
Abstract
Preserving muscle mass and strength is critical for long-term health and longevity. Age-related muscle lipid accumulation has been shown to be detrimental to muscle health. In healthy older individuals, we sought to determine whether muscle lipid content, determined from computed tomography, is associated with self-reported physical function, laboratory-measured performance, and the response to progressive resistance training (PRT), and how metformin may alter these responses (N = 46 placebo, 48 metformin). Using multiple linear regression models adjusted for confounders in a large cohort, we show that intermuscular adipose tissue (IMAT) was not associated with baseline function or response to PRT, contrary to previous reports. On the other hand, thigh muscle density (TMD), as an indicator of intra- and extramyocellular lipid (IMCL and EMCL), remained strongly and independently positively associated with physical function and performance following adjustment. Baseline TMD was inversely associated with gains in strength, independent of muscle mass. Percent change in TMD was positively associated with improved chair stand and increased type II fiber frequency but was not associated with muscle hypertrophy or overall strength gain following PRT. For the first time, we show that metformin use during PRT blunted density and strength gains by inhibiting fiber type switching primarily in those with low baseline TMD. These results indicate that participants with higher muscle lipid content derive the most performance benefit from PRT. Our results further indicate that muscle density may be as influential as muscle size for strength, physical function, and performance in healthy older adults. ClinicalTrials.gov, NCT02308228, Registered on 25 November 2014
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Sarcopenia, the age-associated loss of skeletal muscle mass with accompanying loss of physical function, is frequent among older adults and compromises overall health and quality of life [52]. Loss of muscle mass begins as early as age 40 and can increase to a rate of 1% per year by age 75. Strength declines occur more rapidly, and can decrease at a rate of 3–4% per year in men and 2.5–3% per year in women by age 75 [35]. These declines may be attributed to a multitude of factors including genetic predisposition, hormonal changes, disease, sedentary lifestyle, and poor nutrition with advancing age [6, 17, 21] and seem to be exacerbated in conditions such as insulin resistance [4, 45].
Resistance exercise is a well-established way to defend against reductions in muscle size and strength [22, 28]. However, 25–40% of older persons do not gain muscle mass or strength following resistance exercise training (“poor-responders”) [3]. Additionally, there are age-associated increases in the variability of mass and strength changes in response to resistance exercise training [7, 10]. Following progressive resistance training (PRT) in older adults, our group has observed muscle size changes ranging from losses of approximately 10% to gains of over 20% in thigh muscle cross-sectional area [54]. Similarly, strength change can range from losses of approximately 40% to extreme gains over 150% following long-term resistance training [13]. Factors that drive response heterogeneity have also been observed with aging. These factors may include altered myogenic gene expression, decreased satellite cell numbers and function, and a lower propensity for muscle protein synthesis in response to an anabolic stimulus, now termed anabolic resistance [3, 12, 17, 36, 38, 43, 49]. However, it is unknown whether muscle lipid is associated with heterogeneity in strength or mass responses in healthy older people performing PRT.
Increased ectopic lipid is a consequence of aging and can have profound negative consequences. Lipid-rich muscle tissue leads to decreased muscle density, which can be reliably assessed by computed tomography (CT), and can be caused by increased lipid infiltration within muscle fibers (intramyocellular lipid, IMCL) and/or adipocytes between muscle fibers (extramyocellular lipid, EMCL) [27]. In fact, Goodpaster et al. were the first to show the sensitivity of CT attenuation values with changes in lipid content within skeletal muscle fibers confirmed with lipid-emulsion phantoms, biochemical extraction of triglyceride, and oil-red-o neutral lipid staining [18]. In addition, excess adipose tissue between muscles (intermuscular adipose tissue, IMAT) is associated with aging. Infiltration of lipid within and between muscles has primarily been analyzed in cross-sectional studies showing correlations with impaired physical function, including strength, balance, and gait speed across various populations [8, 14, 24, 47]. Insights from limited prospective studies have related disability costs of lower density muscle and IMAT to higher risk of hospitalizations, loss of mobility, and reduced quality of life [8, 52, 53]. In obese mice, intramuscular diacylglycerols and ceramides increase as a function of IMCL content and contribute to anabolic resistance [42]. Likewise, adipocytes have been shown to secrete inflammatory mediators such as cytokines that are thought to alter metabolic processes and the muscle architecture, which may ultimately compromise physical function and performance [1, 2, 46]. Thus, increased IMCL/EMCL and IMAT may contribute to poor responses to resistance exercise training in older adults via direct effects on muscle fibers [34]. Taken together and given the pathophysiological relevance of lipid accumulation and its unknown physiological role in exercise adaptation, one or all of these depots may be potential targets to restore hypertrophic potential among older individuals most susceptible to sarcopenia and functional decline.
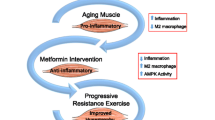
Metformin is the first-choice treatment for lowering glucose in type II diabetes and is the focus of many repurposing investigations. Metformin acts by inhibiting glucose output from the liver, but it also has anti-inflammatory effects in other tissues [44]. Thus, our group hypothesized that metformin would augment muscle growth and strength gain following PRT in a healthy aged cohort (the MASTERS trial) [29]. Contrary to our original hypothesis, metformin blunted the muscle hypertrophic response, did not reverse the poor-responder phenotype, and trended towards inhibiting strength gains. Interestingly, metformin also blunted average muscle density gain following PRT. In addition, transition from slow-twitch, IMCL-rich, type I muscle fibers to fast-twitch, low IMCL, type II fibers normally seen following PRT was blunted by metformin [54].
In this secondary analysis, we used data obtained in the MASTERS trial to assess the independent associations of baseline muscle density and IMAT to pre-training muscle mass, strength, and physical function in this healthy older population. Furthermore, we tested the hypothesis that low muscle density and high IMAT would be associated with reduced muscle mass and strength gains following PRT. Finally, we determined whether changes in IMAT and muscle density would be associated with resistance training outcomes, and whether any of these relationships would be altered by metformin.
Materials and methods
Participants
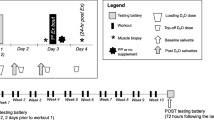
Data were originally acquired from the MASTERS trial (NCT02308228), which was conducted to determine whether metformin could augment hypertrophy in response to 14 weeks of PRT in healthy older adults. Full details of the study protocol, including participant inclusion/exclusion criteria and the overall study/PRT protocol (designed to optimize muscle mass and strength gains), can be found in our previous publications [29, 54]. The trial was conducted at the University of Kentucky and the University of Alabama at Birmingham, and all participants signed an informed consent approved by university IRBs prior to study enrollment. Ninety-four participants, aged ≥ 65, completed the exercise intervention and were included in this analysis. A graphical overview of the study design is shown in Fig. 1. Participants were randomized to receive either placebo or metformin (1700 mg/day) for the duration of the trial. Prior to beginning placebo or metformin, we assessed physical function and performed DXA, thigh CT scans, and muscle biopsies. Next, participants underwent a 2-week drug wash-in period, followed by a 2-week PRT familiarization period, and then baseline strength testing. All participants then continued to perform 12 more weeks of supervised, variable intensity, bilateral, upper, and lower body PRT. DXA, thigh CT scans, muscle biopsies, and physical function and strength testing were repeated 3 days after the final bout of PRT.

Overall study design
Computed tomography imaging
IMAT and thigh muscle density (TMD) were obtained from CT images prior to and following PRT using the methods described previously [30]. Cross-sections of both thighs were obtained at the mid-thigh level (the midpoint between the inguinal crease and superior border of the patella). Participants lay supine, with feet wrapped to avoid movement, wearing non-constrictive clothing to reduce compression of soft tissue. In addition, legs were separated for ease of analysis. NIH ImageJ was used to quantify tissue cross-sectional area and density (attenuation in Hounsfield units, HU). Bone (≥ 151 HU) and bone marrow were excluded from analyses. Skeletal muscle was defined as tissue with a density of − 29 to 150 HU, and adipose tissue was defined as tissue with a density of − 190 to − 30 HU. TMD in HU was used as an indicator of IMCL and EMCL. IMAT was defined as the adipose tissue inside the muscle fascia between the muscles. An individual response from our PRT program is depicted in Fig. 2. For this analysis, all CT variables were calculated from the dominant thigh, defined as the thigh with the higher muscle mass measured by DXA.

Cross-sectional mid-thigh region depicting intermuscular adipose tissue (IMAT), subcutaneous fat, skeletal muscle area (gray), and thigh muscle density (TMD). The black border (tracing of the muscle fascia) distinguishes between subcutaneous fat outside the muscle fascia and IMAT located inside the muscle fascia (both shown in white). Pixels located within the bone marrow were subtracted out for a more accurate analysis of IMAT and muscle. Muscle density was calculated as the mean attenuation of the muscle defined as a Hounsfield unit (HU) range of − 29 to 150
Muscle mass and body composition
Dual-energy x-ray absorptiometry (DXA) was performed to assess thigh muscle mass and body composition (whole body, regional fat, and lean mass) using a Lunar Prodigy (UAB) and an iDXA (UK), with standardized methods for regional partitioning. Custom analyses were performed to determine femur length and thigh muscle and fat mass. Femur length was measured from the center of the junction of the femoral head (at the femoral neck) and the acetabulum to the center of the bottom of the medial condyle. Thigh muscle and fat mass were calculated by subtracting the lower leg from total leg mass by creating a custom region of interest through the center of the knee joint between the tibial plateau and the femoral condyles and encapsulating the lower leg past the toes. Thigh muscle mass was then normalized to femur length. DXA scanner soft tissue percent coefficient of variation with repeat scans was < 1.0% at both sites.
Muscle fiber area
Muscle biopsies were obtained at baseline and after PRT, from the vastus lateralis after administration of local anesthetic (1% lidocaine premixed with bicarbonate) using a 5-mm Bergstrom needle with suction. Fiber type-specific cross-sectional area (CSA) was quantified on 7 μm cryosections, using an antibody recognizing laminin to delineate individual myofibers, followed by incubation with antibodies against myosin heavy chain isoform types I and IIa/IIax. Digital images were captured of the entire cross-section (between 250 and 1200 fibers), and mean myofiber CSA by fiber type was quantified using our validated automated algorithm [55]. CSA was not determined for samples consisting of less than 250 fibers.
Laboratory assessment of performance and self-reported physical function
Participants completed the short physical performance battery (SPPB) in the University of Kentucky Human Performance Laboratory [19]. This battery of tests includes three timed standing balance tests (side-by-side, semi-tandem, and tandem), a 3- or 4-m habitual gait speed test performed twice, and a timed repeated chair sit to stand (five times). Performance for each set of tasks was scored (0–4), with a summary score of 0–12. Individual tasks can also be used for the time to completion. Voluntary, dynamic strength for leg extension exercises was evaluated by testing a one repetition max (1RM), defined as the maximal load that a participant can lift one time with proper form through a full range of motion, via our well-established methods that have been standardized across sites [37]. We also evaluated maximal voluntary isometric contraction (MVIC) knee extension strength with a knee angle of 60° using our established methods on a Biodex System 4 Pro dynamometer at both study sites [48]. To determine knee extension concentric power, participants completed one set of 5 reps with the Biodex set on isotonic mode to allow for variable velocities with a constant external load equal to 40% of MVIC. Peak power was recorded as the highest power recorded during the 5 repetitions over approximately 70–80° range of motion. For an index of muscle quality, strength was normalized to either unilateral or bilateral thigh muscle mass. Baseline strength measures were obtained after 2 weeks of PRT to account for neurological adaptation. For self-reported physical function, we used the Short Form 36 (SF-36) physical function norm-based score and the Patient-Reported Outcomes Measurement Information System (PROMIS) physical function T-score as indicators of independent daily physical function [9, 33].
Statistical analysis
Ninety-four participants completed the original exercise intervention, with 48 randomized to placebo and 46 randomized to metformin. Baseline regression analyses included all participants as a single group. PRT outcome analyses were separated by group because metformin has been shown to blunt hypertrophy, affect strength gains, and impact muscle densities [54]. Data are described as the mean ± standard deviation (SD) and the median for continuous variables. Pearson’s correlation coefficients were used for all univariate associations. Multiple linear regression models were used to estimate the independent effects of IMAT and TMD on changes in muscle mass and physical performance. When appropriate based on the dependent variable, model covariates may have included age, sex, total body percent fat from DXA, and CT thigh muscle area as physiological determinants of muscle mass and growth. Standardized β coefficients and 95% confidence intervals are shown. Thigh muscle area, fiber type frequency, and our indexes of thigh muscle lipid were compared across three groups of SPPB functional scores using one-way analysis of variance (ANOVA) with Fisher’s least significant difference post hoc test. Functional groups were chosen to keep similar numbers of individuals in each group as our cohort was healthy and high functioning overall, making our distribution skewed to the right. Student’s t tests were used to compare dependent variables between high and low IMAT and TMD groups. Statistical analyses were performed using SAS 9.4 software with statistical significance set at p < 0.05.
Gene expression
RNA sequencing (RNA-Seq) was performed on pre- and post-PRT vastus lateralis biopsies from a subset of participants. Approximately 35 mg of muscle biopsy was homogenized in Qiazol (Qiagen, Valencia, CA) and total RNA was isolated using miRNeasy Mini Kits (Qiagen). RNA content and integrity were determined with a 2100 Bioanalyzer (Agilent, Santa Clara, CA). A minimum RNA Integrity Number (RIN) of 6.5 was set for all samples. Library preparation and sequencing were performed at Novogene Corporation (Chula Vista, CA) on an Illumina HiSeq 4000 system, using paired-end 150 bp (PE150) dual indexing. Quality control, trimming, and alignment of raw sequence reads and normalization were performed as previously described [26]. Raw count data are available at https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE157585. Statistical tests were performed using JMP Genomics v. 10 statistical software (SAS Institute). First, we compared baseline gene expression in participants with high versus low TMD (N = 19/group). Next, we identified genes whose fold changes were correlated to changes in TMD in the placebo group following PRT (N = 24) using Pearson’s product-moment correlations. The Benjamini-Hochberg adjustment for multiple comparisons was applied to both RNA-Seq analyses.
Results
Cohort characteristics
Baseline cohort characteristics are given in Table 1. This cohort represents a moderately active, high functioning group of generally healthy older individuals. Forty-three out of 48 individuals scored high (10–12) on the SPPB in the placebo group and 44 out of 46 individuals scored high in the metformin group. Ninety-eight percent of participants were Caucasian and 54% were female. Fourteen participants (15%) were obese with a BMI ≥ 30 kg/m−2. Only three individuals were classified as sarcopenic according to Studenski et al., calculated as the total mineral-free lean mass of the arms and legs adjusted for BMI [50]. We have previously reported that neither total calorie intake nor total protein intake was affected by PRT or PRT with metformin [54]. Likewise, daily physical activity measured by step counts was not affected by PRT or PRT with metformin.
Associations between baseline muscle lipid and body composition, muscle area, and fiber frequency
Baseline IMAT and TMD were significantly negatively associated (r = − 0.63, p < 0.001), providing internal validation to our analyses (Figure s1). Thigh IMAT was positively associated with total body percent fat (r = 0.44, p < 0.01) but did not associate with thigh muscle mass normalized to femur length, muscle area from CT, fiber CSA, or fiber type frequency. On the other hand, TMD was inversely associated with total body percent fat (r = − 0.48, p < 0.01) and positively associated with thigh muscle area (r = 0.23, p < 0.05). These analyses can be seen in Table s1.
Associations between baseline muscle lipid and physical function and performance
Table 2 shows univariate and adjusted multivariate linear regressions between baseline IMAT and TMD and baseline physical function and performance variables. In univariate regressions, IMAT was inversely associated with the SF-36 physical function score (r = − 0.21, p < 0.05), but it was not correlated to any other variable. In the adjusted multivariate model, IMAT was not significantly associated with any physical function or performance outcome, although there were trends towards inverse associations with SF-36 (p = 0.07) and repeated chair stand (p = 0.054). In univariate regressions, TMD was significantly associated with all physical function and performance variables. After adjusting for covariates, TMD remained significantly associated with all variables except gait speed and leg extension.
Relationship between SPPB group, muscle size, fiber frequency, and muscle lipid
To assess associations between muscle size, lipid content, and physical performance, we compared muscle features across three groups of composite SPPB functional scores: two high functioning groups scoring a 12 or 11, and one moderate function group scoring ≤ 10. Skeletal muscle size measures including skeletal muscle mass normalized to femur length and type II fiber CSA did not differ among SPPB groups, but skeletal muscle area (p = 0.07) (Fig. 3a) trended higher in those with higher SPPB scores. For muscle lipid, IMAT area did not differ among SPPB groups (p = 0.20) (Fig. 3b), but TMD was significantly lower in the SPPB ≤ 10 group when compared to both high functioning groups (whole model ANOVA p < 0.01; 11 vs. ≤ 10 post hoc p = 0.007; 12 vs. ≤ 10 post hoc p = 0.001) (Fig. 3c). This relationship was primarily driven by the repeated chair stand component of the SPPB score, since TMD was inversely correlated with chair stand times in both the univariate and adjusted multivariate models (Table 2).

Baseline relationships between SPPB score and muscle phenotype. a Muscle area and b IMAT do not differ between SPPB groups but c thigh muscle density (TMD) is significantly lower in participants with SPPB scores of 7–10; groups not connected by the same symbol are significantly different. N = 48, 23, and 22 for SPPB groups of 12, 11, or 7–10 respectively for CT variables muscle area, IMAT, and TMD
Mean levels of physical function and performance by low versus high TMD
TMD was strongly associated with multiple indicators of physical function and performance, while IMAT was not. We therefore proceeded to compare participants with low TMD (< 43.0 HU) versus high TMD (≥ 43.0 HU), representing those with high or low intramuscular lipid (Table 3). The high TMD group had higher levels of physical function and performance (p < 0.05) for all outcomes except gait speed (p = 0.16) and power (p = 0.09).
Associations between baseline muscle lipid and PRT outcomes in the placebo group
Table s2 shows adjusted multivariate linear regressions between baseline IMAT and TMD and PRT outcomes with placebo. After adjusting for covariates, neither baseline IMAT nor TMD was significantly associated with percent changes in muscle size following PRT. However, baseline IMAT was significantly inversely associated with percent change in PROMIS physical function score (β = − 0.42, p < 0.05) while baseline TMD was significantly inversely associated with percent change in power (β = − 0.38, p < 0.01) following PRT. In addition, baseline TMD trended towards inverse associations with percent change in SF-36 physical function score (β = − 0.29, p = 0.08). Thus, participants with lower baseline TMD tended to derive more functional benefit from PRT.
In participants with higher baseline muscle lipid (low TMD or high IMAT), metformin reduces strength gains in response to PRT
Since lower baseline TMD was associated with greater functional gains in response to PRT, we next determined how baseline TMD affected percent change in TMD following PRT, and whether metformin affected this response. In the placebo group, baseline TMD was inversely correlated to percent change in TMD (r = − 0.49, p < 0.001) (Fig. 4a). However, there was no relationship between baseline TMD and percent change in TMD in the metformin group (Fig. 4b). Since participants with low baseline TMD tended to have larger functional gains in response to PRT, we next determined how metformin affected strength gains in participants with low baseline TMD (< 43.0). While metformin did not affect percent change in power in those with low baseline TMD (Fig. 5a), there was a trend towards a decreased percent change in leg extension in the metformin group (p = 0.053) (Fig. 5b). Furthermore, percent change in MVIC was significantly decreased (p < 0.01) in metformin participants with low baseline TMD (Fig. 5c). Additionally, metformin treatment blunted mean IMAT loss, although the difference was not significant between the groups (Figure s2). We next determined whether metformin affected strength gains in participants with high IMAT at baseline. In participants with high baseline IMAT, metformin did not affect percent change in power. However, metformin participants with high baseline IMAT trended towards reduced gains in leg extension (p = 0.08), and they had significantly reduced gains in MVIC (p < 0.01) (Figure s3a–c).

Baseline thigh muscle density (TMD) is significantly related to changes in thigh muscle density following training in placebo participants (a) but this relationship is blunted with the use of metformin (b)

Metformin preferentially inhibits performance gains in those with low baseline thigh muscle density (TMD) for (b, c) dynamic and isometric strength but not a power with metformin participants seeing less benefit across both low and high baseline densities. **p < 0.01
Associations between changes in muscle lipid and changes in muscle size and function following PRT
We previously reported reduced gains in TMD in participants taking metformin during PRT [54]. Here, we report that mean IMAT loss is also blunted, with placebo participants losing an average of 9.1% ± 12.8 IMAT area vs. 5.7% ± 13.0 in metformin participants. We next used adjusted multivariate linear regressions to determine whether changes in TMD were associated with PRT outcomes in the placebo group (Table 4). Percent change in TMD was not associated with gains in muscle size in the placebo group. However, percent change in TMD was correlated to increased type II fiber frequency (β = 0.40, p < 0.05). TMD gains were also significantly associated with gains in SF-36 physical function score (β = 0.31, p < 0.05) and improved chair stand performance (β = − 0.32, p < 0.05). Nonetheless, percent change in TMD was not associated with percent change in power, leg extension, or MVIC in placebo participants. Percent change in IMAT was not associated with any of the training outcomes in the placebo group (data not shown).
In participants with higher gains in TMD, metformin reduces strength gains in response to PRT
Lower baseline TMD was associated with greater percent change in TMD following PRT, and this relationship was altered by metformin. We therefore determined whether metformin inhibited gains in muscle size and physical function in participants who had the largest gains in muscle density. In participants with the highest TMD gains following PRT, mean percent change in power did not differ between drug groups (Fig. 6a). However, percent change in leg extension (p = 0.09) trended towards a reduction, while percent change in MVIC was significantly reduced (p < 0.05) in participants who were randomized to metformin (Fig. 6b, c). Compared to type I fibers, type II fibers tend to be denser (containing less IMCL) and adaptations in type II fibers drive strength improvements following PRT. Both groups displayed a similar pattern of positive association between percent change in TMD and percent change in type II fiber frequency, with a non-significant trend in the placebo group (p = 0.07) (Fig. 6a), and a significant correlation in the metformin group (p < 0.05) (Fig. 6b). Nonetheless, metformin blunted the typical response of type I fibers switching to type II fibers, as we have previously reported [54]. This reduction in fiber type switching with metformin likely contributed to reduced strength gains (Fig. 7).

Metformin blunts performance in those who had the highest thigh muscle density (TMD) gains following training. *p < 0.05

Percent change in thigh muscle density (TMD) had similar positive associations with percent change in type II fiber frequency in both a placebo and b metformin individuals, although fiber type switching was blunted with metformin use
We did not observe statistically significant relationships between gene expression and TMD
We performed RNA-Seq of muscle biopsies to identify genes that are associated with TMD, and to identify gene expression changes that are correlated to change in TMD in response to PRT. When comparing participants with high versus low TMD at baseline (N = 19/group), only 86 genes differed with unadjusted p < 0.01, and no genes were significantly different following the Benjamini-Hochberg adjustment (Table s3). When gene fold changes were correlated to percent change in TMD following PRT within the placebo group (N = 24), only 190 gene fold changes were correlated with unadjusted p < 0.01, and none was significantly correlated following the Benjamini-Hochberg adjustment (Table s4).
Discussion
In older adults, resistance exercise training is highly beneficial to long-term health. However, to our knowledge, the relationship between muscle lipid content and the response to PRT has not been investigated in detail. In this secondary analysis of the MASTERS trial, we hypothesized that muscle lipid content would be associated with an anabolic resistant state and may be a predictor of the poor-responder phenotype. However, we did not find any association between baseline IMAT or TMD and changes in muscle mass following PRT. Rather, we report that baseline TMD, a proxy of IMCL and EMCL, impairs strength and performance gains in response to PRT in older individuals. In addition, we show that metformin preferentially inhibits PRT responses in those participants with higher baseline lipid profiles (lower TMD or higher IMAT).
Progressively higher IMAT and lower TMD have been observed in older individuals, regardless of changes in weight [15, 16]. Furthermore, higher IMAT area [20, 47] and lower muscle density [2, 14, 16, 24, 32] have both been linked to decrements in self-reported physical function and laboratory-measured performance in various populations. Similar to the result of Khoja et al. studying a rheumatoid arthritis population, we found that TMD, and not IMAT, is independently associated with many of the strength, function, and performance measures we assessed, after adjusting for confounding variables known to affect muscle size and strength [24]. Muscle density was highly related to muscle power and repeated chair stand, which contains elements of both strength and power. This is particularly important since muscle power has been found to be vital to perform tasks of daily living in older individuals [41]. We also show that baseline SPPB score is associated with baseline TMD, with decrements of just 1 SPPB performance point being associated with significantly lower TMD. Furthermore, baseline TMD was a better predictor of poor physical performance than muscle mass or muscle area. However, IMAT did not differ between SPPB groups. These data suggest that muscle density, but not IMAT, may be just as important as muscle size for the performance of physically demanding activities such as PRT.
Mechanisms have been suggested which may explain the connection between increased muscle fat infiltration and performance. Larger quantities of chronic adipose tissue infiltration result in greater inflammation and architectural changes within the muscle micro-environment that produce adverse effects on muscle function [23]. Rahemi et al. used computational models to suggest that intracellular lipid inclusions cause loss in overall force and force per unit size due to mechanical changes in the connective tissue [39]. Changes in pennation angle have also been reported in older women with greater levels of obesity and IMCL [40]. Pro-inflammatory mediators in the muscle can also lead to increased oxidative stress, changes in metabolism due to mitochondrial inefficiency, and reduced production of myofilament proteins, ultimately leading to reduced contractile capacity [11]. However, we found no significant differences in muscle gene expression in participants with high versus low baseline TMD, leading us to hypothesize that muscle lipid storage and metabolism are primarily regulated by lipid and protein signaling. We also found no significant correlations between changes in gene expression and percent change in TMD in the placebo group following PRT, possibly supporting our hypothesis that TMD is largely regulated at the signaling level, or possibly indicating that we missed a crucial time point for gene expression changes that could alter TMD.
Resistance training can have a profound effect on muscle density without changes in muscle volume [51]. This is consistent with our observation that participants with low baseline TMD had the largest strength gains following PRT. However, neither baseline IMAT nor TMD was associated with a hypertrophic response. These data indicate that lipid metabolism may contribute to performance gains with PRT in older individuals with low baseline TMD. Conversely, those with higher baseline muscle density may derive less performance benefit from PRT.
Our primary outcomes report from the MASTERS trial showed that, compared to placebo, metformin blunts hypertrophy, strength, and normal muscle density gains following PRT [54]. Here, we show that metformin preferentially blunted gains in TMD and strength in participants with high baseline muscle lipid (low TMD or high IMAT). Similarly, metformin individuals who had the highest changes in TMD also had blunted strength gains. This suggests that adaptations in lipid metabolism are required to maximize strength gains in response to PRT. These results are consistent with findings from other studies showing that metformin blunted responses to aerobic exercise training [25], resistance training [5], and concurrent aerobic/resistance training [31].
Limitations
This study is not without limitations. Our study population consisted of moderately active, healthy older men and women who engaged in PRT along with metformin or placebo, but it did not include a no-PRT metformin only group. Thus, we are unable to comment on whether metformin alone might affect muscle lipid, size, or strength. Importantly, we did not study the interaction between metformin and PRT in people with pre-diabetes or diabetes, and we cannot comment on whether the combination of metformin and PRT would be appropriate in this population. While we attempted to identify increased infiltration of lipid into the muscle as a possible determinant of anabolic heterogeneity, there are many known other candidates contributing to muscle mass and size loss due to aging such as physical activity, nutrition, and genetic background, which we have not characterized. Additionally, while TMD is a surrogate measure for IMCL and EMCL, it does not elucidate cellular lipid localization, nor does it quantify levels of specific lipid species.
Conclusion
In summary, we report that TMD, as a proxy for IMCL and EMCL, is independently associated with cross-sectional physical performance measures, as well as longitudinal strength and power changes following PRT in a large cohort of healthy older persons. We demonstrate that muscle density is positively associated with cross-sectional physical performance, but it does not predict one’s ability to gain lean mass during PRT. Furthermore, metformin blunts PRT outcomes preferentially in older individuals with lower TMD (higher baseline muscle lipid). Given metformin’s proven safety and efficacy for treating pre-diabetes and diabetes, future work should elucidate the molecular mechanisms responsible for its interactions with exercise. Although future work is also needed to identify molecular mechanisms that drive the relationships between muscle lipid content and resistance exercise adaptation, our results indicate that healthy older adults with higher baseline muscle lipid derive more benefit from PRT than those who have lower baseline muscle lipid. Our results further indicate that muscle density, independent from muscle mass, is strongly associated with function, performance, and strength in older individuals. The work presented here provides opportunities to determine whether modulation of IMAT or TMD could provide an approach to counteract PRT response heterogeneity and restore hypertrophic potential among older individuals.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author and, on reasonable request, a minimal de-identified dataset will be provided.
Abbreviations
- PRT:
-
progressive resistance training
- CT:
-
computed tomography
- IMAT:
-
intermuscular adipose tissue
- TMD:
-
thigh muscle density
- IMCL:
-
intramyocellular lipid
- EMCL:
-
extramyocellular lipid
- DXA:
-
dual-energy x-ray absorptiometry
- HU:
-
Hounsfield unit
- SPPB:
-
short physical performance battery
- SF-36:
-
Short Form 36
- PROMIS:
-
patient-reported outcomes measurement information system
- 1RM:
-
one repetition max
- CSA:
-
cross-sectional area
References
Addison OR (2013) Fit or fat: the relationship of inflammation, intramuscular fat and muscle, and mobility function in older adults. ProQuest Information & Learning.
Addison O, Wende AR, McClain DA, Marcus RL, Drummond MJ, Lastayo PC, Dibble LE (2014) Intramuscular fat and inflammation differ in older adults: the impact of frailty and inactivity [electronic resource] journal of nutrition, health & aging 18:532-538 https://doi.org/10.1007/s12603-014-0019-1.
Bamman MM, Petrella JK, Jeong-su K, Mayhew DL, Cross JM. Cluster analysis tests the importance of myogenic gene expression during myofiber hypertrophy in humans. Journal of Applied Physiology. 2007;102:2232–9.
Barzilay JI, Cotsonis GA, Walston J, Schwartz AV, Satterfield S, Miljkovic I, Harris TB (2009) Insulin resistance is associated with decreased quadriceps muscle strength in nondiabetic adults aged ≥ 70 years Diabetes Care 32:736-738 doi:https://doi.org/10.2337/dc08-1781.
Boule NG, Kenny GP, Larose J, Khandwala F, Kuzik N, Sigal RJ. Does metformin modify the effect on glycaemic control of aerobic exercise, resistance exercise or both? Diabetologia. 2013;56:2378–82. https://doi.org/10.1007/s00125-013-3026-6.
Brook MS, Wilkinson DJ, Phillips BE, Perez-Schindler J, Philp A, Smith K, et al. Skeletal muscle homeostasis and plasticity in youth and ageing: impact of nutrition and exercise. Acta Physiologica. 2016;216:15–41. https://doi.org/10.1111/apha.12532.
Camera DM. Anabolic heterogeneity following resistance training: a role for circadian rhythm? Front Physiol. 2018;9:569. https://doi.org/10.3389/fphys.2018.00569.
Cawthon PM, Fox KM, Gandra SR, Delmonico MJ, Chiou CF, Anthony MS, et al. Do muscle mass, muscle density, strength, and physical function similarly influence risk of hospitalization in older adults? J Am Geriatr Soc. 2009;57:1411–9. https://doi.org/10.1111/j.1532-5415.2009.02366.x.
Cella D et al. (2019) PROMIS ® adult health profiles: efficient short-form measures of seven health domains value in health: the journal of the International Society for Pharmacoeconomics and Outcomes Research 22:537-544 https://doi.org/10.1016/j.jval.2019.02.004.
Chmelo EA, Crotts CI, Newman JC, Brinkley TE, Lyles MF, Leng X, et al. Heterogeneity of physical function responses to exercise training in older adults. Journal of the American Geriatrics Society. 2015;63:462–9. https://doi.org/10.1111/jgs.13322.
Choi SJ, et al. Intramyocellular lipid and impaired myofiber contraction in normal weight and obese older adults. Journals of Gerontology: Series A: Biological Sciences and Medical Sciences. 2016;71:557–64.
Churchward-Venne TA, Breen L, Phillips SM. Alterations in human muscle protein metabolism with aging: protein and exercise as countermeasures to offset sarcopenia. Biofactors (Oxford, England). 2014;40:199–205. https://doi.org/10.1002/biof.1138.
Churchward-Venne TA, Tieland M, Verdijk LB, Leenders M, Dirks ML, de Groot LCPGM, et al. There are no nonresponders to resistance-type exercise training in older men and women. Journal Of The American Medical Directors Association. 2015;16:400–11. https://doi.org/10.1016/j.jamda.2015.01.071.
Cleary LC, Crofford LJ, Long D, Charnigo R, Clasey J, Beaman F, et al. Does computed tomography-based muscle density predict muscle function and health-related quality of life in patients with idiopathic inflammatory myopathies? Arthritis Care & Research. 2015;67:1031–40. https://doi.org/10.1002/acr.22557.
Delmonico MJ, et al. Longitudinal study of muscle strength, quality, and adipose tissue infiltration. American Journal of Clinical Nutrition AJN. 2009;90:1579–85.
Frank-Wilson AW, et al. Associations of quadriceps torque properties with muscle size, attenuation, and intramuscular adipose tissue in older adults. Journals of Gerontology: Series A: Biological Sciences and Medical Sciences. 2018;73:931–8.
Garatachea N, Lucía A. Genes and the ageing muscle: a review on genetic association studies. Age. 2013;35:207–33. https://doi.org/10.1007/s11357-011-9327-0.
Goodpaster BH, Kelley DE, Thaete FL, He J, Ross R. Skeletal muscle attenuation determined by computed tomography is associated with skeletal muscle lipid content. Journal of Applied Physiology (Bethesda, Md: 1985). 2000;89:104–10.
Guralnik JM. Lower extremity function in persons over the age of 70 years as a predictor of subsequent disability. N Engl J Med. 1995;332:556–61.
Hilton TN, Tuttle LJ, Bohnert KL, Mueller MJ, Sinacore DR. Excessive adipose tissue infiltration in skeletal muscle in individuals with obesity, diabetes mellitus, and peripheral neuropathy: association with performance and function. Physical Therapy. 2008;88:1336–44. https://doi.org/10.2522/ptj.20080079.
Horstman AM, Dillon EL, Urban RJ, Sheffield-Moore M. The role of androgens and estrogens on healthy aging and longevity. Journals of Gerontology: Series A: Biological Sciences and Medical Sciences. 2012;67:1140–52.
Hunter GR, McCarthy JP, Bamman MM. Effects of resistance training on older adults. Sports Medicine. 2004;34:329–48.
Jensen GL. Inflammation: roles in aging and sarcopenia. JPEN Journal of Parenteral & Enteral Nutrition. 2008;32:656–9.
Khoja SS, Moore CG, Goodpaster BH, Delitto A, Piva SR. Skeletal muscle fat and its association with physical function in rheumatoid arthritis. Arthritis Care & Research. 2018;70:333–42. https://doi.org/10.1002/acr.23278.
Konopka AR et al. (2018) Metformin inhibits mitochondrial adaptations to aerobic exercise training in older adults Aging Cell:e12880 https://doi.org/10.1111/acel.12880.
Kulkarni AS et al. (2020) Metformin alters skeletal muscle transcriptome adaptations to resistance training in older adults aging accepted.
Larson-Meyer DE, Ravussin E, Newcomer BR, Kelley DE, Smith SR, Heilbronn LK. Muscle-associated triglyceride measured by computed tomography and magnetic resonance spectroscopy. Obesity research. 2006;14:73–87.
Law TD, Clark LA, Clark BC. Resistance exercise to prevent and manage sarcopenia and dynapenia. Annual review of gerontology & geriatrics. 2016;36:205–28.
Long DE, et al. Metformin to Augment Strength Training Effective Response in Seniors (MASTERS): study protocol for a randomized controlled trial. Trials. 2017;18:1–14. https://doi.org/10.1186/s13063-017-1932-5.
Long DE, Villasante Tezanos AG, Wise JN, Kern PA, Bamman MM, Peterson CA, et al. A guide for using NIH Image J for single slice cross-sectional area and composition analysis of the thigh from computed tomography. PLoS ONE. 2019;14:1–11. https://doi.org/10.1371/journal.pone.0211629.
Malin SK, Gerber R, Chipkin SR, Braun B. Independent and combined effects of exercise training and metformin on insulin sensitivity in individuals with prediabetes. Diabetes Care. 2012;35:131–6. https://doi.org/10.2337/dc11-0925.
Marcus RL, Addison O, Dibble LE, Bo Foreman K, Morrell G, LaStayo P (2012) Intramuscular adipose tissue, sarcopenia, and mobility function in older individuals Journal of Aging Research:1-6 https://doi.org/10.1155/2012/629637.
McHorney CA, Ware JE Jr, Raczek AE. The MOS 36-item short-form health survey (SF-36): II. Psychometric and clinical tests of validity in measuring physical and mental health constructs. Medical care. 1993;31:247–63.
Miljkovic N, Lim J-Y, Miljkovic I, Frontera WR. Aging of skeletal muscle fibers. Annals Of Rehabilitation Medicine. 2015;39:155–62. https://doi.org/10.5535/arm.2015.39.2.155.
Mitchell WK, Williams J, Atherton P, Larvin M, Lund J, Narici M. Sarcopenia, dynapenia, and the impact of advancing age on human skeletal muscle size and strength; a quantitative review. Frontiers In Physiology. 2012;3:260. https://doi.org/10.3389/fphys.2012.00260.
Moretti I et al. (2016) MRF4 negatively regulates adult skeletal muscle growth by repressing MEF2 activity Nature Communications 7:12397-12397 https://doi.org/10.1038/ncomms12397.
Petrella JK, Jeong-su K, Tuggle SC, Hall SR, Bamman MM (2005) Age differences in knee extension power, contractile velocity, and fatigability Journal of Applied Physiology 98:211-220.
Petrella JK, Kim J-S, Cross JM, Kosek DJ, Bamman MM. Efficacy of myonuclear addition may explain differential myofiber growth among resistance-trained young and older men and women. American Journal Of Physiology Endocrinology And Metabolism. 2006;291:E937–46.
Rahemi H, Nigam N, Wakeling JM (2015) The effect of intramuscular fat on skeletal muscle mechanics: implications for the elderly and obese Journal Of The Royal Society, Interface 12:20150365–20150365 https://doi.org/10.1098/rsif.2015.0365.
Rastelli F et al. (2015) Effects of muscle composition and architecture on specific strength in obese older women Experimental Physiology 100:1159-1167 https://doi.org/10.1113/EP085273.
Reid KF, Fielding RA. Skeletal muscle power: a critical determinant of physical functioning in older adults. Exercise & Sport Sciences Reviews. 2012;40:4–12.
Rivas DA, McDonald DJ, Rice NP, Haran PH, Dolnikowski GG, Fielding RA (2016) Diminished anabolic signaling response to insulin induced by intramuscular lipid accumulation is associated with inflammation in aging but not obesity American journal of physiology regulatory, integrative and comparative physiology 310:R561-R569 https://doi.org/10.1152/ajpregu.00198.2015.
Robinson MM, Dasari S, Konopka AR, Johnson ML, Manjunatha S, Esponda RR, et al. Enhanced protein translation underlies improved metabolic and physical adaptations to different exercise training modes in young and old humans. Cell Metabolism. 2017;25:581–92. https://doi.org/10.1016/j.cmet.2017.02.009.
Saisho Y (2015) Metformin and inflammation: its potential beyond glucose-lowering effect endocrine, metabolic & immune disorders drug targets 15:196-205.
Sayiner ZA, Öztürk ZA (2019) Relationship between sarcopenia and type 2 diabetes mellitus in elderly patients Yaşlı Hastalarda Sarkopeni ve Tip 2 Diabetes Mellitus Arasındaki İlişki 23:47-52 https://doi.org/10.25179/tjem.2018-62624.
Schering L et al. (2015) Identification of novel putative adipomyokines by a cross-species annotation of secretomes and expression profiles Archives of Physiology & Biochemistry 121:194-205 https://doi.org/10.3109/13813455.2015.1092044.
Scott D et al. (2015) Associations of calf inter- and intra-muscular adipose tissue with cardiometabolic health and physical function in community-dwelling older adults journal of Musculoskeletal & Neuronal Interactions 15:350-357.
Srikuea R et al. (2013) Association of fibromyalgia with altered skeletal muscle characteristics which may contribute to postexertional fatigue in postmenopausal women Arthritis And Rheumatism 65:519-528 https://doi.org/10.1002/art.37763.
Stec MJ, Kelly NA, Many GM, Windham ST, Tuggle SC, Bamman MM. Ribosome biogenesis may augment resistance training-induced myofiber hypertrophy and is required for myotube growth in vitro. American Journal Of Physiology Endocrinology And Metabolism. 2016;310:E652–61. https://doi.org/10.1152/ajpendo.00486.2015.
Studenski SA, Peters KW, Alley DE, Cawthon PM, McLean RR, Harris TB, et al. The FNIH sarcopenia project: rationale, study description, conference recommendations, and final estimates. The Journals Of Gerontology Series A, Biological Sciences And Medical Sciences. 2014;69:547–58. https://doi.org/10.1093/gerona/glu010.
Taaffe DR, Henwood TR, Nalls MA, Walker DG, Lang TF, Harris TB. Alterations in muscle attenuation following detraining and retraining in resistance-trained older adults. Gerontology. 2009;55:217–23. https://doi.org/10.1159/000182084.
Trombetti A, Reid K, Pasha E, Phillips E, Fielding R, Hars M, Herrmann F (2016) Age-associated declines in muscle mass, strength, power, and physical performance: impact on fear of falling and quality of life Osteoporosis International 27:463-471 https://doi.org/10.1007/s00198-015-3236-5.
Visser M, et al. Muscle mass, muscle strength, and muscle fat infiltration as predictors of incident mobility limitations in well-functioning older persons. Journals of Gerontology Series A: Biological Sciences & Medical Sciences. 2005;60:324–33.
Walton RG et al. (2019) Metformin blunts muscle hypertrophy in response to progressive resistance exercise training in older adults: a randomized, double-blind, placebo-controlled, multicenter trial: the MASTERS trial Aging Cell:e13039 https://doi.org/10.1111/acel.13039.
Yuan W et al. (2018) MyoVision: software for automated high-content analysis of skeletal muscle immunohistochemistry Journal of Applied Physiology 124:40-51.
Acknowledgments
The authors would like to thank each of our valuable research participants for their time, effort, and dedication. We would like to thank Tara Bennett PA-C for performing muscle biopsies and Janna Jackson PhD and Cory Dungan PhD for performing immunohistochemistry. We would also like to thank Ameya Kulkarni, PhD and Nir Barzilai, MD, PhD of the Albert Einstein School of Medicine Nathan Shock Center for assistance with RNA sequencing.
Funding
The study was funded by the National Institutes of Health - National Institute on Aging grant AG046920 and supported by the NIH Clinical and Translational Science Awards (CTSA) (UL1TR001998) at the University of Kentucky and the NIH CTSA (UL1TR000165) at the University of Alabama at Birmingham. This study was also supported by an award from the Glenn Foundation for Medical Research.
Author information
Authors and Affiliations
Contributions
DEL, CAP, and RGW wrote the manuscript; DEL, PAK, MMB, CAP, and RGW contributed to the design of the study; DEL, SCT, STW, MMB, PAK, and CAP implemented the clinical protocol; DEL, BCP, and RGW performed analyses; AGV provided statistical analysis of the data. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethics approval
This study was approved by the University of Kentucky institutional review board (IRB 14-0330) and the University of Alabama at Birmingham institutional review board (IRB F140722001) prior to any participants enrolling. Data and safety monitoring was provided by the UK CCTS DSMB on a quarterly basis.
Consent to participate
All participation was on a voluntary basis, and each participant was required to sign an approved IRB consent form and HIPAA authorization prior to any procedures taking place.
Consent for publication
Not applicable.
Disclaimer
The NIH had no role in the design of the study, collection, analysis, or interpretation of data, or in writing the manuscript.
Code availability
Not applicable.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The first author, DEL, is a research coordinator in the University of Kentucky College of Health Sciences, and an exercise physiologist in the UK CCTS Functional Assessment and Body Composition Core Lab.
Supplementary information
Figure s1
Mid-thigh intermuscular adipose tissue (IMAT) is inversely correlated with thigh muscle density (TMD) at baseline HU, Hounsfield Unit (PDF 97 kb)
Figure s2
Metformin blunts loss in IMAT following resistance training (PDF 93 kb)
Figure s3
Metformin preferentially inhibits performance gains in those with high amounts of baseline thigh intermuscular adipose tissue (IMAT) for (b, c) dynamic and isometric strength but not a power. *p < 0.05 (PDF 119 kb)
Table s1
(DOCX 16 kb)
Table s2
(DOCX 18 kb)
Table s3
Baseline gene expression in high vs. low TMD. See electronic supplementary excel file- intended for publication as an online data supplement (XLSX 28 kb)
Table s4
Correlations between gene fold change and % change in TMD in the placebo group following PRT (XLSX 24.8 kb)
About this article

Cite this article
Long, D.E., Peck, B.D., Tuggle, S.C. et al. Associations of muscle lipid content with physical function and resistance training outcomes in older adults: altered responses with metformin. GeroScience 43, 629–644 (2021). https://doi.org/10.1007/s11357-020-00315-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11357-020-00315-9