
Abstract
Carbon (C), nitrogen (N) and phosphorus (P) are the three most important essential elements limiting growth of primary producers. Submerged macrophytes generally absorb nutrients from sediments by root uptake. However, the C:N:P stoichiometric signatures of plant tissue are affected by many additional factors such as taxonomy, nutrient availability, and light availability. We first revealed the relative importance of taxonomy, sediment, and water column on plant C:N:P stoichiometry using variance partitioning based on partial redundancy analyses. Results showed that taxonomy was the most important factor in determining C:N:P stoichiometry, then the water column and finally the sediment. In this study, a significant positive relationship was found between community C concentration and macrophyte community biomass, indicating that the local low C availability in macrophytes probably was the main reason why submerged macrophytes declined in Yangtze floodplain shallow lakes. Based on our study, it is suggested that submerged macrophytes in Yangtze floodplain shallow lakes are primarily limited by low light levels rather than nutrient availability.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Carbon (C):nitrogen (N):phosphorous (P) stoichiometry provides a very powerful way to enhance our understanding of primary production, nutrient cycling, and population dynamics in freshwater and terrestrial ecosystems (Andersen et al. 2004; Elser and Urabe 1999; Evans-White and Lamberti 2006; Sterner and Hessen 2003). C, N and P are three essential elements of organisms and have strong interactions in biochemical functioning (Agren 2008). Many ecological processes such as photosynthesis (Elser et al. 2000; Reich et al. 1997), litter decomposition (Güsewell and Gessner 2009), predator–prey relationships (Ngai and Jefferies 2004; Tibbets and Molles 2005), community composition, and species diversity (Bedford et al. 2008; Gusewell et al. 2005) are related to C, N and P contents or C:N:P mass ratios. In freshwater ecosystems, for instance, N:P ratio can influence the productivity and species composition of plant communities (Hall et al. 2005; Harpole et al. 2011; Willby et al. 2001).
C:N:P stoichiometry of submerged vegetation could be influenced by sediment and water column nutrient availability because submerged macrophytes rely on the surrounding sediment and water to satisfy their N and P requirements (Cao et al. 2011; Madsen and Cedergreen 2002). Previous studies have addressed the relative importance of roots and leaves in nutrient uptake of rooted submerged macrophytes (Carignan and Kalff 1980; Rattray et al. 1991; Robach et al. 1995). Given that the concentrations of nutrients in sediments are usually higher than that of in water column, it is generally accepted that sediments are the major source of N and P for submerged macrophytes (Barko and Smart 1981; Best and Mantai 1978; Bristow and Whitcombe 1971; Carignan and Kalff 1980; Smith and Adams 1986).
The C:N:P stoichiometric signatures in plant tissue, however, depend not only on nutrient supply but also on the light availability in water column. Light availability in water column can affect physiology, morphology, biomass allocation, community structure, or distribution of submersed macrophytes, causing great variation in the C, N, and P concentrations and C:N:P stoichiometry in plant (Cao et al. 2011; Chambers and Kalff 1987; Cronin and Lodge 2003; Xing et al. 2013). For instance, to alleviate low light availability in deep waters, Potamogeton maackianus and Potamogeton malaianus tend to enhance light harvesting by allocating more biomass to the stem, increasing shoot height and specific leaf area, whereas Vallisneria natans tend to allocate more biomass to the leaf than to the rosette (Fu et al. 2012). Furthermore, the C:N:P stoichiometry in plant tissue is not always directly correlated with sediment or water column N and P availability (Güsewell and Koerselman 2002). Indeed, plants will accumulate nutrients in excess of their cellular requirements when their growth is not limited by N and P availability (Cao et al. 2011; Demars and Edwards 2007).
In recent years, eutrophication induced by human activities has changed water and sediment physicochemical conditions extensively. Consequently, these changes have altered nutrient and light availability, leading to variations of nutrient uptake by plants. Thus, current anthropogenic changes in N:P stoichiometry may have important implications for the ecological functioning of lake ecosystems. Additionally, since submerged macrophytes are different in taxonomy, growth forms, and C/N/P metabolic strategies, their stoichiometric signatures are species-specific (Yuan et al. 2016). Thus, which one of the three factors (taxonomy, sediment, and water column) is having the greatest influence on C:N:P stoichiometry of macrophytes is still unclear. In this paper, we aim to (i) explore how sediment and water column affect C, N, and P stoichiometry of submerged macrophytes and (ii) reveal the relative importance of taxonomy, sediment, and water column on the C:N:P stoichiometry of submerged macrophytes.
Materials and methods
Study sites and field sampling
Fourteen lakes along the mid-lower Yangtze River were investigated from June to August 2014 (Fig. 1). The longitudes of the studied lakes range from 112.2° E to 120.4° E and latitudes range from 29.7° N to 33.8° N. All the studied lakes are large shallow floodplain lakes (water depth ranged from 1.1 to 4.1 m) with different nutrient levels (Table 1).

Location of sampled lakes along the middle and lower Yangtze River, China
The species we focused on are six common submerged macrophytes which are widely distributed in the Yangtze floodplain lakes: Potamogeton maackianus, Potamogeton malaianus, Ceratophyllum demersum, Hydrilla verticillata, Myriophyllum spicatum, and V. natans. Distribution of the collected submerged macrophytes at the 14 lakes was shown in Table 2. To reduce environmental stress, these macrophyte species take different growth forms and have various adaptive strategies, including morphological feature and metabolism (Ni 2001; Su et al. 2004; Titus and Adams 1979).
The abundance of submerged macrophytes was measured by random sampling (three replicates) at each site with a long-handled scythe-type sampler (0.2 m2 in sampling area). All macrophytes were cleaned, sorted by species, and then weighed for wet biomass, respectively. Aboveground parts of the six plant species were collected and put into ziplock bags with waterproof labels. Corresponding water was taken from 0.5 m below the water surface. Surface sediments were collected from the top 0–10-cm layers of undisturbed sediments with a columnar sampling instrument (acrylic glass tube, 5 cm inner diameter). All the samples were put into icebox and taken back to laboratory for further analysis. Water depth (WD), transparency (SD), and light attenuation coefficient (K) were also measured at the same locations. WD was determined by a sounding lead. SD was measured by a Secchi disk. Photosynthetically active radiation (PAR) was measured by a LI-COR sensor (at water depth of 0, 0.5, 1.0, and 1.5 m, respectively) coupled with a data logger (Li-1400; LI-COR Company, Lincoln, NE, USA). K in the water column was calculated based on the equation: K = (1 / Z)ln(I 0 / I Z), where I Z is PAR at water depth z and I 0 is PAR at the water surface (Krause-Jensen and Sand-Jensen 1998).
Laboratory analysis
The collected submerged macrophytes were washed carefully to remove epiphytic algae, sediment, and invertebrates. The samples of macrophytes and sediments were oven-dried at 80 °C for 48 h to constant weights and then ground into fine powder using a planetary ball mill (Fritsch, Planetary Micro Mill PULVERISETTE 7 premium line, Germany) before the elemental analyses. The C and N concentrations of plant tissues and sediments were determined by an elemental analyzer (Flash EA 1112 series, CE Instruments, Italy). The total P was measured using a sulfuric acid/hydrogen peroxide digest and the ammonium molybdate ascorbic acid methods (Sparks et al. 1996). Sediment water content was determined gravimetrically by oven-drying at 105 °C to achieve constant weights. Total nitrogen (TN), total phosphorus (TP), and chlorophyll a (Chl a) of water samples were measured according to Chinese standard methods (Huang et al. 1999).
Data analysis
The tissue concentrations and the C:N, C:P, and N:P mass ratios at species level were compared with one-way ANOVA. Turkey’s HSD was used to test the significance between means. Pearson’s correlation analyses were performed using PASW (Predictive Analytics Software) Statistics 18 (SPSS, Inc., Chicago, IL, USA) between tissue elements and corresponding water and sediment parameters, respectively. Redundancy analyses (RDA) were performed with CANOCO for windows (version 5) to elucidate the relationship between plant C:N:P signatures and the environmental parameters. Variance partitioning based on partial redundancy analyses (pRDA) by vegan package in R (R Development Core Team 2014) was used to interpret the relative importance of taxonomy, sediment, and water column on the C:N:P stoichiometry of plant tissues. Monte Carlo permutation tests (Manly 2006) were used to test the significance of each environmental factor and all the constrained factors, respectively.
Results
Environmental conditions in sediment and water column
The average concentrations of sediment C, N, and P were 20.3 ± 14.78, 2.03 ± 1.06, and 0.56 ± 0.16 mg g−1, respectively (Table 3). The average sediment C:N:P ratio was 38:4:1 by mass. Sediment water content ranged from 24.7 to 78.1 % and was significantly correlated with sediment C, N, and P concentrations, respectively (sediment C, r = 0.71, p < 0.001; sediment N, r = 0.79, p < 0.001; sediment P, r = 0.35, p < 0.001).
The mean concentrations of TN, TP, and Chl a in water column were 0.71 ± 0.19 mg L−1, 0.04 ± 0.03 mg L−1, and 24.0 ± 20.18 μg L−1, respectively. The mean water depth where macrophyte sampling occurred and its associated SD was 2.0 ± 0.72 and 0.9 ± 0.45 m, respectively. The mean light extinction coefficient in water was 2.47 ± 1.09 m−1 (Table 3).
Stoichiometric traits of submerged plant tissues
In our study, the average mass ratio of C:N:P in plant tissues was 191:12:1. The mean concentrations of C, N, and P were 380.5 ± 26.60, 25.1 ± 5.44, and 2.50 ± 1.25 mg g−1, respectively (Table 4). The coefficient of variation (CV) of C, N, and P concentrations was 7.0, 21.7, and 50.0 %, respectively. The mean ratio of C:N was 16.0 ± 4.07, the mean ratio of C:P was 191.3 ± 96.63, and the mean ratio of N:P was 11.89 ± 5.01. The lowest N and P concentrations were observed in P. malaianus, whereas the highest N and P concentrations were observed in C. demersum and H. verticillata, respectively. We also found that P. malaianus and M. spicatum had higher C:N and C:P than other submerged macrophyte species.
Effects of taxonomy, sediment, and water column on plant tissue stoichiometry
Sediment P concentrations had significant relationships with P, C:P, and N:P in plant tissue (all p < 0.01), whereas the concentrations of sediment C and sediment N had no significant relationship with all plant stoichiometric signatures (all p > 0.05) (Table 5). As for the factors in water column, both WD and K had significant relationships with P in plant tissues (all p < 0.01), whereas concentrations of TN and TP in water columns had no significant relationship with that of P in plant tissues (all p > 0.05).
The results of RDA were significant (df = 20, F = 7.00, p < 0.001) to explain C, N, and P stoichiometric signatures of plant tissues (Fig. 2). The whole model explained 46.4 % variation in total (adjusted explained variation was 39.7 %). Taxonomy, sediment P, WD, K, and SD:WD have high explanations of plant stoichiometry at significance of 0.001 level (Table 3).

RDA plot of tissue C:N:P stoichiometric signatures and explaining variables
The results of variance partitioning based on pRDA showed that taxonomy, sediment, and water column explained 18.6 %, 6.9 %, and 8.5 % of total variance of plant stoichiometry, respectively (Fig. 3 and Table 6). The interaction effects of sediment and water column explained 7.9 % of plant stoichiometric variance. Thus apart from taxonomy, factors in water column had a higher influence on plant C, N, and P stoichiometry.

Variance partitioning based on pRDA analysis for sediment, water column, and taxonomy on tissue C:N:P stoichiometric signatures
Discussion
C:N:P stoichiometric characteristics of submerged macrophytes
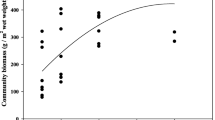
In our study, the mean C:N:P mass ratio of macrophytes was approximately 191:12:1, lower than (except N:P ratio) the ratio of growth rate limitation (272:11:1) reported by Demars and Edwards (2007) for macrophytes in the River Spey in Great Britain. The mean concentration of tissue C in our study was much lower than 438 mg g−1 reported by Demars and Edwards (2007). This result was consistent with the previous study in eastern China and mid-lower Yangtze floodplain lakes (Xia et al. 2014; Xing et al. 2013). The low concentrations of C in macrophytes implied that metabolism and growth may be hampered by the local low C availability. In a community level, tissue C concentration could be used to examine patterns such as variation in productivity and community structure. In the present study, a significant positive relationship (r = 0.49, p < 0.001) was found between community C concentration and macrophyte community biomass (Fig. 4). Therefore, it was suggested that the decline of macrophyte community in Yangtze floodplain shallow lakes was associated with the local low C availability in macrophytes. We inferred this was triggered by low light levels because light could alter the capacity of C fixation by changing the intensity of photosynthesis. This result could potentially occur at the species level and the community level simultaneously. At the species level, biomass of submerged macrophyte species decreased as a result of low capacity of C fixation in low light availability. At the community level, macrophyte species declined generally from high C concentration species to low C concentration species. Previous study (Qiu and Wu 1997) had revealed that succession of submerged macrophyte communities in the process of eutrophication of the Yangtze floodplain shallow lakes is from Potamogeton to species dominated by M. spicatum, V. natans, and C. demersum. This succession pattern supported our result well because C concentration in Potamogeton was significantly higher than that of V. natans and C. demersum (Table 4).

Significant positive relationship between community C concentration and macrophyte community biomass. Macrophyte community biomass was log10 transformed before analysis
Our results did not support the notion that plants were limited by nutrient availability because the obtained tissue N and P concentrations were much higher than the critical concentrations of 13 and 1.3 mg g−1, respectively (Gerloff and Krombholz 1966). Furthermore, significant positive relationships between sediment P and plant tissue P concentration were observed. This suggested that plants would accumulate P even though the P availability in environment was sufficient (Demars and Edwards 2007; Garbey et al. 2004; Rattray et al. 1991). This “luxury” uptake could be an adaptive strategy which would benefit the plant once the limited factors were relieved. Therefore, P concentration of submerged vegetation was generally more variable than the N concentration, which is consistent with previous reports (Fernández-Aláez et al. 1999; Güsewell and Koerselman 2002).
High interspecific variations of C:N:P stoichiometry were found between the six species. The species-specific variation of nutrient composition can be an important trait for identifying the ecological strategies and for predicting the result of competition in plant communities (Tilman 1982). V. natans, for instance, had lowest C concentrations which benefited its tolerance in low light stress and was consistent with its low light compensation point of photosynthesis (Su et al. 2004; Titus and Adams 1979). In addition, V. natans had lower C/N metabolic level and higher carbohydrate storage than other submerged macrophytes (Yuan et al. 2016). On the contrary, P. malaianus and M. spicatum, belonging to canopy formers, had higher C:N and C:P than other submerged macrophyte species. This might be another adaptive strategy which is the result of allocating more C on stem to elongate their shoots toward water surface (Ni 2001; Titus and Adams 1979). C. demersum is a submerged, rootless, free-floating aquatic plant. Unlike rooted vegetation, it requires nutrient uptake from the water (Denny 1987). However, a significant positive relationship (r = 0.41, p = 0.027) was still observed between sediment P and tissue P in C. demersum. This result implied that sediment might play a role in nutrient uptake in C. demersum because it may attach itself to the sediment sometimes. Besides, our results showed that tissue C concentration of C. demersum generally had a higher coefficient of variation than that of other five submerged macrophytes. And previous study (Li et al. 2013) also revealed that the N and P concentrations in leaves and stems of C. demersum were relatively unstable. These results indicated that C. demersum had a low stoichiometric homoeostasis.
Sources of variability in aquatic plant tissue stoichiometry
The results of pRDA revealed that both the sediment and water column had significant effects on C, N, and P concentrations and C:N:P stoichiometry. However, the explained variation was low compared to the variation explained by species composition, indicating that taxonomic effect was more important than that of sediment and water column at the community level. High interaction effects between sediment and water column in explaining stoichiometric variation were observed in this study. It was not surprising because the two components were usually not independent. For instance, resuspension and nutrient release from sediment could alter light attenuation coefficient and stoichiometry of N and P in water column. Plant tissue P had significant positive relationship with sediment P, while it had no significant relationship with TP in water column, implying that submerged macrophyte uptake P was generally from sediment as mentioned in the “Introduction” (Barko and Smart 1981; Best and Mantai 1978; Carignan and Kalff 1980). However, this cannot be regarded as direct evidence that dominance of nutrient uptake cannot be mediated by leaves, especially in oligotrophic streams. For instance, there is no correlation between the total P concentration in sediment and tissues, but a positive relation to water P concentration was observed in Alsace Rhine streams, supporting the view that leaf uptake can be the main pathway for nutrient uptake in some streams (Robach et al. 1995). In water column, our study showed that water depth has a significant negative effect on plant tissue P which is consistent with previous study (Li et al. 2015). Indeed, in deep water, most submerged macrophytes tend to allocate more biomass to stem for shoot elongation to alleviate low light availability (Fu et al. 2012; Strand and Weisner 2001). Changes of this biomass allocation could lead to low tissue P and high C:P because the structural organs generally have higher C and lower P than leaves (Li et al. 2013).
In the present study, sediment had a lower effect on plant C:N:P stoichiometry than water column. All explanations of C:N:P stoichiometric signatures except N:P explained by water column were higher than that of sediment, even though sediment supplies major nutrient to the plants (Best and Mantai 1978; Carignan and Kalff 1980; Smith and Adams 1986). In addition, only factors associated with light availability (SD, K, and SD:WD) in water column were significantly correlated with plant C concentrations. These results suggested that growth of submerged macrophytes in Yangtze floodplain lakes was primarily limited by low light stress rather than nutrient availability. With the aggravation of eutrophication, we infer that this situation will be even worse than the present.
Conclusions
This study revealed the relative importance of taxonomy, sediment, and water column on the C:N:P stoichiometry of submerged macrophytes in Yangtze floodplain shallow lakes. Results showed that submerged macrophytes had a species-specific identity in determining C:N:P stoichiometry. Apart from taxonomy, water column had higher explanations on plant stoichiometry than that of sediment. In the present study, a significant positive relationship was found between community C concentration and macrophyte community biomass, indicating that the local low C availability was probably the main reason why submerged macrophytes declined in Yangtze floodplain shallow lakes. Parameters of WD, K, and SD:WD associated with light availability had significant effects on C:N:P stoichiometry. It is suggested that submerged macrophytes in Yangtze floodplain shallow lakes are primarily limited by low light levels rather than nutrient availability.
References
Agren GI (2008) Stoichiometry and nutrition of plant growth in natural communities. Annu Rev Ecol Evol Syst:153–170
Andersen T, Elser JJ, Hessen DO (2004) Stoichiometry and population dynamics. Ecol Lett 7:884–900
Barko JW, Smart RM (1981) Sediment-based nutrition of submersed macrophytes. Aquat Bot 10:339–352
Bedford BL, Walbridge MR, Aldous A (2008) Patterns in nutrient availability and plant diversity of temperate North American. Wetlands Ecology 80:2151–2169
Best MD, Mantai KE (1978) Growth of Myriophyllum: sediment or lake water as the source of nitrogen and phosphorus. Ecology 59:1075–1080
Bristow JM, Whitcombe M (1971) The role of roots in the nutrition of aquatic vascular plants. Am J Bot 58:8–13
Cao T, Ni L, Xie P, Xu J, Zhang M (2011) Effects of moderate ammonium enrichment on three submersed macrophytes under contrasting light availability. Freshw Biol 56:1620–1629
Carignan R, Kalff J (1980) Phosphorus sources for aquatic weeds: water or sediments? Science 207:987–989
Chambers PA, Kalff J (1987) Light and nutrients in the control of aquatic plant community structure. I In situ experiments Journal of Ecology 75:611–619
Cronin G, Lodge DM (2003) Effects of light and nutrient availability on the growth, allocation, carbon/nitrogen balance, phenolic chemistry, and resistance to herbivory of two freshwater macrophytes. Oecologia 137:32–41
Demars BOL, Edwards AC (2007) Tissue nutrient concentrations in freshwater aquatic macrophytes: high inter-taxon differences and low phenotypic response to nutrient supply. Freshw Biol 52:2073–2086
Denny P (1987) Mineral cycling by wetland plants—a review. Arch Hydrobiol Beih 27:1–25
Elser JJ et al. (2000) Biological stoichiometry from genes to ecosystems. Ecol Lett 3:540–550
Elser JJ, Urabe J (1999) The stoichiometry of consumer-driven nutrient recycling: theory, observations, and consequences. Ecology 80:735–751
Evans-White MA, Lamberti GA (2006) Stoichiometry of consumer-driven nutrient recycling across nutrient regimes in streams. Ecol Lett 9:1186–1197
Fernández-Aláez M, Fernández-Aláez C, Bécares E (1999) Nutrient content in macrophytes in Spanish shallow lakes. Hydrobiologia 408-409:317–326
Fu H, Yuan G, Cao T, Ni L, Zhang M, Wang S (2012) An alternative mechanism for shade adaptation: implication of allometric responses of three submersed macrophytes to water depth. Ecol Res 27:1087–1094
Güsewell S, Gessner MO (2009) N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms. Funct Ecol 23:211–219
Güsewell S, Koerselman W (2002) Variation in nitrogen and phosphorus concentrations of wetland plants. Perspectives in Plant Ecology, Evolution and Systematics 5:37–61
Garbey C, Murphy K, Thiébaut G, Muller S (2004) Variation in P-content in aquatic plant tissues offers an efficient tool for determining plant growth strategies along a resource gradient. Freshw Biol 49:346–356
Gerloff G, Krombholz P (1966) Tissue analysis as a measure of nutrient availability for the growth of angiosperm aquatic plants. Limnol Oceanogr 11:529–537
Gusewell S, Bailey K, Wj BB (2005) Nutrient limitation and botanical diversity in wetlands: can fertilisation raise species richness? Oikos 109:71–80
Hall SR, Smith VH, Lytle DA, Leibold MA (2005) Constraints on primary producer N:P stoichiometry along N:P supply ratio gradients. Ecology 86:1894–1904
Harpole WS et al. (2011) Nutrient co-limitation of primary producer communities. Ecol Lett 14:852–862
Huang X, Chen W, Cai Q (1999) Survey, observation and analysis of lake ecology standard methods for observation and analysis in Chinese Ecosystem Research Network, Series V. Standards Press of China, Beijing in Chinese
Krause-Jensen D, Sand-Jensen K (1998) Light attenuation and photosynthesis of aquatic plant communities. Limnol Oceanogr 43:396–407
Li W, Cao T, Ni L, et al. (2013) Effects of water depth on carbon, nitrogen and phosphorus stoichiometry of five submersed macrophytes in an in situ experiment. Ecol Eng 61:358–365
Li W, Cao T, Ni L, et al. (2015) Size-dependent C, N and P stoichiometry of three submersed macrophytes along water depth gradients. Environmental Earth Sciences 74:3733–3738
Madsen TV, Cedergreen N (2002) Sources of nutrients to rooted submerged macrophytes growing in a nutrient-rich stream. Freshw Biol 47:283–291
Manly BF (2006) Randomization, bootstrap and Monte Carlo methods in biology, 3rd edn. Chapman & Hall/CRC, London
Ngai JT, Jefferies RL (2004) Nutrient limitation of plant growth and forage quality in Arctic coastal marshes. J Ecol 92:1001–1010
Ni L (2001) Growth of Potamogeton maackianus under low-light stress in eutrophic water. J Freshw Ecol 16:249–256
Qiu D, Wu Z (1997) On the decline and restoration of submerged vegetation in eutrophic shallow lakes. Journal of Lake Science 9:82–88
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Rattray M, Howard-Williams C, Brown J (1991) Sediment and water as sources of nitrogen and phosphorus for submerged rooted aquatic macrophytes. Aquat Bot 40:225–237
Reich PB, Walters MB, Ellsworth DS (1997) From tropics to tundra: global convergence in plant functioning. Proc Natl Acad Sci 94:13730–13734
Robach F, Hajnsek I, Eglin I, Trémolières M (1995) Phosphorus sources for aquatic macrophytes in running waters: water or sediment? Acta Botanica Gallica 142:719–731
Smith CS, Adams MS (1986) Phosphorus transfer from sediments by Myriophyllum spicatum. Limnology & Oceanography 31:1312–1321
Sparks DL et al (1996) Methods of soil analysis. Part 3—Chemical methods. Soil Science Society of America, Madison, pp 869–919
Sterner RW, Hessen DO (2003) Algal nutrient limitation and the nutrition of aquatic herbivores. Annual Review of Ecology & Systematics 25:1–29
Strand JA, Weisner SE (2001) Morphological plastic responses to water depth and wave exposure in an aquatic plant (Myriophyllum spicatum). J Ecol 89:166–175
Su W, Zhang G, Zhang Y, Xiao H, Xia F (2004) The photosynthetic characteristics of five submerged aquatic plants. Acta hydrobiologica Sinica 28:391–395
Tibbets TM, Molles MC (2005) C:N:P stoichiometry of dominant riparian trees and arthropods along the Middle Rio Grande. Freshw Biol 50:1882–1894
Tilman D (1982) Resource competition and community structure. Princeton University Press, Princeton
Titus JE, Adams MS (1979) Coexistence and the comparative light relations of the submersed macrophytes Myriophyllum spicatum L. and Vallisneria americana Michx. Oecologia 40:273–286
Willby NJ, Pulford ID, Flowers TH (2001) Tissue nutrient signatures predict herbaceous-wetland community responses to nutrient availability. New Phytol 152:463–481
Xia CX, Yu D, Wang Z, Xie D (2014) Stoichiometry patterns of leaf carbon, nitrogen and phosphorous in aquatic macrophytes in eastern China. Ecol Eng 70:406–413
Xing W, Wu H, Hao B, Liu G (2013) Stoichiometric characteristics and responses of submerged macrophytes to eutrophication in lakes along the middle and lower reaches of the Yangtze River. Ecol Eng 54:16–21
Yuan G, Fu H, Zhong J, et al. (2016) Growth and C/N metabolism of three submersed macrophytes in response to water depths. Environ Exp Bot 122:94–99
Acknowledgments
This work was supported by grant from the State Key Laboratory of Freshwater Ecology and Biotechnology (grant number 2014FBZ02).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Additional information
Responsible editor: Thomas Hein
Rights and permissions
About this article

Cite this article
Su, H., Wu, Y., Xie, P. et al. Effects of taxonomy, sediment, and water column on C:N:P stoichiometry of submerged macrophytes in Yangtze floodplain shallow lakes, China. Environ Sci Pollut Res 23, 22577–22585 (2016). https://doi.org/10.1007/s11356-016-7435-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-016-7435-1