
Abstract
Great efforts have been devoted to assessing the effects of straw managements on greenhouse gas (GHG) emissions, global warming potential (GWP), and net economic budget in rice monoculture (RM). However, few studies have evaluated the effects of straw managements on GHG emissions and net ecosystem economic budget (NEEB) in integrated rice-crayfish farming (RC). Here, a randomized block field experiment was performed to comprehensively evaluate the effects of aquatic breeding practices (feeding or no feeding of forage) and straw managements (rice straw returning or removal) on soil NH4+–N and NO−3–N contents, redox potential (Eh), CH4 and N2O emissions, GWP, and NEEB of fluvo-aquic paddy soil in a rice-crayfish co-culture system in Jianghan Plain of China. We also compared the differences in CH4 and N2O emissions, GWP, and NEEB between RM and RC. Straw returning significantly increased CH4 and N2O emissions by 34.9–46.1% and 6.2–23.1% respectively compared with straw removal. Feeding of forage decreased CH4 emissions by 13.9–18.7% but enhanced N2O emissions by 24.4–33.2% relative to no feeding. Compared with RM treatment, RC treatment decreased CH4 emissions by 18.1–19.6% but increased N2O emissions by 16.8–21.0%. Moreover, RC treatment decreased GWP by 16.8–22.0% while increased NEEB by 26.9–75.6% relative to RM treatment, suggesting that the RC model may be a promising option for mitigating GWP and increasing economic benefits of paddy fields. However, the RC model resulted in a lower grain yield compared with the RM model, indicating that more efforts are needed to simultaneously increase grain yield and NEEB and decrease GWP under RC model.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The global warming caused by greenhouse gas (GHG) emissions is a serious environmental problem in the world. Under current emission scenarios, the global surface temperature is expected to increase by an average of 1.0–3.7 °C by the year of 2100 (IPCC 2013). Agroecosystem is an important source of CH4 and N2O emissions in the atmosphere (IPCC 2007). Flooded rice field, as a primary CH4 source, can emit 33 to 40 Tg CH4 per year, accounting for approximately 15–20% of the total anthropogenic CH4 emissions (Sass and Fisher 1997; Yan et al. 2005, 2009). Rice field can provide a suitable environment for a wide range of aquatic animals, such as fish, freshwater prawns, shrimp, crabs, and turtles (Xie et al. 2011; Halwart 2006), and it has been reported that the conversion of rice paddies to aquaculture could significantly reduce CH4 and N2O emissions by 48% and 56%, respectively (Li et al., 2008; Liu et al. 2016). Thus, co-culture of rice and aquatic animals has been developed as a sustainable agricultural practice for mitigating the global warming potential (GWP) (Datta et al. 2009; Li et al. 2009; Sheng et al. 2018).
Integrated rice-crayfish co-culture, a symbiotic planting-breeding model, has been rapidly popularized in China due to its high economic benefits (Guo et al. 2017; Si et al. 2017). To date, the crayfish breeding area has surpassed 6 × 105 hm2 in China, and rice-crayfish model accounts for about 70% of the total (Guo et al. 2017). The RC model allows crawfish to live and prey in the rice fields, where the rice stubbles serve as the food basis for crawfish after the rice is harvested (Yuan et al. 2007). In addition, a peripheral trench is excavated and used as a refuge for the crayfish (Si et al. 2017). The model is characterized by the advantages of increasing resource utilization efficiency, improving soil carbon pool and microbial community structure, and contributing to high yield due to the activity of crayfish in the field (Si et al. 2017; Xu et al. 2017b). Many studies have been focused on the effects of rice-aquatic farming on CH4 and N2O emissions (Huang et al. 2005; Zhan et al. 2011; Bhattacharyya et al. 2013; Xu et al. 2017a, b). In general, most studies have reported on the mitigation of CH4 emissions and enhancement of N2O emissions in paddy field under rice-aquatic farming. However, few studies were focused on the effects of rice-crayfish co-culture on CH4 and N2O emissions (Cao et al. 2017; Xu et al. 2017b; Li et al. 2018).
Straw returning and feeding are the most common measures for boosting crayfish growth in the rice-crayfish model (Cao et al. 2017; Si et al. 2017). However, the effects of such agricultural practices on GHG emissions remain unclear. Therefore, this study aims to investigate the CH4 and N2O emissions from the rice-crayfish model through a two-year field experiment. We hypothesized that straw returning and feeding could significantly affect CH4 and N2O emissions from the rice-crayfish model. We also expected that the rice-crayfish co-culture could mitigate GWP compared with rice monoculture.
Materials and methods
Site description
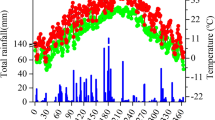
The experimental site is located in the Huazhong Agricultural University Research Farm in Houhu farm, Qianjiang City, Hubei Province, China (30°39′ N, 112°71′ E), where the soil was derived from the alluvial soil of the Yangtze River and classified as a fluvo-aquic paddy soil (PRC classification). This region has a winter static groundwater level of 40–55 cm and a northern, humid, subtropical monsoon climate. The mean annual precipitation is 1100 mm, and the mean annual temperature is 16.1 °C, with a frost-free period of 246 days. The monthly rainfall and air temperature during the experimental period from 2016 to 2017 are shown in Fig. 1. The soil properties of the experimental site (0–20 cm depth) are as follows: pH, 7.12; organic C, 26.43 g kg−1; total N, 2.41 g kg−1; total P, 0.44 g kg−1; and total K, 19.0 g kg−1.

Monthly precipitation and air temperatures during the 2016 and 2017 rice growing seasons. Tmin, monthly minimum air temperature; Tmean, monthly mean air temperature; Tmax, monthly maximum air temperature
Experimental design
The treatments were prepared with a random block design. Each treatment was of three replicates. The experiment included four treatments of rice-crayfish co-culture (RC) with an area of 1000 m2 and two treatments of rice monoculture (RM) with an area of 100 m2. The RC treatments included straw managements [rice straw returning (S) or removal (NS)] and feeding [feeding (F) or no feeding (NF) of forage], resulting in four combinations of treatments designated as RC + S + F, RC + NS + F, RC + S + NF, and RC + NS + NF; whereas the RM treatments included the RM with rice straw returning (RM-S) and removal (RM-NS). For all treatments, rice stubbles (5 cm above ground) were retained in the field after harvesting. For S treatment, rice straws were chopped to approximately 4–6 cm in length, which were subsequently incorporated into the soils, whereas the chopped straws were removal for NS treatment. The C/N ratio of rice straws was 45. For F treatment, supplementary forage (1500 kg ha−1) for the crayfish was supplied daily from April to June. The N, P2O5, and K2O contents of the crayfish forage were 46.6 g kg−1, 11.0 g kg−1, and 10.5 g kg−1, respectively.
The plots were plowed to 30 cm depth by a SNH554 tractor (Shanghai New Holland Agriculture Machinery Co., Ltd.) on June 12, 2016 and June 19, 2017. Taiyou390 (Oryza sativa L.) was mechanically transplanted by WP60D rice transplanter (Kubota China Holdings Co., Ltd.) at the density of 180,000 hills ha−1 on June 15, 2016 and June 24, 2017, and harvested on September 22, 2016 and October 23, 2017. Commercial compound fertilizer (26%–10%–15%, N–P2O5–K2O), urea (46% N), and potassium chloride (60% KCl) were used to provide 167 kg N ha−1, 37.5 kg P2O5 ha−1, and 123.7 kg K2O ha−1 during the rice growing seasons. N fertilizers were used at two doses, with 68% at the basal and 32% at the tillering stage. P fertilizers were only used as basal fertilizers, and K fertilizers were applied with 48% at the basal and 52% at the tillering stage.
For RC model, a peripheral trench (2.0 m wide and 1.2 m deep) was excavated as refuge for the crayfish (Procambarus clarkii) in August 2015. The excavated soil was used to construct a dyke (1.0 m high) surrounded by a nylon net (0.45 m high) to prevent the escaping of the crayfish. Crayfish larvae (5–6 g tail−1) were stocked at a density of 300 kg ha−1 in September 2015, and the crayfish was allowed to self-propagate inside the rice paddies. The immature crayfish would migrate to the peripheral trench during field puddling, seedling planting, drying, and re-watering, and would re-enter the field after these agricultural practices. Manure crayfish were harvested using crayfish cages (length = 5 m, diameter = 0.5 m) in early June and before the rice harvest in 2016 and 2017. For the RC model, the burrow density was observed to be 0.6 burrows·m−2 in the paddy field. For all treatments, the surface water in the paddy was kept at the level of 5–10 cm during rice growing seasons except for a drainage for 5–7 days in the vigorously tillering stage and 2 weeks before rice harvest.
Soil sampling and analytical methods
During rice growing seasons, soil samples at 0–20 cm depth were collected for the determination of NH4+–N and NO3−–N contents using a soil sampler (5 cm of inner diameter) in eight random positions about every 1 month. Soil NH4+–N and NO3−–N were extracted with 2 mol·L−1 KCl in a 1:3 ratio and determined by a FLAStar-5000 continuous flow analyzer (Foss, Sweden).
Soil Eh was determined five times (at about 3–4 week intervals) during rice growing seasons by FJA-4 portable oxidation-reduction potential analyzer (Nanjing Zhuan-Di Instrument and Equipment Company Limited, China).
CH4 and N2O emissions
Static chamber method was used to measure CH4 and N2O fluxes as described by Li et al. (2013). The gas samples were collected 15–16 times at about 7-day intervals during rice growing seasons. The CH4 and N2O concentrations were measured using a chromatograph meter (Shimadzu GC-14B), and the detailed measurement procedures were described by Li et al. (2013).
The seasonal total CH4 and N2O emissions were calculated by interpolating the gas emission during the sampling interval, assuming that the computation derived from measured fluxes represents the average daily flux (Li et al. 2013).
Grain yield measurement
Rice grains at three random positions in each plot were collected using a 2 m × 3 m frame. Rice within the frame was threshed, and the rice grains were adjusted to a moisture content of 14%.
GWP calculation
GWP was calculated by multiplying the seasonal total gas emissions by their respective radiative forcing potential (Bayer et al. 2014). On a 100-year time frame, the radiative forcing potentials of CH4 and N2O are 30 and 268, respectively (IPCC 2013). Therefore, the GWP was calculated according to the following equation:
NEEB calculation
NEEB can be used to evaluate the economic feasibility of eco-agriculture, and was calculated according to the following equation:
In the equation, yield gain was calculated from the current price of rice grains and crayfish (rice, 2620 CNY t−1; crayfish, 25,510 CNY t−1). Agricultural activity cost consisted of the costs of mechanical tillage and harvesting (1200 CNY ha−1), rice seeds (1000 CNY ha−1), fertilizers, pesticides and herbicides (5000 CNY ha−1), and forage (4500 CNY ha−1). The GWP cost was the product of carbon-trade price (103.7 CNY t−1 CO2-eq) and GWP (Li et al. 2015).
Data analysis
Data were statistically analyzed by SPSS 22.0. One-way NANOVA was used to assess the effects of straw management, feeding method, and the stocking of crayfish on NH4+–N and NO3−–N, Eh, GHG emissions, and NEEB. Duncan’s multiple range tests were performed to examine whether the differences in the mean values between treatments were statistically significant at the level of 0.05.
Results
Soil NH4 +–N and NO3 −–N
Soil NH4+–N contents ranged from 1.09 to 44.70 mg kg−1 under RC treatments (RC + S + F, RC + S + NF, RC + NS + F, and RC + NS + NF), and from 1.25 to 10.55 mg kg−1 under RM treatments (RM + S and RM + NS, Fig. 2a). The soil NO3−–N contents were lower than NH4+–N contents. The NO3−–N contents were 1.36–3.96 mg kg−1 under RC treatments, and 1.55–3.83 mg kg−1 under RM treatments (Fig. 2b).

Changes in soil NH4+–N (a) and NO3−–N (b) contents under different treatments in 2016 and 2017 rice seasons. RC, rice-crayfish co-culture; RM, rice monoculture; S, straw returning; NS, straw removal; F, feeding; NF, no feeding. Bars indicate standard error of the means
The stocking of crayfish significantly increased the content of NH4+–N in the soil, but did not affect that of NO3−–N (Fig. 2). RC treatments increased the mean NH4+–N content by 167.9–196.4% relative to RM treatments. Straw returning and feeding significantly affected soil NH4+–N content, but showed no influence on NO3−–N content. Straw returning increased the mean NH4+–N content by 6.0–19.4% compared with straw removal. Moreover, feeding of forage increased the mean NH4+–N content by 8.8–17.6% compared with no feeding.
Soil Eh
The introduction of crayfish into the field improved the soil oxidation potential (Fig. 3). Compared with RM treatments, RC treatments significantly increased the mean soil Eh by 10.8%. In addition, straw returning and feeding also had significant effects on soil Eh. Straw returning reduced the mean soil Eh by 8.5–15.8% relative to straw removal, and feeding led to 12.7–18.0% higher mean soil Eh than no feeding.

Changes in soil Eh under different treatments in 2017 rice seasons. RC, rice-crayfish co-culture; RM, rice monoculture; S, straw returning; NS, straw removal; F, feeding; NF, no feeding. Bars indicate standard error of the means
CH4 and N2O emissions
Seasonal changes in CH4 fluxes from paddy fields are shown in Fig. 4. Two peaks of CH4 fluxes were observed at the vigorously tillering and boosting stages in both 2016 and 2017 rice growing seasons. For RC treatments, the CH4 fluxes were 0.25–29.46 mg m−2 h−1 in 2016 and 0.65–30.60 mg m−2 h−1 in 2017; for RM treatments, the fluxes varied from 0.34 to 34.92 mg m−2 h−1 in 2016 and 1.23 to 33.91 mg m−2 h−1 in 2017.

Changes in CH4 fluxes under different treatments in 2016 and 2017 rice seasons. RC, rice-crayfish co-culture; RM, rice monoculture; S, straw returning; NS, straw removal; F, feeding; NF, no feeding. Bars indicate standard error of the means
Seasonal changes in N2O fluxes under different treatments are shown in Fig. 5. Peaks of N2O fluxes were found just immediately after each N fertilization and after drainage in 2016 and 2017 rice seasons. For RC treatments, the N2O fluxes ranged from 17.35 to 146.09 μg m−2 h−1 in 2016 and from 8.36 to 140.08 μg m−2 h−1 in 2017; for RM treatments, the fluxes were 14.75–63.39 μg m−2 h−1 in 2016 and 8.46–16.39 μg m−2 h−1 in 2017.

Changes in N2O fluxes under different treatments in 2016 and 2017 rice seasons. RC, rice-crayfish co-culture; RM, rice monoculture; S, straw returning; NS, straw removal; F, feeding; NF, no feeding. Bars indicate standard error of the means
The stocking of crayfish in the paddy fields significantly influenced cumulative CH4 and N2O emissions (Table 1). RC treatments decreased cumulative CH4 emissions by 18.1–19.6% compared with RM treatments, but increased cumulative N2O emissions by 16.8–21.0%. Moreover, straw returning and feeding showed significant impacts on cumulative CH4 and N2O emissions. Straw returning significantly enhanced cumulative CH4 and N2O emissions by 34.9–46.1% and 6.2–23.1% respectively relative to straw removal. Feeding decreased cumulative CH4 emissions by 13.9–18.7% but increased cumulative N2O emissions by 24.4–33.2% compared with no feeding.
Grain yield and GWP
The introduction of crayfish into the paddy fields also significantly affected rice grain yield and GWP (Table 2). RC treatments decreased the grain yield and GWP by 30.8–30.9% and 16.8–22.0% respectively relative to RM treatments. Straw returning and feeding did not affect grain yield, while had significant effects on GWP. Straw returning elevated GWP by 33.4–63.6% compared with straw removal, and feeding enhanced GWP by 14.5–29.4% compared with no feeding.
The raising of crayfish in the paddy fields significantly affected NEEB (Table 3). RC treatments increased NEEB by 26.9–75.6% relative to RM treatments. In addition, feeding significantly enhanced NEED by 45.1–123.0% compared with no feeding, while straw returning did not show any effect on NEEB.
Discussion
To date, few studies have described the effects of RC model on GHG emissions compared with RM model (Xu et al. 2017b). The aim of this study was to compare the differences in GHG emissions between RC and RM models, and also to investigate the effects of straw returning and feeding on GHG emissions under RC model. Our results indicate that straw returning and feeding of forage have significant effects on GHG emissions, and RC model could mitigate GWP and increase NEEB compared with RM model. Our results provide a scientific basis for assessing the ecological effects of RC model.
GHG emissions
Two peaks of CH4 fluxes were observed at the vigorously tillering and boosting stages during rice growing seasons in this study (Fig. 4). Similar results were reported by Li et al. (2013). The observed peaks may be ascribed to the increase in available organic carbon from the decomposition of fresh crop residues incorporated into the soil and the release of root exudates (Li et al. 2013) at the two stages. Moreover, high temperature (25–30 °C) at the two stages (Fig. 1) is suitable for the activities of methanogens (Kim et al. 2012; Sanchis et al. 2012), which leads to the rapid production of CH4. Peaks of N2O fluxes were found just immediately after each N fertilization (Fig. 5), possibly due to the increase in substrate availability from fertilizer N hydrolysis (Li et al. 2009; Ussiri et al. 2009). Other peaks were observed just after the fields were drained (Fig. 5), possibly because high soil O2 availability caused by drainage resulted in high nitrification in this stage (Sheng et al. 2018).
In the present study, straw returning led to higher CH4 emissions than straw removal (Table 2). Similar results have been widely reported (Zou et al. 2005; Bhattacharyya et al. 2013; Zhang et al. 2015). The returned straws could provide C substrates for methane-producing bacteria, thus promoting CH4 emissions. Moreover, the decomposition of rice straws can consume the O2 in the soils and thus increase soil anaerobic conditions (Fig. 3; Bayer et al. 2014), leading to the inhibition of CH4 oxidation. Straw returning can provide C and N substrates for denitrification or nitrification (Zou et al. 2005; Sanchis et al. 2012), as partly indicated by the higher NH4+–N content under straw returning, and thus higher N2O emissions under straw returning than under straw removal were observed in this study (Table 2).
Raising of crayfish led to the decrease in CH4 emissions and increase in N2O emissions under RC model in this study (Table 1). Mineralization of the forage residues in the field increased soil NH4+ contents (Fig. 2), which can facilitate CH4 oxidation (Bodelier et al. 2000, Bodelier 2011; Ellen and Frenzel 2001) and reduce CH4 emissions subsequently. The increased soil NH4+ contents by feeding (Fig. 2) can boost nitrification, thus increasing N2O emissions. Moreover, feeding can boost the foraging activity of crayfish, and thus accelerate the gas exchange among soil, water, and atmosphere, which would result in the increase in soil Eh (Fig. 3; Si et al. 2017; Xu et al. 2017b). The increase in soil Eh would decrease the activities of soil methanogens to reduce CH4 emissions. Higher Eh also enhances soil nitrification, and thus increases N2O emissions.
The stocking of crayfish into the fields decreased CH4 emissions (Table 1), which is similar to the result previously reported (Cao et al. 2017; Xu et al. 2017b). This result may be explained by the following reasons. Firstly, the crayfish can dig burrows for refuge, and disturb soil surface and plow the pan of paddy fields, thus improving soil permeability and Eh (Fig. 3; Si et al. 2017; Xu et al. 2017b). The elevated soil Eh will inhibit CH4 emissions. Secondly, the crayfish, an omnivore animal, can suppress weeds in the paddy fields, and thus reduce the consumption of floodwater dissolved O2 by weeds, resulting in an increase in soil Eh (Fig. 3) and CH4 oxidation subsequently. Thirdly, the foraging of crayfish in the fields increases the contact opportunity between soil and O2, which will improve soil Eh (Fig. 3) and thus lead to a decrease in CH4 emissions. Finally, the crayfish can eat rice stubbles retained in the fields, which accordingly reduces CH4 production from rice stubble decomposition under flooding conditions. In contrast, RC treatments increased N2O emissions relative to RM treatments (Table 1) possibly due to the increase in soil Eh (Fig. 3). The activities of digging burrows and foraging of crayfish improve the gas exchange among soil, water, and O2, as well as enhance soil Eh (Fig. 3), which thereby results in more N2O production from nitrification. Moreover, the excretions of crayfish can provide C and N substrates for nitrification and denitrification to promote N2O emissions (Si et al. 2017; Xu et al. 2017b).
Grain yield
In the present study, straw returning did not affect grain yield (Table 2), which is similar to the finding of Zhang et al. (2015). Although increased soil fertility due to straw returning has the potential to improve rice productivity (Pan et al. 2009), the negative effects of straw decomposition on soil ecological processes suggest that straw returning may inhibit rice root growth and development (Zavalloni et al. 2011), which might thereby offset the positive effects of increased soil fertility on rice yield. Moreover, feeding had no effect on grain yield (Table 2). The forage used in RC model consisted of fishbone dust, rapeseed cake, cotton kernel cake and corn meal, and so on. The application of forage may not be sufficient to increase rice yield due to the relatively low nutrient supply of the organic fertilizer (Bayu et al. 2006).
RC model resulted in a lower rice grain yield than RM model (Table 2), which is different from the result of Xu et al. (2017b) that similar rice grain yields were obtained between RC and RM models. This discrepancy may be attributed to the difference in the peripheral trench excavated as a refuge for crayfish. In this study, the area of peripheral trench (accounting for about 15–17% of the total area) was included in the calculation of grain yield in the RC model, which might decrease the grain yield of per unit area, while Xu et al. (2017b) did not consider the area of the peripheral trench when determining the grain yield.
GWP and NEEB
The GWP based on CH4 and N2O emissions under RC model was 5397–8590 kg CO2-eq. ha−1 in 2016 and 3847–7205 kg CO2-eq. ha−1 in 2017, which were comparable to the results of Xu et al. (2017b), who indicated 4960 kg CO2-eq. ha−1 of GWP under RC model in the same region of Jianghan Plain. In the present study, RC model decreased GWP by 16.8–22.0% relative to RM model (Table 2) owing to the effective inhibition of CH4 emissions. Similar result was reported by Xu et al. (2017b), who found 11.1–21.1% of reduction in GWP under RC model relative to RM model. Similar mitigating effects of integrated rice-breeding farming on GWP have been widely reported (Xu et al. 2017a). For example, Sheng et al. (2018) reported that integrated rice-duck farming could decrease the GWP by 28.0–28.1% compared with conventional rice monoculture in central China. Xu et al. (2017a) pointed out that rice-duck farming could lower the GPW by 6.7–14.6% relative to conventional rice monoculture in Nanjing city. Our results further indicate the mitigating effects of integrated rice-breeding farming on GWP from paddy fields.
NEEB is an indicator to integratedly assess agricultural economic and ecological benefits, and has far-reaching significance in guiding government decision-making and agricultural activities (Li et al. 2015; Zhang et al. 2015). In the present study, high economic benefits of crayfish resulted in high NEEB under RC model, especially under straw returning and feeding conditions (Table 3), suggesting that RC model may be an effective strategy to simultaneously increase economic benefits and mitigate GWP. However, RC model at the same time decreased rice grain yield obviously relative to RM model, suggesting that this model cannot increase rice grain yield to meet the food demand in the world. Therefore, more efforts should be made to maintain rice grain yield at the same time of mitigating GWP under RC mode (Cao et al. 2017).
Conclusions
This study shows that straw returning and feeding significantly affect CH4 and N2O emissions from paddy fields. RC model mitigates CH4 emissions, and thus reduces GWP and increases NEEB. Our results indicate that the RC model might be an effective practice to increase economic benefits and mitigate GWP at the same time. However, the RC model would result in a lower grain yield relative to RM model, suggesting that more efforts should be made to simultaneously maintain rice grain yield and mitigate GWP under RC model.
References
Bayer C, Costa FDS, Pedroso GM, Zschornack T, Camargo ES, Lima MAD, Frigheto RTS, Gomes J, Marcolin E, Macedo VRM (2014) Yield-scaled greenhouse gas emissions from flood irrigated rice under long-term conventional tillage and no-till systems in a humid subtropical climate. Field Crop Res 162:60–69
Bayu W, Rethman NFG, Hammes PS, Alemu G (2006) Effects of farmyard manure and inorganic fertilizers on sorghum growth, yield, and nitrogen use in a semi-arid area of Ethiopia. J Plant Nutr 29:391–407
Bhattacharyya P, Sinhababu DP, Roy KS, Roy KS, Dash PK, Sahu PK, Dandapat R, Neogi S, Mohanty S (2013) Effect of fish species on methane and nitrous oxide emission in relation to soil C, N pools and enzymatic activities in rainfed shallow lowland rice-fish farming system. Agric Ecosyst Environ 176:53–62
Bodelier PLE (2011) Interactions between nitrogenous fertilizers and methane cycling in wetland and upland soils. Curr Opin Environ Sustain 3:379–388
Bodelier PLE, Roslev P, Henckel T, Frenzel P (2000) Stimulation by ammonium-based fertilizers of methane oxidation in soil around rice. Nature 403:421–424
Cao CG, Jiang Y, Wang JP, Yuan PL, Chen SW (2017) “Dual character” of rice-crayfish culture and strategy for its sustainable development. Chin J Eco-Agric 25:1245–1253 (In Chinese)
Datta A, Nayak DR, Sinhababu DP, Adhya TK (2009) Methane and nitrous oxide emissions from an integrated rainfed rice–fish farming system of Eastern India. Agric Ecosyst Environ 129:228–237
Ellen G, Frenzel P (2001) Changes in activity and community structure of methane-oxidizing over the growth period of rice. Appl Environ Microbiol 67:2395–2403
Guo YF, Zhu ZW, Ma DW, Tang JQ (2017) The development report of China’s crayfish industry in 2017. China Fisheries 13:8–17 (In Chinese)
Halwart M (2006) Biodiversity and nutrition in rice-based aquatic ecosystems. J Food Compos Anal 19:747–751
Huang Y, Wang H, Huang H, Feng ZW, Yang ZH, Luo YC (2005) Characteristics of methane emission from wetland rice-duck complex ecosystem. Agric Ecosyst Environ 105:181–193
IPCC (2007) The physical science basis. Contributions of working group I to the fourth assessment report of the intergovernmental panel on climate change: the physical science basis. Cambridge University Press. Cambridge and New York
IPCC (2013) Climate change: the physical science basis: working group I contribution to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press. Cambridge and New York
Kim SY, Gutierrez J, Kim PJ (2012) Considering winter cover crop selection as green manure to control methane emission during rice cultivation in paddy soil. Agric Ecosyst Environ 161:130–136
Li B, Fan CH, Zhang H, Chen ZZ, Sun LY, Xiong ZQ (2015) Combined effects of nitrogen fertilization and biochar on the net global warming potential, greenhouse gas intensity and net ecosystem economic budget in intensive vegetable agriculture in southeastern China. Atmos Environ 100:10–19
Li CF, Cao CG, Wang JP, Zhang M, Yuan WL, Shahrear A (2009) Nitrous oxide emissions from wetland rice-duck cultivation systems in southern China. Arch Environ Contam Toxicol 56:21–29
Li C, Zhang Z, Guo L, Cao ML, Cao CC (2013) Emissions of CH4 and CO2 from double rice cropping systems under varying tillage and seeding methods. Atmos Environ 80:438–444
Li Q, Xu L, Xu L, Qian Y, Jiao Y, Bi Y, Zhang T, Zhang W, Liu Y (2018) Influence of consecutive integrated rice-crayfish culture on phosphorus fertility of paddy soils. Land Degrad Dev 29:3413–3422. https://doi.org/10.1002/ldr.3107
Liu S, Zou J, Wu S, Li S, Li Z, Zou J (2016) Methane and nitrous oxide emissions reduced following conversion of rice paddies to inland crab−fish aquaculture in Southeast China. Environ Sci Technol 50:633–642
Pan G, Smith P, Pan W (2009) The role of soil organic matter in maintaining the productivity and yield stability of cereals in China. Agric Ecosyst Environ 129:344–348
Sanchis E, Ferrer M, Torres AG, Cambra-López M, Calvet S (2012) Effect of water and straw management practices on methane emissions from rice fields: a review through a meta-analysis. Environ Eng Sci 29:1053–1062
Sass RL, Fisher FM (1997) Methane emissions from rice paddies: a process study summary. Nutr Cycl Agroecosyst 49:119–127
Sheng F, Cao CG, Li CF (2018) Integrated rice-duck farming decreases global warming potential and increases net ecosystem economic budget in central China. Environ Sci Pollut R 25:22744–22753
Si G, Peng C, Yuan J, Zhao S Xu D, Wu J (2017) Changes in soil microbial community composition and organic carbon fractions in an integrated rice–crayfish farming system in subtropical China. Sci Rep 7:2856. https://doi.org/10.1038/s41598-017-02984-7
Ussiri DAN, Lal R, Jarecki MK (2009) Nitrous oxide and methane emissions from long-term tillage under a continuous corn cropping system in Ohio. Soil Tillage Res 104:247–255
Xie J, Hu LL, Tang JJ, Wu X, Li NN, Yuan YG, Yang HS, Zhang JE, Luo SM, Chen X (2011) Ecological mechanisms underlying the sustainability of the agricultural heritage rice-fish coculture system. P Natl Acad Sci USA 108(50):1381–1387
Xu G, Liu X, Wang Q, Yu X, HangY (2017a) Integrated rice-duck farming mitigates the global warming potential in rice season. Sci Total Environ 575:58–66
Xu X, Zhang M, Peng C, Si G, Zhou J, Xie Y, Yuan J (2017b) Effect of rice-crayfish co-culture on greenhouse gases emission in straw-puddled paddy fields. Chin J Eco-Agric 25:1591–1603 (In Chinese)
Yan X, Akiyama H, Yagi K, Akimoto H (2009) Global estimations of the inventory and mitigation potential of methane emissions from rice cultivation conducted using the 2006 Intergovernmental Panel on Climate Change Guidelines. Global Biogeochem Cy. https://doi.org/10.1029/2008GB003299
Yan XY, Yagi K, Akiyama H, Akimoto H (2005) Statistical analysis of the major variables controlling methane emission from rice fields. Glob Chang Biol 11:1131–1141
Yuan Y, Bingner RL, Theurer FD, Kolian S (2007) Water quality simulation of rice/crawfish field ponds within annualized AGNPS. Appl Eng Agric 23:585–595
Zavalloni C, Alberti G, Biasiol S, Delle Vedove G, Fornasier F, Liu J, Peressotti A (2011) Microbial mineralization of bio-char and wheat straw mixture in soil: a short-term study. Appl Soil Ecol 50:45–51
Zhan M, Cao CG, Wang JP, Jiang Y, Cai ML, Yue LX, Shahrear A (2011) Dynamics of methane emission, active soil organic carbon and their relationships in wetland integrated rice-duck systems in Southern China. Nutr Cycl Agroecosyst 89:1–13
Zhang ZS, Guo LJ, Liu TQ, Li CF, Cao CC (2015) Effects of tillage practices and straw returning methods on greenhouse gas emissions and net ecosystem economic budget in rice–wheat cropping systems in central China. Atmos Environ 122:636–644
Zou J, Huang Y, Jiang J, Zheng X, Sass RL (2005) A 3-year field measurement of methane and nitrous oxide emissions from rice paddies in China: effects of water regime, crop residue, and fertilizer application. Global Biogeochem Cy 19. https://doi.org/10.1029/2004GB002401
Funding
This work is funded by the National Key Research and Development Project of China (2016YFD0300905), Hubei Agricultural Innovation Post Project of China, Hubei Modern Agricultural Industry Technology System of China, and Natural Science Foundation of Hubei Province (2016CFA017).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Philippe Garrigues
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Sun, Z., Guo, Y., Li, C. et al. Effects of straw returning and feeding on greenhouse gas emissions from integrated rice-crayfish farming in Jianghan Plain, China. Environ Sci Pollut Res 26, 11710–11718 (2019). https://doi.org/10.1007/s11356-019-04572-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-019-04572-w