
Abstract
To evaluate the possible effects of habitat type on crustacean plankton (hereafter zooplankton) biomass and body size, a 5-year study (2011 to 2015) was conducted during wet seasons in three habitats in Lake Poyang, China. The lacustrine habitat with the most stable hydrologic regime had the highest zooplankton biomass coinciding with the highest phytoplankton biomass. The riverine habitat with the lowest Secchi depth overall had the largest zooplankton body size, but high zooplankton biomass only in high water level years. The seasonally inundated floodplain habitats had the lowest zooplankton biomass and the smallest individual body size, coinciding with the lowest phytoplankton biomass and the highest predation pressure, the latter indicated by a low zooplankton: phytoplankton biomass ratio (ZB:PB). Multiple linear regression analyses indicated that pelagic zooplankton assemblages were primarily influenced by phytoplankton biomass in lacustrine habitat, by advection and turbidity in riverine regions, and by predation pressure in seasonally inundated floodplain region. We conclude that the importance of bottom-up and top-down effects on zooplankton biomass and body size varied with habitat type in Lake Poyang.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Numerous studies of the spatial distribution of crustacean plankton (hereafter zooplankton, including cladoceran and copepod plankton) have been conducted in lakes (Patalas and Salki 1984, 1992; Romare et al. 2003), rivers (Bass et al. 1997; Kim and Joo 2000; Thorp and Mantovani 2005; Wahl et al. 2008), and reservoirs (Bernot et al. 2004; Bini et al. 1997; Horn 1991; Masundire 1997; Naselli-Flores and Barone 1997) during the past decades. As zooplankton is food for fish and also grazers for phytoplankton in the trophic web, the effect of phytoplankton biomass (bottom-up) and predation (top-down) on the zooplankton assemblage structure have been studied extensively (Brooks and Dodson 1965; Havens et al. 2009; Jeppesen et al. 2000; Zaret and Kerfoot 1975). It is well known that phytoplankton and zooplankton biomass overall are positively related and that zooplankton community assemblages and size structure can be highly influenced by predation (Brooks and Dodson 1965; Dumont 1994; Jeppesen et al. 2000). The biomass ratio of zooplankton to phytoplankton (ZB:PB) has been widely used as an indicator of predation pressure in both temperate and subtropical lakes (Hart 2011; Havens and Beaver 2013; Havens et al. 2009; Jeppesen et al. 2003). Based on a study of 466 lakes, Jeppesen et al. (2003) argued that the ZB:PB (or predation risk for zooplankton) is unimodal related to total phosphorous in the north temperate zones, being low (high predation) in both the most nutrient-rich and the nutrient-poor lakes and high at intermediate nutrient levels. Compared with temperate lakes, however, the ZB:PB in subtropical lakes is relatively low and dominated by small size zooplankton taxa, inefficiently filtering large-sized phytoplankton, such as cyanobacteria (Havens et al. 2009).
Large lakes in subtropics and tropic subjected zones are often have high variable hydrological regimes spatially (e.g., flood pulse, current velocity, water retention time) (Patalas and Salki 1992), which affecting zooplankton community assemblages (Baranyi et al. 2002; G-Tóth et al. 2011; Thorp and Mantovani 2005). While the role of macrophytes for zooplankton distribution in lakes have been extensively studied (Meerhoff et al. 2012), much less is known about the effects of top-down and bottom-up forces on zooplankton assemblages in lakes that are highly influenced by variation in hydrology, both spatially within the lakes and among years (Otto et al. 2014).
We conducted a 5-year study of zooplankton biomass and body size in different habitats in the wet summer season 2011–2015 in China’s largest freshwater lake, Lake Poyang, which is a river-connected, integrated floodplain ecosystem. Our hypothesis was that zooplankton were influenced significantly by both food sources (phytoplankton) and predation in the most lentic part of the lake, while physical factors would be of higher importance in the riverine part. Finally, predation would be of key importance in seasonally inundated areas covered by macrophytes, as these plants expectedly would attract numerous small fish, as seen in other warm lakes (Meerhoff et al. 2012).
Description of Lake Poyang and the three habitats
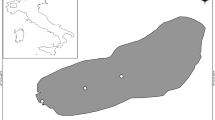
Lake Poyang is located in the Jiangxi province, China, at the south bank of Yangtze River (Fig. 1). It is the largest freshwater lake in China with a catchment area of 162,225 km2 and links to the Yangtze River (Ye et al. 2013). The bottom elevation of the lake ranges between 7 and 19 m (above sea level (a.s.l.)), with ~ 80% of the area being between 12 and 17 m a.s.l., while the low-elevation (7–12 m a.s.l.) regions only account for 12% (Feng et al. 2011; Li et al. 2014). The elevation of the lakebed generally decreases from south to north (Fig. 1(a)), with a difference of about 6.5 m (Li et al. 2014). The fluctuations in water level may be as high as 10 m within a year, and, consequently, the surface area may change from 3000 km2 in the wet seasons but shrinks from a lake-like system to a river-like one in the dry seasons (about 600 km2).

(a) Overview of the three habitat types during the low water level season in Lake Poyang, China. (b) The location of the 67 sampling sites in the high water level season. The photos are typical representations of four regions in summer and were taken in July 2014
During wet seasons, three typical types of habitats can be found in Lake Poyang: main channels (riverine), backwater lakes (lacustrine), and large areas of seasonally inundated alluvial lands (floodplain) (Fig. 1(a)). We, however, divided the lake into four zones, by subdividing the riverine part into two, the northern channel (NC) and the southern channel (SC), both subjected to high current velocities and busy traffic and being inundated for the whole year. The largest difference between NC and SC is their connection with lentic areas. SC divides the south of Lake Poyang into two well-connected parts (east region and west region). All inflow rivers run directly into SC, converging into the narrower NC connected to the Yangtze River. The west region (WR) consisting of seasonally inundated alluvial areas. The gross annual water discharges of the five inflow rivers account for 85% of the total water input to the lake (Gao et al. 2014). Ganjiang, Fuhe, Xinjiang, Raohe, and Xiushui contribute in average, respectively, with 55%, 12%, 14%, 9.3%, and 9.2% of the inflow sources (Zhu and Zhang 1997). Most of the water from SC run through WR, forming the seasonally inundate wetland. It is also recognized as a wetland of international importance for its remarkable biodiversity and habitat for various rare or endangered species, such as Yangtze finless porpoise, Siberian crane, and Oriental stork (Lai et al. 2014). Now, WR is the only region of the lake that has macrophytes in summer (Huang et al. 2002; Jian et al. 2015). Finally, the east region (ER) includes many backwater lakes that are connected to the main lake during high water level seasons and are relatively independent from the main channel during low water level seasons. ER once had dense macrophytes (Guan et al. 1987; Guan and Zhang 1987), but they disappeared several years ago.
Sampling was conducted over a 5-year period (2011–2015). As the water level and lake surface varied between years, the number of sampling sites (Fig. 1(b)) ranges from 34 to 67 (34, 56, 57, 66, and 67 from year 2011 to 2015). The NC was not sampled in 2011.
Sampling and analysis
Two sub-surface water samples at 0–1 m depth were obtained using a 5 liters (l) hydrophore (APHA 2005). This sampling procedure is not accounting for depth segregation (vertical migration). Our sampling procedure may potentially lead to incorrect biomass estimation, although we would not expect so as the water current is relatively high in the riverine lake and the water furthermore is turbid (less need for avoiding predation by staying deep in the daytime). The sampling strategy was selected to avoid anchoring-up at each site (strong current) when sampling. To assess the effect, we conducted vertical sampling at 11 sites (see appendices). We found no significant gradients in zooplankton biomass (Appendix 1), body length (Appendix 2), and community structure (Appendix 3). This may result from high speed of water velocity, which averaged > 0.2 m/s making passive movement more likely than active movement. The samples (10 l) were filtered through a 64-μm mesh net, and preserved in 50-ml polyethylene plastic bottles with 4% formaldehyde solution. Zooplankton with few numbers (< 200) were counted entirety under a dissecting microscope (at x 40-100 magnification), otherwise, sub-samples were used until 200 individuals were counted. Most Cladocera zooplankton were identified to species level following the keys (Jiang and Du1979). Species which could not be distinguished in routine counting (Appendix 4) were identified to genus level. The copepod zooplankton were separated into adult Calanoida, adult cyclops, copepodites, and nauplii. Zooplankton body lengths were measured for the first 20 individuals (if enough, otherwise measure all of them) of each species per sample using the “S-EYE” software. Size dry weight relations (Dumont et al. 1975) were used to calculate the dry biomass of zooplankton in 2012, 2014, and 2015. In 2011 and 2013, when zooplankton size was not measured, the biomasses were calculated from the mean value for each species in the other years (Appendix 4). The zooplankton biomass and body size in 2011 and 2013 may be estimated biasedly. While considering zooplankton mostly dominant by Bosmina and juvenile copepod, the system error may have a minor effect on horizontal distribution patterns.
We measured water temperature, pH, conductivity, water depth, Secchi depth, chlorophyll a (Chl a), total nitrogen (TN), total phosphorous (TP), and chemical oxygen demand (COD) according to standard methods as described by (Liu et al. 2015a, b; Wu et al. 2013). Water level at Hukou station was recorded from the website of Water Resources Department of Jiangxi Province (http://www.jxsl.gov.cn/slxxhw/jhsq/index.html) by Lake Poyang Laboratory for Wetland Ecosystem Research.
A two-way analysis of variance (ANOVA) with nonparametric tests was performed in order to compare the spatial and temporal differences of environmental parameters, zooplankton biomass, and body size. While only main effects of spatial differences were used for environmental parameters, of temporal differences were used for zooplankton biomass and body size in the result. Horizontal distribution patterns of zooplankton dry weight biomass and mean individual body weight were visualized by a geostatistical method of inverse distance weighted (IDW) interpolation. Data was first explored using linear regression with scatter plot. In addition, we ran multiple linear regression analysis with forward selection to identify the most important environmental variables influencing zooplankton biomass and mean body size. Before and during the process of multiple linear regression analysis, collinearity diagnostics was checked by Spearman correlations (ρ < 0.60) and variance inflation factor (VIF < 5) respectively. If necessary, loga (bx + 1) transformation was undertaken to obtain normal distribution. ArcGIS 10.3.1 (USA), Statistical Program for Social Sciences (SPSS-IBM, New York, NY) 20.0 and OriginLab9.0 (USA) software packages were used in the data analyses.
Results
Limnetic and hydrological conditions in wet seasons
NC and SC had the highest water depth and WR the lowest (Fig. 2). The water level was by far lowest in the dry year 2011, being about 4 m below the 10-year average (2004–2014), while it was higher than this average in 2012, 2014, and 2015.

a Variation in mean water level in the sampling period at the Hukou station based on weekly measurements, normalized to the average water level of July from the 10-year period 2004–2014. b Variation of mean water depth in the five study years and four regions (NC, northern channel; SC, southern channel; ER, eastern region; WR, western region) of Lake Poyang in summer
Significant differences were found in Chl a, Secchi depth, COD concentration, and pH between the different regions (Fig. 3), but not in water temperature, conductivity, and nutrient concentrations. ER had the highest Chl a concentration, COD concentration, and pH, while WR had the highest Secchi depth. NC and SC had the lowest Secchi depths and Chl a concentration. The lowest Secchi depth were caused by turbulent in NC and SC. No differences between the two channel regions, NC and SC, were found for any of the chemical variables analyzed.

Mean and standard deviations of chlorophyll a (Chl a), water temperature, Secchi depth, chemical oxygen demand (COD), pH, conductivity, total phosphorus (TP), and total nitrogen (TN) in the four study regions (NC, northern channel; SC, southern channel; ER, eastern region; WR, western region). Results of an ANOVA (df = 3) with Bonferroni correction and Student-Newman-Keuls (SNK) test are shown. Different letters indicate significant differences (P < 0.05), “*log” indicates the test was performed on log-transformed data
Horizontal distribution in zooplankton biomass and body size
The zooplankton biomass varied between the five study years and regions (Fig. 4). Zooplankton biomass was significantly (ANOVA, F = 10.9, df = 4, P = 0.001) higher in 2013 than in the other years. Horizontally, the highest biomass was reached in ER (134 μg/L), followed by NC (85 μg/L) and SC (46 μg/L), and the lowest in WR (30 μg/L). The biomass was overall higher in ER than in WR. The lowest biomass was always found in WR, whereas the highest value occurred in ER in 2011 and 2013, and in NC and ER in 2012, 2014, and 2015. NC had relatively high biomasses only when the water level was higher than average in normal years (Fig. 2a). The copepod and cladoceran biomass distributions were rather similar to each other (Fig. 4).

Inverse distance weighting interpolation of a cladoceran and b copepod biomass in the high water level season of Lake Poyang from 2011 to 2015. The northern channel was not sampled in 2011
The zooplankton body size was overall larger in low water level years such as 2011 and 2013, and there was a decreasing trend in body size of copepods from 2011 to 2015 (Fig. 5). Horizontally, WR overall had the smallest body size and NC the largest. The low water level year, 2011, had the largest copepod body size and smallest cladoceran body size.

Inverse distance weighting interpolation of a cladoceran and b copepod body size (mean individual body weight) in the high water level season of Lake Poyang from 2011 to 2015. The northern channel was not sampled in 2011
Regression analysis
Water depth only had significantly positive relationship with zooplankton biomass in NC and WR (Fig. 6); in a multiple regression, however, the water depth relationship with cladoceran biomass became significant also for ER (Table 1). The slopes of the relationship between cladoceran and copepods versus water depth were significantly higher in NC than in the other regions. Cladoceran biomass exhibited a significantly negative relationship with Secchi depth in ER and SC, but for copepod biomass only in SC (Fig. 6); in a multiple regression, the relationship in NC became significant too, but positively (Table 1). On the contrary, cladoceran biomass exhibited a significant positive relationship with phytoplankton biomass in ER and SC, while for copepod biomass only in ER (Fig. 6); in a multiple regression, the relationship becomes significantly negative related in NC.

Scatterplots and linear regressions of biomass of cladocerans and copepods with water depth, Secchi depth, and phytoplankton biomass
Body size of copepod in SC, ER, and WR had significantly negative relationship with water depth (Fig. 7); in a multiple regression, the relationship became significant negative for copepods in SC and ER, while it was positive for cladocerans in SC (Table 2). Body size of cladocerans had significantly negative relationship with Secchi depth in WR, of copepods in NC and ER (Fig. 7); in a multiple regression, the relationship for cladocerans was not significant, however (Table 2). Body size of copepods exhibited a significant positive relationship with phytoplankton biomass in ER and WR, but negative in NC (Fig. 7), while in a multiple regression analysis, only significant for ER (Table 2).

Scatterplots and linear regressions of body size (individual body weight) of cladocerans and copepods with water depth, Secchi depth, and phytoplankton biomass
ZB:PB was significantly negative related with phytoplankton biomass except for ER, but had significantly positive relationship with cladoceran and copepod biomass in all regions (Fig. 8). The ZB:PB ratio was better explained by the zooplankton biomass (R2 = 0.69~0.80) than by the phytoplankton biomass (R2 = 0~0.46).

Scatterplots and linear regressions of zooplankton to phytoplankton biomass ratio (ZB:PB) versus cladocerans, copepod biomass and body size (individual body weight), and phytoplankton biomass
Discussion
Our results showed dramatically horizontal differences of zooplankton biomass and body size within one very large and habitat diverse lake. The influences of bottom-up, top-down, and hydrological process on zooplankton biomass and body size patterns in this habitat multiple lake, however, had deviant results with individual habitat simplex lakes. Specifically, our observation will be helpful in creating a new understanding of zooplankton distribution in habitat diverse lakes.
We found the largest mean zooplankton body size (though still relatively small-sized) in the lotic riverine habitats (SC and NC) with lowest Secchi depths perhaps because the fish foraging efficiency is lower for many fish species in turbid water (Estlander et al. 2009; Zaret and Kerfoot 1975). This observation is consistent with most reports (Brooks and Dodson 1965; Zhou et al. 2018) but inconsistent with some field reports and experiments (Chen and Chen 2017; Schulze 2011). High abundances of small-sized zooplankton have earlier been found in turbulent and high turbidity environmental habitats (Bernot et al. 2004; Vijverberg and Boersma 1997) like floodplains (Baranyi et al. 2002; Bernot et al. 2004; Masundire 1997; Patalas and Salki 1984) and rivers (Davidson Jr et al. 1998; Massicotte et al. 2013; Thorp and Mantovani 2005; Wahl et al. 2008) in different climate zones as well as in experiments (Francesc and Marrasé 2000; G-Tóth et al. 2011). By contrast, Baranyi et al. (Baranyi et al. 2002) found the larger species, like Daphnia cucullata and Ceriodaphnia pulchella, to be more abundant in lentic than in lotic habitats in the Danube River floodplain system. The differences probably due to habitat diversity, because fish can move among habitats, for example, compare with turbid riverine habitat, fish prefer to forage in floodplain. In our lake, the fish community was dominated by potential planktivorous fish. Carassius auratus, Cyprinus carpio, Silurus asotus, and Pelteobagrus fulvidraco are most dominant species in floodplain areas, while Coilia auratus, Hemiculter bleekeri, and Pseudobrama simoni are most dominant species in riverine areas (Yang et al. 2015).
Accordingly, floodplain WR had the lowest zooplankton biomass, the smallest zooplankton body size, and the lowest ZB:PB ratio during all years, all indicating that this region faced the highest predation pressure (Havens and Beaver 2013; Havens et al. 2009; Jeppesen et al. 2007). However, large-bodied cladocerans such as Daphnia rarely occurred in any of the regions indicating an overall high predation pressure on zooplankton (Brooks and Dodson 1965; David and Joseph 1990) in summer in this lake, but then particularly high in WR. In addition, Yang et al. (2015) found the WR area had the highest fish density, biomass, and species richness, while ER the lowest, which was also confirmed by Fang et al. (2016). Evidence that predation was responsible for the lack of large-bodied zooplankton in the lake can also be derived from fish-manipulated Lake Junshan (separated from Lake Poyang via a dam built for the purpose of aquaculture). Here, relatively large-bodied Daphnia cucullata was found to dominate the zooplankton community after lake restoration by fish removal (Liu et al. 2015c).
Interestingly, we found ER to have the highest zooplankton biomass during all years and it was the only region where this biomass was positively related with the phytoplankton biomass. Moreover, ER was the only region where the ZB:PB ratio was not negatively related with phytoplankton biomass, indicating altogether that bottom-up control was comparatively most important in ER (Brett et al. 2009; Jeppesen et al. 1999). This result is well documented in studies of lentic lakes (Sommer et al. 2012; Straile 2015; Yoshida et al. 2003), especially for studies emphasizing eutrophication (Gagneten and Paggi 2009; Li et al. 2016; Shi et al. 2016). We found a significantly negative relationship between ZB:PB ratio and phytoplankton biomass in the riverine SC and NC, and the slopes of ZB:PB versus phytoplankton biomass were, accordingly, significantly larger (absolute value) than in the other habitats. Turbulence, high turbidity, and high water velocity may reduce zooplankton feeding, growth, and reproduction (Wahl et al. 2008) and thus weaken bottom-up effect in lotic habitats (Havens et al. 2009; Sterner et al. 1996).
Horizontally, distribution of zooplankton biomass and body size is also controlled by inter-year variation of water level, which was also observed in particulate organic matter (Liang et al. 2016) and phytoplankton (Liu et al. 2015a, b). Relatively high zooplankton biomass occurred in outlet NC only in high water level season is consistent with observation of Bini et al. (1997) and Marzolf (1990). Besides, we found water depth to have a significant negative relationship with zooplankton body size but positive relationship with zooplankton biomass in NC and WR, which may due to inter-year variation of flow distribution and lentic habitat’s size (Burdis and Hirsch 2017; G-Tóth et al. 2011; Ning et al. 2010). An expanding lake littoral area with increasing water level may support larger zooplankton production. An increasing water level may, however, also offer better sites for fish spawning with a shoreline covered by macrophytes (Naselli-Flores and Barone 1997; Wang et al. 2011). Correspondingly, in accordance with the latter, reports of “People’s Daily” (http://cppcc.people.com.cn/GB/34958/15349412.html) and statements by local fisherman, fish yield of Lake Poyang in the very dry year 2011 drops to less than half of the yield in normal years.
In conclusion, our study suggests that habitat heterogeneity and hydrology play important roles in modulating the response of zooplankton communities to top-down and bottom-up control factors: Bottom-up effect turned out to be strongest in lacustrine habitat and weakest in lotic riverine habitat; top-down effects appeared to be highest in seasonally inundated floodplain lands and low in yearly inundated riverine channels. We also found water level variation leads to major inter-year zooplankton body size and biomass of zooplankton.
References
APHA (2005) Standard methods for the examination of water and wastewater. American Public Health Association (APHA), Washington, DC
Baranyi C, Hein T, Holarek C, Keckeis S, Schiemer F (2002) Zooplankton biomass and community structure in a Danube River floodplain system: effects of hydrology. Freshwat Biol 47:473–482
Bass JAB, Pinder LCV, Leach DV (1997) Temporal and spatial variation in zooplankton populations in the River Great Ouse: an ephemeral food resource for larval and juvenile fish. Regul Rivers Res Manag 13:245–258
Bernot RJ, Dodds WK, Quist MC, Guy CS (2004) Spatial and temporal variability of zooplankton in a great plains reservoir. Hydrobiologia 525:101–112
Bini LM, Tundisi JG, Matsumura-Tundisi T, Matheus CE (1997) Spatial variation of zooplankton groups in a tropical reservoir (Broa Reservoir, São Paulo State-Brazil). Hydrobiologia 357:89–98
Brett MT, Kainz MJ, Taipale SJ, Seshan H (2009) Phytoplankton, not allochthonous carbon, sustains herbivorous zooplankton production. Proc Natl Acad Sci 106:21197–21201
Brooks JL, Dodson SI (1965) Predation, body size, and composition of plankton. Science 150:28–35
Burdis RM, Hirsch JK (2017) Crustacean zooplankton dynamics in a natural riverine lake, Upper Mississippi River. J Freshw Ecol 32:240–258
Chen M, Chen F (2017) Effect of suspended solids on interaction between filter-feeding fish Aristichthys nobilis and zooplankton in a shallow lake using a mesocosm experiment. J Freshw Ecol 32:214–222
David W, Joseph S (1990) Refuge availability: a key to understanding the summer disappearance of Daphnia. Freshw Biol 24:43–62
Davidson NL Jr, Kelso WE, Rutherford DA (1998) Relationships between environmental variables and the abundance of cladocerans and copepods in the Atchafalaya River Basin. Hydrobiologia 379:175–181
Dumont HJ (1994) On the diversity of the Cladocera in the tropics. Hydrobiologia 272:27–38
Dumont HJ, Van de Velde I, Dumont S (1975) The dry weight estimate of biomass in a selection of Cladocera, Copepoda and Rotifera from the plankton, periphyton and benthos of continental waters. Oecologia 19:75–97
Estlander S, Nurminen L, Olin M, Vinni M, Horppila J (2009) Seasonal fluctuations in macrophyte cover and water transparency of four brown-water lakes: implications for crustacean zooplankton in littoral and pelagic habitats. Hydrobiologia 620:109–120
Fang CL, Chen WJ, Zhou HM, Zhang YP, Fu PF, He G, Wu B, Wang S (2016) Suggestions on utilization of fishery resources in Lake Poyang. JAAS 44:233–243
Feng L, Hu CM, Chen XL, Li RF, Tian LQ, Murch B (2011) MODIS observations of the bottom topography and its inter-annual variability of Poyang Lake. Remote Sens Environ 115:2729–2741
Francesc P, Marrasé C (2000) Effects of turbulence on plankton: an overview of experimental evidence and some theoretical considerations. Mar Ecol Prog Ser 205:291–306
Gagneten AM, Paggi JC (2009) Effects of heavy metal contamination (Cr, Cu, Pb, Cd) and eutrophication on zooplankton in the lower basin of the Salado River (Argentina). Water Air Soil Pollut 198:317–334
Gao JH, Jia JJ, Kettner AJ, Xing F, Wang YP, Xu XN, Yang Y, Zou XQ, Gao S, Qi SH, Liao FQ (2014) Changes in water and sediment exchange between the Changjiang River and Poyang Lake under natural and anthropogenic conditions, China. Sci Total Environ 481:542–553
G-Tóth L, Parpala L, Balogh C, Tàtrai I, Baranyai E (2011) Zooplankton community response to enhanced turbulence generated by water-level decrease in Lake Balaton, the largest shallow lake in Central Europe. Limnol Oceanogr 56:2211–2222
Guan SF, Zhang B (1987) Biomass of macrophytes of the Poyang Lake with suggestions of its rational exploitation. Acta Hydrobiologica Sinica 3:219–227
Guan SF, Lang Q, Zhang B (1987) Aquatic vegetation of Poyang Lake. Acta Hydrobiologica Sinica 1:9–21
Hart R (2011) Zooplankton biomass to chlorophyll ratios in relation to trophic status within and between ten South African reservoirs: causal inferences, and implications for biomanipulation. Water SA 37:513–522
Havens KE, Beaver JR (2013) Zooplankton to phytoplankton biomass ratios in shallow Florida lakes: an evaluation of seasonality and hypotheses about factors controlling variability. Hydrobiologia 703:177–187
Havens KE, Elia AC, Taticchi MI, Fulton RS (2009) Zooplankton–phytoplankton relationships in shallow subtropical versus temperate lakes Apopka (Florida, USA) and Trasimeno (Umbria, Italy). Hydrobiologia 628:165–175
Horn W (1991) The influence of biomass and structure of the crustacean plankton on the water transparency in the saidenbach storage reservoir. Hydrobiologia 225:115–120
Huang Y, Pu Y, Li W, Wang C (2002) A study on communities with Potamogeton malaianus MIQ. in Poyang Lake Nature Reserve of People’s Republic of China. Thaiszia - J. Bot. 12:51–60
Jeppesen E, Jensen JP, Søndergaard M, Lauridsen T (1999) Trophic dynamics in turbid and clearwater lakes with special emphasis on the role of zooplankton for water clarity, Shallow lakes’ 98. Springer, Berlin, pp 217–231
Jeppesen E, Peder Jensen J, SØndergaard M, Lauridsen T, Landkildehus F (2000) Trophic structure, species richness and biodiversity in Danish lakes: changes along a phosphorus gradient. Freshw Biol 45:201–218
Jeppesen E, Jensen JP, Jensen C, Faafeng B, Hessen DO, Søndergaard M, Lauridsen T, Brettum P, Christoffersen K (2003) The impact of nutrient state and lake depth on top-down control in the pelagic zone of lakes: a study of 466 lakes from the temperate zone to the arctic. Ecosystems 6:313–325
Jeppesen E, Meerhoff M, Jacobsen B, Hansen R, Søndergaard M, Jensen J, Lauridsen T, Mazzeo N, Branco C (2007) Restoration of shallow lakes by nutrient control and biomanipulation—the successful strategy varies with lake size and climate. Hydrobiologia 581:269–285
Jian MF, Wang SC, Yu HP, Li LY, Jian MF, Yu GJ (2015) Fluorescence properties of submerged macrophytes in Nanjishan wetland, southern Poyang Lake. J Resour Ecol 6:52–59
Jiang XZ, Du NS (1979) Fauna sinica Crustacea Freshwater Cladocera. Science Press, Academia Sinica, Beijing
Kim HW, Joo GJ (2000) The longitudinal distribution and community dynamics of zooplankton in a regulated large river: a case study of the Nakdong River (Korea). Hydrobiologia 438:171–184
Lai X, Huang Q, Zhang Y, Jiang J (2014) Impact of lake inflow and the Yangtze River flow alterations on water levels in Poyang Lake, China. Lake Reservoir Manage 30:321–330
Li YL, Zhang Q, Yao J, Werner AD, Li XH (2014) Hydrodynamic and hydrological modeling of the Poyang Lake catchment system in China. J Hydrol Eng 19:607–616
Li Y, Xie P, Zhao DD, Zhu TS, Guo LG, Zhang J (2016) Eutrophication strengthens the response of zooplankton to temperature changes in a high-altitude lake. Ecol Evol 6:6690–6701
Liang Y, Liu XZ, Xiao HY, Gao XF, Li WH, Xiong J (2016) Impact of high water level fluctuations on stable isotopic signature of POM and source identification in a floodplain lake-Bang Lake (Poyang Lake). Environ Earth Sci 75:12
Liu X, Li YL, Liu BG, Qian KM, Chen YW, Gao JF (2015a): Cyanobacteria in the complex riverconnected Poyang Lake: horizontal distribution and transport. Hydrobiologia 768:95–110
Liu X, Qian K, Chen Y (2015b) Effects of water level fluctuations on phytoplankton in a Changjiang River floodplain lake (Poyang Lake): implications for dam operations. J Great Lakes Res 41:770–779
Liu BG, Tan GL, Xing JS, Li M, Chen YW (2015c) Effect of pen culture on community structure of planktonic crustaceans in Lake Junshan. Journal of Ecology and Rural Environment 1:82–87
Marzolf G (1990): Reservoirs as environments for zooplankton. IN: Reservoir limnology: ecological perspectives. John Wiley & Sons, New York. 1990. p 195–208, 1 fig, 43 ref.
Massicotte P, Frenette JJ, Proulx R, Pinel-Alloul B, Bertolo A (2013) Riverscape heterogeneity explains spatial variation in zooplankton functional evenness and biomass in a large river ecosystem. Landsc Ecol 29:67–79
Masundire HM (1997) Spatial and temporal variations in the composition and density of crustacean plankton in the five basins of Lake Kariba, Zambia-Zimbabwe. J Plankton Res 19:43–62
Meerhoff M, Teixeira-de Mello F, Kruk C, Alonso C, Gonzalez-Bergonzoni I, Pacheco JP, Lacerot G, Arim M, Beklioğlu M, Brucet S (2012) Environmental warming in shallow lakes: a review of potential changes in community structure as evidenced from space-for-time substitution approaches. Adv Ecol Res 46:259–349
Naselli-Flores L, Barone R (1997) Importance of water-level fluctuation on population dynamics of cladocerans in a hypertrophic reservoir (Lake Arancio, south-west Sicily, Italy). Hydrobiologia 360:223–232
Ning NSP, Nielsen DL, Hillman TJ, Suter PJ (2010) The influence of planktivorous fish on zooplankton communities in riverine slackwaters. Freshw Biol 55:360–374
Otto SA, Diekmann R, Flinkman J, Kornilovs G, Mollmann C (2014) Habitat heterogeneity determines climate impact on zooplankton community structure and dynamics. PLoS One 9:1–11
Patalas K, Salki A (1984) Effects of impoundment and diversion on the crustacean plankton of Southern Indan Lake. Can J Fish Aquat Sci 41:613–637
Patalas K, Salki A (1992) Crustacean plankton in Lake Winnipeg-variation in space and time as a function of lake morphology, geology, and climate. Can J Fish Aquat Sci 49:1035–1059
Romare P, Berg S, Lauridsen T, Jeppesen E (2003) Spatial and temporal distribution of fish and zooplankton in a shallow lake. Freshw Biol 48:1353–1362
Schulze PC (2011) Evidence that fish structure the zooplankton communities of turbid lakes and reservoirs. Freshw Biol 56:352–365
Shi H, Chen G, Lu H, Wang J, Huang L, Wang L, Zhao S, Liu X (2016) Regional pattern of Bosmina responses to fish introduction and eutrophication in four large lakes from Southwest China. J Plankton Res 38:443–455
Sommer U, Adrian R, De Senerpont Domis L, Elser JJ, Gaedke U, Ibelings B, Jeppesen E, Lürling M, Molinero JC, Mooij WM (2012) Beyond the Plankton Ecology Group (PEG) model: mechanisms driving plankton succession. Annu Rev Ecol Evol Syst 43:429–448
Sterner R, Kilham S, Johnson F, Winner R, Keeling T, Yeager R, Farrell M (1996) Factors regulating phytoplankton and zooplankton biomass in temperate rivers. Limnol Oceanogr 41:1572–1577
Straile D (2015) Zooplankton biomass dynamics in oligotrophic versus eutrophic conditions: a test of the PEG model. Freshw Biol 60:174–183
Thorp JH, Mantovani S (2005) Zooplankton of turbid and hydrologically dynamic prairie rivers. Freshw Biol 50:1474–1491
Vijverberg J, Boersma M (1997) Long-term dynamics of small-bodied and large-bodied cladocerans during the eutrophication of a shallow reservoir, with special attention for Chydorus sphaericus. Hydrobiologia 360:233–242
Wahl DH, Goodrich J, Nannini MA, Dettmers JM, Soluk DA (2008) Exploring riverine zooplankton in three habitats of the Illinois River ecosystem: where do they come from? Limnol Oceanogr 53:2583–2593
Wang Y, Yu X, Li W, Xu J, Chen Y, Fan N (2011) Potential influence of water level changes on energy flows in a lake food web. Chin Sci Bull 56:2794–2802
Wu Z, Cai Y, Liu X, Xu CP, Chen Y, Zhang L (2013) Temporal and spatial variability of phytoplankton in Lake Poyang: the largest freshwater lake in China. J Great Lakes Res 39:476–483
Yang SR, Li MZ, Zhu QG, Wang MR, Liu HZ (2015) Spatial and temporal variations of fish assemblages in Poyang Lake. Resources and Environment in The Yangtze Basin 24:54–64
Ye XC, Zhang Q, Liu J, Li XH, Xu CY (2013) Distinguishing the relative impacts of climate change and human activities on variation of streamflow in the Poyang Lake catchment, China. J Hydrol 494:83–95
Yoshida T, Urabe J, Elser JJ (2003) Assessment of 'top-down' and 'bottom-up' forces as determinants of rotifer distribution among lakes in Ontario, Canada. Ecol Res 18:639–650
Zaret TM, Kerfoot WC (1975) Fish predation on Bosmina longirostris: body-size selection versus visibility selection. Ecology 56:232–237
Zhou J, Qin B, Han X (2018) The synergetic effects of turbulence and turbidity on the zooplankton community structure in large, shallow Lake Taihu. Environ Sci Pollut Res 25:1168–1175
Zhu H, Zhang B (1997) Poyang Lake. Press of University of Science and Technology of China, Hefei
Acknowledgements
We would like to express our deep thanks to Anne Mette Poulsen from Aarhus University for editorial assistance.
Funding
This study was financially supported by the National Natural Science Foundation of China (Grant 41671096) and the MARS project (Managing Aquatic ecosystems and water Resources under multiple Stress) funded under the 7th EU Framework Programme, Theme 6 (Environment including Climate Change), Contract No. 603378 (http://www.mars-project.eu).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Thomas Hein
Electronic supplementary material
ESM 1
(DOCX 57 kb)
Rights and permissions
About this article

Cite this article
Liu, B., Liu, J., Jeppesen, E. et al. Horizontal distribution of pelagic crustacean zooplankton biomass and body size in contrasting habitat types in Lake Poyang, China. Environ Sci Pollut Res 26, 2270–2280 (2019). https://doi.org/10.1007/s11356-018-3658-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-018-3658-7