
Abstract
This study describes the different parameters used to derive the allometric equation for calculating the biomass of an invasive woody shrub Lantana camara L. from the subtropical conditions of western Himalaya. It identifies the most accurate and convenient method for biomass calculation by comparing destructive with non-destructive methodology. Different parameters were measured on a wide range of Lantana from different community levels for the non-destructive calculation of total above-ground biomass. Different explanatory variables were identified and measured such as basal diameter either as a single independent variable or in combination with plant height. The other suitable combinations of available independent variables include crown length, crown width, crown area, crown volume and coverage of the plant. Amongst the wide range of allometric equations used with different variables, the equation with D2H as a variable was found to be the most suitable estimator of biomass calculation for Lantana. Sahastradhara, being the most disturbed area due to its high tourist activity round the year, showed maximum coverage (58.57 % ha−1), highest biomass (13,559.60 kg ha−1) and carbon density (6,373.01 kg ha−1) of Lantana. The degree of Lantana’s invasiveness in subtropical conditions was also calculated on the basis of importance value index (IVI). The maximum IVI (22.77) and mean coverage (26.8 % ha−1) was obtained from the areas near Jolly Grant airport, indicating that physically disturbed areas are more suitable for the growth of Lantana, which may significantly increase shrub biomass. The importance of incorporating allometric equations in calculation of shrub biomass, and its role in atmospheric carbon assimilation has thus been highlighted through the findings of this study.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Shrubs are a type of vegetal group with a specific structure and appearance (de la Torre 1990). These woody non-trees are of great importance to the biophysical and biodynamic processes of different ecosystems (Di Castri et al. 1981). The exotic plant species vary from their native counterpart, based on requirements such as modes of resource acquisition and more consumption which may ultimately result in changing their soil structure, soil profile, rate of decomposition, nutrient content and moisture. It results in noteworthy unwelcome impacts on the biodiversity and ecosystems. Thus, invasive species are a serious obstruction for conservation and sustainable use of biodiversity. Biological invasions now operated on global level and it will undergo rapid increase due to increasing globalization of markets, rise in global trade, travel and tourism (Eisworth et al. 2005). One such species is Lantana camara L. (henceforth lantana), native to tropical America, this invasive shrub has naturalized in most suitable habitats of tropical and subtropical Africa, Asia, and Australia. Lantana can grow up to 1.8 m in height with an equal spread depending on cultivar. The impacts of lantana infestations range widely across landuse situations, affecting individuals, landholders and land managers, locals, state and federal governments, community groups, and industry.
In natural ecosystems, lantana infests forest edges, coastal zones and riparian areas, penetrates disturbed rainforest and invades open eucalypt woodland. Its dense thickets exclude native species through smothering and allelopathic effects (i.e., toxicity to other plants), dominate understoreys, and reduce biodiversity. Lantana thickets increase the intensity of wildfires, which can have disastrous effects on native flora and fauna. The forests and fallow lands of Doon Valley is heavily infested by this invasive shrub due to many reasons such as large forest gaps created because of round the year tourist activity and availability of resources due to heavy industrialization after its becoming capital of the state Uttarakhand. There is no second opinion that lantana is a threat to the nature, biomass estimation of lantana on the other hand can be very useful for the carbon assimilation process in the deciduous forests of the valley, also a shrub level carbon sequestration has never been calculated in Doon Valley, which can give more accurate estimation of forest biomass. Biomass is a key structural variable in invasive studies and researches into the dynamics of different ecosystems, the type of bio geographic conditions and biodiversity they sustain, their role in the atmospheric carbon cycle, and their resistibility in becoming an invasive species (Waring and Running 1996). Understanding the role of biomass in terms of nutrient cycles, fuel properties and their adaptation to different climatic conditions is also important (Terradas 1991; Rapp et al. 1999; Mary et al. 2001).
Calculation of above-ground biomass and other variables are required for better forest, scrubland, and grassland ecosystem management; development and execution of different models that help in the mapping of different characteristics of forest fuels (Nuñez-Regueira et al. 2000); and effects of invasive species on forest hydrological planning (Pastor-López and Martin 1995). Large-scale evaluation, mapping, and strategic values are necessary for accurate modelling of the dynamics of ecosystems (Oyonarte and Navarro 2003). There are a range of methods, from aerial photography and imagery to destructive sampling, for biomass estimation. Several general and local biomass tables and species-specific allometric equations have been developed for this reason; however, these are limited to tree species. Biomass analysis in forest ecosystems has been obtained in many studies through direct (destructive) or indirect (through dimensional analysis of different attributes of plants) sampling methods (Etienne 1989; Wharton and Griffith 1993).
Destructive methods involve harvesting the plants and weighing in the sample plots. This approach gives very accurate estimates of the biomass but harvesting and weighing a large number of samples is laborious and costly (Uresk et al. 1977). Indirect methods involve the use of allometric and mathematical models, which have been developed for estimation of total biomass and are based on the measurements of different plant attributes like shrub height, stem diameter at breast height (Smith and Brand 1983; Tietema 1993; Buech and Rugg 1989), crown diameter (Navarro and Oyonarte 2006), crown area and crown volume (Maraseni et al. 2005). These indirect methods produce estimates that can approach the prediction accuracy of the destructive methods and allow us to observe a large number of physical parameters at a relatively low cost (Castro et al. 1996; Uso et al. 1997; Montés et al. 2000). Shrub biomass is an important component of the total biomass, especially in the natural forests but, because of the lack of methodology and difficulty in calculation, they are omitted in some cases, which results in underestimation of the total biomass (Khanal 2001; Karki 2002). There are some limitations to the application of these equations. Equations developed for the same species in different sites would be different because of different climatic and edaphic conditions; e.g., the equations developed for Eucalyptus resinifera, at two different sites are different (Ward and Pickersgill 1985) and so are equations for Acacia aneura (Harrington 1979; Burrows et al. 2002). Also, in some cases, the range of applicable diameters or heights is not quantified (Specht and West 2003). In others, the range of diameters or heights is too narrow to apply even for the same species found in the same locality (Burrows et al. 2002; Margules Groome Pöyry Pty Ltd. 1998).
An accurate allometric equation for the biomass estimation of lantana, especially in a subtropical condition like the Doon Valley, is essential for two reasons. First, lantana is rapidly invading in areas such as roadsides, suburban areas, abandoned residential plots, pastures, even the cultivated woodlands. Second, measurement of shrub level carbon flux which is not only important in regional but also in the study of global CO2 cycle and therefore, estimation of shrub biomass can help to produce more accurate estimates of both forest biomass and biomass from other areas. Our primary objective was to compare the two previously used allometric equations for the estimation of shrub biomass proposed by Maraseni et al. (2005) and Navarro and Oyonarte (2006), and to see the compatibility of these allometric equations for lantana in a subtropical deciduous forest. We also wanted to develop a suitable allometric equation for the estimation of lantana biomass by using the different measured plant attributes from different study sites. The allometric regression equation developed for this study includes a variety of physical measurements of the different plant attributes, including circumference of the stem (which was later converted to diameter), the plant height, the maximum and minimum crown diameter (used to calculate the mean crown diameter), crown area, crown volume, phytovolume, and shrub coverage on the plot. Harniss and Murray (1976) used plant height and circumference to calculate shrub biomass while Dean et al. (1981) used maximum and minimum crown diameter, crown depth, and crown denseness.
Peek (1970) measured the maximum crown diameter, and the diameter at right angles to that, calculated canopy area for both an ellipse and a circle, and then calculated plant volume as area versus height. Bryant and Kothmann (1979) calculated crown volume as an inverted cone for four desert species. Murray and Jacobson (1982) developed many independent variables which include circumference, surface area, plant volume, diameter and height of the plant to predict leaf and twig biomass. Ferguson and Marsden (1977) were able to predict total twig length and weight for bitterbrush (Purshia tridentata) from the diameter at the base of the twig. Keeping the above facts in mind we decided to set our objective of the present study under the following domain; (1) To establish a dimensional relationship between different measured attributes of lantana such as D (basal diameter, cm), H (plant height, cm), C (crown diameter, cm), CA (canopy area, m2), CV (canopy volume, m3) and total biomass; (2) To establish an allometric equation for the calculation of lantana biomass from the dimensional relationship, as total harvesting of a large population of the shrub is impractical; (3) To compare the biomass of shrubs calculated by using the allometric equation and existing methods; and (4) To understand the present status of lantana in a native community through a vegetation analysis, which included the calculation of frequency, coverage, abundance and importance value index (IVI).
Materials and methods
Study area
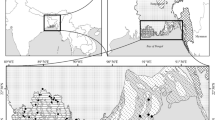
The present study was carried out in the Doon Valley, situated in western Himalayan part of India. The Doon Valley is surrounded by hills on all the sides and has a varied range of subtropical deciduous forests mainly dominated by Shorea robusta, Syzygium spp., Terminalia spp., Ehertia spp., Litsea spp. and others. It is lying between latitudes 29°55′ and 30°30′N and longitudes 77°35′ and 78°24′E. It is a saucer-shaped valley about 20 km wide and 80 km long, with a geographical area of about 2,100 km2 (Figs. 1 and 2). The valley is longitudinal, intermontane, synclinally depressed boulder (Thakur and Pandey 2004; Kumar et al. 2007) filled with coarse clastic fan-Doon gravel of the late Pleistocene and Holocene (Puri 1950). The valley is uniformly oriented in northwest-southeast direction, with lesser Himalayas in the northwest and the Siwalik ranges in the southwest. The two major hydrologic basins of the valley are the Ganga (or Ganges) in the southeast, with the Song and Suswa as its main tributaries, and the Yamuna in the northwest, with the river Asan as its main tributary. We selected eight sampling sites covering all parts of the Doon Valley: (1) Golatappar (open canopy areas), (2) Railway tracks (abandoned lands), (3) Asarori forest (dense forest areas dominated by Sal and Syzygium spp.), (4) Sahastradhara (a tourist place and highly disturbed areas), (5) Rajpur forest periphery (Sal-dominated forest), (6) Selaqui/Jhajra (abandoned residential plots and riverside forests dominated by Sal), (7) Jolly Grant airport (protected urban area), and (8) Mothronwala (swampy areas). The criteria for selecting these sites were to create a variation in physical factors, including different ranges of altitude, topography, habitat, and biotic interference level to calculate lantana infestation rate from the Doon Valley more effectively.

Pictorial representation of present study location in Indian western Himalaya

Potential distribution of Lantana camara in the invasive range of Doon valley as per Genetic algorithm for rule set prediction (GARP). (Source Google maps)
Estimation of shrub biomass
For destructive biomass, we determined biomass of lantana both destructively and non-destructively. For the destructive estimation of biomass, we followed the methods used by Navarro and Oyonarte (2006) in their study of calculating shrub biomass. We used transects in combination with the distance method in its closest individual mode, which permits the calculation of density, coverage, phytovolume, and phytomass in a relatively expeditious manner (Wharton and Griffith 1993; Navarro and Oyonarte 2006). A combination of the above mentioned variables is the best choice for obtaining an accurate description of a plant formation. The technique was as follows:
(1) Two transects were established from a randomly selected starting point in the chosen plot that was preferably located in the central region of each sampled zone. (2) Plot centres were located in the field using GPS. Transects were 50 m in length; the first transect was run in the direction of the slope gradient and the second transect perpendicular to the first at its mid-point. Fifty observations per plot were made, each 1 m apart, to encompass as high a variety of shrubs as possible. From each observation point, the distance to the nearest plant was measured, and the species of the plant recorded. (3) Four morphological variables [plant height (H, cm), largest crown diameter (C1, cm), smallest diameter (C2, cm) and diameter at bottom (C3, cm)] were measured for each specimen using a measuring tape (reading range 0–5 m, precision 1 cm, observation error, 0.5 cm). These data were used to calculate the mean crown diameter (CM, cm) and the phytovolume of each individual (Etienne 1989; Blanco and Navarro 2003). (4) After all the measurements were made in all plots, a sample of lantana was cut at ground level. The samples thus obtained were weighed in situ, without separating the stems and leaves, using an electric balance (Voda Scale, India, maximum weighing capacity 30 kg, minimum weighing capacity 1 kg, accuracy 2 g and a platter size of 400 × 240 mm stainless steel with auto calibration facility). (5) Whole individuals were transferred to the laboratory for drying in an air stove oven at 70 °C for 72 h until constant weight. The dried material was then weighed on the same balance to determine the dry weight of each fraction. (6) A set of formulas were then used to calculate the biomass of lantana using the oven dried mass already calculated by the methods described above. The detailed set of formulas is as follows:
The plant density per unit surface, D was calculated from
where, S is the estimated area (10,000 m2) in this study, d is the average distance between individual plants and points in a sampling, and δ is a correction factor depending upon the particular method used and equal to 2 in the closest individual method (Wharton and Griffith 1993).
The density of each species was calculated from
where, D (spi) is the density of individual species, D is the density per unit surface.
The mean diameter, CM, was used to obtain the coverage C of each plant associated with the ideal circle area:
where, C = Coverage of each plant, \(\pi = \text{3}\text{.142}\), \(\varPhi_{\text{M}}\) = mean crown diameter
The total coverage for species i was obtained from
The biomass per hectare for each species was calculated from the density values obtained in the samplings:
All the above mentioned equations were adopted from Navarro and Oyonarte (2006).
For non-destructive estimation of biomass
The dependant variable, shrub biomass, depends upon many independent variables. The identified independent variables to establish the allometric equation were total plant height (H), maximum basal diameter (D1) and minimum basal diameter (D2), maximum crown width (C1) and minimum crown width (C2), which were measured to calculate the average values before harvesting. Average values of crown width was calculated by C = (C1 + C2)/2 and used to calculate plant canopy area (CA, m2) and plant canopy volume (CV, m3) as (CA = πC2/4), and (CV = CA × H). However, average of D = (D1 + D2)/2 was used to establish the allometric equation along with the plant height.
Dry weight was already measured for each component by keeping it in oven at 70 °C for 72 h. The dry weight of lantana with its diameter (D) and D2H were tested in different regression models. We tested and applied allometric equations described in FAO (1999). The following equations were tested for its best fit for the calculation of shrub biomass.
\(y = a + bD^{\text{2}} H^{\ast}\) (equation used in the present study) \(ln y = a + bD^{\text{2}} H\)
In all of the above equations; y = the biomass, D = the diameter, H = the plant height, CA = canopy area, CV = canopy volume, a, b, c are regression coefficients, and ln indicates natural logarithm.
Comparison of models
The different models were compared to determine which one has better fit compared to others. R 2 (coefficient of determination) has long been used to compare models and hence chosen one of the criteria in this study. However, for models with different set of variables, R 2 gives misleading results (Parresol 1999). The use of root mean square error (RMSE) to compare models was also not found logical in the present case because some models considered were using transformed variables. We chose Furival Index as one of the main criteria along with R 2 to compare the models. Furnival’s Index was introduced by Furnival (1961) as a fit index that can be used for comparing models of different variables or weights based on the maximum likelihood concept. The advantage of this index is that it can reflect the size of the residuals and possible departures from linearity, normality and homoscedasticity (vector of random variables). Furnival Index is one of the recommended tools in statistics to compare different models (Parresol 1999; Montagu et al. 2005; Garber and Maguire 2005). The calculation is as follows:
where R 2 is the coefficient of determination obtained as the ratio of regression sum of squares to the total sum of squares, n is the number of observations on the dependant variables and p is the number of parameters in the model.
The Furnival Index was computed as follows:
-
(a)
The value of the square root of error mean square was obtained for each model under consideration through analysis of variance.
-
(b)
The geometric mean of the derivative of the dependant variable with respect to y was obtained for each model from the observations.
-
(c)
The geometric mean of a set of n observations was defined by the nth root of the product of the observations.
The Furnival Index for each model is then obtained by multiplying the corresponding values of the square root of mean square error with the inverse of the geometric mean.
For example, if the derivative of ln y is (1/y) and the Furnival Index in that case would be:
If the derivative of y is 1 then the Furnival Index would be:
The model y = a + bD 2 H was found having a lower Furnival Index compared to the other sample equations used in this study, and therefore given preference over other models (as model with lower Furnival Index and higher adjusted R 2 is considered more fit and accurate than the others).
Population Dynamics
Data on different parameters were collected from all sites. To study plant population dynamics, a quadrat of 5 m2 in size was laid at random after collecting the field data. Parameters like relative frequency, relative density, relative dominance, and the IVI of species were calculated using the formulae given below and used in previous studies (Oosting 1958; Phillips 1959; Hanson and Churchill 1961; Knox et al. 2011).
Carbon density was calculated by multiplying the biomass with the IPCC (Intergovernmental Panel on Climate Change 2006) default value for carbon conversion (0.47). Atmospheric carbon dioxide (CO2) removal was calculated by multiplying the carbon density or carbon fraction with 44/12 (the ratio of CO2:C).
We assigned the following numbers to the equations that were compared for accuracy, suitability, and compatibility with lantana for subtropical deciduous forests (Maraseni et al. 2005; Navarro and Oyonarte 2006).
where W = biomass of shrub, G10 = girth of plant at 10 cm, Cr V = crown volume
where, D(spi) = density of individual shrubs
where, y = biomass of shrub, D = diameter at breast height, H = plant height, a and b = coefficients
All the data were stored and processed for statistical analysis by using Microsoft Excel 2010 and SPSS (v 10).
Result
A comparison of the biomass calculated by the previously available methods and the equation developed for this study was made to check the compatibility of the equations under the present scenario. We chose an equation given by Maraseni et al. (2005) (Eq. 1), although it was not exclusively made for lantana in subtropical conditions, and then calculated the biomass of lantana by using both Eqs. 1 and 3 and compared the results. The estimated dry weights were closer to the predicted dry weight of lantana when we used Eq. 3 (Table 1). However, there was much more disparities in the estimated dry weight and the predicted dry weight of lantana when we used Eq. 1 and as a result did not match.
Since Eq. 1 did not work in our case, we had to find a suitable one that can be exclusively used for lantana in subtropical Himalaya. Our next step was to check the compatibility of already obtained oven dried mass of the whole plant with the independent variables such as D and H individually. A high adjusted R 2 values (R2 = 0.91) for D and (R2 = 0.86) for H (Figs. 3 and 4) were recorded. The individual correlation of these variables with oven dried mass was good but in search of a better one we then used the combined variables as D 2 H and tested its compatibility with the dry mass, a comparatively better value of adjusted R2 was obtained (R2 = 0.88). Based on R 2, it was found that the best independent variable to use in the regression model was D 2 H. This was not the only criteria used for choosing the best fitted allometric equation. When all the equations were tested for their best suitability, we followed more criteria to choose the best one i.e., the adjusted R 2, the Furnival Index, computed F value, RMSE, and the significance level of the values at 5 %. The regression model for the biomass calculation is considered more accurate and best fitted if it gives a lower RMSE, higher value of adjusted R 2, and less value of Furnival Index. Out of all the variables we used, D 2 H was found to be the best fitted variable considering all these above criteria with adjusted R2 = 0.88 the maximum, Furnival Index (0.04123), and RMSE (157) lowest amongst all the equations (Table 2c).

Scatter plot to show the correlation between shrub height and calculated biomass (Eq. 3)

Scatter plot to show correlation between stem diameter and calculated biomass (Eq. 3)
However, when the computed value of F was compared with the tabular value at (1, n−2) degrees of freedom, we found that the calculated value of F (137.59) were greater than the tabular value (4.41) at (1, 18) degrees of freedom at 5 % significance level (Table 2a). This result shows that the regression coefficient, β is significantly different from 0 and the value is statistically significant. Some independent variables had considerably higher influencing power over the dependent variables in one combination of independent variables (high t values and low significance level), but in other combinations, the influencing power was very poor (low t values but high significance level). Therefore, considering all the facts we found that the regression model with D 2 H as a variable was most suitable for the estimation of shrub biomass and hence chosen for this study (Fig. 5).

Scatter plot to show correlation between dried weight and D 2 H
The findings of this study (using Eq. 3) show that Rajpur forest periphery has the greatest per-plant biomass (0.764 kg/plant), followed by Mothronwala (0.637 kg/plant), which is slightly ahead of Sahastradhara (0.622 kg/plant). The per-hectare biomass of shrub was found highest in Sahastradhara (13,559.60 kg ha−1), followed by Mothronwala (12,956.58 kg ha−1) and Rajpur forest periphery (11,154.40 kg ha−1) (Fig. 6). Sahastradhara has the highest density of the shrub than the other sampling sites (21,800 plants·ha−1) because the per-hectare biomass was highest here. These values were closer to the biomass values obtained by Eq. 2 with a marginal difference, but the biomass values obtained by Eq. 1 were found significantly different from the other two values (Table 3).

Comparison of lantana biomass with all the three equations considered in this study
Carbon density was highest in Sahastradhara with (0.637 kg m−2) and (6,373.01 kg ha−1); followed by Mothronwala (0.609 kg m−2) and (6,090 kg ha−1); and Rajpur forest (0.524 kg m−2) and (5,242.56 kg ha−1). The least was recorded from Jolly Grant (0.317 kg m−2) (3,170.99 kg ha−1). The above carbon density shows that lantana’s CO2 removal capacity was greatest in Sahastradhara at the rate of 23,367.71 kg ha−1, followed by in Mothronwala at the rate of (22,328.50 kg ha−1), and in Rajpur forest (19,222.74 kg ha−1). The lowest CO2 removal site was found to be the Jolly Grant airport with a rate of 11,626.98 kg ha−1 (Table 4).
From the above results, it was clear that Sahastradhara has the maximum density of lantana and the maximum CO2 removal site amongst the entire study sites. Sahastradhara was the most disturbed site because of round the year tourist activities. High anthropogenic activity and abundant resource availability with less competition level probably favoured the growth of lantana at a tremendous rate in this site (Table 4). Incidentally, the shrub canopy area from Sahastradhara was also the highest (2.68 m−2); followed by Rajpur forest periphery (2.64 m−2); and Mothronwala (2.45 m−2).
The coverage of lantana was calculated from all sites and was found highest in Sahastradhara (5.86 m−2); followed by Mothronwala (5.00 m−2); and in Golatappar (3.96 m−2). The smallest canopy area was calculated from Jolly Grant airport (1.67 m−2); and railway tracks (1.66 m−2) (Fig. 7). Some findings of this study were found surprising because the railway tracks and most of the areas near Jolly Grant airport are fallow abandoned lands with more available resources and light availability, but these sites still showed less canopy area and phytovolume than other areas. However, the density of lantana in these areas was found satisfactory with 18,100 plants ha−1 and 20,200 plants ha−1 respectively. Phytovolume of lantana from all of the sites was also calculated and was found highest in Rajpur forest periphery (201,363.20 m−3), followed by Sahastradhara (155,584.42 m−3), and Mothronwala (144,932.67 m−3).

Site wise comparison of canopy coverage (% ha−1) and canopy volume (m3) of lantana in Doon Valley
Despite the fact that the density of lantana (21,800 plants ha−1) was found maximum in Sahastradhara, the phytovolume was highest in Rajpur forest because the average shrub height (5.08 m) and average crown diameter (180.28 cm) per shrub was highest in Rajpur forest peripheries, which were found essential parameters for calculating phytovolume of lantana (Table 5). During the plant community analysis for the assessment of current status of lantana under the present climatic conditions, we found that lantana was dominant in all the study sites with maximum coverage and IVI in its class and was not favouring the growth of all the species in its vicinity. Only a few species were co-dominantly growing with lantana including Parthenium hysterophorus L., Ageratum conyzoides L., Chromolaena odorata (L.) King and Robinson, Callicarpa macrophylla Vahl., Opuntia dillenii (Ker Gawl.) Haw., Adhatoda zeylanica L., Murraya koenigii (L.) Spreng., Cassia occidentalis L., Arundinella spicata Dalzell., Floscopa scandens Lour., Achyranthes aspera L., and Reinwardtia indica Dum. The maximum IVI of lantana (27.43) was calculated at Jolly Grant airport, which was followed by Sahastradhara (22.77); Rajpur forest areas (21.93); Selaqui/Jhajra forest areas (18.72); and Asarori forest areas (17.51). However, the IVI of Parthenium was dominant in the areas like Golatappar (18.94); railway tracks (24.12); and Mothronwala swamp (19.14). The frequency of lantana in all the study areas was 100 %. Table 6 shows a detailed account of five co-dominant species that were found growing along with lantana frequently without any problem, with 100 % frequency from all the sites.
Discussion
Estimation of shrub biomass is very important as it can provide more accurate estimates of forest biomass and carbon sequestration. A rapid, nondestructive method is needed to make biomass estimates because the labour and expense necessary to clip and weigh large plants are more and impractical to preserve the ecosystem (Ludwig et al. 1975). Methods to establish a relationship between easily obtained plant measurements and plant biomass include a technique termed dimension analysis (Whittaker 1970) and further its utilization in shrub biomass estimation through regression analysis (Rutherford 1979). In a study by Rana et al. (1989), the herbs and shrubs of each quadrat were harvested and the litter from each quadrat was placed in paper bags and brought to the laboratory. The samples were oven dried at 80 °C to constant weight and then the net primary production for the tree, shrub and herb layers was calculated from each site. In the present study we give more emphasis in using the allometric equation for non-destructive biomass calculation of lantana over the destructive calculation as it is time saving and easy to use, this was advocated by some previous study where power model and quadratic models were used and found to be the most appropriate equations to describe size biomass relationships (Zeng et al. 2010). Some workers gave more emphasis in using a quadratic equation as a common model to estimate shrub biomass (Sah et al. 2004; Foroughbakhch et al. 2005), and the present results are in line with previous reports that the biomass of shrub populations can be well described with measured architectural variables such as diameter at breast height (DBH) and plant height (H) (Foroughbakhch et al. 2005).
In some other studies, the carbon stock and carbon sequestration rates were estimated as 50 % of the dry weight of biomass and 50 % of net primary productivity, respectively (Hamberg 2000; Brown 2001). The fuel value of a biomass is solely dependent on its calorific value. The amount of heat generated by any fuel depends on the quantitative conversion of carbon and hydrogen present in the fuel to water and carbon dioxide, and is a function of chemical composition of the fuel (Kumar et al. 2009). In our study, the maximum above-ground biomass of lantana amongst all the sites of Doon Valley is 0.764 kg/plant and was recorded from the Rajpur forest 11,154.40 kg ha−1. This is a great source of fuel in an ecosystem characterized as deciduous because its invasion rates are extremely high in all kinds of geographical conditions except the native areas where it originally grows.
However, in some studies the biomass is also calculated and its importance is recognized by using the wood density and logarithmic regression (Barasa et al. 2010; Chaturvedi et al. 2011). The previous method is accurate but it is laborious and time consuming: in a place like the Doon Valley, where the infestation of lantana is very high, it is difficult to calculate the biomass by harvesting methods and oven-dried weight for all the areas. The model developed and used in this study makes it possible to calculate the biomass of lantana in the subtropical deciduous conditions of the Doon Valley by nondestructive means. The accuracy is high and when compared and tested with the oven-dried mass of lantana gave a very positive result. The method will surely save time and labour when applied on a large scale.
The present study shows that the biomass of lantana is best predicted by one (basal diameter), (plant height) or two (the basal diameter and total plant height) variables. In some recent studies, allometric equations are used for environmental studies (Satoo and Madgewick 1982; Araújo et al. 1999; Montés et al. 2000). The findings of this study also suggest that allometric equations are the most trustworthy method for estimating the biomass of shrubs which is consistent with the previous studies (Foroughbakhch et al. 2005; Maraseni et al. 2005; Zeng et al. 2010).
Out of all the study sites, Sahastradhara, Rajpur forest periphery, Jolly Grant airport and its adjacent areas were found very disturbed due to tourist activities; construction works and heavy grazing, thus the lantana density, frequency and IVI from these areas were found elevated. In contrast, places like Golatappar, Railway tracks, and Mothronwala were less disturbed and therefore IVI of lantana was recorded less from these sites. Our results on the basis of high IVI from Jolly Grant airport and Rajpur forest periphery clearly show that lantana is found not only in the fallow abandoned land and in the human disturbed areas but has also started invading in a very strong way to the periphery of the forests and the protected areas.
Though the present study is mainly focused on the calculation of above-ground lantana biomass from the subtropical conditions of the Doon Valley, a lesser attempt was made to understand the invasive nature of lantana in these conditions on the basis of site characteristics, IVI, coverage of plants, and other community analyses. But these findings are not sufficient as the suitability of lantana depends not only on physical disturbance of the site but also on other factors like topography, altitude, soil profile, role of herbivory, and plant attributes.
One interesting finding was lantana’s tremendous power of competition with native trees; e.g., shrub height was greater in the forest periphery (Rajpur forest periphery) and adjacent places than that of the fallow and abandoned land. This observation also shows that lantana, due to its aggressive nature and competitive ability, can very well increase its height and further growth to obtain the resources.
In recent times, the biomass and carbon budget from all types of forests have gained considerable attention of researchers across the globe. However, the role of invasive shrubs in the carbon budget has been elusive due to the lack of proper methodology and resources. Taking a broader look, India is the third largest emitter of CO2 in the world, with emissions of 1,427.6 million tones of CO2 per year (India Climate Portal 2010); China and USA are on the first and second positions, respectively. According to the data, percent change in CO2 emissions from 1990 to 2008, emissions have shot up 146 % in India, second only to China. In contrast, the United States and Japan have only increased in proportion to 14.9 and 8.2 % respectively, which is still manageable. These results show that the conditions in India are very alarming and these forests and shrub-lands can be very useful to sequester CO2.
Lantana invasion in different areas is a much more recent phenomenon and is reducing species richness of native understory species while also causing compositional changes in the herbs and shrubs, as well as tree seedlings (Sharma and Raghubanshi 2010; Sharma and Raghubanshi 2011). Nevertheless its enormous potential to be an abundant fuel source and sequester CO2 from the atmosphere should not be overlooked.
There are many alien and invasive species in the Doon Valley forests (see Table 6), growing with lantana. Together these alien influences could be gradually changing these tropical and subtropical forests, causing significant structural and compositional changes in the vegetation which is likely to have cascading effects on consumer communities, in addition to altering the dynamics and function of this endangered ecosystem. According to 2006 data from the Wildlife Institute of India, India has 778,229 sq. km forest area, which is 23.68 % of the total area of the country (Wildlife Institute of India 2006). Of that total forest area, 20.04 % comes under the protected areas network, of which 24.37 % are national parks and 75.59 % are wildlife sanctuaries. These protected areas are prime wildlife habitats and harbor diverse forest communities. A number of trees and shrubs in these forests are invasive and threatening the native species. The biomass value of these invasive species will not only divulge the accurate carbon sequestration potential of these forests but also reveal their positive aspects for which they are not very well known. The biomass values of forest stands in the Himalayan forests tend to cluster around two very different levels, from a low of approximately 200 t ha−1 for early successional communities such as pine, to a high of about 400 t ha−1 for late successional communities such as the oaks and sal (Singh et al. 1992).
Carbon storage in the Uttarakhand Himalayan forests (the present study sites) ranges from an average of about 175 t C ha−1 for pine forests to approx 300 t C ha−1 for oak and sal dominated forests (Singh et al. 1985). Our findings clearly show how important shrubs are in these areas because they greatly contribute to carbon storage (Table 4). The eight study sites of the present study are storing 75,442.28 kg ha−1 carbon, which were found sequestered by only invasive shrub lantana. It is therefore very important to calculate the biomass and to find out the fuel properties of lantana so that it can be used in the restoration of ecosystems. From the previous literature, it has been clear that the biomass of this shrub in most of cases was calculated or measured by the harvesting methods (Sharma and Raghubanshi 2011) but seldom by using allometric regression models where the measured attributes of plants are considered.
The surroundings of Doon Valley include Rajaji National Park, (which is famous for its mammalian and bird species). Conservation of this endangered forest and its wildlife perhaps hinges on addressing the role of these pervasive factors in driving ecological change. Forests, particularly tropical forests, contribute more than other terrestrial biomes to climate- relevant cycles and processes. This particular forest is a great habitat of the natural sal trees, though it is a protected forest but human activities are still in progress. Heavy human activities in recent years resulted in large gaps in these forests further providing corridors to the success invasive understory species. It would be possible to increase the carbon storage capacity of these forests if we positively consider sal trees and the understory invasive vegetation as a good carbon sink. Proper forest management is required which include adoption of the methods for afforestation, reduced deforestation techniques and invasive species management (Shukla 2006; Sharma et al. 2006). In India, very little information exists about the rate at which different forest ecosystems sequester carbon. Although details on annual net carbon uptake of biomass are available for a few specific ecosystems, they are usually carried out for small plots and do not exist for all the forest types in India.
Therefore, the need to accurately estimate carbon in shrubs will become even more vital in the future as land use and global climate changes increasingly alter both forested and non-forested ecosystems. Such changes will have implications for wildlife, biodiversity, nutrient cycling, fire, and other management issues where carbon sequestration, especially in soils, may become key objectives. We have summarized three methods that might be used to estimate carbon in shrubs but the compatibility and fitness of the equation depends on type of shrubs, growth patterns, physiographic conditions, soil types and disturbance level. The job now is to develop appropriate equations and workable field measurement techniques that together will enable researchers and managers to measure carbon in shrubs quickly, consistently, and rigorously in the future.
Conclusion and recommendation
Our findings show that because of lantana’s good fuel properties and ability to store carbon, it does have some role to play in the ecosystem restoration. There is a clear need to calculate the biomass of lantana and it is possible to estimate biomass nondestructively. The different variables used in this study reveal its close relationships between total biomass and diameter, height, and, up to some extent the canopy area, canopy volume and their combinations. These combinations enabled us to develop and use allometric equations to estimate total biomass of lantana in subtropical Himalaya. This equation was developed and used exclusively for lantana found in a similar climatic region of the study area but because of its effectiveness, we strongly recommend the equation (y = a + b D2H) for the nondestructive estimation of lantana.
References
Araújo TM, Higuchi N, Carvalho JA Jr (1999) Comparison of formulae for biomass content determination in a tropical rain forest site in the state of Para, Brazil. For Ecol Manag 117:43–52
Barasa B, Majaliwa MGJ, Lwasa S, Obando J, Bamutaze Y (2010) Estimation of the aboveground biomass in the Trans-Boundary River, Sio Sub-catchment in Uganda. J Appl Sci Environ Manag 14(2):87–90
Blanco P, Navarro RM (2003) Aboveground phytomass models for major species in shrub ecosystems of Western Andalucia. Investigaciones Agrarias: Sistemasy Recursos Forestales 12(3):47–55
Brown S (2001) Measuring and monitoring carbon benefits for forest-based projects: experience from pilot projects, can carbon sinks be operational? Resources for the future (RFF) workshop proceedings, Washington DC, p 119
Bryant FC, Kothmann MM (1979) Variability in predicting edible browse from crown volume. J Range Manag 32(2):144–146
Buech RR, Rugg DJ (1989) Biomass relation of shrub components and their generality. For Ecol Manag 26:257–264
Burrows WH, Henry BK, Back PV, Hoffmann MB, Tait IJ, Anderson ER, Menke N, Danaher T, Carter JO, McKeon GM (2002) Growth and carbon stock change in Eucalypt woodlands in Northeast Australia: ecological and greenhouse sink implication. Glob Change Biol 8:769–784
Castro I, Casado MA, Ramarez Sanz L, De Miguel JM, Costa M, Diaz F (1996) Funciones de estimacion de la fitomasa aérea de varias species del matorral mediterráneo del centro de la peninsula ibérica. Orsis 11:107–116
Chaturvedi RK, Raghubanshi AS, Singh JS (2011) Carbon density and accumulation in woody species of tropical dry forest in India. For Ecol Manag 262:1576–1588
Dean S, Burkhardt JW, Meeuwig RO (1981) Estimating twig and foliage production of sagebrush, bitterbrush and rabbitbrush in the Great Basin. J Range Manag 34(3):224–227
Di Castri F, Goodall DW, Specht RL (1981) Mediterranean type shrubland. Elsevier Scientific Publishing Company, Amsterdam, p 56
Eiswerth ME, Darden TD, Johnson WS, Agapoff J, Thomas RH (2005) Input-output modeling, outdoor re-creation, and the economic impacts of weeds. Weed Sci 53(1):130–137
Etienne M (1989) Non-destructive methods for evaluating shrub biomass: a review. Acta Oecol 10:115–128
FAO (1999) Forestry research support programme for Asia and the Pacific: a statistical manual for forestry research. Food and Agriculture Organization of the United Nations Regional Office for Asia and the Pacific Bangkok, pp 181–191
Ferguson RB, Marsden MA (1977) Estimating overwinter bitterbrush utilization from twig diameter-length-weight relations. J Range Manag 30(3):231–236
Foroughbakhch R, Reyes G, Alvarado-Vázquez MA, Hernández-Piñero J, Rocha-Estrada A (2005) Use of quantitative methods to determine leaf biomass on 15 woody shrub species in north-eastern Mexico. For Ecol Manage 216:359–366
Furnival GM (1961) An index for comparing equations used in constructing volume tables. For Sci 7:337–341
Garber SM, Maguire DA (2005) The response of vertical foliage distribution to spacing and species composition in mixed conifer stands in central Oregon. For Ecol Manag 211(3):341–355
Hamburg SP (2000) Simple rules for measuring changes in ecosystem carbon in forestry-offset projects. Mitig Adapt Strat Glob Change 5:25–37
Hanson HC, Churchill ED (1961) The plant community. Reinhold Publishing Corporation, New York, p 155
Harniss RO, Murray RB (1976) Reducing bias in dry leaf weight estimates of big sagebrush. J Range Manag 29(5):430–432
Harrington G (1979) Estimation of above-ground biomass of trees and shrubs in a Eucalyptus populnea F. Mull. woodland by regression of mass on trunk diameter and plant height. Aust J Bot 27:135–143
India Climate Portal (2010) SAARC summit: promise of possibility?, vol 13. India Climate Watch, India
IPCC (2006) Agriculture, forestry and other land use. In: Eggleston HS, Buendia L, Miwa K, Ngara T, Tanabe K (eds) IPCC guidelines for national greenhouse gas inventories, prepared by the national greenhouse gas inventories programme. IGES, Japan, p 120
Karki D (2002) Economic assessment of community forestry in inner terai of Nepal: a case study from Chitwan district. Asian Institute of Technology press, Thailand, p 35
Khanal KP (2001) Economic evaluation of community forestry in Nepal and its equity distribution effect. The Royal Veterinary and Agricultural University Press, Denmark, p 68
Knox J, Jaggi D, Paul MS (2011) Population dynamics of Parthenium hysterophorus (Asteraceae) and its biological suppression through Cassia occidentalis (Caesalpiniaceae). Turkish Journal of Botany 35:111–119
Kumar R, Suresh N, Sangode SJ, Kumaravel V (2007) Evolution of the quaternary alluvial fan system in the Himalayan Foreland Basin: implications for tectonic and climatic decoupling. Quatern Int 159:6–20
Kumar R, Chandrashekar N, Pandey KK (2009) Fuel properties and combustion characteristics of Lantana camara and Eupatorium spp. Curr Sci 97(6):930–935
Ludwig LJ, Charles-Edwards DA, Withers AC (1975) Tomato leaf photosynthesis and respiration in various light and carbon dioxide environments. In: Marcell R (ed) Environmental and biological control of photosynthesis. Dr. W. Junk b. v., Publishers, The Hague, pp 17–28
Maraseni TN, Cockfield G, Apan A, Mathers N (2005) Estimation of shrub biomass: development and evaluation of allometric models leading to innovative teaching methods. Int J Bus Manag Edu (Special Issue):17–32. http://core.kmi.open.ac.uk/download/pdf/11036840.pdf
Margules Groome Pöyry Pty Ltd (1998) Wood fibre yields for South East Queensland RFA region (Draft Report): prepared for CRA unit. Department of Natural Resources, Brisbane and BRS, DPIE, Canberra
Mary AA, Steven PH, Siccama T (2001) Validating allometric estimates of aboveground living biomass and nutrient contents of a Northern hardwood forest. Can J For Res 31:11–17
Montagu KD, Duttmer K, Barton CVM, Cowie AL (2005) Developing general allometric relationship for regional estimates of carbon sequestration—an example using Eucalyptus pilularis from seven contrasting sites. For Ecol Manag 204(1):115–129
Montés N, Gauquelin T, Badri W, Bertaudière V, Zaoui EH (2000) A non-destructive method for estimating above-ground forest biomass in threatened woodlands. For Ecol Manag 130:37–46
Murray RB, Jacobson MQ (1982) An evaluation of dimension analysis for predicting shrub biomass. J Range Manag 35(4):451–454
Navarro RM, Oyonarte PB (2006) Estimation of above ground biomass in shrubland ecosystems of Southern Spain. Investigación Agraria. Sistemasy Recursos Forestales 15(2):197–207
Nuñez-Regueira L, Proupin-Castineiras J, Rodriguez-Anon JA (2000) Design of risk index maps as a tool to prevent forest fires in the hill side zone of Galicia (NW Spain). Bioresour Technol 73(2):123–131
Oosting HJ (1958) The study of plant communities. Freeman WH and Corporation, San Francisco
Oyonarte PB, Navarro RM (2003) Aboveground phytomass models for major species in shrub ecosystems of western Andalusia. Investigación Agraria. Sistemas y Recursos Forestales 12(3):47–55
Parresol R (1999) Assessing tree and stand biomass: a review with examples and critical comparisons. For Sci 45:573–593
Pastor-López A, Martin J (1995) Ecuaciones de biomasa para Pinus halepensis en repoblaciones de la provincial de Alicante. Studia Oecologica 12:79–88
Peek JM (1970) Relation of canopy area and volume to production of three woody species. Ecology 51(6):1098–1101
Phillips EA (1959) Methods of vegetation study. Holt H & Corporation, New York, p 96
Puri GS (1950) Soil pH and forest communities in the Sal (Shorea robusta) forests of the Dehradun Valley, U.P. India. Indian For 76(7):292–303
Rana BS, Singh SP, Singh RP (1989) Carbon and energy dynamic of seven Central Himalayan forests. Trop Ecol 30(2):253–264
Rapp M, Regina IS, Gallego HA (1999) Biomass nutrient content, litter fall and nutrient return to the soil in Mediterranean oak forest. For Ecol Manag 119:39–49
Ruiz de la Torre J (1990) Mapa Forestal de Espana. Memoria general. ICONA. Ministerio de Agricultura, Pescay Alimentacion, Madrid, p 191
Rutherford MC (1979) Plant-based techniques for determining available browse and browse utilization: a review. Bot Rev 45:203–228
Sah JP, Ross MS, Koptur S, Snyder JR (2004) Estimating aboveground biomass of broadleaved woody plants in the understory of Florida Keys pine forests. For Ecol Manag 203:319–329
Satoo T, Madgewick HAI (1982) Forest biomass. Junk, Boston, The Hague, p 152
Sharma GP, Raghubanshi AS (2010) How Lantana invades dry deciduous forest: a case study from Vindhyan highlands, India. Trop Ecol 51(2S):305–316
Sharma GP, Raghubanshi AS (2011) Lantana camara L. invasion and impact on herb layer diversity and soil properties in a dry deciduous forest of India. Appl Ecol Environ Res 9(3):253–264
Sharma S, Bhattacharya S, Garg A (2006) Greenhouse gas emissions from India: a perspective. Curr Sci 90(3):326–333
Shukla PR (2006) India’s GHG emission scenarios: aligning development and stabilization paths. Curr Sci 90(3):384–395
Singh JS, Tewari AK, Saxena AK (1985) Himalayan forests: a net source of carbon to the atmosphere. Environ Conserv 12:67–69
Singh SP, Negi GCS, Pant MC, Singh JS (1992) Economic considerations in Himalayan agroecosystems. In: Agrawal A (ed) The Price of Forests. Centre for Science and Environment, New Delhi, p 296
Smith WB, Brand GJ (1983) Allometric biomass equations for 96 species of herb, shrub and small trees. USDA Forest Service, North Central Forest Experimental Station, Saint Paul, p 299
Specht A, West PW (2003) Estimation of biomass and sequestered carbon on farm forest plantation in Northern New South Wales, Australia. Biomass Bioenergy 25:363–379
Terradas J (1991) Mediterranean woody plant growth forms, biomass and production in the eastern part of the Iberian Peninsula. Oecologia 10:337–349
Thakur VC, Pandey AK (2004) Late quaternary tectonic evolution of Doon in fault bend/propagated fold system. Garhwal Sub Himalaya. Curr Sci 87(11):1567–1576
Tietema T (1993) Biomass determination of fuel wood trees and bushes of Botswana, Southern Africa. For Ecol Manag 60:157–269
Uresk DW, Gilbert RO, Rickard WH (1977) Sampling big sagebrush for phytomass. J Range Manag 30:311–314
Uso JL, Mateu J, Karjalainen T, Salvador P (1997) Allometric regression equations to determine above ground biomass of Mediterranean shrubs. Plant Ecol 132:59–69
Ward SC, Pickersgill GE (1985) Nutrient distribution in two eucalypt plantations growing on rehabilitated bauxite mines. Aust J Ecol 10:111–124. doi:10.1111/j.1442-9993.1985.tb00873.x
Waring RH, Running SW (1996) Forest ecosystems Analysis at multiple scales. Academic Press, Harcourt Brace and Company, New York, p 370
Wharton EH, Griffith DM (1993) Methods to estimate total forest biomass for extensive inventories: applications in the north-eastern U.S. USDA Forest service Research Paper, USA, p 681
Whittaker RH (1970) Communities and ecosystems. Macmillan, New York, p 162
Wildlife Institute of India (2006) Summary of existing protected area network in India. Wildlife Institute of India, National Wildlife Database Cell, New Delhi
Zeng HQ, Liu QJ, Feng ZW, Ma ZQ (2010) Biomass equations for four shrub species in subtropical China. J For Res 15:83–90
Acknowledgments
We thank Ecology Research Lab and Botany Department of DAV (PG) College, Dehradun, India for providing necessary help in collecting samples and analysis. We give our sincere regards to the staff of the Department of Botany and National Forest Library, ICFRE, Dehradun, India for providing necessary laboratory and literary help during this work. We also thank Bharti Vidyapeeth University, Pune, India for providing statistical support during this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
The online version is available at http://www.springerlink.bibliotecabuap.elogim.com
Corresponding editor: Hu Yanbo
Rights and permissions
About this article

Cite this article
Mandal, G., Joshi, S.P. Estimation of above-ground biomass and carbon stock of an invasive woody shrub in the subtropical deciduous forests of Doon Valley, western Himalaya, India. J. For. Res. 26, 291–305 (2015). https://doi.org/10.1007/s11676-015-0038-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-015-0038-8