
Abstract
Lipids produced from agricultural and industrial residues using oleaginous microorganisms for use as biofuels are attracting the attention of researchers due to their environmental benefits. However, low efficiencies and high costs limit their application to a certain extent. The present study is the first to use inositol as an enhancer to improve the production and accumulation of lipids during fermentation by the microalga Schizochytrium sp. SR21. The study aimed to maximize the production of lipids and docosahexaenoic acid (DHA) by optimizing the conditions of inositol addition into the fermentation medium. The corresponding key enzyme and metabolite profiles of SR21 were evaluated. The results indicated that the addition of 120 mg L−1 of inositol to the medium at 48 h improved lipid and DHA production by 13.90 and 20.82%, resulting in total concentrations of 22.86 and 8.53 g/L, respectively. Moreover, the ratio of unsaturated fatty acids (UFAs) to saturated fatty acids (SFAs) increased by 23.38% and is consistent with the results of the metabolomic analysis. The activity of enzymes (i.e., PC, G6PDH, NADPH-ME, and ACL) related to fatty acid synthesis in strain SR21 also increased significantly (43.38%, 28.68%, 37.47%, and 19.87%, respectively). Metabolomic analysis also showed that inositol promoted lipid synthesis in SR21 and significantly increased the relative proportion of UFAs by affecting the citrate cycle and SFA and UFA metabolic pathways. Thus, inositol is an ideal enhancer of lipid production and accumulation by oleaginous microorganisms.

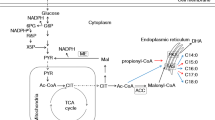
Graphical abstract
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Energy crises and environmental effects of petroleum fuels have driven people to develop alternative fuel sources. Lipid production from agricultural and industrial residues using oleaginous microorganisms has attracted interest due to its many advantages, especially those related to environmental protection. In recent years, marine microbes have emerged as an alternative source for the production of high-quality lipids at a low cost (Garay et al. 2014). Considerable studies have focused on developing techniques for the production of high-value lipids and metabolites for use as biofuels and for nutrition and health (Ratledge 1992). Docosahexaenoic acid (DHA) belongs to the omega-3 group of fatty acids, which is important for various physiological functions in the human body (Brennan and Owende 2010; Taoka et al. 2011). DHA may have beneficial effects in the prevention of diseases, such as high blood pressure, thrombosis, arthritis, depression, and some cancers (Danaei et al. 2009; Janssen and Kiliaan 2014; Mohajeri et al. 2015). Historically, marine fish oil has been regarded as the primary source of commercial DHA, but environmental pollution and fishing restrictions directly influence the quantity and quality of DHA. Single-cell oils (SCOs) do not have the disadvantages of fish oils and their production can be readily scaled up using biotechnological methods. Among the oleaginous microorganisms, Schizochytrium limacinum SR21, a heterotrophic marine microorganism, can produce lipids with a high percentage of DHA (Yokochi et al. 1998; Morita et al. 2006). Multiple studies have demonstrated the safety of DHA-rich lipids produced by Schizochytrium sp. (Kroes et al. 2003; Fedorova-Dahms et al. 2011). However, the low bioconversion rate to DHA is a major problem. Thus, extensive research is currently focused on reducing the costs of microbial oil production.
To improve the feasibility of DHA production from single-celled microorganisms, efficient approaches are required. Few studies have identified enhancers that can stimulate microalgae growth and DHA yield (Fig. 1). Phytohormones and their derivatives also play an important role in many metabolic pathways. Gibberellin and 6-benzylaminopurine have been proven to enhance lipid and DHA production in Aurantiochytrium sp. (Adarme-Vega et al. 2012). Addition of gibberellin (4 mg/L) caused increases in biomass, lipid, and DHA production of up to 14.4%, 43.6%, and 79.1%, respectively, when compared to that in the control group (without gibberellin). Similarly, lipid and DHA yields were significantly enhanced by 6-BAP treatment (3 mg/mL), as observed at 120 h of fermentation (Yu et al. 2016a, b). Besides, Taoka et al. (2011) have reported that 1% Tween-80 supplement in the culture medium can induce a 2-fold increase in biomass and a 1.15-fold increase in lipid content. In addition, Zhang et al. (2016) found that DHA production increased by 47% and reached a concentration of 5.51 g/L when 4 g/L malate was added during the rapid lipid accumulation stage in shake flasks. In Schizochytrium sp. B4D1 strains, some polyols, such as myo-inositol, were also reported to greatly contribute to environmental stress responses. Inositol, an important biological growth factor, was found to modify the fatty acid composition and dry cell weight of Schizochytrium sp. during fermentation (Li et al. 2013). Previous studies from our lab had extensively investigated and discussed the effect of fermentation using defatted silkworm pupae hydrolysate on lipid and DHA yields (Shi et al. 2017). To achieve high productivity with low cost, it is vital to find a new enhancer to improve lipid accumulation and production in the large-scale cultivation of Schizochytrium sp.

Biomass and DHA yield by the strains Schizochytrium and Aurantiochytrium in shake flasks. Gibberellin, 6-BAP, Tween-80, and malate are added at concentrations of 4 mg/L, 3 mg/mL, 1%, and 4 g/L, respectively
DHA has great application prospects in the field of medicine and health and its biosynthesis in microbial cells is extremely complicated (Song et al. 2014). The traditional fatty acid synthesis (FAS) system and the polyketide synthase (PKS) pathway are generally used to synthesize DHA in microbial cells (Metz et al. 2001). In the FAS system, two saturated fatty acids (SFAs), either C16:0 or C18:0, are synthesized after which these fatty acids undergo a series of elongations and desaturations (condensation, reduction, dehydration, and re-reduction) to produce unsaturated fatty acids (UFAs) and polyunsaturated fatty acids (PUFAs) (Ratledge 2004). The transformation of C22:5 to C22:6 (DHA) is catalyzed by the enzyme delta-4 desaturase in this pathway. In the PKS pathway, acyl carrier protein (ACP), produced by coenzyme A (CoA) is used as a covalent attachment point for chain elongation, which is carried out in reiterative cycles. Enzymes such as enoyl reductase (ER), dehydrase/isomerase (DH), 3-ketoacyl-ACP reductase (KR), and 3-ketoacyl synthase (KS) are necessary for the production of fatty acids through the PKS pathway (Yuanmei et al. 2018). In-depth studies of lipid metabolism pathways in microbial cells are critical for the improvement of microbial lipid synthesis efficiency. Metabolomic strategies, therefore, provide an opportunity for metabolic phenotyping of marine microorganism and permit the investigation of influences of enhancers and interactions with the microbe (Xia et al. 2009).
In our study, inositol was used as a fermentation enhancer to promote the lipid yield by Schizochytrium sp. Furthermore, the effects of inositol on growth, lipid, and DHA production of SR21 were studied. Epifluorescence micrographs of SR21, stained with Nile red after 5 days of cultivation, were generated. Moreover, the effect of inositol on the levels of the lipogenic enzymes in SR21 at different time points during fermentation was investigated. Until now, there have been no studies involving the use of inositol for enhancing the production of DHA and lipid in SR21. Thus, the aim of our study was to investigate the effects of inositol on cell growth, lipid accumulation, fatty acid composition, and, particularly, DHA production in SR21.
Materials and methods
Strains and cultures
Schizochytrium sp. SR21 (ATCC MYA-1381) was purchased from the American Type Culture Collection (ATCC) and was maintained on a medium consisting of 5 g L−1 glucose, 1 g L−1 yeast extract, 1 g L−1 tryptone, 20 g L−1 agar, and 30 g L−1 artificial sea salt. The algal inoculum medium comprises 20 g L−1 glucose, 4 g L−1 yeast extract, 4 g L−1 tryptone, and 40 g L−1 artificial sea salt. All chemical reagents were procured from Shanghai Chengjie Chemical Co., Ltd. (Shanghai, China), and enzyme activity test kits were purchased from Beijing Suolaibao Technology Co. Ltd. (Beijing, China). The inoculum medium was cultured on an orbital shaker (160 rpm) at 25 °C for 24 h and was inoculated at 10% (v/v). The fermentation medium consists of 100 g L−1 glucose, 5 g L−1 yeast extract, 5 g L−1 tryptone, and 40 g L−1 artificial sea salt. The shake flask was placed on an orbital shaker at 25 °C and 160 rpm for 5 days. Batch cultivation was performed in a BioFlo/CelliGen 115 reactor (Eppendorf, Hamburg, Germany) with a total volume of 3 L at 25 °C, dissolved oxygen (DO) at 20%, aeration rate at 1.25 VVM, pH at 6.5, and working volume of 1.5 L.
Inositol treatment effect on SR21 growth
The effects of the initial inositol concentration on SR21 growth and lipid production were investigated by the addition of different amounts of inositol to the culture medium to reach a final concentration of the range 0–200 mg L−1 and medium without inositol served as the control group.
The effects of different inositol feeding times on algae growth and DHA productivity were also measured by adding inositol to the media during the cultivation phase at 5 different incubation times (when fermentation time reached 0, 24, 48, 72, 96, and 120 h). The fermentation time was 120 h, and each group was sampled every 24 h. The analysis of biomass, lipid content, fatty acid profile, and metabolic profile was carried out through the methods described below.
Analysis of culture supernatant
Concentration of glucose in the fermentation broth was measured using a bioanalyzer (SBA-40C, Institute of Biology, Shandong Academy of Sciences, China) (Xu and Xu 2014).
Biomass assay
The biomass was measured by centrifuging the cell suspension at 7000g, the cells were washed twice with millipore water and dried overnight with a freeze dryer (Lyoquest-55 Telstar). Biomass is expressed as dry cell weight (DCW).
Extraction and fatty acid analysis of lipids
Freeze-dried cells (1 g) were re-suspended in 10 mL 20% HCl (20 g of HCl in 100 mL of H2O) for 30 min assisted by ultrasonic irradiation and then centrifuged for 10 min at 6000 rpm at 4 °C. The process was repeated 2–3 times and then the supernatants were collected and evaporated in a vacuum furnace to dry it to a constant weight. Subsequently, the total fatty acids (TFAs) were measured on the basis of DCW.
A two-step methylation process was applied for the determination of fatty acid composition in S. limacinum SR21 (Bowyer et al. 1963).
Enzyme assays and protein estimation
Five key lipogenic enzymes were determined during the lipogenic phase of cell culture (at 48, 72, 96, and 120 h), ATP-citrate lyase (ACL), lipase, glucose-6-phosphate dehydrogenase (G-6-PDH), and NADP-malic enzyme (NADP-ME). SpectraMax® iD3 Multi-Mode Microplate Reader (Molecular Devices, San Jose, CA, USA) was employed to measure the activity of the enzymes at 25 °C.
The protein concentration was estimated from the supernatant using the Bradford method with fat-free BSA as a reference standard. All the enzyme and protein assays were done in triplicates.
Microstructure epifluorescence micrograph of SR21
SR21 cells were stained with Nile red for the detection of total cellular lipids (fluorescent yellow). Epifluorescence micrographs of the stained cells were taken on the 5th day of cultivation as reported by Bradford MM (Bradford 1976).
Metabolomic analysis of samples
Internal standard (20 μL) was added to the sample, then 1 mL of 4:1 methanol-water (v/v) was added to a glass vial, mixed with 200 μL chloroform and the supernatant was carefully removed with a pipette. The liquid was transferred to a centrifuge tube, ultrasonicated for 15 min, and then centrifuged for 15 min (10,000g, 4 °C). Next, 1 mL of the supernatant was transferred to a 1.5-mL centrifuge tube and the liquid was allowed to evaporate. Then 250 μL of methanol-water (7:3, v/v) solution was added and the mixture was vortexed for 30 s and sonicated for 2 min. It was then centrifuged for 15 min (10,000g, 4 °C) and 180 μL of the supernatant was filtered through a 0.22-μm organic phase pin filter (Nantong Feiyu Biological Technology Co., Ltd., Nantong, China). The filtered supernatant was transferred to an LC injection vial and stored at − 80 °C until LC-MS analysis was performed. Preparation of quality control (QC) samples was done by mixing equal volumes of the extracts of all the samples. Each QC had the same volume as the sample. All extraction reagents were pre-cooled at − 20 °C before usage.
Waters ACQUITY UPLC System (Waters Corp. Milford, USA) was employed to perform chromatographic analysis. The samples were run on an ACQUITY UPLC BEH C18 (100 mm × 2.1 mm, 1.7 μm). The columns were eluted at a flow rate of 0.45 mL/min and maintained at 45 °C. The mobile phase for samples consisted of water (A) and acetonitrile (B), each containing 0.1% formic acid. The gradient program was optimized as follows: 0–2 min, 5% B to 20% B; 2–4 min, 20% B to 60% B; 4–11 min, 60% B to 100% B; 11–13 min, washing with 100% B, and 13.5–14.5 min, equilibration with 5% B.
Statistical analysis
All cultivations were conducted in duplicates. All statistical calculations were done at least in duplicates and the final values were expressed as mean ± standard deviation. Analysis of variation (ANOVA) of single factors was carried out to analyze the variance of results with significance levels above p < 0.05.
Results and discussion
Effects of inositol on biomass, lipid, and DHA production in SR21
In this study, SR21 was grown in a culture medium containing different concentrations of inositol, ranging from 0 to 200 mg L−1 for 120 h. There was no significant impact of the different inositol concentrations on cell growth and biomass (Fig. 2a). However, lipid and DHA accumulation of SR21 was enhanced upon the addition of inositol to the culture medium. Increased amounts of lipid and DHA were observed concomitantly with an increase in inositol concentration from 0 to 120 mg L−1. The lipid content decreased slightly and DHA levels increased at inositol concentrations of 120 up to 200 mg L−1. The peak production of lipid and DHA (56.03% biomass and 7.99 g L−1 respectively) were achieved when 120 mg L−1 of inositol was added to the culture medium. Compared to the control sample (no inositol), supplementation with inositol resulted in 7.58% and 13.17% increase in lipid and DHA production, respectively. In contrast, final yield of DHA reaches 5.39 and 4.6 g L−1 only when gibberellin and 6-benzylaminopurine were added to Aurantiochytrium sp. respectively (Yu et al. 2016a, b). Our results indicate that inositol is an excellent enhancer, which can greatly improve the accumulation of lipids and increase DHA yield in SR21.

Effects of different concentrations of inositol and different feeding times on biomass, lipid content, and DHA yield in Schizochytrium sp. SR21. (a different concentrations of inositol; b different feeding time of inositol). All data are represented as means of three replicates; vertical error bars represent the standard error of the mean
Lipid and DHA production of SR21 was significantly different from the control when 120 mg L−1 inositol was added to the fermentation medium at different time points (0 to 96 h) (Fig. 2b). Lipid and DHA production had the steepest increase from 0 to 48 h and the maximum DHA yield (8.53 g L−1) was achieved with inositol when the fermentation time was 48 h. The results indicate that the lipid content and DHA yield of SR21 were 13.90 and 20.82% higher than that of the control and reached 22.86 and 8.53 g L−1 respectively (with 120 mg L−1 inositol in the medium at 48 h). Thus, the addition of inositol at 48 h not only considerably improves lipid content but also increases DHA production.
The effects of inositol on biomass, lipid, and DHA production were monitored throughout the fermentation process of SR21. Figures 3b and c show that both lipid and DHA production (with inositol) were significantly higher than those of the control group. However, inositol has little impact on the biomass of SR21 (Fig. 3a). This is the first report of inositol promoting fatty acid accumulation in marine DHA-producing Schizochytrium sp. Thus, increased DHA yields of Schizochytrium sp. can be achieved by adding inositol to the culture medium.

a Biomass, b lipid content, c and DHA yield in Schizochytrium sp. SR21 with or without inositol addition at 48 h. All data are represented as means of three replicates; vertical error bars represent the standard error of the mean
In order to verify that the addition of inositol to the medium promotes the synthesis and accumulation of intracellular lipids in the fermentation process of SR21, epifluorescence micrographs of cells stained with Nile red for the detection of total cellular lipids (fluorescent yellow) were done at 96 h of cultivation. As shown in Fig. 4, the fluorescent yellow of SR21 cells was found to be more intense in the inositol supplemented samples compared to the controls. At the end of 96 h of fermentation (Fig. 4b), the cells cultivated in inositol medium (added at 48 h) had the highest intensity of the lipid staining dye (Fig. 4c). Therefore, this further indicates that inositol can enhance lipid accumulation in Schizochytrium sp.

Epifluorescence micrographs of Schizochytrium sp. SR21 cells stained with Nile red. For detection of total cellular lipids, the intensity of fluorescent yellow was measured on day 5 of the culture period. The strain was cultivated on a medium without inositol; b medium with inositol, added before cultivation; and c medium with inositol supplementation at 48 h. Culture conditions: initial glucose concentration 120 g L−1; DO 20%; incubation temperature 26 °C
Effect of inositol on fatty acid composition
The production of PUFAs by marine or freshwater microalgae has attracted increased attention as they are important components of fatty acids, whose composition is an important indicator for evaluating edible oil products. In the present study, the fatty acid content in different experimental groups was identical, and the main components of SR21 fatty acids included myristic acid (C14:0), palmitic acid (C16:0), docosapentaenoic acid (C22:5), and DHA (C22:6) (Table 1). After adding inositol to the culture medium, the composition of SR21 fatty acids, including UFAs, such as linolenic acid (C18: 3), docosapentaenoic acid (C20: 5, EPA), and DHA (C22: 6), increased to varying degrees compared to the control group. Moreover, the percentage reductions of myristic acid (C14:0) and palmitic acid (C16:0) were most obvious among those of all SFAs, while the percentage increase of DHA (C22:6) was most prominent among those of all UFAs. Therefore, the results showed that the addition of inositol to the medium partly changes the saturation of fatty acids in the SR21 lipids and increases the levels of UFAs in the cells, mainly those of PUFAs. These are the key determinants of whether lipids can be used as high-quality edible oils or fuel feedstock (Ratledge 2014).
Naturally, PUFAs in various microbial cells are synthesized by enzymes, such as polyketide synthases (PKS) or fatty acid synthases (FAS) (Ratledge 2012). In this study, the content of microbial lipids increased after the addition of inositol to the medium and this may be due to inositol stimulating either PKS or FAS in the microbial cells. This eventually led to an increase in the accumulation of intracellular lipids, and the ratio of UFAs/SFAs was augmented by 23.38%.
Effect of inositol on lipid-related enzyme activity in SR21
Inositol was added to the culture medium at a final concentration of 120 mg L−1 before the fermentation of SR21. The concentration of residual sugar, cell biomass, and oil production in the medium was monitored every 12 h to map the growth of microalgae and the yield of lipids during fermentation (Fig. 5). As shown in Fig. 6, after the addition of inositol to the culture medium, the growth curve of the microbial cells is sigmoidal. The cells enter a logarithmic growth phase rapidly after the start of fermentation and grow exponentially between 12 and 72 h. The glucose content of the media reduces with cell growth. Accumulation of lipids occurs between 24 and 108 h and the cells enter a stationary phase after 108 h.

Changes in Schizochytrium sp. SR21 biomass (squares), residual sugar (triangle), DHA yield (circle), and lipid content (inverted triangle) in batch fermentation are shown. Culture conditions: initial glucose concentration 120 g L−1; inositol concentration 120 mg L−1, DO 20%; incubation temperature 26 °C

Specific activity (nmol∙mg−1∙min−1) of the enzymes in vitro at different cultivation stages of Schizochytrium sp. SR21 and different treatments: Control without inositol (black line with white boxes), with inositol being added before the culture at 0 h (black line with white circles) and with inositol being supplemented at 48 h in a shake flask (black line with white triangles). aPC, pyruvate carboxylase; bNADH-ME, NADP-malic enzyme; cICDHm, isocitrate dehydrogenase mitochondrial; dG6PDH, glucose-6-phosphate dehydrogenase; eACL, ATP-citrate lyase. Different letters mean significant difference at p < 0.05
In order to reveal the mechanism of inositol promoting lipid and DHA production by SR21, five key enzymes of lipogenesis were analyzed under different culture stages of Schizochytrium sp. (Fig. 6). As shown in Fig. 6a, pyruvate carboxylase (PC) increases during the lipid accumulation phase. Moreover, the addition of inositol to the medium resulted in an increase in PC activity and peaked when inositol was added during the fermentation process after 48 h. This resulted in a 43.38% activity increase compared to the control group at 96 h of fermentation. PC plays an important role in the oxaloacetate cycle (OAA); oxaloacetate is an important intermediate in the synthesis of acetyl-CoA and NADPH. Wang et al. (2015) reported that the overexpression of PC in Yarrowia lipolytica resulted in a significant increase in intracellular lipid content. Thus, the increase in lipid content in SR21 may be due to inositol-related enhancement of intracellular PC activity.
The reaction of oxidative decarboxylation of malic acid in cells catalyzed by NADP-malic enzyme (NADP-ME) is reversible (Conn et al. 1949). As shown in Fig. 6b, the NADP-ME activity increased continuously between 48 and 72 h and then decreased significantly. The specific activity (847.68 U mg−1) of NADP-ME was highest in the inositol group at 48 h. During the lipid synthesis period (48–96 h), especially between 72 and 96 h, the NADP-ME activity of the inositol group was significantly higher than that of the control group. Compared to the control group, the NADP-ME activity of the experimental group (addition of inositol at 48 h) increased by 37.47%, at 72 h of fermentation. However, after 72 h, the activity of NADP-ME decreased rapidly, which may be due to depletion of nitrogen source in the medium (Rosa et al. 2010). In addition, as shown in Fig. 6c, the specific activities of isocitrate dehydrogenase, mitochondrial (ICDHm, EC 1.1.1.41) in the three treatment groups all showed a downward trend, and the addition of inositol did not significantly affect the activity of ICDHm. ICDHm is a key enzyme involved in the TCA cycle. During the culture period of 48 to 120 h, the concentration of glucose in the medium continuously decreased, resulting in insufficient raw materials for the TCA cycle and the activity of ICDHm continuously decreased, weakening the metabolism.
In the lipid synthesis process of eukaryotic microbial cells, the supply of NADPH is mainly related to NADP-ME and the enzymes in the pentose phosphate pathway (HMP), such as glucose-6-phosphate dehydrogenase (G6PDH) and 6-phosphate glucose acid dehydrogenase (6PGDH) (Ratledge 2014). As shown in Fig. 6d, the activity of G6PDH in SR21 showed an upward trend during the fermentation between 48 and 96 h, and the G6PDH activity of the experimental group (addition of inositol at 48 h) increased by 28.68% compared with the control group at 96 h of cultivation. The results indicate that NADPH is mainly synthesized from HMP and inositol can enhance the activity of G6PDH, thereby promoting the synthesis of intracellular lipids. In addition, the ATP-citrate lyase (ACL) is a key rate-limiting enzyme in lipid synthesis in oil-producing microbial cells. As shown in Fig. 6e, the ACL activity of the samples with inositol was higher than that of the control group. The ACL activity of the experimental group (addition of inositol at 48 h) increased by 19.87% compared with the control group at 72 h of fermentation. The specific activity of ACL is as high as 100.29 U mg−1, which is higher than the ACL activity (80 U/mg) of Y. lipolytica (Ochoa-Estopier and Guillouet 2014). Thus, due to high specific activity of ACL, Schizochytrium sp. has a stronger acetyl-CoA synthesis capacity and higher lipid production than other oil-producing microorganisms.
As shown in Figs. 5 and 7, the ME activity is high at the mid-stage (72 h) of fermentation. The G6PDH activity gradually increases between 48 and 96 h of fermentation until the activity peaks at the mid-stage of the fermentation. This phenomenon indicates that the production of NADPH at the early stage of fermentation is mainly related to ME and that the late stage of fermentation is mainly related to G6PDH. Metabolic pathway map shows that the higher the activity of G6PDH is, the faster the HMP pathway proceeds, to eventually produce more NADPH. Similarly, highly active NADP-ME also produces more NADPH through the hydrogenase cycle system. In addition, high activity of ACL promotes the formation of acetyl-CoA. Adequate NADPH and acetyl-CoA levels are essential for large-scale synthesis of fatty acids, which ultimately leads to inositol-based enhancement of lipid production levels in Schizochytrium sp.

Metabolic pathway of fatty acid biosynthesis and the related key enzymes in Schizochytrium sp. SR21
Metabolic profiles of SR21 grown in inositol medium
Metabolic profiles of samples (with inositol addition after 48 h or without inositol addition) were generated by LC/MS in positive and negative ion mode. Figure 8a shows a principal component analysis (PCA) model employed to discriminate between the tested groups. The distribution of QC was very concentrated, indicating parallelism between the samples. Although one treatment sample deviated from the confidence interval, its data can be eliminated without affecting the quality of the results. To achieve better visualization of the discriminated sample groups from PCA, a supervisory protocol was chosen. The metabolic patterns of SR21 with and without inositol treatment were evaluated by partial least squares-discriminant analysis (PLS-DA). There was no complete separation between the two treatment groups and control groups in the PCA analysis and this might be due to individual differences. It can be seen that the parallelism between the samples is very good and stable based on the QC distribution and the PLS distribution. It is clear from the PLS-DA (Fig. 8b) analysis that the metabolic pathway of SR21 with inositol addition is different from that of the control group, suggesting that inositol induces significant biochemical changes in SR21.

a PCA score plot of samples collected from different treatment groups of Schizochytrium sp. SR21 (R2X = 0.572, Q2 (cum) = 0.207); b PLS-DA score plot of samples collected from different treatment groups of SR21 (R2X = 0.598, R2Y = 0.869, Q2 (cum) = 0.690); c, d OPLS-DA score plot and S-Plot of control and inositol-added group. (C) control group, (T) treatment group
Identification of potentially different metabolites associated with inositol
Figure 8c revealed a sharp separation between the inositol group and the control group in PLS-DA employed by the orthogonal to partial least squares-discriminant analysis (OPLS-DA). The S-plot and variable important in projection (VIP) were adopted to select potentially different metabolites (Fig. 8d). Compounds far from the origin with a VIP > 1 are considered to be potentially different metabolites responsible for the metabolic profile of inositol-added strain SR21. As shown in Fig. 9, the p values of fatty acids and UFAs exceed 0.01 and 0.05, respectively, which demonstrates that the addition of inositol causes a significant increase in the fatty acid biosynthesis of SR21. In addition, the metabolite analysis shows that the expression levels of 44 lipids, including 10 SFAs and 34 UFAs, in the experimental group, were upregulated compared to the control group and their retention time, m/z, VIP, and F/C values are shown in Table S1. As shown in Fig. S1, inositol may inhibit the activity of palmitoyl-CoA hydrolase in the biosynthesis pathway of UFAs (EC 3.1.2.2), leading to a decrease of palmitic acid and stearic acid synthesis, which ultimately leads to the accumulation of C16:0 and C18:0 content, thus providing more precursors for the synthesis of UFAs (López-Ferrer et al. 2001). Hence, SFAs can form UFAs under the catalysis of fatty acyl desaturase and carbon chain elongase (Moon et al. 2001). In addition, Fig. S2 indicates that the addition of inositol may inhibit the progression of the tricarboxylic acid cycle, which reduces the consumption of acetyl-CoA (Ratledge 2004). Large amounts of acetyl-CoA can contribute to the synthesis of lipids, eventually leading to increased fatty acid synthesis. Besides, in the biosynthesis pathway of fatty acids, the amount of SFAs, including tetradecanoic acid, hexadecanoic acid, and octadecanoic acid, was lowered in SR21 in the inositol group (Fig. S3). This might be the reason for the upward trend of the UFA fraction of the total lipid.

Metabolic pathway enrichment map: The portion of p < 0.05 in the metabolic pathway, with the metabolic pathway name as the abscissa and the –log (p value) as the ordinate for the T/C metabolic pathway enrichment map. The red line indicates a p value of 0.01 and the blue line indicates a p value of 0.05. Signaling pathways represented below the blue or above the red line are significant
Conclusions
This is the first report of inositol enhancing DHA and lipid production in Schizochytrium sp. SR21. It was found that 120 mg L−1 of inositol added to the fermentation medium at 48 h results in a lipid and DHA yield of 22.86 g L−1 and 8.53 g L−1, which are 13.90% and 20.82% higher than those of the control group, respectively. In addition, the content of UFAs in the lipid fraction increased significantly. It was found that inositol enhanced the activities of PC, G6PDH, NADP-ME, and ACL during the lipid accumulation stage. Moreover, the metabolomic analysis confirmed that inositol can enhance lipid accumulation in Schizochytrium sp. SR21 and can change fatty acid composition. Inositol can be used as an enhancer of SR21 fermentation and has broad applications in the field of bioenergy. However, the detailed mechanism of the inositol mediated enhancement of lipid synthesis in strain SR21 requires further research.
References
Adarme-Vega TC, Lim DK, Timmins M, Vernen F, Li Y, Schenk PM (2012) Microalgal biofactories: a promising approach towards sustainable omega-3 fatty acid production. Microb Cell Factories 11:96
Bowyer DE, Leat WM, Howard AN, Gresham GA (1963) The determination of the fatty acid composition of serum lipids separated by thin-layer chromatography; and a comparison with column chromatography. Biochim Biophys Acta 70:423–431
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Brennan L, Owende P (2010) Biofuels from microalgae-a review of technologies for production, processing, and extractions of biofuels and co-products. Renew Sust Energ Rev 14:557–577
Conn E, Vennesland B, Kraemer LM (1949) Distribution of a triphosphopyridine nucleotide-specific enzyme catalyzing the reversible oxidative decarboxylation of malic acid in higher plants. Arch Biochem 23:179–197
Danaei G, Ding EL, Mozaffarian D, Taylor B, Rehm J, Murray CJL, Ezzati M (2009) The preventable causes of death in the United States: comparative risk assessment of dietary, lifestyle, and metabolic risk factors. PLoS Med 6:e1000058
Fedorova-Dahms I, Marone PA, Bauter M, Ryan AS (2011) Safety evaluation of DHA-rich algal oil from Schizochytrium sp. Food Chem Toxicol 49:3310–3318
Garay LA, Boundymills KL, German JB (2014) Accumulation of high-value lipids in single-cell microorganisms: a mechanistic approach and future perspectives. J Agric Food Chem 62:2709–2727
Janssen CI, Kiliaan AJ (2014) Long-chain polyunsaturated fatty acids (LCPUFA) from genesis to senescence: the influence of LCPUFA on neural development, aging, and neurodegeneration. Prog Lipid Res 53:1–7
Kroes R, Schaefer EJ, Squire RA, Williams GM (2003) A review of the safety of DHA45-oil. Food Chem Toxicol 41:1433–1446
Li J, Ren LJ, Sun GN, Qu L, Huang H (2013) Comparative metabolomics analysis of docosahexaenoic acid fermentation processes by Schizochytrium sp. under different oxygen availability conditions. OMICS 17:269–281
López-Ferrer S, Baucells MD, Barroeta AC, Galobart J, Grashorn MA (2001) n-3 Enrichment of chicken meat. 2. Use of precursors of long-chain polyunsaturated fatty acids: linseed oil. Poult Sci 80:753–761
Metz JG, Roessler P, Facciotti D, Levering C, Dittrich F, Lassner M et al (2001) Production of polyunsaturated fatty acids by polyketide synthases in both prokaryotes and eukaryotes. Science 293:290–293
Mohajeri MH, Troesch B, Weber P (2015) Inadequate supply of vitamins and DHA in the elderly: implications for brain aging and Alzheimer-type dementia. Nutrition 31:261–275
Moon YA, Shah NA, Mohapatra S, Warrington JA, Horton JD (2001) Identification of a mammalian long chain fatty acyl elongase regulated by sterol regulatory element-binding proteins. J Biol Chem 276:45358–45366
Morita E, Kumon Y, Nakahara T, Kagiwada S, Noguchi T (2006) Docosahexaenoic acid production and lipid-body formation in schizochytrium limacinum SR21. Mar Biotechnol 8:319–327
Ochoa-Estopier A, Guillouet SE (2014) D-stat culture for studying the metabolic shifts from oxidative metabolism to lipid accumulation and citric acid production in Yarrowia lipolytica. J Biotechnol 170:35–41
Ratledge C (1992) Microbial lipids: Commercial realities or academic curiosities. Industrial Applications of Single Cell Oils. AOCS Press
Ratledge C (2004) Fatty acid biosynthesis in microorganisms being used for single cell oil production. Biochimie 86:807–815
Ratledge C (2012) Omega-3 biotechnology: errors and omissions. Biotechnol Adv 30:1746–1747
Ratledge C (2014) The role of malic enzyme as the provider of NADPH in oleaginous microorganisms: a reappraisal and unsolved problems. Biotechnol Lett 36:1557–1568
Rosa SM, Soria MA, Vélez CG, Galvagno MA (2010) Improvement of a two-stage fermentation process for docosahexaenoic acid production by Aurantiochytrium limacinum SR21 applying statistical experimental designs and data analysis. Bioresour Technol 101:2367–2374
Shi XY, Li TY, Wang M, Wu WW, Li WJ, Wu QY, Wu FA, Wang J (2017) Converting defatted silkworm pupae by Yarrowia lipolytica for enhanced lipid production. Eur J Lipid Sci Technol 119:1600120
Song Y, Salbu B, Teien HC, Heier LS, Rosseland BO, Høgåsen T et al (2014) Hepatic transcriptomic profiling reveals early toxicological mechanisms of uranium in Atlantic salmon (Salmo salar). BMC Genomics 15:1–21
Taoka Y, Nagano N, Okita Y, Izumida H, Sugimoto S, Hayashi M (2011) Effect of tween 80 on the growth, lipid accumulation and fatty acid composition of Thraustochytrium aureum ATCC 34304. J Biosci Bioeng 111:420–424
Wang GY, Zhang Y, Chi Z, Liu GL, Wang ZP, Chi ZM (2015) Role of pyruvate carboxylase in accumulation of intracellular lipid of the oleaginous yeast Yarrowia lipolytica ACA-DC 50109. Appl Microbiol Biotechnol 99:1637–1645
Xia JF, Liang QL, Ping HU, Wang YM, Luo GA (2009) Recent trends in strategies and methodologies for metabonomics. Fresenius J Anal Chem 37:136–143
Xu K, Xu P (2014) Betaine and beet molasses enhance L-lactic acid production by Bacillus coagulans. PLoS One 9:e100731
Yokochi T, Honda D, Higashihara T, Nakahara T (1998) Optimization of docosahexaenoic acid production by Schizochytrium limacinum SR21. Appl Microbiol Biotechnol 49(1):72–76
Yu XJ, Sun J, Sun YQ, Zheng JY, Wang Z (2016a) Metabolomics analysis of phytohormone gibberellin improving lipid and DHA accumulation in Aurantiochytrium sp. Biochem Eng J 112:258–268
Yu XJ, Sun J, Zheng JY, Sun YQ, Wang Z (2016b) Metabolomics analysis reveals 6-benzylaminopurine as a stimulator for improving lipid and DHA accumulation of Aurantiochytrium sp. J Chem Technol Biotechnol 91:1199–1207
Yuanmei L, Ying L, Jie T, Jiong M, Jay JC, Maurycy D (2018) Transcriptomic profiling and gene disruption revealed that two genes related to PUFAs/DHA biosynthesis may be essential for cell growth of Aurantiochytrium sp. Mar Drugs 16:310
Zhang Y, Min Q, Xu J, Zhang K, Chen S, Wang H, Li D (2016) Effect of malate on docosahexaenoic acid production from Schizochytrium sp. B4D1. Electron J Biotechnol 19:56–60
Funding
This work was supported by the Key Research and Development Programs (Modern Agriculture) of Jiangsu Province (grant number BE2013405, BE2017322), the Six Talent Peaks Project of Jiangsu Province (grant number 2015-NY-018), the Key Research and Development Program (Modern Agriculture) of Zhenjiang City (grant number NY2017010), the 333 High-level Talent Training Project of Jiangsu Province (2018), and the Shen Lan Young scholars program of Jiangsu University of Science and Technology (2015).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Responsible editor: Ta Yeong Wu
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOC 196 kb)
Rights and permissions
About this article

Cite this article
Liu, ZX., You, S., Tang, BP. et al. Inositol as a new enhancer for improving lipid production and accumulation in Schizochytrium sp. SR21. Environ Sci Pollut Res 26, 35497–35508 (2019). https://doi.org/10.1007/s11356-019-06056-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-019-06056-3