
Abstract
Litter production, litter standing crop, and potential nutrient return via litterfall to soil were studied during a 4-year period (January 2004–December 2007) in a Chinese fir (Cunninghamia lanceolata (Lamb.) Hook) plantation and a secondary broadleaved forest in Hunan Province in subtropical China. Mean annual litterfall in the sampling sites varied from 358 g m−2 in the pure plantation to 669 g m−2 in the secondary broadleaved forest. Total litterfall followed a bimodal distribution pattern for both forests. Amount of litterfall was also related to the air temperature in both forests. During the period under this study, annual variation in the total litterfall in the pure plantation was significantly higher than that in the secondary broadleaved forest. Litterfall was markedly seasonal in the both forests. Leaf proportions of litterfall in the pure plantation and secondary broadleaved forest were 58.1 and 61.7%, respectively. Total potential nutrient returns to the soil through litterfall in the pure plantation were only 46.2% of those in the secondary broadleaved forest. Total litter standing crop was 913 and 807 g m−2 in the pure plantation and secondary broadleaved forest, respectively. Our results confirm that conversion from a secondary broadleaved forest into a pure coniferous plantation changes the functioning of the litter system.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The area of natural/secondary forests has been shrinking on an average of 16 million ha annually during 1990–2000 due to the demand for forest products (West 2006). In subtropical China, extensive areas of native broadleaved forests have also been converted to mostly pure coniferous plantations such as Chinese fir (Cunninghamia lanceolata (Lamb.) Hook), covering over 1.21 million ha and accounting for 6.5% of all plantation forests in the world (Chen and Wang 2004; FAO 2006). Since the 1980s, decline in yield and soil fertility of pure Chinese fir stands has been observed due to successive planting, short-rotation time, and stem-only harvesting (Chen et al. 1990). As such, there has been increasing concern regarding the effects of conversion of native broadleaved forests to plantations on ecosystem functions such as nutrient cycling, and soil fertility, particularly from the perspective of sustainability of plantation forests (Lian and Zhang 1998; Goma-Tchimbakala and Bernhard-Reversat 2006; Pandey et al. 2007; Wang and Wang 2007).
Litterfall is closely related to the productivity of managed forests. It is the main pathway for the return of nutrients from plants to soil and for the formation of soil organic matter in all forest ecosystems (Spain 1984). In small regions, litterfall is strongly influenced by tree species composition, density, successional stage, and soil fertility (Meentmeyer 1982; Arunachalam et al. 1998). Many reports have suggested that the conversion of natural forests to plantations strongly affects litterfall, litter standing crop, litter quality, and decomposition (Lian and Zhang 1998; Yang et al. 2004; Goma-Tchimbakala and Bernhard-Reversat 2006; Pandey et al. 2007). Some studies have revealed that natural forests have higher litterfall than pure plantations (Lian and Zhang 1998; Yang et al. 2004; Pandey et al. 2007). However, the impacts of the conversion of secondary broadleaved forests to pure Chinese fir plantations on litter production in subtropical China are still not clearly known.
On soil, litter acts as a temporary sink of nutrients, which are released slowly. Its decomposition rate regulates nutrient cycling and maintains soil fertility. While a number of previous studies regarding Chinese fir have emphasized litter amount, composition, and distribution (Lian and Zhang 1998; Wu and Hong 2002; Fang et al. 2005), data on the ecological role of Chinese fir litterfall in nutrient cycling are scarce. In recent years, Mlambo and Nyathi (2008) evaluated the ecological role of Colophospermum mopane litterfall in nutrient cycling in semi-arid southern African savanna woodland. Wang et al. (2008) also evaluated the ecological function of Chinese fir litterfall in nutrient cycling. However, little information is available regarding the impacts of secondary broadleaved forests conversion to coniferous plantations on the ecological role of litterfall in nutrient cycling in subtropical China. This has also limited the research linking nutrient return through litterfall and soil nutrient, although in a previous study these authors reported that soil organic carbon and nitrogen pools were higher in a secondary broadleaved forest than in a Chinese fir plantation (Wang and Wang 2007).
This study attempts to explore the effects of forest transformation on litter production, litter standing crop, litter quality, and potential nutrient return to the soil via litterfall in a Chinese fir plantation and a secondary broadleaved forest in the subtropical region of southern China. We test for the following hypotheses: (1) there is significant difference in the litter dynamics and composition between the pure plantation and the secondary broadleaved forest; (2) the Chinese fir plantation has higher litter standing crop than the secondary forest; and (3) potential nutrient returns through litter to the soil are higher in the secondary broadleaved forest than in the pure coniferous plantation.
Materials and methods
Study sites
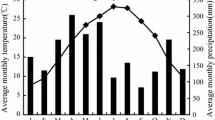
The study site is located at the Huitong Experimental Station of Forest Ecology, Chinese Academy of Sciences (26°45′ N, 109°30′ E, 510 m a.s.l.), in Hunan Province, Southern China. The climate of this region is humid mid-subtropical monsoon, with the wet season from March to July and the dry season from August to February. The site receives an annual average rainfall of 1200 mm with 60–72% falling between March and July. It has a mean annual temperature of 16.5°C (Fig. 1). The mean maximum temperature varies from 3.4 to 31.9°C, while the mean minimum ranges from 0.8 to 23.6°C from January 2004 to December 2007.

Changes in monthly rainfall (mm) and temperature (°C) during the course of the study in Huitong Experimental Station of Forest Ecology, Chinese Academy of Sciences, in Hunan Province, China
In subtropical China, native vegetation is evergreen broadleaved forests, which have almost been extirpated by human activities. In this study, the relatively undisturbed secondary forest (SF), which is about approximately 45 years old, is covered by subtropical evergreen broadleaved species dominated by Castanopsis hystrix, Cyclobalanopsis glauca, Machilus Pauhoi, Liquidambar formosana, and Juglans cathayensis (Deng et al. 2000). This forest was destroyed by a man-made disturbance in 1958 and has since then been protected and restored through natural regeneration. The secondary broadleaved forest covers 2.5 ha. In 2005, its measured tree density was 937 stems ha−1 with 9.0 m for tree height and 13.9 cm for tree diameter at breast height (DBH). The pure plantation of Chinese fir (PC) with 2,000 trees ha−1 was established in the early spring of 1983 after the clear-cutting of secondary broadleaved forests in the autumn of 1982 and burning in the winter. For the 3.2 ha Chinese fir plantation, tree density, height, and DBH were 1050 stems ha−1, 16.9 m, and 21.65 cm, respectively. In both stands, the soil, which has been categorized as Oxisol according to US Taxonomy, were developed from the same parent material. The physical and chemical properties of the soil are listed in Table 1.
Litterfall and standing litter sampling collection
Litterfall was collected for a 4-year period starting from January 2004 to December 2007 in a secondary broadleaved forest and a pure Chinese fir plantation. Litter collection was conducted within the three permanent plots of 15 × 15 m established in 1998 for long-term research of forest ecology in each stand. Ten 1 × 1 m litter traps made of nylon mesh (1 mm mesh size) at 50 cm above the ground were randomly placed in each permanent plot (30 traps in each stand). Overall, 60 traps were placed in both stands. Litterfall was collected semi-monthly. The collected litters at each time were taken to the laboratory and categorized into leaves, branches, barks, reproductive parts (e.g., flowers and fruits), and miscellaneous materials (e.g., insect fecal and unidentified plant parts) and were oven-dried at 70°C to constant weight. Each month, the dried litters in each plot were combined. The mean monthly litter mass for each component of each plot was calculated on a unit area basis (g m−2).
Litter standing crop was sampled at the beginning of each year by collecting litter from the surface of the forest floor (L horizon). Ten samples of 0.25 m2 each were randomly taken at each permanent 15 m × 15 m plot. The litter samples were then transported to the laboratory, sorted into leaves, branches, reproductive parts, and miscellaneous materials. Samples were then dried to constant weight at 70°C.
Chemical analyses
The oven-dried litter samples were ground, sieved through a 0.5 mm mesh, and analyzed for total N, P, K, Ca, and Mg concentration. Total N concentration was determined by the micro-Kjeldahl method by digesting 0.5-g samples in 10 ml concentrated H2SO4 using a catalyst mixture (CuSO4, K2SO4 and selenium powder) and distillation. In order to measure P, 0.2-g litter samples were digested in 10 ml triacid mixture (nitric, perchloric, and sulphuric acid; 5:1:1) and then cooled. Total P was colorimetrically determined in the digested samples by ammonium molybdate stannous chloride method (Olsen and Sommers 1982). Concentrations of K, Ca, and Mg in the digested solution were determined with a flame atomic absorption spectrophotometer (Hitachi Z-8100, Tokyo, Japan) following HClO4–HNO3 digestion (Jones and Case 1990).
All chemical analyses of the litter samples were carried out in triplicate on the each sample, and the mean of each triplicate was taken. There were no significant differences among the triplicates.
Statistical analyses
The monthly potential input of each nutrient to the soil through each litter fraction was obtained by multiplying the values of each fraction mass by its corresponding nutrient concentrations (Lian and Zhang 1998). Annual nutrient input was the sum of the monthly nutrient inputs based on 12 monthly estimations. Data on the mean annual litterfall, potential nutrient return, litter standing crop, and its nutrient storage were analyzed through a one-way ANOVA using SPSS version 17.0 for Windows. Tukey’s Honestly Significant Difference test was then used for multiple comparisons at a significant level of 0.05 between the pure plantation and secondary broadleaved forest. Pearson’s linear correlations were used to assess the relationships between litterfall amount and abiotic variables (temperature and rainfall).
Results
Litter production
Litterfall mass was significantly lower in the pure Chinese fir plantation than in the secondary broadleaved forest (Table 2). The average annual litterfall was 358 g m−2 in the Chinese fir plantation, with leaves, branches, barks, reproductive parts, and miscellaneous parts accounting for 58.1, 23.5, 1.1, 14.5, and 2.6%, respectively, of total litterfall. In the secondary broadleaved forest, annual litterfall was 669 g m−2, with leaves, branches, and reproductive parts accounting for approximately 95% of total litterfall.
Total litterfall conformed to a bimodal distribution pattern in the both forests (Fig. 2). However, the peaks of litterfall between both forests occurred in different periods. In the secondary broadleaved forest, one peak occurred in April–May and the other in November from 2004 to 2007. In the pure Chinese fir plantation, one peak occurred in February during the 4-year period, while the others occurred in November 2004–2006, and in December 2007. The amount of litterfall in the pure plantation was negatively correlated with the abiotic variables, whereas it was positively correlated in the secondary broadleaved forest (Figs. 1 and 2). The amount of litterfall was more strongly related to air temperature in the pure plantation (r = 0.40 to 0.43, P < 0.05) than in the secondary broadleaved forest (r = 0.16 to 0.21, P < 0.05).

Monthly litterfall (mean ± SE) from the secondary broadleaved forest (SF) and pure Chinese fir plantation (PC) stands. SE is standard error
Annual variation in total litterfall of the pure plantation (variation coefficient at 38.0%) was significantly higher than that of the secondary broadleaved forest (variation coefficient at 9.8%) during the period of this study. The annual litterfall varied from 257 g m−2 in 2005 to 558 g m−2 in 2007 in the pure plantation and from 573 g m−2 in 2006 to 715 g m−2 in 2004 in the secondary broadleaved forest.
Litterfall was markedly seasonal, although it occurred continuously throughout the year in the both forests (Fig. 3). Seasonal patterns of total litterfall differed between the forests. For the secondary broadleaved forest, the highest litterfall occurred in spring, accounting for approximately 39%. This was followed by autumn, which accounted for approximately 28%. Summer and winter accounted for approximately 17 and 16%, respectively. In the pure Chinese fir plantation, litterfall occurred mainly in winter, accounting for over 47%. This was followed by spring and autumn, which accounted for approximately 25 and 21%, respectively. Summer accounted for less than 7%. Approximately 64% of the total litterfall in the pure plantation was recorded during the cool and dry period (from November to February), while approximately 30% was recorded in the secondary broadleaved forest during the same period.

Seasonal variations (mean ± SE) in litterfall from January 2004 to December 2007 in the secondary broadleaved forest (SF) and pure Chinese fir plantation (PC). Spring, March–May; summer, June–August; autumn, September–November; winter, December–February. SE is standard error
Nutrient content in litterfall fractions and potential returns to the soil
The concentrations of N, P, K, Ca, and Mg in the litterfall differed among the five fractions (Table S1). The concentrations of N and Ca were the highest in reproductive litter in the pure plantation and in leaf in the secondary broadleaved forest. Among all litter fractions, reproductive litter had the highest P and K concentrations in both pure plantation and secondary broadleaved forest. P concentration was 2.2 and 2.6 times greater in reproductive litter than in branch in the pure plantation and secondary broadleaved forest, respectively, while K concentration was 2.6 and 3.2 times greater. Mg concentration was the highest in branch and the lowest in bark in the pure plantation, while being highest in leaf and the lowest in miscellaneous in the secondary broadleaved forest.
Annual potential nutrient returns to the soil through litter were significantly lower in the pure plantation than in the secondary broadleaved forest (Table 3). Mg approximately 6.3 g m−2 in the pure plantation to 12.4 g m−2 in the secondary broadleaved forest was generally returned to the soil in the highest amount, followed by N with a range of 1.9–5.7 g m−2, Ca with a range of 2.0–4.6 g m−2, and K with a range of 1.7–2.9 g m−2. The P return to the soil through litterfall was much less than the other nutrients, ranging from 0.32 g m−2 in the pure plantation to 0.65 g m−2 in the secondary broadleaved forest. The percent contribution of leaf litter to total nutrient return was significantly lower in the pure plantation ranging from 50% (Mg) to 69% (K) than in the secondary broadleaved forest ranging from 69% (P) to 86% (N). In terms of nutrient return, leaf and branch fractions were the main components of litterfall, accounting for over 80% in both forests.
Annual nutrient returns via litterfall in the secondary broadleaved forest were relatively stable across the 4-year observation period compared with the pure plantation. The yearly return ratios for 2004, 2005, 2006, and 2007 for all nutrients were 1:0.78:0.99:1.79, respectively, in the pure plantation and 1:1.02:0.80:0.90, respectively, in the secondary broadleaved forest. The pattern of nutrient returns through litter was similar to litterfall in both forests.
Litter standing crop and nutrient storage
In general, the pure plantation showed a higher mean standing crop of litter than the secondary broadleaved forest, although no significant difference was found between both forests (Table 4). The proportions of litter represented by leaves, branches, reproductive litter, and miscellaneous were different between the two forests and among years. Leaves were the highest in proportion, comprising over 50% of the standing crop of litter, followed by branches.
The concentrations of all the studied nutrients in the standing crop of litter differed between the sampling forests (Table S2). Among the different fractions, P and Mg concentrations in the standing crop of litter were significantly or slightly lower than those in litterfall, and K concentration was lower in the standing crop of litter than in litterfall, except for leaves, in the secondary broadleaved forest. In contrast, the concentration of N was higher in the standing crop of litter, except for the miscellaneous fraction, than in litterfall in the secondary broadleaved forest; N was only higher in branch fraction in the pure plantation. The storage of N, K, and Mg in the standing crop of litter was significantly lower in the pure plantation than in the secondary broadleaved forest (Table 5).
Discussion
Based on the analysis of the effects for 4 years on a number of forest plots, we have considerable confidence in our findings. Results of this study suggest that litter production is significantly reduced due to the forest conversion of the secondary broadleaved forest to a coniferous plantation, which are consistent with other observations regarding other tree species in some regions (Lian and Zhang 1998; Yang et al. 2004; Pandey et al. 2007). Lian and Zhang (1998) found that annual litterfall was significantly lower in a 31-year-old pure Chinese fir plantation (4.8 t ha−1) than in a native forest of broadleaved evergreen Castanopsis kawakamii (13.3 t ha−1), while Pandey et al. (2007) observed that a managed plantation had a slightly lower litterfall than a natural Quercus serrata forest in a subtropical region in Northeastern India. In contrast, Goma-Tchimbakala and Bernhard-Reversat (2006) found that annual litterfall was slightly but not significantly higher in a Terminalia superba plantation than in a natural tropical forest in Mayombe, Congo.
Litter production in forest ecosystems mainly depends on site fertility and global climatic conditions (Jorgensen et al. 1975; Arunachalam et al. 1998); however, at a local or small scale with the same climatic condition, it is mostly limited by tree species composition and characteristics, nutrient availability, and age structure (Parrotta 1999; Sundarapandian and Swamy 1999; Polyakova and Billor 2007). In subtropical China, short-rotation plantation of Chinese fir generally reaches the mature stage approximately 20–25 years after planting, after which the trees are cut by farmers for timber products. Chinese fir trees had been 21 years old at the beginning of this investigation. Thus, in this study, litter production of Chinese fir had been relatively stable at the mature stage unless unusual weather occurred. The decrease in litterfall is not merely a transitory impact restricted to wood age, as was previously suggested by Lian and Zhang (1998) and Lisanework and Michelsen (1994). Several possible mechanisms can partly explain the effects of the conversion from a native broadleaved forest to a coniferous plantation on litter production. In order to begin with, compared with the native broadleaved forest, the observed decline in litterfall in the pure Chinese fir plantation can be likely attributed, at least in part, to tree species composition, although the decreased litterfall in the pure plantation in this study is not easily explained. Some studies in subtropical regions have observed a more significant increase in litter production in mixed plantations than in coniferous monoculture plantation (Trofymow et al. 2002; Wang et al. 2007; Wang et al. 2008). In this study, the secondary broadleaved forest featured several tree species, but the Chinese fir plantation featured only one tree species. Secondly, the reduction in litter production in the pure plantation may be also partly explained by the competitive production principle (Kelty 2006). It is known that tree species with substantially different characteristics may use site resources more completely and efficiently in producing biomass, resulting in higher total biomass production, than species with similar characteristics. In other words, different species have complementary use of resources (Haggar and Ewel 1997) or good ecological combing ability. In the study, the broadleaved tree species in the secondary forest had different characteristics such as crown structure, height growth rate, leaf area density, foliar phenology, and root depth and phenology. Thus, in the secondary forest, broadleaved species with substantially different characteristics may have used site resources more efficiently. Compared to the pure Chinese fir, the broadleaved species may have synthesized more materials due to these complementary characteristics, resulting in greater litterfall. Finally, differences in soil fertility or available nutrients between both forests may be partly explained by the decline in litterfall in the pure plantation. Wang et al. (2006) found that soil total N, P and K, and available N, P, and K in the pure Chinese fir plantation were significantly lower than those in secondary broadleaved forests in the same studied sites.
Conversion from a secondary broadleaved forest to a pure coniferous plantation did not significantly affect litter composition, although it significantly influenced litterfall. From our results, leaf and branch components accounted for over 80% of total litterfall. These components also mainly determined the pattern of total litterfall, which is consistent with other investigations in subtropical forests or plantations (Zhang et al. 1993; Wang et al. 2007, 2008). However, these estimates are lower than the estimates from other species in subtropical plantations (Liao et al. 2000; Pandey et al. 2007).
The litterfall pattern changed due to forest transformation. The peaks of the litterfall were different in both the secondary broadleaved forest and pure plantation; however, there was a bimodal distribution pattern in both forests. Yang et al. (2004) also found a significant difference in litterfall pattern between pure a Chinese fir plantation with a multi-peak and a native broadleaved forest of Castanopsis kawakamii with a unimodal distribution pattern in Fujian, Southern China. In addition, a similar litterfall pattern was observed among one native broadleaved forest, one mixed, and one pure Chinese fir plantation in subtropical China by Lian and Zhang (1998). Correspondingly, our results indicate that the seasonal pattern of litterfall is also significantly affected by forest conversion. From our results, one main peak of total litterfall in the secondary broadleaved forest was observed in spring over the 4-year period, which is in agreement with some studies on evergreen broadleaved forests (Deng et al. 1993; Liao et al. 2000; Yang et al. 2004). The occurrence of litterfall for the evergreen broadleaved forest in spring is likely due to the replacement of senescent leaves with new ones. In the pure Chinese fir plantation, litterfall was concentrated during the cold and dry period (from November to February) of the year, with approximately 64% of total litterfall occurring during this period. This is also comparable to other results in subtropical plantation ecosystems (Zhang et al. 1993; Pandey et al. 2007; Sundarapandian and Swamy 1999). The tendency of litterfall to be concentrated in the cool and dry season may be associated with the shedding of needles induced by low temperature stress and/or low soil moisture during this period (Jackson 1978). As per the analysis of the coefficient between litterfall and air temperature, we found that the amount of litterfall in the pure Chinese fir plantation was significantly affected by air temperature. Another mechanism or hypotheses by Moore (1980) is that water stress causes the synthesis of abscisic acid in the foliage of plants, which in turn stimulates the senescence of leaves and other parts. However, we did not find litterfall to be closely related to rainfall in this study.
Litter standing crop in forest ecosystems is primarily controlled by annual litterfall mass and litter decomposition rate. The results of this study show that litter standing crop in the pure plantation was slightly but not significantly higher than that in the secondary broadleaved forest, which is consistent with the reports of Goma-Tchimbakala and Bernhard-Reversat (2006), who found significant differences in the standing crop with a trend towards a lower amount in a native broadleaved forest than in T. superba plantations. Numerous studies have shown that a large amount of standing litter is accumulated in subtropical and tropical plantations, especially when the planted species has litter with low quality and decomposition rates (Stohlgren 1988; Bernhard-Reversat 1993; Mlambo and Nyathi 2008), although litter may have a high decomposition rate due to the sufficient rainfall and high temperature. In this study, high litter standing crop in the pure plantation is partly attributed to the low decomposition rate of Chinese fir litterfall, which is generally controlled by litter quality and litter species composition within the same climatic region (Taylor et al. 1989; Hoorens et al. 2003; Schere-Lorenzen et al. 2007). Our previous studies also showed that Chinese fir litter with a high C/N ratio and lignin content and a low initial content of N had a lower decomposition rate than the litter of broadleaved tree species (Wang et al. 2008).
The potential nutrient inputs of different elements to the soil via litterfall in the secondary broadleaved forest were 1.7–3.0 times greater than those in the pure plantation. The differences in the nutrient returns observed between both forests are a result of both lower litterfall mass and lower nutrient concentrations in the litter. The results confirm the previous studies reported by Lian and Zhang (1998), who observed that N, P, K, and Ca returns through litterfall in a subtropical native broadleaved forest were significantly higher than in a pure coniferous plantation. The input of nutrients through leaf litterfall dominated the total litterfall nutrient input, although the nutrient contribution of leaf litterfall to total nutrient return was different between the two forests (Lian and Zhang 1998; Wang et al. 2007; Mlambo and Nyathi 2008; Wang et al. 2008). This may be due to the higher proportion of leaf litterfall to total litterfall and the higher nutrient concentrations. These results also indicate that a decline in litterfall may be partially responsible for lower soil nutrients in the surface layer in pure plantations; some studies also suggest that it is crucial for root litter and soil fertility (Liao et al. 1999; Xiao et al. 2008). Unfortunately, the inputs of root litter to soil were not quantified in this study. Liao et al. (1999) found that root production and turnover of Chinese fir was lower than that in the broadleaved tree species in the same region. Low soil fertility in the pure plantation is likely to be due, at least in part, to low production of the root.
Long-term effects of forest conversion on litterfall and nutrient return can be reflected by soil fertility or site productivity. Soil properties investigated in 2005 were different in the secondary broadleaved forest and in the Chinese fir plantation; however, we did not delve into the impacts of forest transformation on soil fertility in this study (Table 1). Soil organic C, total N, and K were higher in the secondary broadleaved forest than in the pure plantation, and soil bulk density and clay content were lower in the secondary forest. This confirms the results of Yang et al. (2005), Wang et al. (2006), and Wang and Wang (2007) in this and other subtropical regions. Xu and Xu (2003) also found that the total organic carbon content of samples from Chinese fir stands was 27.3% lower than that form the broadleaved forest in Huzhou, Zhejiang Province, in East China. The changes in soil fertility are mainly due to the differences in litterfall amount, pattern, and nutrient return.
In conclusion, forest conversion from a secondary broadleaved forest to a pure Chinese fir plantation significantly reduced litterfall mass and potential nutrient inputs through litterfall to the soil, and changed litterfall patterns. In contrast, standing litter was slightly higher in the pure plantation than in the secondary broadleaved forest. This is attributed to the low decomposition rate of Chinese fir litter with low quality. This can explain, at least in part, the decline in soil fertility resulting from forest conversion of a secondary broadleaved forest to a pure coniferous plantation.
References
Arunachalam A, Arunachalam K, Pandey HN, Tripathi RS (1998) Fine litterfall and nutrient dynamics during forest regrowth in the humid subtropics of north-eastern India. For Ecol Manage 110:209–210
Bernhard-Reversat F (1993) Dynamics of litter and organic matter at the soil-litter interface in fast-growing tree plantations on sandy ferrallitic soils (Congo). Acta Ecol 14:179–195
Chen C, Wang S (2004) Ecology of mixed plantation forest. Science Press, Beijing, p 3
Chen CY, Zhang JW, Zhou CL, Zheng HY (1990) Researches on improving the quality of forest land and the productivity of artificial Cunninghamia lanceolata stands. J Appl Ecol 1:97–106
Deng C, Hou J, Li S, Zhao H, Fu J (1993) Study on forest litter in Ailao Mountain, North China. Acta Phytoecol Geobot Sin 17:364–370
Deng S, Liao L, Wang S, Gao H (2000) Bioproductivity of Castanopsis hystrix-Cyclobalanopsis glauca-Machilus Pauhoi community in Huitong, Hunan. Chin J Appl Ecol 1:651–654
Fang X, Tian D, Xiang W, Lei P (2005) Litter production and carbon release ratios in the litter decomposition process of litter in Chinese fir plantations. J Center South For Univer 25:12–16
FAO (2006) Global forest resource assessment 2005. Food and Agricultural Organization of the United Nations, Rome
Goma-Tchimbakala J, Bernhard-Reversat F (2006) Comparison of litter dynamics in three plantations of an indigenous timber-tree species (Terminalia superba) and a natural tropical forest in Mayombe, Congo. For Ecol Manage 229:304–313
Haggar JP, Ewel JJ (1997) Primary productivity and resource portioning in model tropical ecosystems. Ecology 78:1211–1221
Hoorens B, Aerts R, Stroetenga M (2003) Does initial litter chemistry explain litter mixture effects on decomposition. Oecologia 137:578–586
Jackson JF (1978) Seasonality of flowering and leaf fall in a Brazilian subtropical lower montane moist forest. Biotropica 10:38–42
Jones JB, Case VW (1990) Sampling, handling, and analyzing plant tissue samples. In: Westerman RL (ed) Soil testing and plant analysis. Soil Science Society of America, Inc., Madison, WI, pp 390–428
Jorgensen JR, Well CG, Metz LJ (1975) The nutrient cycle. Key to continuous forest production. J For 73:400–403
Kelty MJ (2006) The role of species mixtures in plantation forestry. For Ecol Manage 233:195–204
Lian Y, Zhang Q (1998) Conversion of a natural broad-leafed evergreen forest into pure and mixed plantation forests in a subtropical area: effects on nutrient cycling. Can J For Res 28:1518–1529
Liao L, Yang Y, Wang S, Gao H (1999) Distribution, decomposition and nutrient return of the fine root in pure Cunninghamia lanceolata, Michelia macclurei and the mixed plantations. Acta Ecol Sin 19:342–346
Liao L, Wang S, Chen C (2000) Dynamics of litterfall in the mixed plantation of Cunninghamia lanceolata and Michelia macclurei: a ten-year’s observation. Chin J Appl Ecol 11(Supp):131–136
Lisanework N, Michelsen A (1994) Litterfall and nutrient release by decomposition in three plantations compared with a natural forest in the Ethiopian highland. For Ecol Manage 65:149–164
Meentmeyer V (1982) World patterns and amounts of terrestrial plant litter production. Bioscience 32:125–128
Mlambo D, Nyathi P (2008) Litterfall and nutrient return in a semi-arid southern African savanna woodland dominated by Colophospermum mopane. Plant Ecol 196:101–110
Moore TC (1980) Biochemistry and physiology of plant hormones. Narosa Publishing House and Springer Verlag, New Delhi pp. 274
Olsen SR, Sommers LE (1982) Phosphorus. In: Page AL, Miller RH, Keeney DR (eds) Methods of soil analysis, part 2. Agronomy Society of America and Soil Science Society of America, Madison, WI, pp 403–430
Pandey RR, Sharma G, Tripathi SK, Singh AK (2007) Litterfall, litter decomposition and nutrient dynamics in a subtropical natural oak forest and managed plantation in northeastern India. For Ecol Manage 240:96–104
Parrotta JA (1999) Productivity, nutrient cycling, and succession in single- and mixed-species plantation of Casuarina equisetifolia, Eucalyptus robusta, and Leucaena leucocephala in Puerto Rico. For Ecol Mnaage 124:45–77
Polyakova O, Billor N (2007) Impact of deciduous tree species on litterfall quality, decomposition rates and nutrient circulation in pine stands. For Ecol Manage 253:11–18
Schere-Lorenzen M, Bonilla JL, Potvin C (2007) Tree species richness affects litter production and decomposition rates in a tropical biodiversity experiment. Oikos 116:2108–2124
Spain AV (1984) Litterfall and the standing crop of litter in three topical Australian rainforests. J Ecol 72:947–961
Stohlgren TJ (1988) Litter dynamics in two Sierran mixed conifer forests. I. Litterfall and decomposition rates. Can J For Res 18:1127–1135
Sundarapandian SM, Swamy PS (1999) Litter production and leaf-litter decomposition of selected tree species in tropical forests at Kodayar in the Western Ghats, India. For Ecol Manage 123:231–244
Taylor BR, Parkinson D, Parsons WFJ (1989) Nitrogen and lignin content as predictors of litter decay rates: a microcosm test. Ecology 70:97–104
Trofymow JA, Moore TR, Titus B, Prescott C, Morrison I, Siltanen M, Smith S, Fyles J, Wein R, Camiré C, Duschene L, Kozak L, Kranabetter M, Visser S (2002) Rates of litter decomposition over 6 years in Canadian forests: influence of litter quality and climate. Can J For Res 32:789–804
Wang Q, Wang S (2007) Soil organic matter under different forest types in southern China. Geoderma 142:349–356
Wang Q, Wang S, Feng Z (2006) Comparison of active soil organic carbon pool between Chinese fir plantations and evergreen broadleaved forests. J Beijing For Univ 28:1–6
Wang Q, Wang S, Fan B, Yu X (2007) Litter production, leaf litter decomposition and nutrient return in Cunninghamia lanceolata plantations in south China: effect of planting conifers with broadleaved species. Plant Soil 297:201–211
Wang Q, Wang S, Huang Y (2008) Comparison of litterfall, litter decomposition and nutrient return in a monoculture Cunninghamia lanceolata and a mixed stand in southern China. For Ecol Manage 255:1210–1218
Wu C, Hong W (2002) Dynamics of litterfall in Cunninghamia lanceolata forest using wave-type series analysis. J Trop Subtrop Bot 10:201–206
Xiao C, Sang W, Wang R (2008) Fine root dynamics and turnover rate in an Asia white birch forest of Donglingshan Mountain, China. For Ecol Manage 255:765–773
Xu QF, Xu JM (2003) Changes in soil carbon pools induced by substitution of plantation for native forest. Pedosphere 13:271–278
Yang Y, Guo J, Chen G, Xie J, Cai L, Lin P (2004) Litterfall, nutrient return, and leaf-litter decomposition in four plantations compared with a natural forest in subtropical China. Ann For Sci 61:465–476
Yang Y, Guo J, Lin P, He Z, Chen G (2005) Carbon and nutrient pools of coarse woody debris in a natural forest and plantation in subtropical China. Sci Sil Sin 41:7–11
Zhang J, Liao L, Li J, Su Y (1993) Litter dynamics of Pinus massoniana and Michelia macclurei mixed forest and its effect on soil nutrients. Chin J Appl Ecol 4:359–363
Acknowledgments
This research was supported by the National Natural Science Foundation of China (30590381-07) and the Chinese Academy of Science Program (No. KZCX2-YW-405). We sincerely appreciate the efforts of the two anonymous reviewers who provided comments for the improvement of our manuscript. We are also grateful to the Huitong National Research Station of Forest Ecosystem for providing the study sites and laboratory facilities, and to Xiuyong Zhang for his assistance during sampling.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Wang, Q., Wang, S., Xu, G. et al. Conversion of secondary broadleaved forest into Chinese fir plantation alters litter production and potential nutrient returns. Plant Ecol 209, 269–278 (2010). https://doi.org/10.1007/s11258-010-9719-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-010-9719-8