
Abstract
Anatomical characteristics have been proven useful for extracting climatic signals. To examine the climatic signals recorded by tree-ring cell features in the Changbai Mountains, we measured cell number and cell lumen diameter, in addition to ring widths, of Korean pine (Pinus koraiensis) tree rings at sites of varied elevation, and we developed chronologies of cell number (CN), mean lumen diameter (MLD), maximum lumen diameter (MAXLD) and tree-ring width (TRW). The chronologies were correlated with climatic factors monthly mean temperature and the sum of precipitation. As shown by our analysis, the cell parameter chronologies were suitable for dendroclimatology studies. CN and TRW shared relatively similar climatic signals which differed from MLD and MAXLD, and growth-climate relationships were elevation-dependent, as shown by the following findings: (1) at each elevation, MLD and MAXLD recorded different monthly climatic signals from those recorded by TRW for the same climatic factors; and (2) MLD and MAXLD recorded climatic factors that were absent from TRW at lower and middle elevations. Cell lumen diameter proved to be an effective archive for improving the climate reconstruction for this study area.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Tree rings are known to be a useful archive for studying climate variability (Esper et al. 2002; Cook et al. 2004). The dendroclimatology method uses tree-ring variables to examine radial growth-climate relationships by correlation analysis (Fritts 1974; Gagen et al. 2006; Lebourgeois et al. 2012). Ring width, because it easily can be measured and cross-dated, has widely been used as a climate proxy since tree-ring dendrochronology principles and methods were first established (Fritts 1976). Tree-ring width is a measure of the sum of a growing season’s cell division and cell enlargement. The width of rings is primarily a function of three basic growth processes, viz. cell division, cell enlargement and cell differentiation (Vaganov et al. 2006). Thus, anatomical features such as cell number, cell size and cell wall thickness may carry higher-resolution climatic information than ring widths (Kirdyanov et al. 2003; Panyushkina et al. 2003) and therefore are of importance in examining growth-climate relationships.
The use of cell features in dendroclimatology is strongly related to the development of the applicable acquisition methods (Fonti et al. 2010). Initially, researchers used field monitoring to record changes in cell features and climate over a given period to analyze the relationships between cell features and climate (Knigge and Schulz 1961; Fritts 1966; Denne 1974; Ford et al. 1978; Antonova and Stasova 1993; Camarero et al. 1998). With the development of computerized image-analysis systems, more tree growth-climate relationships were investigated at the cellular scale (Yasue et al. 2000; Wang et al. 2002; Kirdyanov et al. 2003; Eilmann et al. 2006, 2009; Drew et al. 2012; Olano et al. 2012; Noval et al. 2013). In China, some studies attempted to examine the climatic information carried by the cell features of tree rings (Li et al. 2008; Wang et al. 2011; Xu et al. 2015). All of these studies reported that the cell feature variables carried more climatic information than did ring widths. Dendroclimatology studies using cell anatomical characteristics are in their initial developmental stages in China but have bright prospects.
The Changbai Mountains, a region in northeastern China where climate warming became obvious early on with a warming rate of about 0.3 °C (10a)−1 (Guo et al. 2005; Dong and Wu 2007; Zhao et al. 2009), are of importance in climate reconstructions for China. They include a large area of undisturbed temperate old-growth forests (Shao and Zhao 1998). Korean pine (Pinus koraiensis) is the main coniferous species in the Changbai Mountains and many tree-ring studies have reported ring widths of P. koraiensis for this area (Shao and Wu 1997; Zhu et al. 2009; Gao et al. 2011; Yu et al. 2011; Wang et al. 2013). These studies concluded that temperature is the main climatic factor limiting radial growth. One study based on cell feature variables found that cell size carried a more strongly correlated precipitation signal than ring-width (Wang et al. 2011). Additionally, elevation is one of the main factors to affect tree radial growth, and many dendroclimatology studies have reported that tree growth-climate relationships were affected by or strongly dependent on elevation (Brookhouse and Bi 2009; Zhang et al. 2012). The Changbai Mountains are characterized by vertical vegetation zonation (Editorial Committee for Forestry of Jilin 1988; Hao et al. 2007). Previous studies found that growth-climate relationships were elevation dependent for Larix olgensis (Yu et al. 2005; Chen et al. 2011) and Picea jezoensis (Yu et al. 2006), the other two dominant coniferous species in the Changbai Mountains. Korean pine might also have an elevation-dependent growth-climate relationship.
Therefore, we examined Korean pine tree-ring samples from various elevations of the Changbai Mountains to (1) identify the consistent climatic signals contained in cell parameter chronologies and (2) quantify the effect of elevation on tree-growth relationships indicated by cell parameters.
Materials and methods
Study area and sampling sites
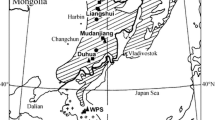
Our study area was on the northern slope of Changbai Mountain National Nature Reserve in northeastern China, in Jilin Province (42° to 42°45′E, 127°30′ to 128°30′N). The area attains its highest elevation of 2189 m above sea level (asl) at Tianchi Lake (Fig. 1). This area has not been extensively affected by anthropogenic activity, because Changbai Mountain did not become a tourist attraction until in the 1980s and was then designated a protected nature reserve (Yu et al. 2012). This area is characterized by a temperate continental monsoon climate (Editorial Committee for Forestry of Jilin 1988). Based on the average meteorological data from the Donggang and Erdao stations (data from the National Meteorological Information Center of China, Fig. 1), the mean annual temperature was 3.7 °C, with a mean temperature of 15.6 °C in January and 20.1 °C in July during the period 1971–2000 (Fig. 2). Mean annual precipitation was 811 mm, and mainly occurred from May to September. The vegetation of the area showed typical vertical zonation. Korean pine and broad-leaved mixed forest extended from 500 to 1100 m asl. A spruce-fir forest continued from 1100 to 1800 m asl. Soils in the coniferous forest zone are mountainous brown forest soil (Editorial Committee for Forestry of Jilin 1988).

Locations of tree-ring sampling sites and meteorological stations in the study area

Long-term monthly mean temperature (line/squares) and precipitation (bars) at the Donggang and Erdao stations, based on data for the period 1971–2000
Tree-ring cores were sampled from three sites designated Site 1, Site 2 and Site 3 (Fig. 1) at three elevations (Table 1). Site 1 and Site 2 were located at the lower and upper elevational range of distribution of Korean pine and broad-leaved mixed forest, respectively. Site 3, at 1258 m asl, sampled spruce-fir forest. All sites were located on a north-facing slope of 0–10. Approximately 30 of the largest and presumably oldest trees were selected at each site for sampling. Using increment borers two cores were extracted at breast height from each tree.
Laboratory work and chronology development
All cores were air-dried and then mounted and sanded to produce clearly visible tree-ring boundaries. They were visually cross-dated (Stokes and Smiley 1968) under stereoscopes. Total ring width was measured using a LINTAB-station (Frank Rinn, Heidelberg, Germany) at 0.01 mm resolution. The COFECHA program (Holmes 1983) was used to check the quality of cross-dating and measurements. Based on the statistics from COFECHA, only individual samples with consecutive segments that correlated well with the master series and had high sensitivity were included in development of cell parameter chronologies. Twelve to 15 cores of different trees were selected for cell parameters measurements. The number of samples was sufficient to obtain credible cell features (Fonti et al. 2009). These cores were further polished to make cell boundaries clearly visible for identification. We used the MIVNT image analysis system (Guilin, Guangxi, China) to measure cell number and cell lumen diameter. The software is widely used to process and analyze images in various fields, such as material science, biology and chemistry. Cell numbers and lumen diameters were measured along each of five radial files of each growth ring. To allow direct comparisons of cell dimensions patterns between rings with different cell numbers, we converted all size measurement series to the average numbers of all annual rings using the method of Vaganov (1990). The cell variable values were averaged within each tree ring to develop chronologies.
Tree-ring chronologies were developed using the ARSTAN program (Cook 1985). A cubic smoothing spline with 67 % of the series length was used to remove the age-related growth trends of raw ring-width chronologies and cell number chronologies. Lumen diameter chronologies have no obvious long-term growth trends; therefore, they were standardized using straight lines ((value-mean)/mean) (Conkey 1986). The resulting ratio series was then computed as a biweight robust mean of the detrended and standardized individual series (Cook et al. 1990). Finally, the chronologies of ring-width (TRW), cell number (CN), maximum lumen diameter (MAXLD) and mean lumen diameter (MLD) were constructed. To show the strength of common signals in the chronologies, we performed a within-chronology common interval analysis for each chronology. The statistical quality of each chronology was measured using several coefficients commonly used in dendrochronology, including standard deviation (SD), mean sensitivity (MS), first-order autocorrelation (AC1), mean correlation between trees (R), variance in the first principal component (PC1) (Fritts 1976) and the expressed population signal (EPS) (Wigley et al. 1984). The EPS indicates the chronology confidence level (Briffa and Jones 1990), and a threshold value of 0.85 is often used to evaluate the useful time span of the final chronologies (Wigley et al. 1984). Three types of chronologies (residual, standard or arstan) were established (Cook and Holmes 1986), and the standard chronologies were used to analzye growth-climate relationships. To measure the similarity between any two chronologies, we used Pearson correlation analysis and principal component analysis for the common time span from 1958 to 2002.
Climatic data
Climatic data were obtained from the Erdao (591 m asl), Donggang (774 m asl) and Tianchi (2623 m asl) meteorological stations near the sites in the study area (Fig. 1). The Tianchi station data were not used because winter observations were terminated in 1989. The monthly mean temperature and precipitation correlation coefficients of Erdao and Donggang stations in corresponding months from 1958 to 2007 were 0.95 and 0.79, respectively. Therefore, we averaged the meteorological data from the Erdao and Donggang stations to improve the regional representation, and we used the average values to analyze the growth-climate relationships.
Analysis of tree growth-climate relationships
Pearson correlation analysis was used to identify the growth-climate relationship (Fritts 1974). Because the width of an annual ring can be affected by climatic conditions over a long period, the climatic conditions in the prior year may affect the growth in the current year (Fritts 1976). Thus, the monthly mean temperature and total precipitation from the prior September to the current August were included in the analysis. Meteorological data were limited from 1958 to 2007, therefore we analyzed the growth-climate relationship for the period 1959–2007 at Site 1 and Site 3 and for the period 1959–2002 at Site 2.
Results
Chronology characteristics
The statistics of the cell chronologies of each site were lower in value than were those for the TRW, especially for MLD and MAXLD (Table 2). The SD and MS values, which indicate the dispersion degree of indices and the annual variability, respectively, were especially low.
The characteristics of the chronology changed with elevation. MS, SD and EPS were highest for the Korean pine chronology at Site 1 and smallest at Site 2. The result means that the annual variability of ring variables and the population signal of the sample declined with increasing elevation. The AC1 of the chronologies at Site 2, was lowest, particularly for the MAXLD, and there were fewer common signals at Site 2 chronologies with lower R and PC1.
The annual variation of indices revealed the correlations between the chronologies of each variable (Fig. 3). Synchronous behavior of the variables at Sites 1 and 2 was visible in specific pointer years (e.g., 1965, 1974, 1975, 1983, 1990 and 1994). The correlation coefficients shown in Table 3 illustrated this phenomenon. The correlation coefficients of the chronologies for every variable except MAXLD were greater than 0.439 (P < 0.01) at Sites 1 and 2. The similarities between Site 3 and the other two sites were very low for all variables, except for the correlation coefficient of MLD between Sites 2 and 3, which was 0.321 (P < 0.05).

Annual variations of ring width (TRW), cell number (CN), mean lumen diameter (MLD) and maximum lumen diameter (MAXLD) during 1958–2007 for Sites 1 and 3 and during 1958–2002 for Site 2
The chronologies of all variables were compared at each site to determine whether they contained the same kind of information (Table 4). Generally, TRW and CN were very closely related, displaying a correlation higher than 0.96 (P < 0.01) at all three sites. The correlations were also high between MLD and MAXLD with the value higher than 0.60 (P < 0.01). However, the correlations were lower between cell size chronologies and TRW or CN, although some were significant. This result can be best expressed by principal component analysis at each site (Fig. 4), which sorts the variables according to their correlations. The two first principal components retained more than 90 % of the total variance. Ordinations along the two axes all showed that TRW and CN were significantly different from the two cell size variables.

Scatter plots of principal component analysis loading of the four chronologies of each site for Pinus koraiensis plotted along the axes of the two first principal components (the two first principal components retained more than 90 % of the total variance)
Relationships between cell variable chronologies and climatic factors
Climatic signals recorded by cell lumen diameters
MLD and MAXLD were correlated with monthly mean temperature and the sum of precipitation (Fig. 5). At Site 1, MLD was positively correlated with March temperature while MAXLD was not. Both of these chronologies were negatively and significantly correlated with May temperature. MLD and MAXLD were both positively correlated with precipitation in May, and MLD was highly correlated with June precipitation in the current year. MLD and MAXLD were positively correlated with March temperature and June precipitation at Site 2; additionally, MLD at this site was positively correlated with July precipitation. At the highest elevation, MLD and MAXLD only reflected the temperature signal and were positively correlated with the previous October or current June temperatures.

Correlation coefficients between monthly climate elements and tree-ring indices during 1959–2007 for Sites 1 and 3 (1959–2002 for Site 2). Sizes of the circles indicate the strengths of the correlations. P9 the previous September, P10 the previous October, P11 the previous November, P12 the previous December, T temperature, P precipitation
Comparison of the growth-climate relationship for cell variables and ring widths
The relationships at each site between CN and climatic factors were similar to the relationships between TRW and climatic factors (Fig. 5). However, the relationships between lumen diameters or ring widths and climate factors revealed that lumen diameters can record different climatic signals from those recorded by ring widths.
At Site 1, TRW was not related to monthly temperature; it was positively correlated with precipitation in the previous September and in the current June. This result was different from that found for the precipitation signal recorded by MLD and MAXLD. At Site 2, TRW was positively correlated with April temperature, contrasting with the temperature signals recorded by MLD and MAXLD. TRW at this site was unrelated to precipitation. TRW at Site 3 was positively correlated with July temperature and precipitation during the previous September when there was no precipitation signal in MLD or MAXLD.
Discussion
Evaluation of the chronology characteristics of cell variables
The established chronology characteristics of the cell variables showed substantially lower sensitivities and common signals than did ring widths. This phenomenon was reported by others (Yasue et al. 2000; Wang et al. 2002; Corcuera et al. 2004). The common signal indicates the statistical quality of each chronology, so growth ring width is more suitable for dendroclimatology study. Ring width cannot, however, inform regarding analogies or differences in the ecological information that the chronology records. The identification of such information requires a comparison of the chronologies with each other and with climate data (Wimmer and Grabner 2000; Fonti and García-González 2004). Previous studies on the mean vessel area of ring-porous species showed mean sensitivities and correlations between samples ranging from 0.05 to 0.20 and from 0.2 to 0.4, respectively (Tardif and Conciatori 2006; Fonti et al. 2007). Nevertheless, the correlations with climatic parameters reached values >0.60 in certain cases. For coniferous species, Yasue et al. (2000) found that the mean sensitivity and correlation between samples of Picea glehnii cell wall thickness were 0.05 and 0.14, respectively, but 37 % of the variance could be explained by climatic factors. Although the statistical quality of MLD was lower than that for TRW at Site 1 in our study, MLD was positively correlated with March and May temperature that could not be recorded by TRW. Similar results were recorded for Sites 2 and 3. In order to improve the statistical quality of cell variables, we suggest to increase sample sizes. For example, when the sample size of MLD at Site 1 increased to 16 cores, EPS improved to 0.85.
The chronologies of TRW and CN showed almost the same relationships with climatic factors at all three elevations. This suggests that TRW was correlated with CN, meaning that ring width depended primarily on cell numbers, a common phenomenon in coniferous species. Camarero et al. (1998) reported that the ring widths and cell numbers of Pinus uncinata Ram. and Pinus sylvestris L. in the Central Spanish Pyrenees were functionally correlated (P < 0.001). The correlation coefficient of these two variable chronologies was 0.78 for Larix cajanderi. (P < 0.05), as calculated by Panyushkina et al. (2003), and 0.907 for Picea crassifolia (P < 0.01), as calculated by Li et al. (2008). This phenomenon can be attributed to the processes governing cell anatomy; Cuny et al. (2012) monitored the intra-annual wood formation dynamics of three coniferous species and estimated that approximately 75 % of the annual radial increment variability was attributable to the rate of cell production, whereas only 25 % was attributable to its duration. For this reason, cell number has been used relatively infrequently to analyze the growth-climate relationship (Deslauriers and Morin 2005).
Limiting climatic factors for the radial growth of Korean pine at different elevations
The combination of the relationships between cell variables/ring widths and climatic factors showed that both temperature and precipitation were limiting factors for the radial growth of Korean pine in the study area.
March temperature and previous September precipitation were the common climatic factors limiting tree growth at all three sites. MLD was positively correlated with March temperature because a high temperature before the growing season can promote cambial activity and can improve photosynthesis, both of which are positively related to cell enlargement. Greater precipitation in September leads to greater soil-moisture availability and this is advantageous to Korean pine growth in the following year.
The low correlations of each variable chronology between sites (Table 3) indicated that there were also differences in the growth-climate relationship related to elevation. At the low elevation site, TRW was also limited by precipitation in the current June. This finding is consistent with the results of Yu et al. (2011). MLD and MAXLD can record May precipitation because the largest cells are formed during the early growing season and these are a more reliable proxy for climatic conditions during that period than is TRW. At the middle elevation site, MLD and MAXLD correlated well with June precipitation, complementing the climatic signals furnished by TRW. At the high elevation site, July temperature was the limiting factor for ring width. This finding verifies the conclusion of Gao et al. (2011). With increasing elevation, lower temperatures can delay the onset of the growth period and terminate growth before the end of the growing season at lower elevations. MLD at this elevation was significantly correlated with June temperature. This verified that cell lumen diameter can record climate factors for earlier months. Nevertheless, at high elevation, lumen diameter chronologies did not respond significantly to the precipitation signal. The reason for this outcome might be the increase in precipitation at this elevation.
Since the MLD and MAXLD supplied different climatic information from that yielded by TRW, they can improve climate reconstruction in the Changbai Mountains. However, because the climatic data used to analyze the growth-climate relationships were expressed on a monthly basis, it cannot be determined whether lumen diameters in this area can offer a higher temporal resolution than ring widths. In further studies, climatic data at a scale of days or pentads should be used to examine the climatic signals recorded by cell features. Additionally, mechanistic models such as TREERING (Fritts et al. 1999), which uses daily climatic data to simulate processes affecting the formation of cell features, may be effective methods for studying the growth-climate relationship at the cellular scale.
Conclusions
It is possible to build variable chronologies for cells of Korean pine in the Changbai Mountains for dendroclimatology studies. Although the statistics of the cell lumen diameter chronologies were lower in value than the statistics of the ring-width chronologies, they can record different climatic signals from those encoded in ring widths. Both temperature and precipitation are limiting factors for Korean pine radial growth, and the growth-climate relationships change with elevation. The significance of cell features in reconstruction of spring temperature at low elevation, summer precipitation at middle elevation, and autumn temperature at high elevation of the Changbai Mountains has been demonstrated, but the relationship between cell features and climate should be further investigated to assess the climatic reconstruction potential offered by cell features.
References
Antonova GF, Stasova VV (1993) Effects of environmental factors on wood formation in Scots pine stems. Trees 7:214–219
Briffa KR, Jones PD (1990) Basic chronology statistics and assessment. In: Cook EA, Kairiukstis LA (eds) Methods of dendrochronology: applications in the environmental sciences. Kluwer Academic Publishers, Dordrecht, pp 137–152
Brookhouse MT, Bi H (2009) Elevation-dependent climate sensitivity in Eucalyptus pauciflora Sieb. ex Spreng. Trees 23:1309–1320
Camarero JJ, Guerrero-Campo J, Gutiérrez E (1998) Tree-ring growth and structure of Pinus uncinata and Pinus sylvestris in Central Spanish Pyrenees. Arct Antarct Alp Res 30:1–10
Chen L, Wu SH, Pan T (2011) Variability of climate-growth relationships along an elevation gradient in the Changbai Mountain, northeastern China. Trees. doi:10.1007/s00468-011-0588-0
Conkey LE (1986) Red sprue tree-ring width and densities in eastern North America as indicators of past climate. Quat Res 26:232–243
Cook ER (1985) A time series approach to tree-ring standardization. Ph.D. thesis, University of Arizona
Cook ER, Holmes RL (1986) User’s manual for Program ARSTAN (chronology series VI). University of Arizona
Cook ER, Briffa KR, Shiyatov SG, Mazepa VS (1990) Tree-ring standardization and growth-trend estimation. In: Cook ER, Kairiukstis LA (eds) Methods of dendrochronology: applications in the environmental sciences. Kluwer Academic Publishers, Dordrecht, pp 104–123
Cook ER, Woodhouse CA, Eakin CM, Meko DM, Stahle DW (2004) Long-term aridity changes in the western United States. Science 306:1015–1018
Corcuera L, Camarero JJ, Gil-Pelegrin E (2004) Effects of a severe drought on Quercus ilex radial growth and xylem anatomy. Trees 18:83–92
Cuny HE, Rathgeber CBK, Lebouegeois F, Fortin M, Fournier M (2012) Life strategies in intra-annual dynamics of wood formation: example of three conifer species in a temperate forest in north-east France. Tree Physiol 32:612–625
Denne MP (1974) Effect of light intensity on tracheid dimensions in Picea sitchensis. Ann Bot 38:337–345
Deslauriers A, Morin H (2005) Intra-annual tracheid production in balsam fir stems and the effect of mereorogical variables. Trees 19:402–408
Dong MY, Wu ZF (2007) Variations trend of annual mean temperature and precipitation for the last 50 years in Jilin Province. J Northeast Normal Univ (Nat Sci) 39:114–119
Drew DM, Allen K, Downes GM, Evans R, Battaglia M, Baker P (2012) Wood properties in a long-lived conifer reveal strong climate signals where ring-width series do not. Tree Physiol. doi:10.1093/treephys/tps111
Editorial Committee for Forestry of Jilin (1988) Forest of Jilin. China Forestry Press, Beijing
Eilmann B, Weber P, Rigling A (2006) Growth reactions of Pinus sylvestris L. and Quercus pubescens Willd. to drought years at a xeric site in Valais,Switzerland. Dendrochronologia 23:121–132
Eilmann B, Zweifel R, Buchmann N, Fonti P, Rigling A (2009) Drought-induced adaptation of xylem in Scots pine and pubescent oak. Tree Physiol 29:1011–1020
Esper J, Cook ER, Schweingruber FH (2002) Low-frequency signals in long tree-ring chronologies for reconstructing past temperature variability. Science 295:2250–2253
Fonti P, García-González I (2004) Suitability of chestnut earlywood vessel chronologies for ecological studies. New Phytol 163:77–86
Fonti P, Solomonoff N, García-González I (2007) Earlywood vessels of Castanea sativa record temperature before their formation. New Phytol 173:562–570. doi:10.1111/j.1469-8137.2006.01945.x
Fonti P, Eilmann B, García-González I, von Arx G (2009) Experditious building of ring-porous earlywood vessel chronologies without loosing signal information. Trees 23:665–671
Fonti P, von Arx G, Garcia-Gonzalez I, Eilmann B, Sass-Klaassen U, Gärtner H, Eckstein D (2010) Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytol 185:42–53
Ford ED, Robards AW, Piney MD (1978) Influence of environmental factors on cell production and differentiation in the earlywood of Picea sitchensis. Ann Bot 42:683–692
Fritts HC (1966) Growth rings of trees: their correlation with climates. Science 154:973–979
Fritts HC (1974) Relationships of ring-widths in arid-site conifers to variations in monthly temperature and precipitation. Ecol Monogr 44:411–440
Fritts HC (1976) Tree rings and climate. Academic Press, London
Fritts HC, Shashkin AV, Downes GM (1999) A simulation model of conifer ring growth and cell structure. In: Wimmer R, Vetter RE (eds) Tree-ring analysis. Cambridge University Press, Cambridge, pp 3–32
Gagen M, McCarroll D, Edouard JL (2006) Combining ring width, density and stable carbon isotope proxies to enhance the climate signal in tree-rings: an example from the southern French Alps. Clim Change 78:363–379
Gao LS, Wang XM, Zhao XH (2011) Response of Pinus koraiensis and Picea jezoensis var. komarovii to climate in the transition zone of Changbai Mountain,China. China J Plant Ecol 35:27–34
Guo ZM, Miao QL, Li X (2005) Variation characteristics of temperature over north-ern China in recent 50 years. Sci Geograph Sin 25:448–454
Hao ZQ, Zhang J, Song B, Ye J, Li BH (2007) Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest. For Ecol Manag 252:1–11
Holmes RL (1983) Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull 43:69–78
Kirdyanov AV, Hughes M, Vaganov EA, Schweingruber FH, Silkin P (2003) The importance of early summer temperature and date of snow melt for tree growth in the Siberian Subarctic. Trees 17:61–69
Knigge W, Schulz H (1961) Einfluss der Jahreswitterung 1959 auf Zellartenverteilung, Faserlänge und Gefässweite verschiedener Holzarten. Holz Roh Werkst 19:293–303
Lebourgeois F, Mérian P, Courdier F, Ladier J, Dreyfus P (2012) Instablility of climate signal in tree-ring width in Mediterranean mountains: a multi-species analysis. Trees 26:715–729
Li Y, Liang EY, Shao XM (2008) Variations of Picea crassifolia tree-ring cell structure and their impliacations to past climate in eastern margin of Qaidam Basin, Northwest China. China J Plant Ecol 19:524–532
Noval K, Luǐs MD, Raventós J, Ćufar K (2013) Climatic signals in tree-ring widths and wood structure of Pinus halaepensis in contrasted environmental conditions. Trees 27:927–936
Olano JM, Eugenio M, García-Cervigón AI, Folch M, Rozas V (2012) Quantitative tracheid anatomy reveals a complex environmental control of wood structure in continental Mediterranean climate. Int J Plant Sci 173:137–149
Panyushkina IP, Hughes MK, Vanganov EA, Munro MAR (2003) Summer temperature in northeastern Siberia since 1642 reconstructed from tracheid dimensions and cell numbers of Larix cajanderi. Can J For Res 33:1905–1914
Shao XM, Wu XD (1997) Reconstruction of climate change on Changbai Mountain, Northeast China using tree-ring data. Quat Sci 1:76–85
Shao G, Zhao G (1998) Protecting versus harvesting of old-growth forests on the Changbai Mountain (China and North Korea): a remote sensing application. Nat Areas J 18:358–365
Stokes MA, Smiley TL (1968) An introduction to tree-ring dating. University of Chicago Press, Chicago
Tardif JC, Conciatori F (2006) Influence of climate on tree ring and vessel features in red oak and white oak growing near their northern distribution limit, southwestern Quebec. Can J For Res 36:2317–2330. doi:10.1139/X06-133
Vaganov EA (1990) The tracheidogram method in tree-ring analysis and its application. In: Cook ER, Kairiukstis LA (eds) Methods of dendrochronology: applications in the environmental sciences. Kluwer Academic Publishers, Netherlands, pp 63–67
Vaganov EA, Hughes MK, Shashkin AV (2006) Growth dynamics of conifer tree rings: images of past and future environments. Springer, Berlin, Heidelberg
Wang LL, Payette S, Bégin Y (2002) Relationships between anatomical and densitometric characteristics of black spruce and summer temperature at tree line in northern Quebec. Can J For Res 32:477–486
Wang H, Shao XM, Fang XQ, Yin ZY, Chen L, Zhao DS, Wu SH (2011) Responses of Pinus koraiensis tree ring cell scale parameters to climate elements in the Changbai Mountains. China J Plant Ecol 22:2643–2652
Wang H, Shao XM, Jiang Y, Fang XQ, Wu SH (2013) The impacts of climate change on the radial growth of Pinus koraiensis along elevations of Changbai Mountain in northeastern China. For Ecol Manag 289:333–340
Wigley TML, Briffa KR, Jones PD (1984) On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J Clim Appl Meteorol 23:201–213
Wimmer R, Grabner M (2000) A comparison of tree-ring features in Picea abies as correlated with climate. IAWA J 12:403–416
Xu JM, Lu JX, Evans R, Downes GM (2015) Climatic signal in cellulose microfibril angle and tracheid radial diameter of Picea crassifolia at different altitudes of Tibetan plateau, northwest China. Wood Sci Technol. doi:10.1007/s00226-015-0753-5
Yasue K, Funada R, Kobayashi O, Ohtani J (2000) The effects of tracheid dimensions on variations in maximum density of Picea glebnii and relationships to climatic factors. Trees 14:223–229
Yu DP, Wang SZ, Tang LN, Dai LM, Wang QL, Wang SX (2005) Relationship between tree ring chronology of Larix olgensis in Changbai Mountains and the climate change. Chin J Appl Ecol 16:14–20
Yu DP, Wang QL, Wang GG, Dai LM (2006) Dendroclimatic response of Picea jezoensis along an altitudinal gradient in the Changbai Mountains. Sci China Ser E 49:150–159. doi:10.1007/S11434-006-8116-0
Yu DP, Wang QW, Wang Y, Zhou WM, Ding H, Fang XM, Jiang SW, Dai LM (2011) Climatic effects on radial growth of major tree species on Changbai Mountain. Ann For Sci 68:921–933
Yu DP, Liu JQ, Lewis BJ, Zhou L, Zhou WM, Fang XM, Wei YW, Jiang SHW, Dai LM (2012) Spatial variation and temporal instability in the climate–growth relationship of Korean pine in the Changbai Mountain region of Northeast China. For Ecol Manag. doi:10.1016/j.foreco.2012.06.032
Zhang WT, Jiang Y, Dong MY, Kang MY, Yang HC (2012) Relationship between the radial growth of Picea meyeri and climate along elevations of the Luyashan Mountain in North-Central China. For Ecol Manag 265:142–149
Zhao CY, Ren GY, Zhang YF, Wang Y (2009) Climate change of the Northeast China over the past 50 years. J Arid Land Resour Environ 23:25–30
Zhu HF, Fang XQ, Shao XM, Yin ZY (2009) Tree ring-based February–April temperature reconstruction for Changbai Mountain in Northeast China and its implication for East Asian winter monsoon. Clim Past 5:661–666
Acknowledgments
This research was supported by the National Science-Technology Support Plan Projects (2012BAC19B02) and the Beijing Natural Science Foundation Projects (8154046).
Author information
Authors and Affiliations
Corresponding author
Additional information
Project founding: This research was supported by the National Science-Technology Support Plan Projects (2012BAC19B02) and the Beijing Natural Science Foundation Projects (8154046).
The online version is available at http://www.springerlink.com
Corresponding editor: Chai Ruihai
Rights and permissions
About this article

Cite this article
Wang, H., Shao, X., Fang, X. et al. Relationships between tree-ring cell features of Pinus koraiensis and climate factors in the Changbai Mountains, Northeastern China. J. For. Res. 28, 105–114 (2017). https://doi.org/10.1007/s11676-016-0292-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-016-0292-4