
Abstract
The Huayanghe Lakes play an important role in the Yangtze floodplain in China and had extremely high water levels during the summer of 2016. Monitoring data was collected in an effort to understand the impact of this change on the crustacean zooplankton composition and abundance and the biomass variation in the Huayanghe Lakes between a regular hydrological cycle (RHC) and an extreme hydrological cycle (EHC). The crustacean zooplankton community composition, abundance, and biomass in the floodplain lakes were markedly affected by the water-level disturbance. The number of species was lower in the RHC, but the mean density and biomass decreased from 93.84 ± 13.29 ind./L and 6.11 ± 0.89 mg/L, respectively, in the RHC to 66.62 ± 10.88 ind./L and 1.22 ± 0.26 mg/L, respectively, in the EHC. Pearson correlations and redundancy analyses revealed the environmental factors with the most significant impact on the crustacean zooplankton community differed between the RHC and EHC cycles. Little previous information exists on the zooplankton in these lakes, and the present study provides data on the zooplankton composition, abundance, and biomass, both at baseline and in response to hydrological changes.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The responses of aquatic organisms to altered environmental conditions should be determined to effectively manage water resources. The dynamic ecosystems of natural lakes are ultimately influenced by hydrological fluctuations. Focusing on the mechanisms of a succession of biological communities is an important facet of ecological research (Mittelbach and Schemske 2015). Hydrological variation greatly influences the composition, abundance, and dynamics of the zooplankton community (Wetzel 2001; Van den Brink et al. 1994; Masundaire 1994). Furthermore, zooplankton can also exhibit a wide tolerance for various drivers in lakes (Elliott and Quintino 2007).
There is a broad range of limnological literature regarding the zooplankton of floodplain lakes; these organisms are highly responsive to changes in environmental factors, including flood pulses and connections to other waterways (Goździejewska et al. 2016), water levels (Schriver et al. 1995; Havens et al. 2007; Deboer et al. 2015), and oxygen levels (Aka et al. 2000; Jones et al. 2016). Zooplankton species composition, abundance, and biomass of zooplankton in floodplain lakes vary significantly during different water-level phases in response to environmental changes. Flooding or drought causes the limnological parameters to change (Thomaz et al. 2007), substantially affecting the interactions between plankton in the lakes (Aoyagui and Bonecker 2004). Such knowledge is critical to understanding and predicting the changes in floodplain lake communities resulting from water-level fluctuations. Information on the role of hydrology on organisms in floodplain lakes connected to the Yangtze River has focused mainly on phytoplankton (Wang et al. 2015; Qian et al. 2016). Surprisingly, with little attention paid to zooplankton communities (Wang et al. 2010), the Huayanghe Lakes play an important ecological function in the Yangtze River basin, which also has the characteristics of nature, typicality, scarcity, and diversity. The area experiences rainy season from mid-June to mid-July, and in the summer of 2016, it experienced the highest rainfall recorded in the previous three decades. The Huayanghe Lakes exceeded their guaranteed level of 0.27 m, reaching a new high of 17.07 m (Fig. 1, data from www.ncc-cma.net).

Annual rainfall in the Huayanghe Lakes basin. The solid and dashed lines denote the daily average rainfall in 2015 and 2016, respectively
To study the impact of extremely high flooding on the zooplankton community in the Huayanghe Lakes, multivariate analyses were performed to evaluate the zooplankton composition, abundance, and biomass in three sub-lakes (Lake Bohu, Lake Huangda, and Lake Longgan) in relation to environmental factors. We hypothesized zooplankton abundance, and biomass decreased in the extreme hydrological cycle. Furthermore, we hypothesize that the zooplankton community structure differed between the RHC and EHC; the given results will be compared to the results of zooplankton in regular hydrological cycle.
Material and methods
Study area
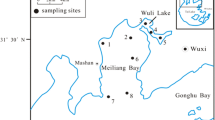
Huayanghe Lakes (29°52′48.8″~30°08′07.6″N; 116°04′29.2″~116°33′19.8″E), involving Lake Bohu, Lake Haungda, and Lake Longgan, is located at the connection of the middle and lower reaches of the Yangtze River (Fig. 2). The lakes have a monsoon-influenced humid subtropical climate with four distinct seasons and abundant rainfall in the spring and summer of each year. The lakes provide a unique habitat because of its long shoreline. During the winter, the lakes become home to a large number of migratory birds. Also, in recent years, due to the rapid development of a fishery (Zhang et al. 2008), the lakes have gradually seen an influx of a type of algae. The ground of the lakes is a local fault depression which formed and separated shallow lakes by silting from the Yangtze River during the long-term flood. When the water level reaches 17 m, an occupied area is nearly 800 km2 (Du et al. 2015). The water flows to the Yangtze River mainly through the Huayang and Yangwan rivers.

Map of Huayanghe Lakes group and distribution of sampling stations
We sampled 26 sites in three sub-lakes according to the type of lake (seven sites in Lake Bohu, nine sites in Lake Huangda, and ten sites in Lake Longgan) during the period of April 2015 to January 2017. The sampling sites are shown in Fig. 2.
Sampling and analyses
At each sampling site, several water column parameters were measured in the field: Secchi depth (SD), turbidity (NTU, Hach, America); water temperature (WT) (°C); pH (EL2, Mettler Toledo, Switerland); and dissolved oxygen (mg/L, Hach, America). The samples were analyzed in the laboratory and involved the following: chlorophyll a (Chl a), using the 90% acetone extraction method according to the Surface Water Environment Quality Standard (GB3838-2002 2002), total nitrogen (TN), and total phosphorus (TP) were analyzed according to standard methods (SEPB 2002).
Crustacean zooplankton samples were taken with a 5-L modified Patalas’ bottle sampler. Each sample was a mixture of several sub-samples collected from the surface of the water to the bottom at 0.5 or 1 m intervals. Cladocerans and copepods were collected from the remaining 10 L of mixed lake water with 64-μm mesh plankton net. Crustacean zooplankton organisms were fixed in 4% formaldehyde solution prior to identification. The crustacean identifications were conducted at the species level when possible according to the methods of Jiang and Du (1979) for Cladocerans and Sheng (1979) for copepods under a light microscope (BX53, Olympus, Tokyo, Japan) at × 4 magnification. The mean biomass (net weight) of the first 30 of each crustacean species was estimated according to a length-weight relationship (Zhang and Huang 1991; Bolduc et al. 2016).
Data analyses
In this study, we defined a regular hydrological cycle (RHC) as a period from April 2015 to January 2016 and further defined the period of May 2016 to January 2017 as an extreme hydrological cycle (EHC). The dominant species of crustacean zooplankton, determined based on the dominance value of each species, was computed as
where ni is the abundance of individual species i, N is the total number of all species, and fi is the occurrence frequency of species i. Detrended correspondence analysis (DCA) was used to determine if the length of crustacean zooplankton dominant gradient was low (< 3). The redundancy analysis (RDA) was completed using CANOCO 4.5, and the data was transformed into log10 (x + 1) expected pH before running the program. Pearson correlation analysis was performed by the SPSS package (Version 20.0, IBM http://www.ibm.com) to evaluate the influence of the limnological variables on crustacean zooplankton densities. Plotting of the diagrams was done using GraphPad Prism 5. ANOVA was also conducted to test the significant difference between the RHC and the EHC by the soft. The interpolation map was constructed by ArcGIS software (Version 10.0, ESRI http://www.esri.com) using the inverse distance weighting method (Wang et al. 2015; Jiang et al. 2014).
Results
Water quality
The physicochemical parameters of the Huayanghe Lakes were examined (Figs. 3 and 4). The samples were taken from 26 sampling sites from April 2015 to January 2017. The water depth of the lakes increased beginning in March 2016 and rose more than 4 m; flooding then began. There was unexpected extreme flooding (mean water depth, 5.85 m) in the summer of 2016, with a water depth of 1.70 m higher than that of 2015 (ANOVN, F1, 40 = 4.281, p < 0.001). Subsequently, the depth gradually decreased. WT and transparency, as well as WD, had an increasing and then decreasing trend. But there were no significant differences between the RHC and EHC (F1, 170 = 1.362, p = 0.155, F1, 170 = 1.378, p = 0.141, respectively). The pH value was generally > 7 (except in January 2017), indicating alkaline conditions. DO and turbidity in the summer reached lower values than those in the other seasons between two hydrological cycles. Chl a, TP, and TN concentrations changed significantly in the two periods (p < 0.05).

Water depth of the Huayanghe Lakes during each sampling period

Physicochemical parameters measured in the Huayanghe Lakes during each sampling period
Crustacean zooplankton composition, abundance, and biomass
A total of 22 genera and 25 species of crustacean zooplankton were observed in the present study. Among them, there were 14 species of crustacean zooplankton in the RHC, and 23 species were found in the EHC. The dominant species included Bosmina longirostris, Diaphanosoma brachyurum, Moina micrura, Sinocalanus dorrii, Limnoithona sinensis, and copepod nauplii in the RHC sampling season and D. brachyurum, M. micrura, B. longirostris, Bosmina coregoni, S. dorrii, Pseudodiaptomus forbesi, Mesocyclops leuckarti, Limnoithona sinensis, and copepod nauplii. The number of dominant species decreased in the EHC compared to the RHC. Figure 5 shows the density and biomass of the mixed crustacean zooplankton assemblage at different sampling sites. The density and biomass of crustacean zooplankton during the RHC were 93.84 ± 13.29 ind./L and 6.11 ± 0.89 mg/L, respectively (mean ± standard error), and 66.62 ± 10.88 ind./L and 1.22 ± 0.26 mg/L, respectively, during the EHC. The density of crustacean zooplankton was significantly different (F1, 170 = 1.584, p = 0.040) between the sampling periods, as was the biomass (F1, 170 = 12.41, p < 0.001).

The density and biomass of crustacean zooplankton collected from Lake Bohu, Lake Huangda, and Lake Longgan. The lines in the box indicate the mean level
Relationship between crustacean zooplankton and environmental factors
Pearson correlation analysis showed that WT and WD were both significantly positively correlated with the total density of crustacean zooplankton (p < 0.05) in both of the periods investigated (Table 1). There was a negative significant relationship between the pH (r = − 0.229, p = 0.032) and DO level(r = − 0.443, p < 0.01) and the total density of crustacean zooplankton sampling during RHC; however, pH had the reverse effect (r = 0.300, p = 0.007) on the total density during the EHC. Furthermore, Chl a level had a positive influence on the total density (r = 0.297, p = 0.005) in the RHC, and TN and TP had a significant effects on the total density (p < 0.05) during the EHC sampling period. The total biomass showed a significant positive correlation with the WT and the Chl a level (p < 0.01), but a negative relationship with the DO level (r = − 0.308, p = 0.004) during the RHC. However, the EHC total biomass had a positive correlation with the WT and pH (p < 0.05), and a negative correlation with TP (r = − 0.304, p = 0.006). The results indicated that the environmental factors most affecting crustacean zooplankton differed in the two hydrological periods.
We performed a redundancy analysis (RDA) to summarize the relationship between the crustacean zooplankton species and environmental variables (Fig. 6). The Huayanghe Lakes RDA ordination triplot included different dominant species and nine environmental variables. The eigenvalues for RDA axis 1 (0.341) and axis 2 (0.116) explained 79.8% of the variance in the crustacean zooplankton during the RHC. The species-environment correlation of axis 1 was 0.790, and axis 2 was 0.827—indicating a significant relationship between the nine limnological variables and the dominant crustacean zooplankton taxa. However, the eigenvalues for the first RDA axis (0.248) and the second axis (0.110) explained only 60.6% of the variance in the species during the EHC. The species-environment correlation of the first axis was 0.741, and the second axis was 0.678. We found that the WT, SD, WD, and DO levels were the main factors that affected the zooplankton community structure in the RHC, but changed differently in the EHC.

Redundancy analysis of a major zooplankton species (original point) in relation with environmental variables (arrows). Abbreviations used in the figure: Bl Bosmina longirostris, Bf Bosmina fatalis, Db Diaphanosoma brachyurum, Mm Moina micrura, Sd Sinocalanus dorrii, Ml Mesocyclops leuckarti, Cn copepod nauplii, Ls Limnoithona sinensis, Sf Schmackeria forbesi. To allow a better graphical representation, only the most abundant species are shown
Discussion
Extreme flooding as a driving force for the changes in the Huayanghe Lakes variables
Biotic conditions in aquatic systems are impacted by water-level fluctuations, which usually play a synergetic role in maintaining the prevailing community structure in floodplain lakes. The limnology parameters are markedly affected, and therefore, excessive water levels in floodplain lakes have significant implications (Okogwu 2010; Simões et al. 2012). The extreme rain experienced in the Huayanghe Lakes area in June of 2016 submerged a large number of farmlands, creating a surface hydrological connection between the river channel and different areas, and lasted until the beginning of August. Consequently, there were changes in the environmental factors of the lakes, especially since the mean WD was 1.5 times higher than usual (Fig. 3).
Extreme flooding in the floodplain systems is regarded as a vital element for the nutrient status of lakes. Although the nutrients showed similar trends between the RHC and EHC, there were significantly higher TN and TP concentrations (Fig. 6) in the EHC than in the RHC. This may have been due to increased accumulation of organic matter from terrestrial plants and other natural inputs. The Chl a content represents the phytoplankton biomas, and higher Chl a levels were observed in the EHC, especially after the flood. This was because the nutrients concentration was higher than during in the same sampling seasons of the RHC. Thus, the phytoplankton abundance increased with the nutrients concentration.
The response of crustacean zooplankton to different environment disturbances
Crustacean zooplankton assemblage composition is affected by lateral connectivity, which is easily disturbed by water-level fluctuation. Okogwu (2010) observed that the highest numbers of Cladocera species during in a relatively low water-level period. However, we observed the opposite result, with increased species richness in the EHC than in the RHC. These species may have increased due to the increased water volume during the flood period. In addition, Zu et al. (2012) also reported that the continuous increasing of water level increased the richness of crustacean zooplankton Lake Bohu (one of Huaynghe lakes).
It is a monsoon-influenced humid subtropical climate with four distinct seasons in Huayanghe Lake. WT had an increasing and then decreasing trend during the two periods, but there were no significant differences between the RHC and EHC. In the present study, WT was the main factor that affected the zooplankton community structure in the RHC and EHC. This indicates that there are seasonal changes in the community structure of zooplankton. This is in agreement with the different water levels of Nafi’u and Ibrahim (2017) in Thomas Dam.
Zooplankton abundance and biomass were both influenced by some environmental factors and were dramatically affected by the extreme water-level changes. The physical and chemical variables that affected crustacean zooplankton abundance and biomass were varied during the different periods. In the RHC, the DO and Chl a levels were the key factors affecting abundance and biomass. However, pH and TP were important factors affecting the density and biomass of zooplankton in the EHC. In addition, TN and SD also effected on the zooplankton community in the EHC.
The peak of mean density was observed during the RHC season, inconsistent with reports from Ologe Lagoon Floodplain Lake (Okogwu and Ugwumba 2006). Extreme flooding of lakes may have affected the crustacean zooplankton population. Bozelli et al. (2015) postulated that crustacean zooplankton density might decrease during the flooding phase of the Trombetas River floodplain due to dilution effects. Consequently, individual organisms may be lost from the lakes into the rivers. In addition, the increased lake volume could also expand the habitat availability of crustacean zooplankton and enhanced the swimming ability of certain (Medley and Havel 2007), and flooding could also introduce new species.
Hydrological alterations can result in the loss of aquatic plants (Van Geest et al. 2005). Eight different species of aquatic plants covered more than 35% of the Huyanghe Lakes group in the summer of 2015 (Table 2); however, very few aquatic plants when the water level was extremely high during the summer of 2016. In the present study, the decrease in the density of zooplankton may be due to the change of refuge habitat during the EHC compared to the density of the RHC. It also suggests a reduction in the refuge effect for crustacean zooplankton against predation by fish, which have a well-known affection for the crustacean zooplankton community structure (Carvalho 1984; Havens et al. 2007). Although the Chl a concentration was the highest during the EHC phase, significantly lower density in the crustacean zooplankton occurred during this period. The changes in crustacean zooplankton densities could be caused by differences in feeding habits among different crustacean zooplankton. Okogwu (2010) also has suggested that differences in predation pressure on crustacean zooplankton during the different phases might contribute to changes in the floodplain lakes.
References
Aka M, Pagano M, Saint-Jean L, Bouvy M, Cecchi P, Corbin D, Thomas S (2000) Zooplankton variability in 49 shallow tropical reservoirs of Ivory Coast (West Africa). Internat Rev Hydrobiol 85:439–504
Aoyagui ASM, Bonecker CC (2004) Rotifers in different environments of the upper Paraná River floodplain (Brazil): richness, abundance and the relationship with connectivity. Hydrobiologia 522(1):281–290
Bolduc P, Bertolo A, Pinel-Alloul B (2016) Does submerged aquatic vegetation shape zooplankton community structure and functional diversity? A test with a shallow fluvial lake system. Hydrobiologia 778:151–165
Bozelli RL, Thomaz SM, Padial AA, Lopes PM (2015) Floods decrease zooplankton beta diversity and environmental heterogeneity in an Amazonian floodplain system. Hydrobiologia 753:233–241
Carvalho ML (1984) Influence of predation by fish and water turbidity on a Daphnia gessneri, population in an Amazonian floodplain lake, Brazil. Hydrobiologia 113(1):243–247
Deboer JA, Webber CM, Dixon TA, Pope KL (2015) The influence of a severe reservoir drawdown on springtime zooplankton and larval fish assemblages in Red Willow Reservoir, Nebraska. J Freshw Ecol 31(1):1–16
Du HW, Zhang HJ, Fan ZY, Zhi JY, Xiao JE (2015) Study on characteristics of sediments in Huayang River-Lake group. Environ Sci Technol 38(1):128–132 (in Chinese)
Elliott M, Quintino V (2007) The estuarine quality paradox, environmental homeostasis and the difficulty of detecting anthropogenic stress in naturally stressed areas. Mar Pollut Bull 54(54):640–645
GB3838-2002 (2002) Environmental quality standard for surface water. State Environmental Protection Administration of China (in Chinese)
Goździejewska A, Glińska-Lewczuk K, Obolewski K, Grzybowski M, Kujawa R, Lew S, Grabowska M (2016) Effects of lateral connectivity on zooplankton community structure in floodplain lakes. Hydrobiologia 774(1):7–21
Havens KE, East TL, Beaver JR (2007) Zooplankton response to extreme drought in a large subtropical lake. Hydrobiologia 589(1):187–198
Jiang XZ, Du NS (1979) Fauna sinica: crustacean, freshwater cladocera. Science Press, Beijing (in Chinese)
Jiang YJ, He W, Liu WX, Qin N, Ouyang HL, Wang QM, Kong XZ, He QS, Yang C, Yang B, Xu FL (2014) The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu). Ecol Indic 40:58–67
Jones S, Perissinotto R, Carrasco NK, Vosloo A (2016) Impact of a flood event on the fooplankton of an estuarine lake. Mar Biol Res 12(2):1–10
Liu JJ (2016) Study on vascular plants in Susong Huayang Lake nature reserves and Shengjin Lake nature reserves, Anhui Province. University of Anhui, Dissertation (in Chinese)
Masundaire HM (1994) Seasonal trends in zooplankton densities in Sanyati basin, Lake Kariba: multivariate analysis. Hydrobiologia 272:211–230
Medley KA, Havel JE (2007) Hydrology and local environmental factor influencing zooplankton communities in floodplain ponds. Wetlands 27:864–872
Mittelbach GG, Schemske DW (2015) Ecological and evolutionary perspectives on community assembly. Trends Ecol Evol 30(5):241–247
Nafi’u SA, Ibrahim S (2017) Seasonal dynamics of zooplankton composition and abundance in Tomas Dam Dambatta, Kano, Nigeria. Bayero J Pure Appl Sci 10(1):268–276
Okogwu OI, Ugwumba OA (2006) The zooplankton and environmental characteristics of Ologe Lagoon, Southwest. Nigeria Zool 3:86–92
Okogwu OI (2010) Seasonal variations of species composition and abundance of zooplankton in Ehoma Lake, a floodplain lake in Nigeria. Rev BiolTrop 58(1):171–182
Qian KM, Liu X, Chen YW (2016) Effects of water level fluctuation on phytoplankton succession in Poyang Lake, China––a five year study. Ecohydrology Hydrobiol 16(3):175–184
Schriver P, Bøgestrand J, Jeppesen E, Søndergaard M (1995) Impact of submerged macrophytes on fish and zooplankton-phytoplankton interactions: large-scale enclosure experiments in a shallow eutrophic lake. Freshw Biol 33:255–270
Sheng JR (1979) Fauna sinica: crustacean, freshwater copepoda. Science Press, Beijing (in Chinese)
Simões NR, Lansac-Tôha FA, Velho LF, Bonecker CC (2012) Intra and inter-annual structure of zooplankton communities in floodplain lakes: a long-term ecological research study. Rev Biol Trop 60(4):1819–1836
State Environmental Protection Bureau (SEPB) (2002) Methods of monitoring and analysis for water and wastewater, 4th edn. China Environmental Science Press, Beijing (in Chinese)
Thomaz SM, Bini LM, Bozelli RL (2007) Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 579(1):1–13
Van den Brink FHB, Van Katwijk MM, Van der Velde G (1994) Impact of hydrology on phyto- and zooplankton community composition in floodplain lakes along the Lower Rhineand Meuse. J. Plankton Res 16:351–373
Van Geest GJ, Wolters H, Roozen FCJM, Coops H, Roijackers RMM, Buijse AD, Scheffer M (2005) Water-level fluctuations effect macrophyte richness in floodplain lakes. Hydrobiologia 539:239–248
Wang L, Wang C, Deng DG, Zhao XX, Zhou ZZ (2015) Temporal and spatial variations in phytoplankton: correlations with environmental factors in Shengjin Lake China. Environ Sci Poll R 22(18):14144–14156
Wang SB, Xie P, Geng H (2010) The relative importance of physicochemical factors and crustacean zooplankton as determinants of rotifer density and species distribution in lakes adjacent to the Yangtze River, China. Limnologica 40(1):1–7
Wetzel RG (2001) Limnology––lake and river ecosystems. Academic Press, New York
Zhang TL, Li ZJ, Guo QS (2008) Investigations on fishes and fishery of four lakes along the middle and lower basins of the Changjiang River. Acta Hydrobiologia Sin 32(2):167–177 (in Chineses)
Zhang ZS, Huang XF (1991) Study methods of freshwater plankton. Science Press, Beijing (in Chinese)
Zu GZ, Bao CH, Hu JH, Rong CZ, Liu X, Duan RH, Wang XX, Wang L, Liu QM, Cheng BS, Liu TW (2012) Investigation on hydrobios resource evolution in Bohu lake ecosystem restoration area. J Anh Agr Univ 39(3):327–335 (in Chinese)
Acknowledgements
This work was supported by the Anhui Provincial Education Department’s Natural Science Research Project of China (Y06061777). We thank Jingjing Liu and Jianxun Wu for their participation in the field work.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Philippe Garrigues
Rights and permissions
About this article

Cite this article
Zhang, K., Xu, M., Wu, Q. et al. The response of zooplankton communities to the 2016 extreme hydrological cycle in floodplain lakes connected to the Yangtze River in China. Environ Sci Pollut Res 25, 23286–23293 (2018). https://doi.org/10.1007/s11356-018-2144-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-018-2144-6