
Abstract
Inadequate substrate availability and increased nitric oxide synthase inhibitor levels attenuate nitric oxide (NO) synthesis, whereas increased vascular oxidative stress may lead to inactivation of NO. We compared markers of NO synthesis capacity and oxidative stress in a bi-ethnic male population. Inter-relationships of ambulatory blood pressure and urinary albumin-to-creatinine ratio with NO synthesis capacity and oxidative stress markers were investigated. NO synthesis capacity markers (L-arginine, asymmetric dimethylarginine (ADMA), and symmetric dimethylarginine (SDMA)) and oxidative stress markers (serum peroxides, total glutathione, glutathione peroxidase (GPx), glutathione reductase (GR), superoxide dismutase (SOD), and catalase) were measured. Black men displayed higher blood pressure and albumin-to-creatinine ratio (all p < 0.001), while NO synthesis capacity was more favorable (higher L-arginine and lower ADMA (p ≤ 0.003)). Antioxidant enzyme activities were similar except for the redox status markers (GR activity and GR/GPx ratio), which were upregulated in black men (p < 0.001). In black men, ADMA was inversely related to GPx activity (R 2 = 0.15; β = −0.20; p = 0.050) and GPx/SOD ratio (R 2 = 0.24; β = −0.37; p < 0.001), but none of these markers related to blood pressure or albumin-to-creatinine ratio. In white men, albumin-to-creatinine ratio was positively associated with ADMA (R 2 = 0.18; β = 0.39; p < 0.001) while ADMA was inversely related to GR activity (R 2 = 0.26; β = −0.29; p = 0.002) and GR/GPx ratio (R 2 = 0.25; β = −0.28; p = 0.003). Black men with elevated blood pressure and albumin-to-creatinine ratio displayed a favorable NO synthesis capacity. This may be counteracted by increased inactivation of NO, although it was not linked to vascular or renal phenotypes. In white men, reduced NO synthesis capacity may lower NO bio-availability, thereby influencing the albumin-to-creatinine ratio.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cardiovascular disease is a health challenge globally (Kearney et al. 2005) and especially within South Africa (Sliwa et al. 2008) with the highest prevalence of hypertension in developing countries reported in adults over 50 years of age (Lloyd-Sherlock et al. 2014). Vascular endothelial dysfunction is associated with lower bio-available nitric oxide (NO) (Endemann and Schiffrin 2004), hypertension, and the subsequent increased prevalence of cardiovascular disease (Dharmashankar and Widlansky 2010).
Since NO is essential in the regulation of endothelial function (Tousoulis et al. 2012), factors influencing its bio-availability are frequently investigated. Elements involved in reduced NO synthesis include lower nitric oxide synthase activity, lower levels of the substrate L-arginine (Tousoulis et al. 2012), as well as nitric oxide synthase inhibitors such as asymmetric dimethylarginine (ADMA) (Böger 2005) and symmetric dimethylarginine (SDMA) (Siegerink et al. 2013). Moreover, NO bio-availability is suppressed by factors that enhance NO inactivation such as increased vascular oxidative stress (Wang et al. 2009) and decreased antioxidant defense (Sözmen et al. 1998).
Previous studies in South Africa showed similar ADMA levels in black and white men, but that ADMA was positively associated with arterial stiffness only in black men (Schutte et al. 2010). In another study of black South African men, systolic blood pressure associated positively with reactive oxygen species (Kruger et al. 2012) and attenuated total glutathione levels were linked to subclinical atherosclerosis in hypertensive black men (Schutte et al. 2009). Taken together, these studies suggest that factors known to influence NO bio-availability may contribute to vascular deterioration in black South Africans. Furthermore, we have not yet investigated the association of NO synthesis capacity with oxidative stress-related markers in this population group. We therefore investigated associations of NO synthesis capacity markers with oxidative stress-related markers, and we attempted to link blood pressure and renal microvascular disease with markers affecting NO bio-availability in these groups.
Materials and methods
Study population and protocol
This study is embedded in the Sympathetic activity and Ambulatory Blood Pressure in Africans (SABPA) study, a cross-sectional target population study. The study population and protocol has been described elsewhere (Malan et al. 2014). In the present study, we also excluded women (n = 202), human immunodeficiency virus-infected participants (n = 13), and participants with missing data (n = 5) (NO synthesis capacity markers and oxidative stress-related markers) leaving a total of 88 black and 101 white men. The study was conducted in line with the ethical principles of the Declaration of Helsinki and was approved by the Ethics Committee of the North-West University (Potchefstroom Campus) (NWU-00036-07-S6).
Anthropometric and physical activity measurements
Weight and height were measured using calibrated instruments (Precision Health Scale, A & D Company, Tokyo, Japan, and Invicta Stadiometer, IP 1465, Invicta, London, UK) to the nearest 1.0 cm or 1.0 kg, respectively, and body mass index was calculated. All anthropometric measurements were taken in triplicate using standard international protocols (Marfell-Jones et al. 2012). The Actical® (Mini Mitter, Bend OR, Montréal, Québec), an omnidirectional accelerometer, was worn around the waist and determined physical activity for 24-h taking the resting metabolic rate into account.
Cardiovascular measurements
The ambulatory blood pressure monitoring device was programmed to measure blood pressure at 30-min intervals during the day (07:00–22:00) and 1-h intervals during nighttime (22:00–06:00) (Kohara et al. 1995). The ambulatory blood pressure data was loaded onto a database using the CardioVisions 1.19 Personal Edition (Meditech, Budapest, Hungary). The mean successful inflation rate over the 24-h period was 80.6 %. Hypertensive status was classified from the ambulatory blood pressure measurements as systolic blood pressure ≥130 mmHg and/or diastolic blood pressure ≥80 mmHg according to the 2013 European Society of Hypertension/European Society of Cardiology guidelines for a 24-h period (Mancia et al. 2013).
Biochemical analyses
A registered nurse obtained fasting blood samples with a sterile winged infusion set from the antebrachial vein branches. Blood samples were collected in serum separating tubes and ethylenediaminetetraacetic acid BD Vacutainers (Becton Dickinson (Pty) Ltd., Franklin lakes, NJ), and serum and plasma were obtained, respectively, according to standardized procedures. Urine was collected overnight for a period of 8-h. All samples were stored at −80 °C until analysis.
L-arginine, ADMA, and SDMA were quantified by liquid chromatography tandem mass spectrometry (Schwedhelm et al. 2005). In brief, 25 μL of plasma was diluted with stable isotope-labeled internal standards (i.e., [2H7]-arginine and [2H6]-ADMA). Subsequently, proteins were precipitated with methanol and guanidine compounds were converted to their butyl esters. Concentrations were calculated with calibration curves (four levels, triplicates), and plate-wise quality controls were run (two levels, duplicates). Intra- and inter-assay coefficients of variation were ≤7.5 % for all. Samples were re-analyzed for coefficients of variation and bias of quality controls ≥15 %. L-citrulline was determined with an electrospray ionization tandem mass spectrometry method as previously described (Mels et al. 2013).
One of the measurable reactive oxygen species, namely total peroxides, was determined in serum samples (Hayashi et al. 2007). Total glutathione levels were determined with the BIOXYTECH® GSH/GSSG-412™ kit supplied by OxisResearch™, (Foster City, CA). Glutathione peroxidase (GPx), glutathione reductase (GR), and superoxide dismutase (SOD) activities were determined with assay kits from Cayman Chemical Company (Ann Arbor, MI). Catalase (CAT) enzyme activity was determined with a fluorometric kit from OxiSelect™ (Cell Biolabs Inc., San Diego, CA). Antioxidant enzyme ratios were calculated to assess antioxidant defenses and included the glutathione reductase-to-glutathione peroxidase ratio (GR/GPx ratio) and the glutathione peroxidase-to-superoxide dismutase ratio (GPx/SOD ratio).
Glycated hemoglobin was determined by a turbidimetric inhibition immunoassay (Integra 400, Roche, Basel, Switzerland) in ethylenediaminetetraacetic acid whole blood. Total cholesterol, high-density lipoprotein cholesterol, triglycerides, high-sensitivity C-reactive protein, γ-glutamyltransferase, and creatinine were analyzed in serum (Unicel DXC 800, Beckman and Coulter, Germany and Konelab™ 20I Sequential Multiple Analyzer Computer, Thermo Scientific, Vantaa, Finland). Estimated glomerular filtration rate, a measure of kidney function, was calculated with the abbreviated modification of diet in renal disease equation. Serum cotinine was determined with a homogeneous immunoassay (Automated Modular, Roche, Basel, Switzerland). Creatinine and albumin were analyzed in urine samples (Unicel DXC 800, Beckman and Coulter, Germany) and the albumin-to-creatinine ratio, a measure of renal microvascular disease, was calculated. Both the intra- and inter-assay coefficients of variation for all the assays were less than 10 %.
Statistical analyses
Statistical analyses were performed with Statistica 12 (Statsoft Inc., Tulsa, OK, USA). Data were expressed as arithmetic mean and standard deviation for normally distributed variables. Variables with a non-Gaussian distribution were logarithmically transformed (L-arginine, ADMA, GR activity, SOD activity, GR/GPx ratio, GPx/SOD ratio, C-reactive protein, γ-glutamyltransferase, estimated glomerular filtration rate, and albumin-to-creatinine ratio) and the central tendency and spread represented by the geometric mean and the 5th and 95th percentile intervals. Means and proportions were compared using independent T tests and Chi-square tests, respectively. Interactions of race were tested for the relationship between NO synthesis capacity markers and oxidative stress markers using multiple regression analysis. Associations of NO synthesis capacity markers (L-arginine, ADMA, and SDMA) with oxidative stress-related markers (serum peroxides, GPx activity, GR activity, SOD activity, CAT activity, GR/GPx ratio, and GPx/SOD ratio) were investigated using single, partial (adjusted for age and body mass index), and multiple regression analyses. In multiple regression analyses, L-arginine and ADMA were the dependent variables, with an oxidative stress-related marker as main independent variable. Other covariates included age, body mass index, total energy expenditure, γ-glutamyltransferase, cotinine, glycated hemoglobin, C-reactive protein, and albumin-to-creatinine ratio. In sensitivity analyses, albumin-to-creatinine ratio was replaced with estimated glomerular filtration rate in models with ADMA as the dependent variable, and all the multiple regression analyses were repeated. Additionally, associations between systolic blood pressure, diastolic blood pressure, and albumin-to-creatinine ratio with NO synthesis capacity and oxidative stress-related markers were investigated using single, partial, and multiple regression analyses. Main independent variables included ADMA and an oxidative stress-related marker (GPx, GR, or GR/GPx). Other covariates included age, body mass index, total energy expenditure, γ-glutamyltransferase, cotinine, glycated hemoglobin, C-reactive protein, and estimated glomerular filtration rate. In models with albumin-to-creatinine ratio as the dependent variable, ambulatory systolic blood pressure was also included as an independent variable. In sensitivity analyses, participants using anti-hypertensive medication (n = 45), antioxidant intake (n = 1), and multivitamin intake (n = 8) were excluded and multiple regression analyses were repeated.
Results
Characteristics of the study population
An interaction existed with race for the following relationships: reactive oxygen species and L-arginine (p = 0.021), GPx/SOD ratio and L-arginine (p = 0.027), CAT and ADMA (p = 0.048), and GR/GPx ratio and ADMA (p = 0.005). The group was therefore stratified by race. The characteristics of the study population are presented in Table 1. Black men presented with higher blood pressure (systolic blood pressure and diastolic blood pressure (both p < 0.001)) and worse albumin-to-creatinine ratio (p < 0.001) compared to white men. Contradictory to their cardiovascular profile, the markers indicative of NO synthesis capacity were more favorable in black men, with significantly higher L-arginine (p = 0.003), and L-citrulline (p = 0.020), while the NO synthesis inhibitors (ADMA and SDMA, p < 0.001 for both) were significantly lower. However, assessment of oxidative stress-related markers showed significantly higher reactive oxygen species levels (p = 0.017), while some of the markers indicative of redox status such as total glutathione, GR activity, and the GR/GPx ratio (all p < 0.001) were also significantly higher, suggesting upregulation of this system in the black men. After excluding participants using anti-hypertensive drugs, the favorable NO synthesis capacity profile as well as higher redox status markers remained in black men when compared to white men.
Regression analyses in black men
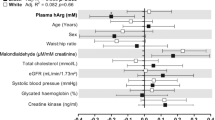
Results of single (data not shown), partial (Table S1) and multiple regression analyses (Fig. 1) in black men indicated a positive association of ADMA with SOD activity (R 2 = 0.25; β = 0.37; p < 0.001) and CAT activity (R 2 = 0.21; β = 0.32; p = 0.002), while ADMA was negatively associated with other antioxidant enzyme activities such as GPx (R 2 = 0.15; β = −0.20; p = 0.050) and the GPx/SOD ratio (R 2 = 0.24; β = −0.37; p < 0.001). When albumin-to-creatinine ratio was replaced with estimated glomerular filtration rate in the multiple regression analyses, the results remained unchanged, except for the negative association of ADMA with GPx, which still tended to be significant (R 2 = 0.19; β = −0.19; p = 0.059). Excluding participants on anti-hypertensive medication, multivitamins and anti-oxidants did not change the result, except for the association of ADMA with GPx, which lost significance (anti-hypertensive medication: R 2 = 0.16; β = −0.15; p = 0.25; multivitamin and anti-oxidant usage: R 2 = 0.13; β = −0.19; p = 0.075).

Forward stepwise multiple regression analyses of ADMA with oxidative stress-related markers in black and white men
Although we managed to demonstrate a link between ADMA and oxidative stress, none of the NO synthesis capacity markers or oxidative stress markers showed any significant associations with blood pressure or renal microvascular disease in this group, except for the previously reported (Kruger et al. 2012) association of systolic blood pressure and diastolic blood pressure with reactive oxygen species. Additionally, systolic blood pressure was identified as the most important contributor to the variance in albumin-to-creatinine ratio in this group (Table 2).
Regression analyses in white men
Negative associations of ADMA with GR activity (R 2 = 0.26; β = −0.29; p = 0.002), GR/GPx ratio (R 2 = 0.25; β = −0.28; p = 0.003), and GPx/SOD ratio (R 2 = 0.23; β = −0.20; p = 0.028) were indicated in single (data not shown), partial (Table S1), and multiple regression analyses (Fig. 1). When albumin-to-creatinine ratio was replaced with estimated glomerular filtration rate in the multiple regression analyses, the results remained unchanged. After excluding participants taking multivitamins or antioxidants, the result remained unchanged, but after excluding participants on anti-hypertensive medication, the association of ADMA with the GPx/SOD ratio (R 2 = 0.14; β = −0.12; p = 0.25) was no longer significant.
A positive association was observed between L-arginine and reactive oxygen species (R 2 = 0.17; β = 0.32; p = 0.001), whereas inverse relationships of L-arginine with GR activity (R 2 = 0.20; β = −0.29; p = 0.004) and the GR/GPx ratio (R 2 = 0.20; β = −0.28; p = 0.005) were found in single (data not shown), partial (Table S1), and multiple regression analyses (Fig. 2). L-Arginine also tended to be positively associated with the GPx/SOD ratio (R 2 = 0.16; β = −0.19; p = 0.052). After excluding participants on anti-hypertensive medication, only the association of L-arginine with GR activity remained significant. After excluding participants taking multivitamins or antioxidants, the association between L-arginine and reactive oxygen species lost significance (R 2 = 0.12; β = 0.21; p = 0.12). All the other results remained unchanged. Lastly, ADMA was found to be positively associated with the albumin-to-creatinine ratio (Table 2).

Forward stepwise multiple regression analyses of L-arginine with oxidative stress-related markers in white men
Discussion
Black men, with elevated blood pressure and albumin-to-creatinine ratio, surprisingly displayed a more favorable NO synthesis capacity than white men. Increased oxidative stress, as indicated by higher reactive oxygen species (serum peroxide) and an upregulated redox system (Lammertyn et al. 2015; Mels et al. 2014) in the black men, may counteract NO bio-availability, by increasing NO inactivation. However, we were unable to link these factors to vascular and renal phenotypes in the black men.
The more favorable NO synthesis capacity (higher L-arginine and lower ADMA and SDMA) observed in the black men in our study (Reimann et al. 2013) contradicts previous findings from Europe indicating higher ADMA levels in black men when compared to white men (Melikian et al. 2007). Within the South African context, similar ADMA and SDMA levels (Schutte et al. 2010) and lower L-arginine levels (Glyn et al. 2012) were observed in black compared to white men. The reason for these inconsistent findings is unknown but may relate to factors such as age, locality, socioeconomic status, and hypertension. The results reported by Melikian (Melikian et al. 2007) were for younger (mean age 32 years), normotensive black men living in Europe, whereas the mean age of the black men in our sample from South Africa was 43 years, with 63 % being hypertensive. Further to this, results obtained from the South African study regarding the role of Sex, Age and Ethnicity on Insulin sensitivity and Cardiovascular function (SAfrEIC study) comparing black and white men did not match the groups by socioeconomic status (Glyn et al. 2012) which may have contributed to differences in the results observed. In the present study, we tried to obtain more homogenous socioeconomic groupings by including only school teachers.
We further demonstrated that although antioxidant enzyme activities such as GPx, SOD, and CAT were comparable between black and white men, these antioxidant enzyme activities were independently associated with ADMA levels, only in black men. The independent inverse relationship between ADMA and GPx activity is in agreement with previous results obtained in children with primary hypertension and the metabolic syndrome (Śladowska-Kozłowska et al. 2012) and in young healthy adults (Puchau et al. 2009). Our results align with previous experimental work in rats, suggesting that ADMA administration decreases GPx activity (Alacam et al. 2013). However, these authors also indicated that CAT activity and SOD activity decreased after ADMA administration (Alacam et al. 2013), which is contradictory to the positive associations we found between these two enzymes and ADMA levels. Additionally, the GPx/SOD ratio, a marker of oxidative stress with greater discriminating power than the individual enzymes (De Haan et al. 1995), was inversely related to ADMA in both groups.
Since ADMA competes with L-arginine for binding to nitric oxide synthase (Böger 2007), it may lead to electrons being diverted to molecular oxygen, with consequent increased production of superoxide rather than NO (Vásquez-Vivar et al. 1998). The increased superoxide production may in turn call for an increase in SOD activity to facilitate the conversion of superoxide to hydrogen peroxide (Dittmar et al. 2008). High SOD activity along with low GPx activity may lead to the accumulation of hydrogen peroxide and hydrogen peroxide-derived reactive oxygen species such as hydroxyl radicals. The accumulation of hydrogen peroxide may further lead to the inactivation of NO and the formation of singlet oxygen, with highly cytotoxic effects (Noronha-Dutra et al. 1993). Our findings may therefore suggest that the favorable NO synthesis capacity observed in black men may be counteracted by increased inactivation of NO. This is further supported by the absence of any relationship between NO synthesis capacity markers or oxidative stress markers with blood pressure or renal microvascular disease in this group. Instead, higher blood pressure seems to be the driving force behind the increased albumin-to-creatinine ratio in the black men (Schutte et al. 2011). The inverse association of ADMA with the GPx/SOD ratio, but not with the individual enzymes as noted in the white men, confirms the importance of the previously suggested biological optimum ratio between SOD and the combined activity of GPx and CAT, rather than the absolute activities of these enzymes (De Haan et al. 1995).
Another prominent finding from our study is that although white men displayed a lower albumin-to-creatinine ratio, which may also indicate better systemic endothelial function (Pedrinelli et al. 2001), albumin-to-creatinine ratio was positively associated with ADMA levels. This was also found in untreated hypertensive patients (Tsioufis et al. 2010). However, what should be highlighted is the absence of this relationship in our black group.
The inverse relationship of ADMA with measures of the redox cycle, such as GR activity and the GR/GPx ratio, in white men suggests that ADMA degradation may be diminished in this group. These results also support previous experimental findings indicating that glutathione is important in the regulation of ADMA degradation (Fan et al. 2013). The degradation of ADMA is largely dependent on dimethylarginine dimethylaminohydrolase activity (Cooke 2004), and oxidative stress has been implicated in the attenuation of dimethylarginine dimethylaminohydrolase activity (Ito et al. 1999). Sensitivity of dimethylarginine dimethylaminohydrolase to oxidative stress is the result of a critical sulfhydryl group forming part of the active site of the enzyme (Leiper et al. 2002). Since reduced glutathione also contains a sulfhydryl group, it may be protective against oxidation of the dimethylarginine dimethylaminohydrolase sulfhydryl group, thereby preserving enzyme activity, and in turn ADMA degradation. In our study, this is further supported by the lower total glutathione levels, GR activity, and the potentially slower glutathione redox cycle (GR/GPx ratio) in white men when compared to their black counterparts, suggesting that diminished available total glutathione and glutathione regeneration may play a role in the higher ADMA levels observed in white men.
This study has to be interpreted within the context of its limitations and strengths. As a result of the cross-sectional study design, we cannot infer causality. Additionally, some of the non-Gaussian distributed variables were logarithmically transformed to enable the use of parametric statistical analyses. While our results were consistent after multiple adjustments, we cannot exclude residual confounding effects due to unknown factors associated with these variables. This study also lacks dietary data to quantify amino acid and antioxidant intake. However, the major strength of this study lies in the fact that this bi-ethnic cohort shared the same age, geographic location, education, and occupation. We also included various factors involved in the synthesis of NO as well as oxidative stress-related markers to help elucidate the mechanistic relationships between these variables within this population.
Conclusions
Our results suggest that different mechanisms involved in NO bio-availability may be at play in our bi-ethnic population. Although inactivation of NO may be more important in black men, we were unable to link factors involved in NO synthesis to blood pressure or renal microvascular disease, whereas in white men, reduced NO synthesis capacity may lower NO bio-availability thereby influencing the albumin-to-creatinine ratio. The elucidation of different mechanisms influencing NO bio-availability may aid in the development of more targeted treatment strategies.
References
Alacam H et al. (2013) Does ADMA affect the oxidant/antioxidant balance in rats? Turk J Med Sci 43:405–410
Böger RH (2005) Asymmetric dimethylarginine (ADMA) and cardiovascular disease: insights from prospective clinical trials. Vasc Med 10:S19–S25. doi:10.1191/1358863x05vm602oa
Böger RH (2007) The pharmacodynamics of L-arginine. J Nutr 137:1650S–1655S
Cooke JP (2004) Asymmetrical dimethylarginine the über marker? Circulation 109:1813–1818. doi:10.1161/01.CIR.0000126823.07732.D5
De Haan J, Cristiano F, Iannello RC, Kola I (1995) Cu/Zn-superoxide dismutase and glutathione peroxidase during aging. Biochem Mol Biol Int 35:1281–1297
Dharmashankar K, ME W (2010) Vascular endothelial function and hypertension: insights and directions. Curr Hypertens Rep 12:448–455. doi:10.1007/s11906-010-0150-2
Dittmar M, Knuth M, Beineke M, Epe B (2008) Role of oxidative DNA damage and antioxidative enzymatic defence systems in human aging. Open Anthropol J 1:38–45. doi:10.2174/1874912700801010038
Endemann DH, EL S (2004) Endothelial dysfunction. J Am Soc Nephrol 15:1983–1992. doi:10.1097/01.ASN.0000132474.50966
Fan N-C, Tsai C-M, Hsu C-N, Huang L-T, Tain Y-L (2013) N-acetylcysteine prevents hypertension via regulation of the ADMA-DDAH pathway in young spontaneously hypertensive rats Biomed Res Int 2013:1–9 doi:org/10.1155/2013/696317
Glyn M et al. (2012) Ethnicity-specific differences in L-arginine status in South African men. J Hum Hypertens 26:737–743. doi:10.1038/jhh.2011.103
Hayashi I, Morishita Y, Imai K, Nakamura M, Nakachi K, Hayashi T (2007) High-throughput spectrophotometric assay of reactive oxygen species in serum. Mutat Res Genet Toxicol Environ Mutagen 631:55–61. doi:10.1016/j.mrgentox.2007.04.006
Ito A, Tsao PS, Adimoolam S, Kimoto M, Ogawa T, JP C (1999) Novel mechanism for endothelial dysfunction dysregulation of dimethylarginine dimethylaminohydrolase. Circulation 99:3092–3095. doi:10.1161/01.CIR.99.24.3092
Kearney PM, Whelton M, Reynolds K, Muntner P, PK W, J H (2005) Global burden of hypertension: analysis of worldwide data. Lancet 365:217–223. doi:10.1016/S0140-6736(05)17741-1
Kohara K, Nishida W, Maguchi M, Hiwada K (1995) Autonomic nervous function in non-dipper essential hypertensive subjects evaluation by power spectral analysis of heart rate variability. Hypertension 26:808–814. doi:10.1161/01.HYP.26.5.808
Kruger R et al. (2012) Associations between reactive oxygen species, blood pressure and arterial stiffness in black south africans: the SABPA study. J Hum Hypertens 26:91–97. doi:10.1038/jhh.2010.134
Lammertyn L, Mels CM, Pieters M, Schutte AE, Schutte R (2015) Ethnic-specific relationships between haemostatic and oxidative stress markers in black and white South Africans: The SABPA study. Clin Exp Hypertens 28:1–7. doi:10.3109/10641963.2015.1013123
Leiper J, Murray-Rust J, McDonald N, Vallance P (2002) S-nitrosylation of dimethylarginine dimethylaminohydrolase regulates enzyme activity: further interactions between nitric oxide synthase and dimethylarginine dimethylaminohydrolase. Proc Natl Acad Sci 99:13527–13532. doi:10.1073/pnas.212269799
Lloyd-Sherlock P, Beard J, Minicuci N, Ebrahim S, Chatterji S (2014) Hypertension among older adults in low-and middle-income countries: prevalence, awareness and control. Int J Epidemiol 43:116–128. doi:10.1093/ije/dyt215
Malan L, Hamer M, Frasure-Smith N, Steyn HS, Malan NT (2014) COHORT PROFILE: Sympathetic activity and Ambulatory Blood Pressure in Africans (SABPA) Prospective Cohort Study Int J Epidemiol doi:10.1093/ije/dyu199
Mancia G et al. (2013) 2013 practice guidelines for the management of arterial hypertension of the European Society of Hypertension (ESH) and the European Society of Cardiology (ESC): ESH/ESC Task Force for the management of arterial hypertension. J Hypertens 31:1925–1938. doi:10.1097/HJH.0b013e328364ca4c
Marfell-Jones MJ, Stewart A, de Ridder J (2012) International standards for anthropometric assessment. International Society for the Advancement of Kinanthropometry. Wellington, New Zealand
Melikian N et al. (2007) Asymmetric dimethylarginine and reduced nitric oxide bioavailability in young Black African men. Hypertension 49:873–877. doi:10.1161/01.HYP.0000258405.25330.80
Mels C et al. (2013) The link between vascular deterioration and branched chain amino acids in a population with high glycated haemoglobin: the SABPA study. Amino Acids 45:1405–1413. doi:10.1007/s00726-013-1611-0
Mels CM et al. (2014) 8-Oxo-7, 8-dihydro-2′-deoxyguanosine, reactive oxygen species and ambulatory blood pressure in African and Caucasian men: the SABPA study. Free Radic Res 48:1291–1299. doi:10.3109/10715762.2014.951840
Noronha-Dutra AA, Epperlein MM, Woolf N (1993) Reaction of nitric oxide with hydrogen peroxide to produce potentially cytotoxic singlet oxygen as a model for nitric oxide-mediated killing. FEBS Lett 321:59–62. doi:10.1016/0014-5793(93)80621-Z
Pedrinelli R, Dell’Omo G, Penno G, Mariani M (2001) Non-diabetic microalbuminuria, endothelial dysfunction and cardiovascular disease. Vasc Med 6:257–264. doi:10.1177/1358836X0100600410
Puchau B, Zulet MA, Urtiaga G, Navarro-Blasco Í, JA M (2009) Asymmetric dimethylarginine association with antioxidants ntake in healthy young adults: a role as an indicator of metabolic syndrome features. Metabolism 58:1483–1488. doi:10.1016/j.metabol.2009.04.037
Reimann M et al. (2013) Effects of acute and chronic stress on the L-arginine nitric oxide pathway in black and white South Africans: the sympathetic activity and ambulatory blood pressure in Africans study. Psychosom Med 75:751–758. doi:10.1097/PSY.0b013e3182a3e465
Schutte AE et al. (2010) Dimethylarginines: their vascular and metabolic roles in Africans and Caucasians. Eur J Endocrinol 162:525–533. doi:10.1530/EJE-09-0865
Schutte R et al. (2011) Arterial stiffness, ambulatory blood pressure and low-grade albuminuria in non-diabetic African and Caucasian men: the SABPA study. Hypertens Res 34:862–868. doi:10.1038/hr.2011.45
Schutte R et al. (2009) Blood glutathione and subclinical atherosclerosis in African men: the SABPA study. Am J Hypertens 22:1154–1159. doi:10.1038/ajh.2009.158
Schwedhelm E, Tan-Andresen J, Maas R, Riederer U, Schulze F, RH B (2005) Liquid chromatography–tandem mass spectrometry method for the analysis of asymmetric dimethylarginine in human plasma. Clin Chem 51:1268–1271. doi:10.1373/clinchem.2004.046037
Siegerink B et al. (2013) Asymmetric and symmetric dimethylarginine and risk of secondary cardiovascular disease events and mortality in patients with stable coronary heart disease: the KAROLA follow-up study. Clin Res Cardiol 102:193–202. doi:10.1007/s00392-012-0515-4
Śladowska-Kozłowska J et al. (2012) Oxidative stress in hypertensive children before and after 1 year of antihypertensive therapy. Pediatr Nephrol 27:1943–1951. doi:10.1007/s00467-012-2193-x
Sliwa K, Wilkinson D, Hansen C, Ntyintyane L, Tibazarwa K, Becker A, Stewart S (2008) Spectrum of heart disease and risk factors in a black urban population in South Africa (the Heart of Soweto Study): a cohort study. Lancet 371:915–922. doi:10.1016/S0140-6736(08)60417-1
Sözmen B, Kazaz C, TASKIRAN D, Aslan L, Akyol A, EY S (1998) Plasma antioxidant status and nitrate levels in patients with hypertension and coronary heart disease. Turk J Med Sci 28:525–532
Tousoulis D, A-M K, Tentolouris Nikolaos Papageorgiou C, Stefanadis C (2012) The role of nitric oxide on endothelial function. Curr Vasc Pharmacol 10:4–18. doi:10.2174/157016112798829760
Tsioufis C et al. (2010) ADMA, C-reactive protein, and albuminuria in untreated essential hypertension: a cross-sectional study. Am J Kidney Dis 55:1050–1059. doi:10.1053/j.ajkd.2009.11.024
Vásquez-Vivar J et al. (1998) Superoxide generation by endothelial nitric oxide synthase: the influence of cofactors. Proc Natl Acad Sci 95:9220–9225
Wang D, Strandgaard S, Iversen J, Wilcox CS (2009) Asymmetric dimethylarginine, oxidative stress, and vascular nitric oxide synthase in essential hypertension. Am J Physiol Regul Integr Comp Physiol 296:R195–R200. doi:10.1152/ajpregu.90506.2008
Acknowledgments
The SABPA study would not have been possible without the voluntary collaboration of the participants and the Department of Education, North-West Province, South Africa. We gratefully acknowledge the technical assistance of Mariola Kastner, Anna Steenpass, Mrs. Tina Scholtz, Dr. Szabolcs Péter, and Sr Chrissie Lessing. This study was supported by the National Research Foundation; the National Research Foundation Thuthuka (80643); the Medical Research Council, the North-West University, Potchefstroom; Roche Products (Pty) Ltd., South Africa, and the Metabolic Syndrome Institute, France.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflict of interest to declare. Any opinion, findings, and conclusions or recommendations expressed in this material are those of the authors and therefore the National Research Foundation does not accept any liability in regard thereto.
Electronic supplementary material
Table S1
(DOCX 14 kb)
About this article

Cite this article
Mels, C.M.C., Huisman, H.W., Smith, W. et al. The relationship of nitric oxide synthesis capacity, oxidative stress, and albumin-to-creatinine ratio in black and white men: the SABPA study. AGE 38, 9 (2016). https://doi.org/10.1007/s11357-016-9873-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11357-016-9873-6