
Abstract
In skeletal muscles, calorie restriction (CR) preserves muscle mass in middle-aged rats but not younger rats. The underlying mechanisms for this age-specific response are unknown. Skeletal muscle mass depends on several factors, with protein synthesis and degradation playing major roles. Therefore, the purpose of this study was to investigate whether CR affects younger and older animals differently on mTOR signaling and ubiquitin-proteasome pathway (UPP). Four-, 8-, and 16-month-old rats, with or without 40% CR for a duration of 14 weeks, were sacrificed after an overnight fasting. Total protein content and the phosphorylation level of AKT, mTOR, S6K, and 4EBP1 and protein content of key markers in the UPP (FOXO3a, atrogin, MuRF1, ubiquitinated proteins, proteasome subunits alpha 7 and beta 5) were determined. Unlike younger rats, CR decreased the content of phosphorylated mTOR, S6K, phosphorylated S6K, FOXO3a, and ubiquitinated proteins in middle-aged rats. In conclusion, CR-induced reduction of content/ phosphorylation levels of key proteins in mTOR signaling and the UPP occurred in the middle-aged rats but not younger rats. The age-dependent effects of CR on mTOR signaling and the UPP indirectly explained the age-related effects of CR on muscle mass of animals.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Over the past several years, calorie restriction (CR) investigations report many benefits such as improvements in cardiometabolic health, preservation of skeletal muscle mass, and an increase in years without disease or disability (Chen et al. 2015; Colman et al. 2008; Fontana et al. 2010; Mattison et al. 2017; McKiernan et al. 2012). The mechanisms responsible for the beneficial effects of CR include the suppression of mammalian target of rapamycin (mTOR) signaling (Fontana et al. 2010; Kaeberlein et al. 2005; Sharples et al. 2015; Testa et al. 2014) and the reduction of inflammation (Lopez-Lluch and Navas 2016; Mercken et al. 2013; Sharples et al. 2015; Yang et al. 2016). Interestingly, the magnitude of the CR benefit depends on the age at which CR is initiated. For example, compared to old rats, young rats respond to a greater extent in terms of increasing content and receptors of adiponectin following CR (Niemann et al. 2008). In adult monkeys, CR protected against immune cell senescence whereas it was ineffective in young and old monkeys (Messaoudi et al. 2008). In our previous publication, we showed that CR improved skeletal muscle energy metabolism and preserved muscle mass in middle-aged rats but not in young rats (Chen et al. 2015). At this time, the mechanism that explains the age-dependent effects of CR on skeletal muscle mass is unknown.
The determination of skeletal muscle mass depends on several factors, with the balance between protein synthesis and degradation playing the major role. It is well known that muscle mass decreases with aging. In the atrophied muscles of older animals, both mTOR signaling and ubiquitin-proteasome pathway (UPP) were found upregulated compared to younger animals (Clavel et al. 2006; Guillet et al. 2004; Ibebunjo et al. 2013; Kimball et al. 2004; Markofski et al. 2015). The concurrent enhancement of anabolic and catabolic signaling in aging muscles is likely a response to the chronic low-grade inflammatory environment present in older animals (Dalle et al. 2017). Studies have shown that mTOR, ribosomal protein S6 protein kinases 1 (S6K), and forkhead box class O (FOXO) (a transcriptional factor that upregulates UPP) are activated by inflammatory cytokines (Chung et al. 2011; Dalle et al. 2017; Ma and Blenis 2009; Zhang et al. 2008). In addition, it is well established that low-grade inflammation is present in aging skeletal muscles (Combaret et al. 2009; Marzetti et al. 2013; Mercken et al. 2017; Peake et al. 2010; Sakuma et al. 2017). Based on the finding that CR preserved muscle mass in middle-aged rats only (Chen et al. 2015) and knowing that CR is a powerful intervention to decrease inflammation, the current study aimed to identify the age-dependent response of mTOR signaling and UPP to CR in skeletal muscles.
Method
Animals and diet intervention
Eighty male Sprague-Dawley rats aged 4 months (young rats, 100% strain survival) (n = 20), 8 months (adult rats, 100% strain survival) (n = 20), and 16 months old (middle-aged rats, 90% strain survival) (n = 40) were purchased from the Laboratory Animal Center of Chang Gung University and were housed in the research animal facility at Chang Gung University. Rats in each age group were randomized into ad libitum (AL) and calorie restriction (CR) groups. Rats in the CR group received 40% CR for 14 weeks as described previously (Chen et al. 2015). After the 14 weeks of intervention, there were 20 young rats (8 months), 20 adult rats (12 months), and 27 middle-aged rats (20 months; 12 in the AL group and 15 in the CR group). Animals were anesthetized with an intraperitoneal injection of ketamine (37.5 mg/kg) and xylazine (12.5 mg/kg) after overnight fasting. The soleus muscle was taken and stored at − 80 °C for analysis.
In the current study, body weights and muscle weights were not reported. Using the same research design, we reported that soleus muscle mass decreased with aging by 16% between young (8 months) and middle-aged (20 months) rats (5). CR reduced body weight in both young and middle-aged rats by 32%. The soleus muscle weight was reduced by 28% in the young rats, and no change was observed in the middle-aged rats. In the current study, we continued to focus on the soleus muscle in order to fully understand how this muscle responds to CR and aging. The protocol of this study was approved by the Institutional Animal Care and Use Committees of Chang Gung University.
Western blot
Muscle tissue was homogenized in ice-cold buffer (20 mM HEPES, 2 mM EGTA, 50 mM NaF, 100 mM KCl, 0.2 mM EDTA, 50 mM glycerophosphate, 1 mM DTT, 0.1 mM PMSF, 1 mM benzamidine, and 0.5 mM Na3VO4, and phosphatase inhibitor cocktail) using a Teflon pestle. The homogenate was centrifuged at 14,000g for 10 min at 4 °C. Protein concentrations were determined using a bicinchoninic acid assay (Thermo Fisher, Rockford, IL). Proteins (35 μg for AKT, 20 μg for mTOR, alpha 7, and beta 5; 30 μg for S6K, initiation factor 4E binding proteins (4EBP1), atrogin and MuRF1; 30 μg for FOXO3a; 15 μg for ubiquitinated protein) were electrophoresed on 10% (AKT, S6K, FOXO3a, atrogin, MuRF1, and ubiquitinated protein) or 13% (4EBP1, alpha 7, and beta 5) or 4~20% gradient (mTOR) polyacrylamide gels (Bio-Rad Laboratories, Hercules, CA). After resolution, the proteins were transferred to polyvinylidene difluoride membranes at 90 V for 90 min at 4 °C (Bio-Rad Laboratories). Ponceau S staining was used to ensure equal loading and the quality of transfer.
Protein-bound membranes were blocked with 5% low-fat dried milk in Tris-buffered saline containing 0.01% Tween 20 (TBST) for 1 h at room temperature. Membranes were then washed with TBST and incubated with primary antibodies diluted in blocking solution with 5% bovine serum albumin (Sigma-Aldrich, St. Louis, MO, USA.) overnight at 4 °C. Primary antibodies against phospho-AKT (Thr308) (Cell Signaling Technologies, Boston, MA, #9275), mTOR (Cell Signaling Technologies, #2971), phospho-mTOR (Ser2448) (Cell Signaling Technologies, #2972), S6K (Cell Signaling Technologies, #9202), phospho-S6K (Thr389) (Cell Signaling Technologies, #9205), 4EBP1 (Cell Signaling Technologies, #9452), phospho-4EBP1 (Thr37/46) (Cell Signaling Technologies, #9459), FOXO3a (Cell Signaling Technologies, #2497), phospho-FOXO3a (Ser253) (Cell Signaling Technologies, #9466), and alpha 7 (Santa Cruz, sc-58417) were diluted 1:500. Primary antibodies against AKT (Cell Signaling Technologies; #9272), atrogin (Proteintech, Chicago, IL; 12866-1-AP), MuRF1 (Proteintech, Chicago, IL; 55456-1-AP), ubiquitinated protein (Santa Cruz, sc-53509), and beta 5 (Cell Signaling Technologies, #12919) were diluted 1:1000. After incubation with the primary antibodies, membranes were washed in TBST and incubated with anti-mouse/anti-rabbit HRP-conjugated secondary antibody (Sigma-Aldrich, St. Louis, MO) (1:5000 in 3.5% low-fat dried milk blocking buffer) at room temperature for 1 h. Immuno-bound antibodies were detected by electrochemiluminescence (Invitrogen, Carlsbad, CA) and imaged by a ChemiDoc imaging system (Bio-Rad, Hercules, CA). Quantity One (Bio-Rad) was used to quantify the optical density (OD) of each sample. The OD of the samples was normalized to that of a standard sample on the same membrane to compare samples across multiple membranes (Chen et al. 2010, 2015, 2017).
Statistics
SPSS version 24 (SPSS, Chicago, IL, USA) and GraphPad Prism 5.0 (GraphPad software Inc., La Jolla, CA, USA) were used to analyze data and draw the plots. Effects of age and CR on outcome variables were determined by two-way ANOVA. If there was significant interaction between age and CR, independent t test was used to examine the effects of CR in each age group. Data were reported as the mean ± SE. P < 0.05 was considered significant statistically.
Results
Regulators in mTOR signaling pathway
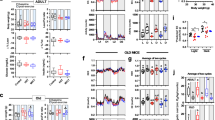
AKT is an up-stream activator of mTOR. AKT belongs to the insulin signaling pathway and is activated by anabolic hormones including insulin and in turn stimulates mTOR signaling (Sandri 2008). We found that neither age nor CR affected the total AKT content in muscles significantly (Fig. 1a). CR did not change phosphorylated AKT content significantly regardless of the age of the rats; however, the phosphorylated AKT content in muscles increased with aging (Fig. 1b).

Effects of age and CR on the content of AKT (a), phosphorylated AKT (b), mTOR (c), and phosphorylated mTOR (d) in soleus muscles of rats. Values were expressed as mean ± SE. *P < 0.05. CR, calorie restriction; mTOR,: mammalian target of rapamycin
mTOR is the hub responding to signals from amino acids, energy status, and anabolic hormones in order to regulate cellular protein synthesis (Sandri 2008). We found that neither age nor CR affected the total mTOR content in muscles significantly (Fig. 1c). The effect of CR on the phosphorylated mTOR content was age-dependent. CR reduced the phosphorylated mTOR content in muscles of middle-aged rats and had no effects on the muscles of adult and young rats. Notably, the phosphorylated mTOR content was greater in muscles of middle-aged rats than that in adult and young rats (Fig. 1d).
Two main downstream targets of mTOR are 4EBP1 and S6K that regulate translation of cells. S6K enhances translation efficiency by increasing RNA helicase activities (Ma and Blenis 2009). We found that the effects of CR on the total S6K content and the phosphorylated S6K content were age-dependent. CR decreased the total S6K and phosphorylated S6K content in muscles of middle-aged rats, whereas CR had no effects on muscles of adult and young rats. Notably, the total and phosphorylated S6K content was greater in muscles of middle-aged rats than that in adult and young rats (Fig. 2a, b).

Effects of age and CR on the content of S6K (a), phosphorylated S6K (b), 4EBP1 (c), and phosphorylated 4EBP1 (d) in soleus muscles of rats. Values were expressed as mean ± SE. *P < 0.05. CR, calorie restriction; S6K, ribosomal protein S6 protein kinases 1; 4EBP1, initiation factor 4E binding proteins
4EBP1 is phosphorylated by mTOR. Phosphorylation of 4EBP1 results in its release from eukaryotic translation initiation factor 4E that allows the translation initiation to proceed (Ma and Blenis 2009). We found that CR did not influence the total and phosphorylated 4EBP1 content regardless of the age of the rats. The total and phosphorylated 4EBP1 content increased with aging (Fig. 2c, d).
Regulators/markers in the ubiquitin-proteasome system
FOXO3a is a transcription factor that regulates the ubiquitin-proteasome proteolysis pathway (Sanchez et al. 2014). Phosphorylated FOXO3a remains in the cytosol and translocates to the nucleus when it is dephosphorylated. When FOXO3a translocates to the nucleus, it triggers protein degradation by increasing the transcription of genes including atrogin and MuRF1 (Sanchez et al. 2014). We found the effects of CR on the content of FOXO3a and phosphorylated FOXO3a were age-dependent. CR reduced the total FOXO3a content only in the middle-aged rats but not in the young and adult rats. Notably, the total FOXO3a content in muscles was greater in middle-aged rats than that in young rats (Fig. 3a). Regarding the content of phosphorylated FOXO3a, we found the effects of CR were age-dependent. However, post hoc analyses did not show any significant difference between the AL and the CR groups within each age group (Fig. 3b).

Effects of age and CR on the content of FOXO3a (a), phosphorylated FOXO3a (b), Atrogin (c), and MuRF1 (d) in soleus muscles of rats. Values were expressed as mean ± SE. *P < 0.05. Black dotted lines indicated noncontiguous gel lanes. AL, ad libitum; CR, calorie restriction; FOXO, forkhead box class O
The ubiquitin-proteasome system degrades misfolded and oxidative-damaged proteins (Lecker et al. 2006). MuRF1 and atrogin are E3 ubiquitin ligases. They are the two most upregulated proteins in conditions of muscle atrophy and are seen as two sensitive markers of protein degradation by the UPP (Sandri 2008). We found that CR did not affect the atrogin and MuRF1 content in muscles of rats, regardless of the age of the rats (Fig. 3c). However, MuRF1 content in muscles increased with aging (Fig. 3d).
The influence of CR on the content of ubiquitinated protein was age-dependent. CR decreased the ubiquitinated protein content in muscles of middle-aged rats whereas no influence was found in adult and young rats. Notably, the ubiquitinated protein content in muscles of middle-aged rats was greater than that in young and adult rats (Fig. 4a, b). The proteasome subunit alpha 7 represents the total proteasome content, and the proteasome subunit beta 5 is the catalytic subunit that performs protease activity (Budenholzer et al. 2017). We found the effects of CR on alpha 7 were age-dependent. CR increased alpha 7 content in muscles of middle-aged rats while CR had no effects on muscles of adult and young rats. Notably, the alpha 7 content in muscles was lower in muscles of middle-aged rats than that in young rats (Fig. 4c). Regarding beta 5 content, neither CR nor age affected it significantly (Fig. 4d).

Effects of age and CR on the content of ubiquitinated proteins (a), alpha 7 (c), and beta 5 (d) in soleus muscles of rats. b The representative blot of ubiquitinated proteins. Values were expressed as mean ± SE. *P < 0.05. Black dotted lines indicated noncontiguous gel lanes. AL, ad libitum; CR, calorie restriction
Discussion
The current study is the first to report age-dependent effects of CR on both mTOR signaling and UPP in skeletal muscles. Figure 5 highlights the major findings of the current study. Specifically, for middle-aged rats, CR reduced phosphorylation of mTOR and S6K and decreased the content of FOXO3a and ubiquitinated proteins in skeletal muscles. In contrast to the middle-aged rats, these pathways in skeletal muscles of young and adult rats did not change with CR.

Both anabolic and catabolic signaling are upregulated in muscles with aging. Calorie restriction (CR) reduced phosphorylation of mTOR and S6K and decreased the content of FOXO3a and ubiquitinated proteins in skeletal muscles in middle-aged rats. CR did not change these pathways in skeletal muscles of young and adult rats. FOXO, forkhead box class O; mTOR, mammalian target of rapamycin; S6K, ribosomal protein S6 protein kinases 1; 4EBP1, initiation factor 4E binding proteins
Over the past 4 decades the morphology and molecular-cellular properties of skeletal muscles with aging have been investigated using various animal models (e.g., flies, mice, rats, nonhuman primates, and humans). One well-studied characteristic is muscle mass. Skeletal muscles decrease in size with aging (Chen et al. 2015; Zengin et al. 2017). Using the same research design as the present study (e.g., same ages), we found muscle mass decreased 10–20% in rats between 8 (young) and 20 months of age (middle-aged) (Chen et al. 2015). Now, we report that during this time period (8 and 20 months), both anabolic and catabolic signaling are upregulated in muscles. The age-associated hyper-phosphorylation of mTOR and S6K and greater content of FOXO3a, MuRF1, and ubiquitinated proteins are consistent with previous reports using the rat model or human muscle biopsy (Baar et al. 2016; Chen et al. 2008; Clavel et al. 2006; Kimball et al. 2004; Markofski et al. 2015). In the current study, we found that CR reversed the aging muscles to a “youthful” state. The findings extend the knowledge in the field of aging by providing a potential explanation by which CR preserves skeletal muscle mass with aging.
It is well accepted that activation of the mTOR pathway facilitates muscle growth (Sandri 2008). However, this general concept may not hold true in aging skeletal muscles. For example, studies found that even though the phosphorylation status of mTOR and S6K is greater in muscles of older subjects compared to younger subjects, muscle protein fractional synthetic rate in both age groups was the same (Guillet et al. 2004; Markofski et al. 2015). This suggests that the efficiency of protein synthesis regulated by the mTOR pathway is altered in aging muscles. The next logical question to ask is how does a reduction of mTOR and S6K phosphorylation (the findings in the current study) contribute to the preservation of muscle mass of older animals. If a reduction in mTOR signaling plays a role in muscle mass maintenance, it is likely through indirect mechanisms. One potential indirect mechanism is the link between S6K and muscle insulin sensitivity. S6K phosphorylates insulin receptor substrate 1, which in turn inhibits the insulin signaling pathway (Tremblay et al. 2005; Um et al. 2004, 2006; Zhang et al. 2008). An impairment in insulin signaling potentially contributes to anabolic resistance (Morais et al. 2018). Anabolic resistance is a common phenomenon observed in muscles of older humans and animals where the sensitivity and capacity to activate mTOR signaling in response to stimuli such as nutrition and exercise are reduced (Francaux et al. 2016; Fujita et al. 2009; Morton et al. 2018; Sharples et al. 2015). Support for this idea is found in the study by Guillet et al. where they reported that although basal phosphorylation of S6K in the elderly is greater than that in young subjects, S6K phosphorylation level in the elderly does not increase in response to insulin and amino acid infusion (Guillet et al. 2004). Therefore, we suggest the CR-induced restoration of mTOR and S6K hyper-phosphorylation improves insulin signaling in muscles of older animals and indirectly maintains their muscle mass. To show a definite link between S6K phosphorylation, insulin signaling, anabolic resistance, and muscle mass in muscles of older animals, further studies are needed.
Although inhibition of mTOR by CR is beneficial, it is important to acknowledge the practice of CR is very challenging in daily life. Recognizing this challenge in the practice of CR, the scientific field sought CR mimetics by screening novel inhibitors of mTOR signaling (Lee et al. 2017). A well-studied CR mimetic is the mTOR inhibitor, rapamycin. It was shown that rapamycin attenuates alveolar bone loss in aged mice (An et al. 2017) and optimizes mitochondrial oxidative phosphorylation and senescence resistance of human cardiac fibroblast (Nacarelli et al. 2018). Thus, it is possible to obtain or achieve the benefits of mTOR signaling inhibition from CR mimetics or other pharmacological agents.
The signaling pathways and their indirect impact within skeletal muscles with age and CR are complex. To our surprise, the results of 4EBP1 did not follow the results of mTOR even though 4EBP1 is another down-stream target of mTOR. We assessed 4EBP1 because its phosphorylation initiates protein translation. One possible explanation for the different changes with CR between mTOR and 4EBP1 is that 4EBP1 is regulated by both mTOR-dependent and mTOR-independent pathways (AKT/glycogen synthase kinase-3β pathway) (Shin et al. 2014). In fact, the regulation of 4EBP1 by mTOR-independent pathway but not mTOR-dependent pathway was shown in another animal model where muscle protein synthesis was disrupted (Mirzoev et al. 2016). Future research is needed to investigate the effects of CR and age on 4EBP1 in both mTOR-dependent and mTOR-independent pathways.
The UPP is one of the key pathways that degrade cellular proteins. Proteins that are misfolded or oxidized are subjected to ubiquitination and then degraded by proteasomes. FOXO is a transcription factor that regulates this pathway (Sanchez et al. 2014). Thus, the content of FOXO and the extent of ubiquitinated proteins reflect the protein degradation state. Greater levels of FOXO and protein ubiquitination were reported in atrophying muscles under conditions such as inactivity (Bonaldo and Sandri 2013; Schiaffino et al. 2013). Our findings of the CR-induced downregulation of UPP in skeletal muscles of middle-aged rats but not young rats are as predicted. In fact, the result supports the published preservation of muscle mass with CR in the middle-aged rats (Chen et al. 2015). In the published CR literature, there are reports using various aged mammals and investigating components of the UPP (Lu et al. 2017; Mercken et al. 2013). The finding in our young and adult rats is also consistent with an investigation using neonatal calves with CR where the expression of E3 ubiquitin ligases did not change (Lu et al. 2017). Whereas the finding of CR-induced decrease in FOXO content in older animals is also similar to rats on lifelong CR (Mercken et al. 2013). Although UPP is one of the key pathways that degrade proteins, other pathways such as calpain, apoptosis, and autophagy-lysosome machinery also involve in cellular protein degradation. The role of other muscle degradation systems in the CR-induced preservation of muscle mass of older animals needs further studies.
The underlying mechanism to explain the responses in mTOR signaling and UPP with CR and age is likely related to the cellular environment. The inflammatory profile in skeletal muscles increases with aging (Kalinkovich and Livshits 2017; Mercken et al. 2017). Several regulators of protein turnover including mTOR, S6K, and FOXO3a are activated by inflammatory cytokines (Chung et al. 2011; Dalle et al. 2017; Ma and Blenis 2009; Zhang et al. 2008). Thus, our finding of the concurrent upregulation of both anabolic and catabolic signaling pathways in aging muscles is likely a response to the inflammatory environment present in skeletal muscles of older animals. Because CR decreases inflammation (Lopez-Lluch and Navas 2016; Mercken et al. 2013; Yang et al. 2016), its impact is significant in the older animals compared to younger animals.
Generally speaking, reduced food intake by CR provides a wide spectrum of beneficial effects including reversion of obesity-related endothelial dysfunction (Csipo et al. 2018), preservation of muscle mass of aging animals (Chen et al. 2015), and prevention of many chronic diseases such as diabetes and cardiovascular diseases (Mattison et al. 2017). Nevertheless, CR has potential adverse effects for certain biological systems and animals with specific genetic backgrounds (Contreras et al. 2018) (Fang et al. 2017). For example, healthy individuals with long-term CR had lower leukocyte count which may compromise the defense function of their immune system compared to age-matched individuals who maintained a Western diet (Contreras et al. 2018). Because the effects of CR are complex, prior to the initiation of CR, consideration of the goals of the CR and the physical status of individuals including age, current body composition, and health status is warranted.
In conclusion, our data indicate that the effects of CR on mTOR signaling and the UPP in skeletal muscles are age-dependent. CR did not alter mTOR signaling and UPP in soleus muscles of young and adult rats whereas CR altered the pathways in middle-aged rats.
References
An JY et al (2017) Rapamycin treatment attenuates age-associated periodontitis in mice. GeroScience 39:457–463
Baar EL, Carbajal KA, Ong IM, Lamming DW (2016) Sex- and tissue-specific changes in mTOR signaling with age in C57BL/6 J mice. Aging Cell 15:155–166
Bonaldo P, Sandri M (2013) Cellular and molecular mechanisms of muscle atrophy. Dis Model Mech 6:25–39
Budenholzer L, Cheng CL, Li Y, Hochstrasser M (2017) Proteasome structure and assembly. J Mol Biol 429:3500–3524
Chen CN, Ferrington DA, Thompson LV (2008) Carbonic anhydrase III and four-and-a-half LIM protein 1 are preferentially oxidized with muscle unloading. J Appl Physiol 105:1554–1561
Chen CN, Brown-Borg HM, Rakoczy SG, Ferrington DA, Thompson LV (2010) Aging impairs the expression of the catalytic subunit of glutamate cysteine ligase in soleus muscle under stress. J Gerontol A Biol Sci Med Sci 65:129–137
Chen CN, Lin SY, Liao YH, Li ZJ, Wong AM (2015) Late-onset caloric restriction alters skeletal muscle metabolism by modulating pyruvate metabolism. Am J Physiol Endocrinol Metab 308:E942–E949
Chen CN et al (2017) Diet-induced obesity accelerates blood lactate accumulation of rats in response to incremental exercise to maximum. Am J Phys Regul Integr Comp Phys 313:R601–R607
Chung HY et al (2011) Molecular inflammation as an underlying mechanism of the aging process and age-related diseases. J Dent Res 90:830–840
Clavel S, Coldefy AS, Kurkdjian E, Salles J, Margaritis I, Derijard B (2006) Atrophy-related ubiquitin ligases, atrogin-1 and MuRF1 are up-regulated in aged rat Tibialis Anterior muscle. Mech Ageing Dev 127:794–801
Colman RJ, Beasley TM, Allison DB, Weindruch R (2008) Attenuation of sarcopenia by dietary restriction in rhesus monkeys. J Gerontol A Biol Sci Med Sci 63:556–559
Combaret L, Dardevet D, Bechet D, Taillandier D, Mosoni L, Attaix D (2009) Skeletal muscle proteolysis in aging. Curr Opin Clin Nutr Metab Care 12:37–41
Contreras NA, Fontana L, Tosti V, Nikolich-Zugich J (2018) Calorie restriction induces reversible lymphopenia and lymphoid organ atrophy due to cell redistribution. GeroScience 40:279–291
Csipo T, Fulop GA, Lipecz A, Tarantini S, Kiss T, Balasubramanian P, Csiszar A, Ungvari Z, Yabluchanskiy A (2018) Short-term weight loss reverses obesity-induced microvascular endothelial dysfunction. GeroScience 40:337–346. https://doi.org/10.1007/s11357-018-0028-9
Dalle S, Rossmeislova L, Koppo K (2017) The role of inflammation in age-related sarcopenia. Front Physiol 8:1045
Fang Y et al (2017) Differential effects of early-life nutrient restriction in long-lived GHR-KO and normal mice. GeroScience 39:347–356
Fontana L, Partridge L, Longo VD (2010) Extending healthy life span--from yeast to humans. Science 328:321–326
Francaux M, Demeulder B, Naslain D, Fortin R, Lutz O, Caty G, Deldicque L (2016) Aging reduces the activation of the mTORC1 pathway after resistance exercise and protein intake in human skeletal muscle: potential role of REDD1 and impaired anabolic sensitivity. Nutrients 8
Fujita S, Glynn EL, Timmerman KL, Rasmussen BB, Volpi E (2009) Supraphysiological hyperinsulinaemia is necessary to stimulate skeletal muscle protein anabolism in older adults: evidence of a true age-related insulin resistance of muscle protein metabolism. Diabetologia 52:1889–1898
Guillet C et al (2004) Impaired anabolic response of muscle protein synthesis is associated with S6K1 dysregulation in elderly humans. FASEB J 18:1586–1587
Ibebunjo C et al (2013) Genomic and proteomic profiling reveals reduced mitochondrial function and disruption of the neuromuscular junction driving rat sarcopenia. Mol Cell Biol 33:194–212
Kaeberlein M et al (2005) Regulation of yeast replicative life span by TOR and Sch9 in response to nutrients. Science 310:1193–1196
Kalinkovich A, Livshits G (2017) Sarcopenic obesity or obese sarcopenia: a cross talk between age-associated adipose tissue and skeletal muscle inflammation as a main mechanism of the pathogenesis. Ageing Res Rev 35:200–221
Kimball SR, O'Malley JP, Anthony JC, Crozier SJ, Jefferson LS (2004) Assessment of biomarkers of protein anabolism in skeletal muscle during the life span of the rat: sarcopenia despite elevated protein synthesis. Am J Physiol Endocrinol Metab 287:E772–E780
Lecker SH, Goldberg AL, Mitch WE (2006) Protein degradation by the ubiquitin-proteasome pathway in normal and disease states. J Am Soc Nephrol 17:1807–1819
Lee MB et al (2017) A system to identify inhibitors of mTOR signaling using high-resolution growth analysis in Saccharomyces cerevisiae. GeroScience 39:419–428
Lopez-Lluch G, Navas P (2016) Calorie restriction as an intervention in ageing. J Physiol 594:2043–2060
Lu Y, Bradley JS, McCoski SR, Gonzalez JM, Ealy AD, Johnson SE (2017) Reduced skeletal muscle fiber size following caloric restriction is associated with calpain-mediated proteolysis and attenuation of IGF-1 signaling. Am J Phys Regul Integr Comp Phys 312:R806–R815
Ma XM, Blenis J (2009) Molecular mechanisms of mTOR-mediated translational control. Nat Rev Mol Cell Biol 10:307–318
Markofski MM et al (2015) Effect of age on basal muscle protein synthesis and mTORC1 signaling in a large cohort of young and older men and women. Exp Gerontol 65:1–7
Marzetti E, Calvani R, Cesari M, Buford TW, Lorenzi M, Behnke BJ, Leeuwenburgh C (2013) Mitochondrial dysfunction and sarcopenia of aging: from signaling pathways to clinical trials. Int J Biochem Cell Biol 45:2288–2301
Mattison JA et al (2017) Caloric restriction improves health and survival of rhesus monkeys. Nat Commun 8:14063
McKiernan SH et al (2012) Cellular adaptation contributes to calorie restriction-induced preservation of skeletal muscle in aged rhesus monkeys. Exp Gerontol 47:229–236
Mercken EM et al (2013) Calorie restriction in humans inhibits the PI3K/AKT pathway and induces a younger transcription profile. Aging Cell 12:645–651
Mercken EM et al (2017) Conserved and species-specific molecular denominators in mammalian skeletal muscle aging. NPJ Aging Mech Dis 3:8
Messaoudi I et al (2008) Optimal window of caloric restriction onset limits its beneficial impact on T-cell senescence in primates. Aging Cell 7:908–919
Mirzoev T, Tyganov S, Vilchinskaya N, Lomonosova Y, Shenkman B (2016) Key markers of mTORC1-dependent and mTORC1-independent signaling pathways regulating protein synthesis in rat soleus muscle during early stages of hindlimb unloading. Cell Physiol Biochem 39:1011–1020
Morais JA, Jacob KW, Chevalier S (2018) Effects of aging and insulin resistant states on protein anabolic responses in older adults. Exp Gerontol 108:262–268
Morton RW, Traylor DA, Weijs PJM, Phillips SM (2018) Defining anabolic resistance: implications for delivery of clinical care nutrition. Curr Opin Crit Care 24:124–130
Nacarelli T, Azar A, Altinok O, Orynbayeva Z, Sell C (2018) Rapamycin increases oxidative metabolism and enhances metabolic flexibility in human cardiac fibroblasts. GeroScience 40:243–256. https://doi.org/10.1007/s11357-018-0030-2
Niemann B, Silber RE, Rohrbach S (2008) Age-specific effects of short- and long-term caloric restriction on the expression of adiponectin and adiponectin receptors: influence of intensity of food restriction. Exp Gerontol 43:706–713
Peake J, Della Gatta P, Cameron-Smith D (2010) Aging and its effects on inflammation in skeletal muscle at rest and following exercise-induced muscle injury. Am J Phys Regul Integr Comp Phys 298:R1485–R1495
Sakuma K, Aoi W, Yamaguchi A (2017) Molecular mechanism of sarcopenia and cachexia: recent research advances. Pflugers Arch 469:573–591
Sanchez AM, Candau RB, Bernardi H (2014) FoxO transcription factors: their roles in the maintenance of skeletal muscle homeostasis. Cell Mol Life Sci 71:1657–1671
Sandri M (2008) Signaling in muscle atrophy and hypertrophy. Physiology 23:160–170
Schiaffino S, Dyar KA, Ciciliot S, Blaauw B, Sandri M (2013) Mechanisms regulating skeletal muscle growth and atrophy. FASEB J 280:4294–4314
Sharples AP, Hughes DC, Deane CS, Saini A, Selman C, Stewart CE (2015) Longevity and skeletal muscle mass: the role of IGF signalling, the sirtuins, dietary restriction and protein intake. Aging Cell 14:511–523
Shin S, Wolgamott L, Tcherkezian J, Vallabhapurapu S, Yu Y, Roux PP, Yoon SO (2014) Glycogen synthase kinase-3beta positively regulates protein synthesis and cell proliferation through the regulation of translation initiation factor 4E-binding protein 1. Oncogene 33:1690–1699
Testa G, Biasi F, Poli G, Chiarpotto E (2014) Calorie restriction and dietary restriction mimetics: a strategy for improving healthy aging and longevity. Curr Pharm Des 20:2950–2977
Tremblay F et al (2005) Overactivation of S6 kinase 1 as a cause of human insulin resistance during increased amino acid availability. Diabetes 54:2674–2684
Um SH et al (2004) Absence of S6K1 protects against age- and diet-induced obesity while enhancing insulin sensitivity. Nature 431:200–205
Um SH, D'Alessio D, Thomas G (2006) Nutrient overload, insulin resistance, and ribosomal protein S6 kinase 1, S6K1. Cell Metab 3:393–402
Yang L et al (2016) Long-term calorie restriction enhances cellular quality-control processes in human skeletal muscle. Cell Rep 14:422–428
Zengin A et al (2017) Associations of muscle force, power, cross-sectional muscle area and bone geometry in older UK men. J Cachexia Sarcopenia Muscle 8:598–606
Zhang J, Gao Z, Yin J, Quon MJ, Ye J (2008) S6K directly phosphorylates IRS-1 on Ser-270 to promote insulin resistance in response to TNF-(alpha) signaling through IKK2. J Biol Chem 283:35375–35382
Acknowledgments
This work was supported by Ministry of Science and Technology (106-2314-B-010 -052 -MY3); and Chang Gung Memorial Hospital (CMPRD3D0213). We thank Yu-Mei Chih for technical assistance and Yu-Chieh Lin for drawing the cartoon pictures.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
About this article

Cite this article
Chen, CN., Liao, YH., Tsai, SC. et al. Age-dependent effects of caloric restriction on mTOR and ubiquitin-proteasome pathways in skeletal muscles. GeroScience 41, 871–880 (2019). https://doi.org/10.1007/s11357-019-00109-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11357-019-00109-8