
Abstract
Purpose
Chinese fir (Cunninghamia lanceolata (Lamb.) Hook) is an important native tree species in China. Consecutive cropping traditionally occurs in Chinese fir plantations (CFPs), but this practice has resulted in productivity declines in subsequent rotations. This study was designed to better understand the change of soil properties in the continuous cropping CFPs.
Materials and methods
We investigated soil pH, soil organic matter (SOM), and nutrient contents in different soil layers and in rhizosphere soil (RS) and non-rhizosphere soil (NRS) under CFPs of different ages and in different rotations.
Results and discussion
In the upper (0–20 cm) soil layer, soil pH decreased, while SOM increased, beneath mature CFPs with consecutive rotations. Total nitrogen (TN), available potassium, and available phosphorus contents in the upper soil layers did not differ significantly with consecutive rotations. Soil pH in RS was significantly lower than in NRS under mature plantations of the third rotation. Soil organic matter, TN, and available nitrogen did not differ between RS and NRS. Available phosphorus in RS was consistently lower than in NRS, and was highly deficient in the third rotation.
Conclusions
We conclude that no severe soil nutrient degradation occurred in the continuous cropping CFPs examined in this study, with soil acidification and phosphorus deficiency being two primary problems observed.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Chinese fir (Cunninghamia lanceolata (Lamb.) Hook) is an indigenous tree species in South China. Due to desirable features such as fast growth, pest and disease-resistance, and good timber quality, it is historically one of the most important timber tree species in South China and has been widely planted for decades in hilly and mountainous tropics and subtropics of China. Many natural forests have been converted into monocultural Chinese fir plantations (CFPs) in South China since the middle of the twentieth century (Xu et al. 2005). Plantations of this species cover more than 1.10 × 107 hm−2, accounting for over 6.66 % of the total forest area in China (State Forestry Administration of the People’s Republic of China 2014).
Consecutive rotation of Chinese fir is a traditional silvicultural practice and has been widely practiced in South China (Yang et al. 2005). A new rotation is usually established after clear-cutting, slash burning, and tillage prior to planting. In recent decades, researchers have found that CFPs in the continuous rotation system exhibited reduced growth rates and yields (Fang et al. 2010). For example, the heights of dominant Chinese fir trees of the second and third rotations decreased by 7 and 23 % relative to the first rotation, respectively (Bi et al. 2007). Additionally, the standing wood volumes of the second and third rotations of CFPs were 30 and 47 % lower than the first rotation, respectively (Bi et al. 2007). Plantation biomass growth rates decreased by 24 % from the first to the second rotation and by 40 % from the second to the third rotation (Zhang et al. 2004). The decline of productivity in continuously cropped Chinese fir ecosystems remains an outstanding problem in plantation management in South China (Yang et al. 2005). Degradation of successively planted forestland of other tree species has caused concerns since the nineteenth century, and extensive studies on mechanisms and countermeasures have been conducted in other parts of the world (Raynaud 2010). Results, however, are by no means consistent. The total volume yield in a second-generation Norway spruce (Picea abies (L.) Karst.) stand was 40 % higher than that of the first generation and dominant tree height at a stand age of 40 years was about 20 % greater in the second generation than in the first (Eriksson and Johansson 1993).
Since the 1980s, continuous cropping of Chinese fir has been reported to cause soil degradation with regard to physiochemical, biological, and biochemical properties (Chen and Wang 2003; Sun et al. 2003; Bi et al. 2007; Zhao et al. 2013). For example, under continuous rotations, water holding capacity and permeability of the surface soil declined and bulk density increased (Yao and Xie 1991); additionally, soil pH decreased and available P and N declined sharply. Soil organic matter (SOM) contents in the second rotation decreased by 10 % relative to the first rotation, and further decreased by 15 % in the third rotation relative to the second rotation (Huang et al. 2004). Soil enzyme activity in the second rotation was significantly lower than that of the first (Chen and Wang 2003), and microbial community diversity, as well as microbial biomass, decreased with successive rotations (Liu et al. 2010). The above aspects of soil degradation were believed to result from successive cropping, short rotations (around 25 years), whole-tree harvest, and tillage (Zhang et al. 2004; Liu et al. 2010). For example, Yang et al. (2005) reported that N removal via clear-cutting from CFPs was 233 kg hm−2 and, compared to pre-burning, topsoil organic C and total nitrogen (TN) contents were reduced by 17 and 19 %, respectively, just two days after slash burning. Also, timber harvest caused substantial loss and redistribution of SOM and N (Wear and Greis 2002), and exerted negative effects the ecosystem C balance (Chen et al. 2013).
Some scientists, however, have reported that soil degradation does not necessarily develop consistently with continuous cropping. Wei et al. (2012) showed that soils under CFPs of the third rotation had higher SOM, total P and K, and available nitrogen contents than those of the second rotation, hypothesizing that sustainable management of CFPs could be attained in the continuous cropping system if plantation density was optimized. Also, soil fertility changed but did not necessarily decrease with plantation development. Sun et al. (2003) observed that comprehensive soil fertility improved under mature plantations relative to the half-mature plantations in the first rotation. According to Wang et al. (2010), contents of SOM, TN, available N, and available K decreased in half-mature plantations, but significantly increased in mature plantations and reached maximum levels in overly mature plantations. Ma et al. (2007) also noticed that SOM contents increased with CFPs growth. Wu et al. (2011) compared soil fertility under CFPs of different ages and found that fertility was highest in overly mature forests and decreased through mature forests, near-mature forests, and young forests to its lowest level in half-mature forests. Zhou et al. (2016) also found that young stands of CFPs depleted soil nutrients and showed low nutrient return, while mature or over-mature stands had greater nutrient return leading to better recovered nutrient pools in forest soil.
Consequently, it remains uncertain whether soil degradation is a universal problem for CFPs managed under continuous rotations, and the manner in which pH, SOM, and nutrient contents change in response to stand age and rotation in CFPs. Additionally, the severity of soil degradation associated with continuous rotation requires more careful and detailed examination. For this purpose, we investigated soil pH, SOM, and nutrient contents under different-aged CFPs in the first, second, and third rotations and compared soil pH, SOM content, and macronutrient availability in the rhizosphere soil (RS) with those of the non-rhizosphere soil (NRS).
2 Materials and methods
2.1 Study area and site description
The study area was located in Xidong Forest Farm, Jian’ou City (27° 03′ N, 118° 20′ E), Fujian Province, one of the central production regions of CFPs in China. According to the space-for-time ecological sequential comparison method (Fukami and Wardle 2005), CFPs in the first rotation, second rotation, and third rotation (1, 2, and 3, respectively) that were either young (y; 5–6 years old), half-mature (h; 12–15 years old), and mature (m; 25–30 years old) were carefully selected for the study, with each rotation and age combination (1-y, 1-h, 1-m, 2-y, 2-h, 2-m, 3-y, 3-h, and 3-m) being represented by a typical plantation. The selected plantations were in close proximity, shared the same soil type, occurred at similar elevation (530–560 m) and exhibited comparable micro-topography (Table 1). The soil was developed from granite parent material and classified as dark red soil in the Chinese soil classification system. Soil depths under the selected plantations were mostly greater than 1 m, with slopes between 20 and 30 °. The study area belonged to the middle subtropical monsoon climatic zone, with a mean annual temperature of 18.7 °C and a mean annual precipitation of 1670 mm, which mainly occurred from March to August. The topsoil bulk density was 1.20 ± 0.03 g cm−3, capillary porosity was 44.8 ± 1.3 %, and field capacity was 421.0 ± 27.9 g kg−1. Before plantation establishment, clear-cutting, slash piling and burning, and complete site tillage were carried out. Immediately after plantation establishment, calcium superphosphate was applied at 100 g tree−1 into a 5-cm deep tunnel circling the tree; no fertilization was conducted before harvesting.
2.2 Soil sampling
In each selected plantation, three plots (each 20 m × 20 m) were established for soil sampling. Within each plot, five soil profile pits were dug on an S-shaped route across the plot. Soil samples of 1 kg were collected from the 0–20, 20–40, and 40–60-cm layers of each profile, respectively. Soils collected from the same layer of the five profiles were mixed evenly and a composite soil sample was extracted using the quartering method (dividing the soil into quarters and collecting from the two quarters facing opposite one another), which was regarded as representative of the given soil layer of the plot.
Additionally, in each plot, a composite soil sample was collected from the 0–40-cm layer of the above-mentioned pits, which was designated as the non-rhizosphere soil (NRS) sample. When collecting NRS soil, only soils > 3 cm from visible roots was collected. Five dominant trees in each plot were selected randomly from which to collect rhizosphere soil (RS). Under each tree, full-length roots with diameter ≤ 2 mm in the 0–40-cm soil layer in the pits were randomly chosen and 200 g soil within 1 cm of the root surface was collected. Soil sampled from the roots of the five trees was mixed to constitute a composite RS sample for the plot.
2.3 Determination of soil properties
The chemical properties of soil were analyzed according to Carter and Gregorich (2008). Soil pH was measured in deionized water (1:2.5 w/v) using a glass-calomel combination electrode. Soil organic carbon (SOC) was first measured by wet oxidation, and then was calculated as soil organic matter (SOM) using the transfer factor 1.724. Both total nitrogen and available nitrogen were measured with the modified Kjeldahl method. Extraction of total and available phosphorus was accomplished by digestion with H2SO4–HClO4 and extraction with HCl–H2SO4, respectively, and then measured using the molybdenum-blue method. Total and available potassium were determined by digestion with HF and extraction with NH4OAc, respectively, and flame photometry.
2.4 Statistical analyses
All data were analyzed using SAS. One-way analysis of variance (ANOVA) was conducted to determine the changes in soil pH, SOM and nutrient contents. If significant effects were detected (α = 0.05), Duncan’s multiple range test (DMRT) procedure was used to separate the mean values of each plantation.
3 Results
3.1 Soil pH and SOM under CFPs of different ages and rotations
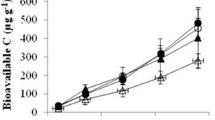
Fluctuations of soil pH among plantations of different rotations and ages were observed in every soil layer. Soil pH values in the 0–20 and 20–40-cm soil layers under the 3-m plantation were significantly lower than their respective counterparts under the 1-m plantation (Fig. 1). However, significant increases in soil pH were observed in the 20–40 and 40–60-cm layers at the transition from 1-m to 2-y plantations. For 0–20-cm layers, no significant change in soil pH were observed at the transition from 1-m to 2-y, or from 2-m to 3-y, indicating that pH did not decrease consistently with the transition of rotation in CFPs.

Mean soil pH (±SE) under different Chinese fir plantations. Bars marked with different letters within the same pane are significantly different (Duncan’s test, α = 0.05)
SOM content in the surface soil generally increased with CFPs ages and rotations (Fig. 2), with SOM content in the 0–20-cm layer under the 3-m plantation increasing 200 % over that of the 1-y plantation. SOM content increases were less dramatic and consistent in the 20–40 and 40–60-cm layers, with SOM contents of the 3-y and 3-h plantations being significantly greater than that of the 2-h plantation in the 20–40-cm layer, and SOM contents of the 3-y plantation being significantly greater than those of the 1-y, 1-h, and 2-h plantations in the 40–60-cm layer.

Mean soil organic matter contents (±SE) under different Chinese fir plantations. Bars marked with different letters within the same pane are significantly different (Duncan’s test, α = 0.05)
3.2 Soil nutrient contents under CFPs of different ages and rotations
Soil TN and available N fluctuated with rotation and age (Figs. 3 and 4, respectively). Available N in the surface soil was lowest under 1-y plantations and highest under 2-m plantations, and this was consistent at other soil depths (Fig. 3). Available N in all soil layers increased with CFPs growth in the first rotation, reached a plateau under 2-m plantations, and then dropped and remained somewhat stable in the third rotation (Fig. 3).

Mean total nitrogen contents (±SE) under different Chinese fir plantations. Bars marked with different letters within the same pane are significantly different (Duncan’s test, α = 0.05)

Mean available nitrogen contents (±SE) under different Chinese fir plantations. Bars marked with different letters within the same pane are significantly different (Duncan’s test, α = 0.05)
Available P was extremely low in each of the three soil layers, with contents below 5, 4, and 2 mg kg−1 for the 0–20, 20–40, and 40–60-cm layers, respectively (data not shown). Available K content in the surface soil was lowest in the 2-m plantations and highest in the 2-h plantations, but overall available K content did not show any obvious pattern in response to plantation age or rotation (data not shown).
3.3 Rhizosphere soil pH, SOM, and nutrients
In our study, mean pH values of the RS were lower than those of the NRS for all plantations; however, only the difference in pH values between RS and NRS in the 3-m plantation was statistically significant. The pH values in RS in 1-m plantations declined from 3.95 to 3.75 in 2-m plantations and further to 3.60 in 3-m plantations, exhibiting notable acidification in RS. Within the second and third rotations, pH in RS dropped sharply with plantation growth (Table 2).
In plantations of the first and second rotations, SOM in RS was higher than that in NRS; in the 3-y plantations, SOM was similar in RS and NRS; in 3-h and 3-m plantations, however, SOM was lower in RS than in NRS (Table 2). In addition, a decrease in SOM in both RS and NRS occurred at the transition from 1-m to 2-y and 2-m to 3-y.
Contents of TN, available N, and available K in RS showed little difference from those of NRS and no differences were significant under any plantation except available K under 3-h (Tables 2 and 3). Mean available P contents, however, were always lower in RS than in NRS, with significant differences occurring under 1-m, 2-m, 3-y, 3-h, and 3-m plantations (Table 3). In this study, it appears that P depletion in the rhizosphere became severe under third-rotation plantations.
4 Discussion
Soil pH is one of the most important soil chemical indices, influencing plant nutrient availability and many other soil properties. Yang et al. (2001) reported that surface soil pH in the first-rotation CFPs dropped from 5.02 to 4.90 in third-rotation stands, while our results revealed a drop from 4.20 to 3.90 over the same period, confirming the previously described acidification trend. However, we observed pronounced pH increases in the 20–40 and 40–60-cm layers at the transition from 1-m to 2-y plantations, which might be a result of fertilization and/or slash burning between rotations (Chen 1998). Burning has been shown to accelerate decomposition of organic acids in soil and to return basic ions to the soil, thereby increasing pH (Tanaka et al. 2005). Subsequent decreases in soil pH likely resulted from base ions being taken up by trees while organic acid root exudations gradually re-accumulated in the rhizosphere.
SOM serves as a storage reservoir for soil nutrients and has important effects on almost all soil chemical, physical, and biological properties (Tanaka et al. 2005); consequently, SOM is generally accepted as an important criterion for assessment of sustainable forest management (Haynes and Beare 1996). Chen et al. (2013) showed that soil organic C (SOC) pools in CFPs increased only between 16 to 21 years, and then stabilized until reaching an over-mature stage. Moreover, increasing rotation length could cause a decrease in SOC (Skjemstad et al. 2008). Our results showed that SOM in the surface soil generally increased with age of CFPs. Further, although non-significant, 2-y and 3-y plantations exhibited a decrease in mean SOM of surface soil (0–20 cm) relative to 1-m and 2-m plantations, respectively, suggesting that SOM in the surface soil declines after harvesting. In accordance with previous studies, we conclude that SOM in CFPs in our study exhibited changes because (1) the increase of both litterfall and fine-root biomass of Chinese fir trees can cause a high accumulation of SOM from young to mature stage in the same rotation (Skjemstad et al. 2008; Chen et al. 2013); however, (2) intensive management practice in forest harvesting, such as clear-cutting, burning and/or removal of harvest residue and forest litter have led to a dramatic decrease in SOM after harvesting or in the young stage of next rotation (Yang et al. 2005; Guo et al. 2016).
Surface soil TN remained relatively stable, with minor fluctuations throughout the experiment. This result was inconsistent with that of Luo and Zhang (2007), who found that nitrogen content in soils under second-rotation CFPs were 8.6 % lower than those of the first rotation, while those under the third rotation were 23.0 % lower than the second. Wang et al. (2010) attributed such decreasing soil nitrogen contents to relatively low nitrogen return through litterfall over the course of consecutive rotations.
The rhizosphere is recognized as one of the most important compartments of terrestrial ecosystems due to its major effect on biogeochemical cycles (Raynaud 2010). Rhizosphere soil has been found to be quite different in chemical properties and nutrient availability from NRS due to root absorption and rhizodeposition activities (Bi et al. 2007). In our study, pH values in the RS were lower than those in the NRS under all plantations (Table 3), although this was only significant in 3-m plantations. This pH difference may be partly because Chinese fir absorbed more nutrient bases such as K+, Ca2+, and Mg2+ and fewer anions from the soil, which consequently resulted in base reduction in the root-soil interface (Hu et al. 2006), and therefore a decrease in pH. Additionally, root-exuded organic acids, CO2, and HCO3 −, as well as secretions by microorganisms in the rhizosphere, have also been shown to contribute to the acidification of the RS (Ingestad 1987). The pH reduction in RS confirmed to some extent the widely reported soil acidification in continuous cropping of CFPs (Ingestad 1987; Guo et al. 2016).
Nutrient deficiency, particularly that of P, has been commonly regarded as an important factor leading to yield decline of CFPs (Chen 2003; Bi et al. 2007). Of all the nutrients studied, P was identified as most likely to be an important limiting factor to CFPs in the third rotation. In the third rotation, available P decreased (especially for RS) compared with the first and the second rotation (Table 3). Moreover, mean available P content was always lower in RS than in NRS, with the differences under 1-m, 2-m, 3-y, 3-h, and 3-m plantations being significant (Table 3). Over 80 % of total P in CFP ecosystems is organic P, which has a large effect on P availability (Chen 2003). Organic P must be hydrolyzed to available P by phosphatases, however, and this does not meet requirements for Chinese fir tree growth (Chen 2003). P fertilizer, such as calcium superphosphate, is therefore applied routinely. Unfortunately, when phosphate is applied as fertilizer in soils, it is rapidly immobilized owing to fixation by bonding with calcium, magnesium, aluminum, and iron (López-Arredondo et al. 2014). The production and root exudation of organic acids is believed to be an important factor in enhancing P availability in soils highly prone to P fixation (López-Arredondo et al. 2014). For example, Yu et al. (2008) reported that organic acid exudation from the roots of Chinese fir increased under stress from low P. Data from Zhao (2012) showed that compared with the first generation CFPs, the content of organic P, Al-P, and Ca-P increased, while the soil P activation coefficient decreased in second-generation CFPs. Moreover, Liao et al. (2001) found that some root exudations, especially HPO4 2−, significantly decreased in the second-generation CFPs. This suggests that low availability of P in the third rotation was principally due to the fact that P is prone to chemical or organic fixation, while both root exudations and microbial activity in soil decreased with succession of CFPs. The extent to which these limitations in P supply during latter rotations of CFPs may be overcome by fertilization and other means should be the target of further research.
In this study, although some nutrients showed a decreasing trend in the consecutive rotation system, no statistically significant decrease in any nutrient content in any of the soil layers was observed under 3-m plantations relative to the beginning of the first rotation. This implies that there may be other factors contributing to the observed yield decline of CFPs. For example, autotoxicity was reported to be a major problem for continuous cropping ecosystems and a drawback for Chinese fir growth (Fernandez et al. 2008). Some cyclic dipeptides have been shown to be highly active allelochemicals and found to be highly concentrated in soils of second-rotation CFPs, where they proved to be phytotoxic to Chinese fir (Kong et al. 2008).
Numerous measures have been investigated as a means to help prevent soil degradation and yield decline in consecutively cropped CFPs systems. Intercropping was conducted in young CFPs and proved a good silvicultural practice for ameliorating nutrient depletion (Wang et al. 2005). Additionally, mixed plantation practices have been recommended for Chinese fir planting by many scientists to maintain soil fertility (Luan et al. 2010). Furthermore, it has been proposed that slash burning and overall tillage should be abandoned and replaced with no-tillage site preparation and herbicide application (Burger 2009). Reasonable and efficient fertilization systems should be identified and alternative cropping methods, other than continuous cropping systems, established in regions where soil degradation has resulted from consecutive rotation practices.
One shortcoming of this study is the lack of stand replicates, associated with the logistical challenge of identifying multiple stands across the landscape. As soil characteristics are highly heterogeneous, especially in hilly regions in south China, the lack of stand replicates might compromise the reliability of our conclusions in examining the effects of continuous cropping on soil fertility. In spite of these limitations, however, this study still provides useful insight into effects of forest management in tree plantations on soil properties.
5 Conclusions
Soil pH in the 0–20 and 20–40-cm layers beneath mature CFPs decreased with consecutive rotations; however, soil pH did not decrease consistently with the transition of CFPs rotation, potentially as a result of anthropogenic activities, such as slash burning and fertilization between rotations. Accumulation of SOM in the surface soil was observed under continuously cropped CFPs, with SOM content under 3-m plantations being two times higher than that under 1-y plantations. TN, available N, and available K did not show any obvious patterns with age and rotation. TN, available N, and available K contents in RS showed little difference from those in NRS; however, pH and available P in RS were consistently lower than in NRS and tended to decrease with continuous cropping. Generally, soil degradation in the continuous cropping system of our study appeared less serious than expected based upon previous reports. Soil acidification and available P deficiency appear to be two of the major potential problems resulting from Chinese fir continuous rotation practices.
References
Bi JBJ, Blanco JABJ, Seely BSB, Kimmins JPKJ, Ding YDY, Welham CWC (2007) Yield decline in Chinese-fir plantations: a simulation investigation with implications for model complexity. Can J For Res 37:1615–1630
Burger JA (2009) Management effects on growth, production and sustainability of managed forest ecosystems: past trends and future directions. For Ecol Manag 258:2335–2346
Carter MR, Gregorich EG (2008) Soil sampling and methods of analysis. CRC Press, Boca Raton
Chen H (1998) Biomass and nutrient distribution in a Chinese-fir plantation chronosequence in Southwest Hunan, China. For Ecol Manag 105:209–216
Chen H (2003) Phosphatase activity and P fractions in soils of an 18-year-old Chinese fir (Cunninghamia lanceolata) plantation. For Ecol Manag 178:301–310
Chen L, Wang S (2003) Preliminary study of allelopathy of root exudates of Chinese fir. Acta Ecol Sin 23:393–398, in Chinese
Chen GS, Yang ZJ, Gao R, Guo JF, Xie JS, Huang ZQ, Yang YS (2013) Carbon storage in a chronosequence of Chinese fir plantations in southern China. For Ecol Manag 300:68–76
Eriksson H, Johansson U (1993) Yields of Norway spruce (Picea abies (L.) Karst.) in two consecutive rotations in southwestern Sweden. Plant Soil 154:239–247
Fang S, Li H, Sun Q, Chen L (2010) Biomass production and carbon stocks in poplar-crop intercropping systems: a case study in northwestern Jiangsu, China. Agrofor Syst 79:213–222
Fernandez C, Voiriot S, Mévy J, Vila B, Ormeño E, Dupouyet S, Bousquet-Mélou A (2008) Regeneration failure of Pinus halepensis mill: the role of autotoxicity and some abiotic environmental parameters. For Ecol Manag 255:2928–2936
Fukami T, Wardle DA (2005) Long-term ecological dynamics: reciprocal insights from natural and anthropogenic gradients. P Roy Soc B-Biol Sci 272:2105–2115
Guo J, Chen G, Xie J, Yang Z (2016) Clear-cutting and slash burning effects on soil CO2 efflux partitioning in Chinese fir and evergreen broadleaved forests in subtropical China. Soil Use Manag. doi:10.1111/sum.12243
Haynes RJ, Beare MH (1996) Aggregation and organic matter storage in meso-thermal, humid soils. Structure and organic matter storage in agricultural soils. CRC Press, Boca Raton, pp 213–262
Hu YL, Wang SL, Zeng DH (2006) Effects of single Chinese fir and mixed leaf litters on soil chemical, microbial properties and soil enzyme activities. Plant Soil 282:379–386
Huang Y, Wang SL, Feng ZW, Ouyang ZY, Wang XK, Feng ZZ (2004) Changes in soil quality due to introduction of broad-leaf trees into clear-felled Chinese fir forest in the mid-subtropics of China. Soil Use Manag 20:418–425
Ingestad T (1987) New concepts on soil fertility and plant nutrition as illustrated by research on forest trees and stands. Geoderma 40:237–252
Kong CH, Chen LC, Xu XH, Wang P, Wang SL (2008) Allelochemicals and activities in a replanted Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook) tree ecosystem. J Agr Food Chem 56:11734–11739
Liao LP, Deng SJ, Yu XJ, Han SJ (2001) Growth, distribution and exudation of fine roots of Chinese fir trees grown in continuously cropped plantations. Acta Ecol Sin 21:569–573, in Chinese
Liu L, Duan ZH, Xu MK, Hu JC, Wang SL, Hu ZG, Zhang QR, Wang SJ (2010) Effect of monospecific and mixed Cunninghamia lanceolata plantations on microbial community and two functional genes involved in nitrogen cycling. Plant Soil 327:413–428
López-Arredondo DL, Leyva-González MA, González-Morales SI, López-Bucio J, Herrera-Estrella L (2014) Phosphate nutrition: improving low-phosphate tolerance in crops. Annu Rev Plant Biol 65:95–123
Luan J, Xiang C, Liu S, Luo Z, Gong Y, Zhu X (2010) Assessments of the impacts of Chinese fir plantation and natural regenerated forest on soil organic matter quality at Longmen mountain, Sichuan, China. Geoderma 156:228–236
Luo YJ, Zhang XQ (2007) The assessment of soil degradation in successive rotations of Chinese fir plantation and the soil amelioration of mixed plantation of Chinese fir and broad-leaved. Acta Ecol Sin 27:715–724, in Chinese
Ma X, Heal KV, Liu A, Jarvis PG (2007) Nutrient cycling and distribution in different-aged plantations of Chinese fir in southern China. For Ecol Manag 243:61–74
Raynaud X (2010) Soil properties are key determinants for the development of exudate gradients in a rhizosphere simulation model. Soil Biol Biochem 42:210–219
Skjemstad JO, Krull ES, Swift RS, Szarvas S (2008) Mechanisms of protection of soil organic matter under pasture following clearing of rainforest on an Oxisol. Geoderma 143:231–242
State Forestry Administration of the People’s Republic of China (2014) Forest resources in China: The 8th National Forest Inventory. http://211.167.243.162:8085/8/book/jiankuang/index.html
Sun QW, Yang CD, Jiao RZ (2003) PCA on the soil degradation of the successive Chinese fir plantation. For Res 16:689–693, in Chinese
Tanaka S, Kendawang JJ, Yoshida N, Shibata K, Jee A, Tanaka K, Ninomiya I, Sakurai K (2005) Effects of shifting cultivation on soil ecosystems in Sarawak, Malaysia IV. Chemical properties of the soils and runoff water at Niah and Bakam experimental sites. Soil Sci Plant Nutr 51:525–533
Wang H, Huang Y, Huang H, Wang KM, Zhou SY (2005) Soil properties under young Chinese fir-based agroforestry system in mid-subtropical China. Agrofor Syst 64:131–141
Wang Q, Wang S, Xu G, Fan B (2010) Conversion of secondary broadleaved forest into Chinese fir plantation alters litter production and potential nutrient returns. Plant Ecol 209:269–278
Wear DN, Greis JG (2002) Southern forest resource assessment: summary of findings. J For 100:6–14
Wei X, Blanco JA, Jiang H, Kimmins JP (2012) Effects of nitrogen deposition on carbon sequestration in Chinese fir forest ecosystems. Sci Total Environ 416:351–361
Wu YL, Wang B, Zhao C, Dai W, Li P (2011) Comprehensive evaluation of soils fertility in different developing stages of Chinese fir plantations. J Northwest A&F Univ 39:69–75, in Chinese
Xu JC, Ai XH, Deng XQ (2005) Exploring the spatial and temporal dynamics of land use in Xizhuang watershed of Yunnan, southwest China. Int J Appl Earth Obs 7:299–309
Yang YS, He ZM, Chen GS, Xi JS, Yu XT (2001) PCA of soil fertility under different gaps of continuously planting Chinese fir. Soil Environ Sci 10:33–38, in Chinese
Yang YS, Guo J, Chen G, Xie J, Gao R, Li Z, Jin Z (2005) Carbon and nitrogen pools in Chinese fir and evergreen broadleaved forests and changes associated with felling and burning in mid-subtropical China. For Ecol Manag 216:216–226
Yao MS, Xie YQ (1991) The effects of undergrowth on productivity of Chinese fir plantation. For Res 3:246–252, in Chinese
Yu YC, Yu J, Shan Q, Fang L, Jiang D (2008) Organic acid exudation from the roots of Cunninghamia lanceolata and Pinus massoniana seedlings under low phosphorus stress. Front For China 3:117–120
Zhang X, Kirschbaum MUF, Hou Z, Guo Z (2004) Carbon stock changes in successive rotations of Chinese fir (Cunninghamia lanceolata (lamb) hook) plantations. For Ecol Manag 202:131–147
Zhao JR (2012) The influence of Chinese fir ecosystem conversion on the soil phosphorus forms and its mechanism. Fujian Agriculture & Forestry University, Dissertation, in Chinese
Zhao MF, Xiang WH, Tian DL, Deng XW, Huang ZH, Zhou XL, Peng CH (2013) Effects of increased nitrogen deposition and rotation length on long-term productivity of Cunningham lanceolata in Southern China. PLoS ONE 8:e55376
Zhou L, Shalom ADD, Wu P, Li S, He Z, Liu C, Ma X (2016) Biomass production, nutrient cycling and distribution in age-sequence Chinese fir (Cunninghamia lanceolate) plantations in subtropical China. J For Res 27:357–368
Acknowledgments
This paper was financially supported by the National Natural Science Foundation of China (No. 31270664, No. 31270675 and No. 30271069), National Key Research and Development Program of China (No. 2016YFNC030025), and Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Hailong Wang
Rights and permissions
About this article

Cite this article
Yu, Y., Yang, J., Zeng, S. et al. Soil pH, organic matter, and nutrient content change with the continuous cropping of Cunninghamia lanceolata plantations in South China. J Soils Sediments 17, 2230–2238 (2017). https://doi.org/10.1007/s11368-016-1472-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-016-1472-8