615–619
Hu WH, Yan XH, Yuan LF, Yang Q, Wu ZY (2011) The role of light intensity in the recovery of photosynthesis in tomato leaves after chilling under low light. B Bot Res 31(2):164–168
Hubbell SP, Foster RB, O′Brien ST, Harms KE, Condit R, Wechsler B, Wright SJ, Loo de Lao S (1999) Light-gap disturbances, recruitment limitation, and tree diversity in a neotropical forest. Science 283:554–557
James G, Landis RM, Ashton MS, Galvão J (2005) Growth response by big-leaf mahogany (Swietenia macrophylla) advance seedling regeneration to overhead canopy release in southeast Pará, Brazil. For Ecol Manag 204:399–412
Jie JM, Yu JH, Huang GB, Jie MH (2008) Effects of light intensity on photoinhibition in pepper seedlings treated under low temperature and poor irradiance. Trans Chin Soc Agric Eng 24(5):231–234
Jin GZ, Liu YY, Liu S, Kim JH (2007) Effect of gaps on species diversity in the naturally regenerated mixed broadleaved-Korean pine forest of the Xiaoxing’an mountains, China. J Ecol Environ 30(4):325–330
Jing X, Duan WB, Chen LX, Wang T, Du S, Zhang YS, Chen QM (2015) Spatial distribution pattern of main populations and gap makers in Picea koraiensis and Abies nephrolepis forest of Xiaoxing′an Mountains, Northeast China. Chin J Appl Ecol 26(10):2928–2936
Jönsson AM, Linderson ML, Stjernquist I, Schlyter P, Bäring L (2004) Climate change and the effect of temperature backlashes causing frost damage in Picea abies. Glob Planet Change 44(1–4):195–207
Laughlin GP, Kalma JD (1990) Frost risk mapping for landscape planning: a methodology. Theor Appl Climatol 42(1):41–51
Li DL, Jin YQ, Xiang QB (2005) Survey of the freezing injury of phoebe chekiangensis seedlings around Nanjing. J Jiangsu For Sci Technol 32(4):12–14
Li M, Duan WB, Chen LX, Liu Y, Gao ZQ (2012) Geostatistical analysis of extreme soil surface temperature of forest gap in Pinus koraiensis dominated broadleaved mixed forest. J Nat Resour 27(10):1688–1695
Li XF, Zhu JJ, Wang QL, Zhang JX, Zhu CY, Liu XF, Liu LM (2013) Research on the disturbance of frost damage to forests. Acta Ecol Sin 33(12):3563–3574
Lindkvist L, Chen D (1999) Air and soil frost indices in relation to plant mortality in elevated complex clear-felled terrain. Clim Res 12(1):65–75
Liu YY, Jin GZ, Li FR (2014) Influence of forest gaps on seedling establishment in a mixed broadleaved-Korean pine (Pinus koraiensis) forest in Xiao Xing′an Mountains. Chin Sci Bull 59(24):2396–2406
Luo Y, Tang HR, Zhang Y (2007) Effect of low temperature stress on activities of SOD and enzymes of ascorbate–glutathione cycle. Acta Hortic Sin 34(6):1405–1410
Muhamed H, Touzard B, Bagousse-Pinguet YL, Michalet R (2013) The role of biotic interactions for the early establishment of oak seedlings in coastal dune forest communities. For Ecol Manag 297(3):67–74
Nagel TA, Svoboda M, Rugani T, Diaci J (2010) Gap regeneration and replacement patterns in an old-growth Fagus–Abies forest of Bosnia–Herzegovina. Plant Ecol 208(2):307–318
Nordlander G, Hellqvist C, Johansson K, Nordenhem H (2011) Regeneration of European boreal forests: effectiveness of measures against seedling mortality caused by the pine weevil Hylobius abietis. For Ecol Manag 262(12):2354–2363
Nunez M, Bowman DMJS (1986) Nocturnal cooling in a high altitude stand of Eucalyptus delegatensis as related to stand density. Aust For Res 16(2):185–197
Rambo TR, North MP (2009) Canopy microclimate response to pattern and density of thinning in a Sierra Nevada forest. For Ecol Manag 257(2):435–442
Ramming A, Jönsson AM, Hickler T, Smith B, Bärring L, Sykes MT (2010) Impacts of changing frost regimes on Swedish forests: incorporating cold hardiness in a regional ecosystem model. Ecol Model 221(2):303–313
Romell E, Hallsby G, Karlsson A (2009) Forest floor light conditions in a secondary tropical rain forest after artificial gap creation in northern Borneo. Agric For Meteorol 149(6–7):929–937
Sane PV, Lvanov AG, Hurry V, Huner NP, Öquist G (2003) Changes in the redox potential of primary and secondary electron-accepting quinones in photosystem II confer increased resistance to photoinhibition in low-temperature-acclimated Arabidopsis. Plant Physiol 132(4):2144–2151
Song Y, Zhang YX, Guo N, Sun GY (2013) Effects of lights on chlorophyll fluorescence characteristics and energy allocation pathways in leaves of Populus simonii × P. nigra seedlings after chilling stress. J Anhui Agric Sci 41(10):4421–4423
Stan AB, Daniels LD (2014) Growth releases across a natural canopy gap-forest gradient in old-growth forests. For Ecol Manag 313:98–103
Tan JJ (2013) Physiological and biochemical responses of Pinus massoniana superior provenance seedlings under artificial low temperature stress. Sci Silvae Sin 49(3):51–55
Tang JM, Zhai MP (2006) Morphological response and biomass allocation of Schima superb seedlings at different habitats in gap. J Huazhong Agric Univ 25(5):559–563
Tian Y, Zheng GY, Zhang HH, Xu N, Wang J, Sun GY (2013) Effects of light intensity and temperature on chlorophyll fluorescence in leaves of basil seedlings. Pratac Sci 30(10):1561–1568
Verduin JJ, Seidlitz A, Keulen MV, Paling EI (2013) Maximising establishment success of Amphibolis antarctica seedlings. J Exp Mar Biol Ecol 449(1770):57–60
Wang BY, Feng YL (2005) Effects of growth light intensities on photosynthesis in seedlings of two tropical rain forest species. Acta Ecol Sin 25(1):23–30
Wang N, Wu J, Xia PY, Su JL (2011) Physiological responses of Ilex latifolia to low temperature and its cold tolerance. J South China Agric Univ 32(3):82–86
Wang Y, Liu J, Zhang JX, Li XF, Zhu CY, Zhao TB, Yue Y, Zhan SY (2015) Spatial distribution characteristics of photosynthetic photon flux density and air temperature in gaps at two different slope directions of secondary forest ecosystem in montane area of eastern Liaoning Province. Chin J Ecol 34(4):933–940
Wu J, Wang ZQ, Liu XF (2002) Frost injury to young Tilia amurensis plantation. J Northeast For Univ 30(6):6–10
Yan T, Lü XT, Yang K, Zhu JJ (2016) Leaf nutrient dynamics and nutrient resorption: a comparison between larch plantations and adjacent secondary forests in Northeast China. J Plant Ecol 9:165–173
Yang K, Shi W, Zhu JJ (2013) The impact of secondary forests conversion into larch plantations on soil chemical and microbiological properties. Plant Soil 368(1–2):535–546
Zenner EK, Sagheb-Talebi K, Akhavan R, Peck JLE (2015) Integration of small-scale canopy dynamics smoothes live-tree structural complexity across development stages in old-growth Oriental beech (Fagus orientalis Lipsky) forests at the multi-gap scale. For Ecol Manag 335(2):26–36
Zhang YD (1992) Liaodong area meteorological disasters and disaster mitigation strategies. Meteorological Press, Beijing, pp 83–98
Zhang YP, Wang JX, Ma YX, Liu YH, Li YR (2002) The temporal-spatial distribution of temperature on the surface roughness of the tropical secondary forest gap in Xishuangbanna, Yunnan Province. Sci Silvae Sin 38(6):1–5
Zhou Q, Wang JZ, Chen XH (2010) Effect of durative low temperature on morphological and physiological characteristics of cucumber seedling. North Hortic 16:1–3
Zhu JJ, Mao ZH, Hu LL, Zhang JX (2007) Plant diversity of secondary forests in response to anthropogenic disturbance levels in montane regions of northeastern China. J For Res 12(6):403–416
Zhu JJ, Tan H, Li FQ, Chen M, Hu LL (2009) Comparison of near-ground air temperature and soil temperature of summer within three gaps of different sizes at secondary forests in Eastern Montane Regions of Liaoning Province. Sci Silvae Sin 45(8):161–165
Zhu JJ, Zhang GQ, Wang GF, Yan QL, Lu DL, Li XF, Zheng X (2015) On the size of forest gaps: can their lower and upper limits be objectively defined? Agric For Meteorol 213(2):64–76
Zhu CY, Wang Y, Li XF, Liu J, Zhang JX, Wang T (2016) Spatiotemporal distribution of the minimum temperature during autumn at different slope directions of secondary forest gap in montane zone of eastern Liaoning Province. Chin J Ecol 35(6):1411–1419
Acknowledgements
This work was supported by the National Key Research and Development Program of China (2016YFC0500302) and the National Nature Scientific Foundation Project of China (31200432). We thank Dr. Li-ning Song, Dr. Qiao-ling Yan, Dr. Kai-Yang and Ming-cai Li for their helpful suggestions on the earlier versions of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Project funding: This work was supported by the National Key Research and Development Program of China(2016YFC0500302)and the National Nature Scientific Foundation Project of China (31200432).
The online version is available at http://www.springerlink.com
Corresponding editor: Chai Ruihai.
Rights and permissions
About this article

Cite this article
Li, X., Wen, Y., Zhang, J. et al. The effect of low-temperature event on the survival and growth of Juglans mandshurica seedlings within forest gaps. J. For. Res. 29, 943–951 (2018). https://doi.org/10.1007/s11676-017-0520-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-017-0520-6