
Abstract
Heavy metals cause serious problems in the environment, and they can be accumulated in organisms, especially in the higher fungi. The concentration of Ni, Cr, Pb, Cd, and Hg in 10 species of edible mushrooms in Medvednica Nature Park, Croatia was therefore determined. In addition, the similarity between the studied species was determined by cluster analysis based on concentrations of the aforementioned metals in the fruiting bodies. The contents of nickel, chromium, lead, cadmium, and mercury in the fruiting bodies of mushrooms were obtained by X-ray fluorescence spectrometry. The highest concentrations of Ni (3.62 mg kg−1), Cr (3.01 mg kg−1), and Cd (2.67 mg kg−1) were determined in Agaricus campestris. The highest concentration of Pb (1.67 mg kg−1) was determined in Macrolepiota procera, and the highest concentration of Hg (2.39 mg kg−1) was determined in Boletus edulis. The concentration of all heavy metals significantly differed (p < 0.001) between examined saprophytic and ectomycorrhizal mushrooms. Considering anatomical part of the fruiting body (cap-stipe), a considerably higher concentration of the analyzed elements was found in the cap for all mushroom species. According to calculated bioconcentration factors, all the examined species were found to be bioexclusors of Ni, Cr, and Pb and bioaccumulators of Cd and Hg. Cluster analysis performed on the basis of the accumulation of the studied metals revealed great phenotypic similarity of mushroom species belonging to the same genus and partial similarity of species of the same ecological affiliation.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Environmental pollution is one of the most serious problems in industrialized countries. Emissions of heavy metals from anthropogenic sources have been constantly increasing in recent decades. Heavy metals are very persistent in the environment and, due to the ability of accumulation, threat to living organisms. Mushrooms are known to accumulate high concentrations of toxic metallic elements, metalloids, and radio-nuclides (Kalač 2001; Vetter 2004; Svoboda et al. 2006). Mushrooms are a distinct group of living organisms of considerable nutritional, pharmaceutical, and ecological value. They play a vital role in the majority of ecosystems in the biosphere because they are able to biodegrade the substrate on which they grow. Also, many low-grade agricultural waste products can be used for cultivation of saprophytic mushrooms, including the stalks of agricultural produce, corn cobs, cotton shells, sugarcane segments, etc. Fruiting bodies of mushrooms are appreciated for their chemical (Işıldak et al. 2004) and nutritional properties (Manzi et al. 1999) and also for texture and flavor (Ouzouni et al. 2009).
The content of metallic elements in many mushroom species is considerably higher (Kalač 2010) than in fruits and vegetables (Turkdogan et al. 2003). This suggests that mushrooms have an effective mechanism to facilitate the adsorption of heavy metals from the ecosystem (Turkekul et al. 2004), though the mechanism of adsorption is still not known. Mushrooms are intimately involved in soil mineral weathering and element cycling in the critical zone (Amundson et al. 2007). Their mycelia colonize both organic and mineral soils and produce a variety of active chemical compounds including enzymes (Baldrian 2008) and various organic acids (van Schöll et al. 2006). Production of enzymes in saprotrophic macrofungi plays an important role in the transformation of humic substances including humus formation and mineralization (Baldrian 2008) and mobilization of metals associated with organic matter. Apart from mycelium, rhizomorphs, which are root-like structures and which are known to be spread by species over a huge area of forest, also take part in the uptake of mineral elements (Falandysz and Borovička 2013). The contribution of rhizomorphs as a highly efficient medium for water and mineral compound uptake from soils seems to explain the presence of Hg in fruiting bodies of Armillaria solidipes (Falandysz et al. 2012). An individual organism of Armillaria bulbosa could spread the mycelium and rhizomorphs over 150,000 m2 of forest (Smith et al. 1992). The density and depth of the mycelium, which lives in the soil for several months or years, influence the metal content in fruiting bodies (García et al. 2009). In addition, species of mushroom various environmental factors and soil properties (pH, organic matter, redox potential, type of substrate, geochemistry of substrate, distance from the source of pollution, etc.) can affect the metal content in mushrooms (Nikkarinen and Mertanen 2004; García et al. 2009; Aloupi et al. 2012; Garcia et al. 2013; Petkovšek and Pokorny 2013). The relationship between abundance and bioavailability of heavy metals from the substrate is very complex and still not known (Kalač 2010). However, by determining the heavy metal content in mushrooms and the substrate on which they grow, and by calculating the factor of bioconcentration, it is possible to determine the suitability of using mushrooms as bioindicators of environmental pollution (Falandysz et al. 2007).
Nickel, chromium, lead, cadmium, and mercury can serve as reliable indicators of environmental pollution. Trace amounts of nickel are beneficial, since they activate some enzyme systems, while their higher levels can lead to serious toxicity. Numerous works (Işıldak et al. 2004; 2007; Soylak et al. 2005; Ouzuni et al. 2007; Sarikurkcu et al. 2011; Aloupi et al. 2012; Chudzynski et al. 2013; Drewnowska and Falandysz 2015; Falandysz and Drewnowska 2015; Stefanović et al. 2016) reported usual nickel content from traces to 15 mg kg−1, while the highest reported level was 58.60 mg kg in Coprinus comatus (Yamaç et al. 2007). In an experiment with Macrolepiota procera, Baptista et al. (2009) reported that nickel uptake increased proportionally to the metal increase in the substrate. Cr is a metallic element that can exist in several oxidation states; the trivalent and hexavalent states are the most important biological forms (Garcia et al. 2013). It is the tenth most abundant element in the earth’s mantle, and it is used extensively for several industrial purposes in both its tri- and hexavalent forms, depending upon the final use of the end product; these industrial applications include electroplating, timber treatment, pulp production, mineral ore, and petroleum refining (Wang et al. 2011). Wild growing mushrooms frequently contain Cr in a concentration from traces to 5 mg kg−1, less frequently between 5 and 10 mg kg−1 (Kalač 2010; Li et al. 2011; Aloupi et al. 2012; Chudzynski et al. 2013; Garcia et al. 2013; Kuldo et al. 2014; Širić et al. 2014; Drewnowska and Falandysz 2015; Falandysz and Drewnowska 2015; Stefanović et al. 2016). Lead is a heavy metal toxic to plants, animals, and humans, and there is no evidence of its biological role in the human body. According to results from papers published until 2016, the usual lead content in most species from unpolluted areas ranges between 0.4 and 5 mg kg−1. In addition, there are also some species of mushroom with extremely high potential for lead accumulation, such as Agaricus campestris, Rozites capertus, Lepista nuda, and M. procera (Kalač and Svoboda 2000; García et al. 2009; Ouzouni et al. 2009; Kalač 2010; Chudzynski et al. 2013; Petkovšek and Pokorny 2013; Drewnowska and Falandysz 2015). Most likely, cadmium is the most commonly studied trace element in mushrooms. Wild-grown mushrooms frequently contain cadmium in a concentration between 0.10 and 7.50 mg kg−1 (Svoboda et al. 2000; Malinowska et al. 2004; Mendil et al. 2004; Cocchi et al. 2006; Yamaç et al. 2007; Kalač 2010; Kojta et al. 2011; Chudzynski et al. 2013; Petkovšek and Pokorny 2013; Drewnowska and Falandysz 2015). The cadmium concentration in mushrooms sampled in unpolluted areas rarely exceeds 10 mg kg−1. However, this concentration is often exceeded in polluted areas (Falandysz and Borovička 2013). Mercury is one of the most toxic and dangerous environmental contaminants and can be efficiently bioaccumulated by many mushrooms (Amanita sp., Armillariela sp., Boletus sp., Coprinus sp., Cortinarius sp., Leccinum sp., Macrolepiota sp., Suillus sp., Xerocomus sp.), even if it is scarcely present in forest soils. For others, biconcentration factors can vary from genus: Amanita 1.2 to 8.7 (Drewnowska et al. 2014), Boletus 7.3 to 130 (Falandysz et al. 2014), 126 to 421 (Melgar et al. 2009), and 2.9 to 71 (Falandysz et al. 2015a), Coprinus 9.5 to 74 (Falandysz 2015), Cortinarius 22 to 150 (Falandysz 2014), Leccinum 2.3 to 35 (Falandysz et al. 2015b), 4.6 to 24 (Krasinska and Falandysz 2015), and 3.4 to 65 (Krasinska and Falandysz 2016), Macrolepiota 16 to 220 (Falandysz and Borovička 2013) and 52 to 140 (Kuldo et al. 2014), Suillus 3.6 to 42 (Saba et al. 2016), and Xerocomus 1.3 to 22 (Chojnacka et al. 2012) and 1.4 to 18 (Dryzalowska and Falandysz 2014). Species of the aforementioned genus are capable of accumulating several fold greater amounts of mercury than any other mushrooms.
Mushroom picking is very popular in Central and Southern Europe, as well as in Croatia (Širić et al. 2014). Medvednica Nature Park is located near the largest urban and industrial center in Croatia, the capital city Zagreb, and there may be increased concentrations of heavy metals in mushrooms. Till now in Medvednica Nature Park, 81 species of mushrooms have been identified date but there has been no study on their metal contents. The objectives of this study were to (i) determine the heavy metal content in wild-growing edible mushroom species and the substrate on which they grow, (ii) determine the accumulation capacity (bioconcentration or exclusion) of heavy metals in fruiting bodies of mushrooms, (iii) determine the distribution of analyzed elements in anatomical parts of fruiting bodies (cap and stipe), and (iv) perform cluster analysis on the mushroom species in relation to their metal content.
Material and methods
Sampling of mushrooms
Sampling was done from September to December 2012 in Medvednica Nature Park, in the north-western part of Zagreb County, Croatia. Within the sampling area, the most prevalent tree species were Quercus sp., Carpinus betulus L., Castanea sativa Mill., Fagus sylvatica L., Picea abies L., Acer pseudoplatanus L., and Fraxinus excelsior L. Levels of heavy metals (Ni, Cr, Pb, Cd, and Hg) were determined in ten edible mushroom species (20 samples per species). Among the sampled species, there were four terrestrial saprophytes (Agaricus campestris (L) Fries, Clitocybe inversa (Scop. ex Fr.) Pat., Clitocybe nebularis Batsch. ex Fr., and M. procera (Scop. ex Fr.) Sing.), one lignicoluous saprophyte (Armillaria mellea (Vahl. ex Fr.) Karst), and five ectomycorrhizal species (Boletus aestivalis Paulet ex Fries, Boletus edulis Bull. ex Fries, Lactarius deterrimus Groger, Tricholoma portentosum (Fr.) Quelet, and Tricholoma terreum (Schff. ex Fr.) Kummer) (Bozac 2005, 2008). The types of fungi are selected for the frequency of the geographical area of research, culinary quality, and commercialization. Completely developed and mature fruiting bodies of the investigated mushrooms were collected randomly. At the same time, soil samples of the forest upper soil horizon (0–10 cm) were collected at appropriate sampling places, according to Garcia et al. (2013). All samples were analyzed in triplicate.
Analysis of heavy metals
Collected specimens were documented, oven dried at 60 °C to constant mass, milled with a laboratory Retch SM2000, and pressed into tablets (r = 16 mm; d = 5 mm) with a Chemplex press for further analysis. X-ray analysis was done by fluorescence spectroscopy (XRF, TwinX, Oxford instruments). In the first step, qualitative analysis was performed. The most frequent pollutants were identified. Calibration curves were prepared for those elements and, in the second step, quantitative analysis was carried out. For calibration curves, Pleurotus ostreatus fruiting bodies and mycelia were used. Fruiting bodies were cultivated on controlled mycelium. This material was homogenized and mixed with laboratory Retch SM2000 mill. In order to obtain standards with known concentrations, milled fungal material (10 g) was supplemented with aqueous solutions of targeted concentrations. Ratio between fungal material and aqueous solution used was 1: 3 (w/w). This enables that all material used for production of respective standard was homogenously treated. Concentration range was determined based on the preliminary data. Each standard contains between two up to five respective elements in order to address possible interference. Treated fungal material was than conditioned for 24 h and afterwards oven dried (60 °C, 48 h). Concentration in standards and analyzed material was verified with atomic absorption spectroscopy. Afterwards, calibration curves were afterwards prepared and verified with built-in software. Most of the measurements were performed on a PIN detector (U = 26 kV, I = 115 μA, t = 300 s). However, mercury concentration was determined with a proportional detector in a helium atmosphere. The values of bioconcentration factors were calculated as a ratio between the heavy metal contents in the mushroom and the element concentrations in the soil in which the mushroom grew.
Soil pH and organic matter
The pH value of the soil substrate was determined potentiometrically in a suspension of the soil substrate and distilled water in a ratio of 1: 5. Measurement was carried out according to methods adapted from “Methods of soil analysis” (Thomas 1996). The pH value was determined using pH meter IQ 150 (IQ Scientific Instruments, USA). Organic matter content was determined gravimetrically after combustion of the soil (2 g air dried) organic matter at 550 °C for 16 h in a furnace horn (Select-Horn Selecta) (García et al. 2009).
Data analysis
Statistical analysis and all chartings were performed within the R program (R Core Team 2014) by using two integral and three external statistical packages. Descriptive statistics, calculation of bioconcentration factors, and pairwise comparisons (t test) of means (concentrations of trace elements) between anatomical parts of the fruiting body were obtained within the package “stats.” Multiple pairwise comparisons (Tukey’s HSD test) of means among species were obtained by the “agricolae” package (de Mendiburu 2014). Extraction of the information required to create and plot compact letter displays of all pairwise comparisons was performed within the “multcomp” package (Hothorn et al. 2008). Plotting of box-whisker plots was performed with the packages “graphics” (integral) and “lattice” (Sarkar 2008).
Cluster analysis was performed within the “stats” package. The distance matrix was computed by using the “Euclidian” distance measure, and hierarchical cluster analysis was performed by using a method of complete linkage, which defines the distance between two clusters to be at the maximum distance between their individual components. At every stage of the clustering process, two nearest clusters were merged into a new cluster, and this process was repeated until the whole data set was agglomerated into a single cluster. The results of cluster analysis were converted into a “phylo” object within the “ape” package (Paradis et al. 2004) and thereafter presented graphically as polar dendograms.
Results
Soil factors
Soil properties (pH value and organic matter content) and average concentrations of heavy metals Ni, Cr, Pb, Cd, and Hg in the area of Medvednica are summarized in Table 1. The mean pH value of the soil substrate at Medvednica was 7.22, ranging between 6.30 and 8.12. Organic matter content of the soil ranged from 2.16 to 12.65 %, with a mean value of 6.48 %. The results of analysis of heavy metal concentrations in the soil substrate show that Cr (17.24–21.40 mg kg−1) and Pb (13.25–20.10 mg kg−1) had the highest content, followed by Ni (11.55–18.31 mg kg−1) and Cd (0.46–0.72 mg kg−1), while Hg (0.08–0.10 mg kg−1) have the lowest content.
Metal concentration and bioconcentration factors
Descriptive statistics on heavy metal concentration and accompanying factors of bioconcentrations (BCF) are given in Table 2.
In general, concentrations of Ni were low and varied among the tested species (Table 2 and Figs. 1 and 2). Values oscillated between 2.34 in the ectomycorrhizal species T. terreum and 3.62 mg kg−1 in A. campestris, a saprophyte species that lives in contact with mineral particles in the soil. The values of BCF were below 1 in all tested mushrooms (Table 2).

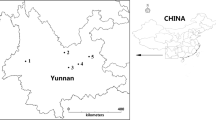
Area of the sampling of mushrooms species in Nature Park Medvednica, Croatia

Box and whisker plots representing the distribution of Ni concentration in 10 mushroom species. The box represents the interquartile range (from first to third), solid line within the box represents the median (or second quartile), and the whiskers represent the extremes of the distribution. Letters represent the results of Tukey’s post hoc comparisons of mean values among the species
The mean Cr concentration in the 10 species of wild edible mushroom was 2.03 mg kg−1. In terms of species, the highest mean chromium concentrations of 3.01 and 2.92 mg kg−1 were found in A. campestris and M. procera, respectively, and the lowest concentrations in T. portentosum (0.88) and T. terreum (0.97) (Fig. 3, 4, 5, and 6). Similarly to nickel, all analyzed species of mushroom had values of BCFs <1.

Box and whisker plots representing the distribution of Cr concentration in 10 mushroom species. The box represents the interquartile range (from first to third), solid line within the box represents the median (or second quartile), and the whiskers represent the extremes of the distribution. Letters represent the results of Tukey’s post hoc comparisons of mean values among the species

Box and whisker plots representing the distribution of Pb concentration in 10 mushroom species. The box represents the interquartile range (from first to third), solid line within the box represents the median (or second quartile), and the whiskers represent the extremes of the distribution. Letters represent the results of Tukey’s post hoc comparisons of mean values among the species

Box and whisker plots representing the distribution of Cd concentration in 10 mushroom species. The box represents the interquartile range (from first to third), solid line within the box represents the median (or second quartile), and the whiskers represent the extremes of the distribution. Letters represent the results of Tukey’s post hoc comparisons of mean values among the species

Box and whisker plots representing the distribution of Hg concentration in 10 mushroom species. The box represents the interquartile range (from first to third), solid line within the box represents the median (or second quartile), and the whiskers represent the extremes of the distribution. Letters represent the results of Tukey’s post hoc comparisons of mean values among the species
The determined content of lead in all mushroom species was lower than in the soil in which they grew (BFC < 1). Among the saprophytes, the highest concentrations of lead were determined in C. nebularis and M. procera, while among the ectomycorrhizal, two species of the genus Boletus have had the highest concentrations of lead (Table 2).
The range of cadmium in the fruiting bodies of tested mushrooms was 0.36–3.33 mg kg−1, while the highest mean value was determined in A. campestris (2.67) and the lowest in L. deterrimus (0.72). The values of bioconcentrations factor were above 1 (BCF > 1) in all analyzed species (Table 2), wherein the highest bioaccumulation potentials of 4.34 and 3.36 were calculated in the saprophytic (terrestrial) species A. campestris and M. procera.
Table 2 shows that mushroom species have varied ability of mercury accumulation. The highest concentrations of mercury were determined in B. edulis (2.39 mg kg−1). In contrast, the wood-decaying saprophytic fungi A. mellea had the lowest mercury content of 0.28 mg kg−1. All mushrooms species in this study accumulated mercury and BCF values were higher than one (Table 2). The highest BCF value of 26.67 was determined in B. edulis, which was 8.52 times higher than the lowest BCF of 3.13 found in A. mellea.
Morphological parts
The distributions of heavy metals between anatomical parts of the fruiting bodies (cap and stipe) of the investigated mushrooms are given in Table 3. In terms of the anatomical part of the fruiting body, a considerably higher accumulation of heavy metals was determined in caps than in stipes in all mushroom species.
Comparison among metal concentrations in saprophytic and ectomycorrhizal fungal species
The concentrations of heavy metals in mushrooms of different ecological habits are shown in Table 4. A statistically higher concentration (p < 0.001) of nickel, chromium, lead, and cadmium was determined in saprophytic species, while mercury had a higher concentration (p < 0.001) in ectomycorrhizal species.
Cluster analysis
A graphic presentation (dendogram) of hierarchical cluster analysis is shown in Fig. 7. Cluster analysis based on the accumulation of heavy metals revealed large similarity among species belonging to the same genus (Fig. 7). In addition to these within-genera similarities, great similarity based on accumulated heavy metals was found between A. mellea and L. deterrimus and between A. campestris and M. procera. Furthermore, the former were found to be more similar to members of the genus Tricholoma, while the latter were found to be more similar to members of the genera Boletus and Clitocybe.

Dendrogram of the cluster analysis on the mushrooms species in relation to their heavy metal contents
Discussion
The lowest nickel value was found in T. terreum, which is considerably higher than the results of Işıldak et al. (2007)). The highest content of nickel in our study was determined in A. campestris. This value is far above the one reported by Sarikurkcu et al. (2011) for the same species sampled in the national park in Turkey. For Ni, our results are a little higher than reported by Polish researcher, who obtained Ni concentrations range 0.82–2.20 mg kg−1 (Drewnowska and Falandysz 2015). For specimens collected in Sweden, this element was found at concentration range 0.56–4.00 mg kg−1, respectively (Jorhem and Sundstrom 1995). Ni concentration at the range of 1.05–6.3 mg kg−1 was obtained in recent Serbian studies (Stefanović et al. 2016), which is close to our results. Furthermore, the results of some research show that most received nickel was accumulated in the caps of fungi (Kalač 2010; Brzostowski et al. 2012; Jarzynska and Falandysz 2012; Chudzynski et al. 2013). The distribution of nickel between the cap and stipe in our study is in agreement in the abovementioned literature. However, available scientific literature does not mention the reason for the higher accumulation of nickel in the cap than in the stipe. A possible explanation is higher biological activity of proteins contained in the cap toward the stipe. Considering that the area of sampling (Nature park Medvednica) is in the vicinity of the main industrial centers in Croatia, it can be potential cause of atmospheric deposition heavy metals. In the present study, the values of BCFs for nickel were below 1 in all tested mushrooms, which is in agreement with the results of Aloupi et al. (2012), Gucia et al. (2012a), Garcia et al. (2013), and Drewnowska and Falandysz (2015). Accordingly, all investigated mushrooms are bioexclusors for nickel.
We determined considerable differences in chromium content among investigated species of mushrooms. The highest content of chromium in our study was found in A. campestris and M. procera but the lower value than those reported by Garcia et al. (2013). However, the authors reported the lowest mean levels in T. portentosum, which is equivalent to our results. In contrast, Işıldak et al. (2007)) showed considerably higher contents of chromium in species from the genus Tricholoma and as a main reason quote high concentrations of chromium in the tested soil samples (88.50 mg kg−1). Furthermore, Işıldak et al. (2004) reported a notably higher concentration of chromium in A. mellea than those found in this study (22.70 versus 2.82 mg kg−1). In studies on Parasol mushroom (M. procera), Jarzynska et al. (2011) reported Cr levels of 0.31 and 0.61 mg/kg dw in caps and stipes, respectively. Drewnowska and Falandysz (2015) also reported lower values of Cr in Cantharellus cibarius. The Cr levels were found to depend on the sampling site and were always less than 1 mg kg−1 dw, although these concentrations were still less than those obtained in the present work for the same species (2.92 mg kg−1 dw). However, Stefanović et al. (2016) reported contents of Cr in M. procera were higher than in this study and literature data (Jarzynska et al. 2011; Gucia et al. 2012a; Kuldo et al. 2014), for samples from site S1 (6.9 ± 4.6 and 5.6 ± 3.2 mg/kg dw for caps and stipes, respectively. Aloupi et al. (2012) argue that the concentration of chromium in mushrooms depends on the sampling site and the geochemistry of the substrate on which the mushrooms grow. In line with this, the authors state significantly higher (p < 0.001) concentrations in mushrooms growing on serpentine soils than if those collected on volcanic soils. With regard to the sampling areas, the mean chromium concentration in soil was 19.35 mg kg−1. The mean BCF value was 0.10, and all species presented BCF less than 0.2. These values indicated that bioexclusion of the mushrooms studied with regard to soil chromium concentration and are in agreement with Figueiredo et al. (2007) for several species (Agaricus sp., Amanita sp., Lactarius sp.): with Chudzynski and Falandysz (2008) for Suillus gravillei, with Garcia et al. (2013) for several species (Agaricus sp., Amanita sp., Boletus sp., Cantharellus sp., Clitocybe sp., Coprinus sp., Hydnum sp., Lactarius sp., Leccinum sp., Lepista sp., Macrolepiota sp., Russula sp., Tricholoma sp.), and with Drewnowska and Falandysz (2015) and Falandysz and Drewnowska (2015) for C. cibarius. This behavior was similar to that of nickel (Drewnowska and Falandysz 2015; Falandysz and Drewnowska 2015 and Stefanović et al. 2016) and lead (García et al. 2009; Drewnowska and Falandysz 2015; Falandysz and Drewnowska 2015 and Stefanović et al. 2016) and in contraposition to the bioaccumulative character of cadmium (Jarzynska et al. 2011; Gucia et al. 2012a; Kuldo et al. 2014; Stefanović et al. 2016) and mercury (Melgar et al. 2009; Chojnacka et al. 2012; Drewnowska et al. 2014; Dryzalowskaa and Falandysz 2014; Falandysz et al. 2014; Falandysz 2014; Kuldo et al. 2014; Falandysz 2015; Falandysz et al. 2015a; Falandysz et al. 2015b; Krasinska and Falandysz 2015; Saba et al. 2016; Krasinska and Falandysz 2016).
Results for lead in this study are in agreement with the most commonly reported results (García et al. 2009; Ouzouni et al. 2009; Kalač 2010; Petkovšek and Pokorny 2013). The lowest mean concentration of lead was found in saprophytic species A. mellea, which grows on wood without contact with mineral particles of soil, which is in agreement to results of Ouzouni et al. (2009) and Petkovšek and Pokorny (2013). Saprophytic species M. procera accumulates the highest mean concentration of lead (Table 2), and established values are in agreement with the reports of García et al. (2009) and Kalač (2010). Similar values of lead were determined 1.50 mg kg in M. procera (Kuldo et al. 2014). The lead content in M. procera was generally lower than values reported by Kojta et al. (2011), Gucia et al. (2012a), and Petkovšek and Pokorny (2013). In the study of Stefanović et al. (2016), results for Pb ranged from 0.24 to 1.22 mg/kg in caps for sites S2, S3, S4, and S5. These values are comparable with results of other researchers (Yamaç et al. 2007; Kuldo et al. 2014). However, Stefanović et al. (2016) reported much higher content of lead from site S1 (8.8 ± 4.5 mg/kg in caps and 5.4 ± 1.7 mg/kg for stipes) but also comparable with the results obtained for unpolluted areas in Poland both for caps (8.5 ± 2.4 mg/kg dw) and stipes (5.2 ± 2.1 mg/kg dw) (Gucia et al. 2012b). The median value of Pb in Finnish study was 1.71 mg kg−1 (Pelkonen et al. 2006). For the samples collected in Sweden, Pb concentration range was 0.14 to 3.3 mg kg−1 (Jorhem and Sundstrom 1995). Pb concentration for the samples collected from unpolluted regions of Poland was from 0.28 to 0.80 mg kg−1 (Drewnowska and Falandysz 2015) and 0.28 to 0.50 mg kg−1 (Falandysz and Drewnowska 2015). Extremly high Pb contents in mushrooms have been determined in the site Upper Meža Valley Slovenia due to past smelting and mining activities, in spite of significnt reductions of Pb emissions in the last two decades (Petkovšek and Pokorny 2013). This variation in lead contents may be attributed to differences in the area of sampling, mushroom species, and ecosystem. A considerably higher content of lead was determined in caps than in stipes (Table 3), which is in accordance with the results of Kojta et al. (2011) and Jarzynska et al. (2011). García et al. (2009) found extremely large differences in the concentration of lead between the hymenophore and rest of the fruiting body, which was especially pronounced in A. campestris, C. nebularis, M. procera, and T. portentosum. However, those differences were found to be statistically insignificant. Increased accumulation of lead in the hymenophore in their study was attributed to the biological activity of proteins contained in the hymenophore. The concentrations of lead in the studied mushrooms were considerably lower than the concentrations in the soil, which is in agreement with the results of Işıldak et al. (2007), García et al. (2009), Kojta et al. (2011), and Drewnowska and Falandysz (2015). Consequently, all calculated bioconcentration factors were below 1, which suggests that all studied species are bioexclusors of lead. A lower efficiency in the accumulation of lead at high concentrations may be due to the toxic effects of metal ions (García et al. 2009).
Cadmium levels determined in this study were within the above mentioned range, with the highest concentration determined in A. campestris, which is in agreement with the results of Petkovšek and Pokorny (2013). They found even higher concentrations (8.93 mg kg−1) when sampling was performed near smelting area. Cd contents in M. procera and Boletus sp., another accumulator of Cd and one of the most important wild-growing mushroom species for human consumption, were much lower than values reported for species collected in Slovenia (Petkovšek and Pokorny 2013) and Serbia (Stefanović et al. 2016). However, considerably lower concentrations of cadmium were found in Poland in C. cibarius species, 0.21–0.41 mg kg−1 (Falandysz and Drewnowska 2015) and 0.20–0.45 mg kg−1 (Drewnowska and Falandysz 2015), according to the site of sampling. The ranges of Cd given by Polish researchers were from 0.21 to 0.41 mg kg−1 (Falandysz and Drewnowska 2015) and from 0.20 to 0.45 mg kg−1 (Drewnowska and Falandysz 2015), according to the site of sampling. For samples collected in Sweden, this range was 0.4–2.57 mg kg-1 (Jorhem and Sundstrom 1995). Significant impact of sampling area on level of cadmium accumulation in mushroom also was found by Kalač and Svoboda (2000) and Sarikurkcu et al. (2011) with hyperaccumulation in mushrooms of the genus Agaricus (10–55 mg kg−1). The lowest cadmium content in this study was established in the ectomycorrhizal species L. deterrimus (0.76 mg kg−1), which is in agreement to the results of Ouzuni et al. (2007) and Aloupi et al. (2012). We suggest that the main cause of such low concentrations of cadmium in these ectomycorrhizal species (in contrast to saprophytes) is the deeper mycelium and symbiotic relationship with spruce (Picea excelsa) which accumulates quite amounts of the metal. A considerably higher concentration of cadmium in the cap than in the stipe determined in all examined species in this study is in major agreement with the results of Thomet et al. (1999), Kojta et al. (2011), and Stefanović et al. 2016). Therefore, all species of mushrooms were found to be bioaccumulators of cadmium, with a considerably higher BCF determined in the saprophytic species and the reason can be inherent properties of cadmium, which is a metal of the top soil and is therefore automatically more available for species which develop mycelium in the upper soil horizon. A. campestris and M. procera had the highest values of BCF, while L. deterrimus and T. terreum had the lowest. Our results are in agreement with values reported by Jarzynska et al. (2011) for the species M. procera. However, Kuldo et al. (2014) reported much higher values of BCF (45–50) in M. procera than in this study. Even though that cadmium is a frequently reported heavy metal in mushroom studies, there are only a few reports on its bioaccumulation potential. The high bioaccumulation potential for cadmium could be explained by high availability of cadmium in soil substrate and also the high potential of some species, e.g., M. procera for uptake of Cd from the substrate (Gucia et al. 2012a and Stefanović et al. 2016).
The results in our study are in accordance with those of Falandysz et al. (2001) and Melgar et al. (2009), who found the highest concentration of mercury in species of the genus Boletus. However, there is still no scientifically based explanation for this phenomenon, although Falandysz et al. (2007) consider that the tubes, which are part of the carpophore, are extremely rich with mercury. The wood-decaying saprophytic mushroom (A. mellea) had the lowest average mercury content, which is in agreement with the results of Żarski et al. (1999) and Falandysz et al. (2012). This is the most commonly attributed to the growth substrate (limited availability of metal ions in stump) of this lignicolous saprophyte. Higher deposition of mercury in the cap versus the stipe determined in this study is in agreement with the results of Melgar et al. (2009). The caps (hymenophore) may generally accumulate higher amounts of mercury because it contains more mercury-binding proteins and enzymes than the rest of fruiting body (Melgar et al. 2009). The BCF of all mushroom species for mercury was greater than 1, which implies that all investigated mushroom species are mercury bioaccumulators. Mentioned results are in major agreement with several published studies on this issue (Falandysz et al. 2007; Melgar et al. 2009; Kojta et al. 2011: Saba et al. 2016; Falandysz et al. 2015a). The Hg contents of soil and the availability of its Hg compounds from soil are a major factor impacting absorption and sequestration of this element in fruiting bodies by mushroom. The atmospheric deposition loads of Hg increased in the industrial era because of emissions from anthropogenic sources (Schuster et al. 2002; UNEP 2013), and this resulted in substantial enrichment of Hg in surface layer of forest soils. Hg deposited from atmosphere is highly retained in the surface layer (litter, fermentation layer, and humus) of soils and when complexed with some organic ligands a small component could also infiltrate into deeper layers of soil mineral horizon (Demers et al. 2007). Fruiting bodies are of ephemeral existence, while mycelium is a vegetative part of the fungus that can live for a long time. Hence, a passive absorption of Hg from the atmosphere by short-lived fruiting bodies, if any, is small; also, the Hg concentration in air in background (unpolluted) areas is small. However, considering that the area of sampling (Nature Park Medvednica) in the vicinity of the main industrial centers in Croatia, which can cause atmospheric deposition investigated metals. These results imply that analyzed fungi have the potential and can be used as a bio-indicator for Hg in soil substratum, especially genus Boletus.
In general, metal levels in fruiting bodies of wild mushrooms are influenced considerably by the age of mycelium, interval between fructifications, and the species of mushrooms. These factors cause wide variability in heavy metal concentrations. Transporter systems for uptake of essential metals are present in the cell membrane while non-essential metals are usually cotransported using the same transporters due to their low specificity. In the ectomycorrhizal fungus Paxillus involutus, the uptake of Cd involves its rapid binding to the cell wall and a slower carrier-mediated transport into the cell (Baldrian 2003). Also uptake partially depends on the membrane potential and is linked to the transport of Ca. On the subcellular level, 50 % of absorbed metals are bound to the cell wall, 30 % remain in the cytoplasm, and 20 % are transported into vacuoles (Blaudez et al. 2000). Furthermore, as reviewed by Bellion et al. (2006), saprophytes and probably mycorrhizal fungal species developed (evolved) mechanisms that enable them to tolerate metal concentrations excess in their environment. These mechanisms allow them to reduce the uptake of metals into the cytosol by extracellular chelation through extruded ligands and by binding into cell wall components. Further mechanisms are intracellular chelation of metals in the cytosol (e.g., by glutathione and metallothioneins), increased efflux from the cytosol out of the cell, or increased free radical-scavenging capacity. Molecular defense system, determined by the activity of antioxidant enzymes superoxide dismutase and catalase and by the content of glutathione and heat shock protein 70 kDa, was induced in B. edulis which grows in areas polluted with Cd, Zn, Cu, and Hg (Collin-Hansen et al. 2005a). It was shown that low concentrations of Hg and, probably, other three metals damaged DNA and lipids in B. edulis (Collin-Hansen et al. 2005b). In caps of the same mushroom species exposed to the excess of these four metals, phytochelatins, a family of cysteine-rich oligopeptides, were found. The efficiency of absorption of heavy metals in fungi in relation to the plant is performed with the specific proton pump that enables up to 10 times higher absorption of minerals in hyphae (Jennings 2007). Since fine fungal filaments called hyphae can grow beyond the root system, they help fungus to acquire mineral nutrients, more efficiently. Therefore, fungi can absorb heavy metals in very low concentrations, which are inaccessible for plants.
Grouping of the species based on the concentration of heavy metals in their fruiting body (hierarchical cluster analysis) revealed that species of the same genus have a similar ability to accumulate metals from their growing environment. Similar nutritional habits of species belonging to the same genus were probably the major reason for such consistent clustering at basic level. This clustering was completely in accordance with our expectations, bearing in mind that species within the same genus share some common physiological characteristics. Grouping of the species in separate clusters on the next level was more or less expected, since the majority of analyzed species (or even whole genera) were separated in clusters in accordance with their ecological affiliation (i.e., ectomycorrhizal and saprophytic species). However, this clustering was not completely consistent, since members of the genus Boletus were found to be more similar to the terrestrial saprophytes than to members of its own ecological affiliation. Moreover, members of the genus Clitocybe were found to resemble more to members of the genus Boletus than the terrestrial saprophytes (A. campestris and M. procera). Regarding the ecological habits of the examined species, the positioning of A. mellea within a cluster composed of ectomycorrhizal species can also be considered to be a partially unexpected result. This species is a lignicoluous saprophyte, and we therefore expected its separation in a single cluster, since this was the only representative of that ecological affiliation among all the examined species. However, this result is not completely surprising since lignicoluous saprophytes live and grow on the stumps of trees, which can limit their metal intake (Melgar et al. 2009). Such a positioning of lignicoluous saprophytes far away from terrestrial one is evidence that lignicoluous saprophytes more resemble to ectomycorrhizal species than the other saprophytic species (in terms of the bioaccumulation of the examined heavy metals). The results of cluster analysis were finally presented graphically as dendrograms.
Conclusion
The presented results are based on analysis of samples originating from Medvednica Nature Park and therefore provide additional data on the background concentrations of Ni, Cr, Pb, Cd, and Hg. The heavy metal contents in the mushrooms are mainly affected by species and their lifestyle. All mushrooms species were bioexclusors of nickel, chromium, and lead. On the other hand, bioaccumulation features were determined in all of the investigated mushroom species for the metals cadmium and mercury. The average concentrations of the investigated metals between the anatomical parts of the fruiting body (cap and stipe) were considerably different. The determined values of analyzed elements in mushrooms correspond to levels in unpolluted areas. Based on the determined concentration of metals in mushrooms and according to the regulations on maximum levels of certain contaminants in foodstuffs (Official Gazette 154/2008), it can be concluded that the consumption of the tested mushrooms cannot be considered to provide a toxicological risk from lead, cadmium, and mercury content for humans. However, to evaluate the possible danger of heavy metals on human health, their level should be analyzed more often in wild edible mushrooms.
References
Aloupi M, Koutrotsios G, Koulousaris M, Kalogeropoulos N (2012) Trace metal contents in wild edible mushrooms growing on serpentine and volcanic soils on the island of Levos, Greece. Ecotoxicol Environ Saf 78:184–194. doi:10.1016/j.ecoenv.2011.11.018
Amundson R, Richter DD, Humphreys GS, Jobbágy EG, Gaillardet J (2007) Coupling between biota and Earth materials in the critical zone. Elements 3:327–332. doi:10.2113/gselements.3.5.327
Baldrian P (2003) Interactions of heavy metals with white-rot fungi. Enzyme Mycrob Tech 32:78–91. doi:10.1016/S0141-0229(02)00245-4
Baldrian P (2008) Chapter 2: enzymes of saprotrophic basidiomycetes. In: Boddy L, Frankland JC, van West P (eds) Ecology of saprotrophic Basidiomycetes, vol 28, British Mycological Society Symposia Series. Elsevier, Amsterdam, pp 19–41
Baptista P, Ferreira S, Soares E, Coelho V, Bastos MD (2009) Tolerance and stress response of Macrolepiota procera nickel. J Agri Food Chem 57:7145–7152. doi:10.1021/jf902075b
Bellion M, Courbot M, Jacob C, Blaudez D, Chalot M (2006) Extracellular and cellular mechanisms sustaining metal tolerance in ectomycorrhizal fungi. FEMS Microbiol Lett 242:173–181
Blaudez D, Botton B, Chalot M (2000) Cadmium uptake and subcellular compartmentation in the ectomycorrhizal fungus Paxillus involutus. Mycrobiology 146:1109–1117. doi:10.1099/00221287-146-5-1109
Bozac R (2005) Mushrooms encyclopaedia 1. Skolska knjiga, Zagreb pp. 598, ISBN 953-0-61413-6
Bozac R (2008) Mushrooms encyclopaedia 2. Skolska knjiga, Zagreb pp. 966, ISBN 978-953-0-61473-4
Brzostowski A, Falandysz J, Jarzynska G, Dan Z (2012) Bioconcentracion potential of metallic elements by Poison Pax (Paxillus involutus) mushroom. J Environ Sci Health Part A 46:378–393. doi:10.1080/10934529.2011.542387
Chojnacka A, Drewnowska M, Jarzynska G, Nnorom IC, Falandysz J (2012) Mercury in Yellow-cracking Boletes Xerocomus subtomentosus mushrooms and soils from spatially diverse sites: assessment of bioconcentration potential by species and human intake. J Environ Sci Health Part A 47:2094–2100. doi:10.1080/10934529.2012.695990
Chudzynski K, Falandysz J (2008) Multivariate analysis of elements content of Larch Bolete (Suillus gravillei). Chemosphere 73:1230–1239
Chudzynski K, Jarzynska G, Falandysz J (2013) Cadmium, lead and some other trace elements in Larch Bolete mushrooms (Suillus grevillei) (Klotzsch) Sing., collected from the same site over two years. Food Adit Contamin 6(4):249–253. doi:10.1080/19393210.2013.807881
Cocchi L, Vescovi L, Petrini LE, Petrini O (2006) Heavy metals in edible mushrooms in Italy. Food Chem 98:277–284. doi:10.1016/jfoodchem.2005.05.068
Collin-Hansen C, Andersen RA, Steinnes E (2005a) Molecular defence systems are expressed in the king bolete (Boletus edulis) growing near metal smelters. Mycologia 97:973–983
Collin-Hansen C, Andersen RA, Steinnes E (2005b) Damage to DNA and lipids in Boletus edulis exposed to heavy metals. Mycolog Res 109:1386–1396
De Mendiburu F (2014) agricolae: statistical procedures for agricultural research. R package version 1.2-1. http://CRAN.R-project.org/package=agricolae
Demers JD, Driscoll CT, Fahey TJ, Yavitt JB (2007) Mercury cycling in litter and soil in different forest types in the Adirondack region, New York, USA. Ecol Appl 17:1341–1351
Drewnowska M, Falandysz J (2015) Investigtion on mineral composition and accumulation by popular edible mushroom common chanterelle (Cantharellus cibarius). Ecotox Env Saf 113:9–17. doi:10.1016/j.ecoenv.2014.11.028
Drewnowska M, Nnorom CI, Falandysz J (2014) Mercury in grisette. Amanita vaginata FR and soil below the fruiting bodies J Environ Sci Health Part B 49:521–526. doi:10.1080/03601234.2014.896677
Dryzalowskaa A, Falandysz J (2014) Bioconcentration of mercury by mushroom Xerocomus chrysenteron from the spatial distinct locations: levels, possible intake and safety. Ecotox Env Saf 107:97–102. doi:10.1016/j.ecoenv.2014.05.020
Falandysz J (2015) Mercury bio-extraction by fungus Coprinus comatus: a bioindicator and mycoremediator of polluted soils. Environ Sci Pollut Res 23(8):7444–741. doi:10.1007/s11356-015-5971-8
Falandysz J, Borovička R (2013) Macro and trace mineral constituents and radionuclides in mushrooms: health benefits and risks. Appl Microbiol Biotechnol 97:477–501. doi:10.1007/s00253-012-4552-8
Falandysz J, Gucia M, Mazur A (2007) Content and biconcentration factors of mercury by Parasol Mushrooms Macrolepiota procera. J Environ Sci Health Part B 42:735–740
Falandysz J, Szymczyk K, Ichihashi H, Bielawski L, Gucia M, Frankowska A, Yamasaki SI (2001) ICP/MS and ICP/AES elemental analysis (38 elements) of edible wild mushrooms growing in Poland. Food Addit Contam 18:503–513
Falandysz J, Mazur A, Kojta AK, Jarzynska G, Drewnowska M, Dryzalowska A, Nnorom IC (2012) Mercury in fruiting bodies of dark honey fungus (Armillaria solidipes) and beneath substratum soils collected from spatially distant areas. J Sci Food Agric 93:853–858. doi:10.1002/jsfa.5807
Falandysz J (2014) Distribution of mercury in Gypsy Cortinarius caperatus mushrooms from several populations: an efficient accumulator species and estimated intake of element. Ecotox Environ Safe 110:68–72. doi:10.1016/j.ecoenv.2014.08.018
Falandysz J, Krasinska G, Pankavec S, Nnorom CI (2014) Mercury in certain boletus mushrooms from Poland and Belarus. J Environ Sci Health Part B 49:690–695. doi:10.1080/03601234.2014.922853
Falandysz J, Zhang J, Wang Y, Krasinska G, Kojta A, Saba M, Shen T, Li T, Liu H (2015a) Evaluation of mercury contamination in mushrooms of genus Leccinum from two different rgions of the world: accumulation, distribution and probalbe dietary intake. Sci Total Environ 537:470–478. doi:10.1016/j.scitotenv.2015.07.159
Falandysz J, Zhang J, Wang YZ, Saba M, Krasinska G, Wiejak A, Li T (2015b) Evaluation of mercury contamination in fungi Boletus species from latosols, lateritic red earths, and red and yellow earths in the Circum-Pacific Mercuriferous Belt of Southwestern China. PlosOne, 1–19. doi: 10.371/journal.pone.0143608.
Falandysz J, Drewnowska M (2015) Macro and trace elements in common chanterelle (Cantharellus cibarius) mushroom from the European background areas in Poland: composition, accumulation, dietary exposure and data review for species. J Environ Sci Health Part B 50:374–387. doi:10.1080/03601234.2015.1000190
Figueiredo E, Soares ME, Baptista P, Castro M, Bastos ML (2007) Validation of an electrothermal atomatization absorption spectrometry method for quantification of total chromium and chromium IV in wild edible mushrooms and underlying soils. J Agric Food Chem 55:7192–7198
García MÁ, Alonso J, Melgar MJ (2009) Lead in edible mushrooms. Levels and bioaccumulation factors J Hazard Mater 167:777–783. doi:10.1016/j.jhazmat.2009.01.058
Garcia MA, Alonso J, Melgar MJ (2013) Bioconcentration of chromium in edible mushrooms: influence of environmental and genetic factors. Food Chem Toxicol 58:249–254. doi:10.1016/j.fct.2013.04.049
Gucia M, Jarzynska G, Kojta A, Falandysz J (2012a) Temporal variability in 20 chemical elements content of Parasol Mushroom (Macrolepiota procera) collected from two sites over a few years. J Environ Sci Health Part B 47:81–88. doi:10.1080/03601234.2012.611433
Gucia M, Jarzynska G, Rafal E, Roszak M, Kojta AK, Osiej I, Falandysz J (2012b) Multivariate analysis of mineral constituents of edible Parasol Mushroom (Macrolepiota procera) and soils beneath fruiting bodies collected from Northern Poland. Environ Sci Pollut Res 19:416–431. doi:10.1007/s11356-011-0574-5
Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biometrical J 50(3):346–363
Işıldak Ö, Turkekul I, Elmastas M, Aboul-Enein HY (2007) Bioaccumulation of heavy metals in some wild-grown edible mushrooms. Anal Lett 40:1099–1116. doi:10.1080/00032710701297042
Işıldak Ö, Turkekul I, Elmastas M, Tüzen M (2004) Analysis of heavy metals in some wild-grown edible mushrooms from the middle Black Sea region, Turkey. Food Chem 86:547–552. doi:10.1016/j.foodchem.2003.09.007
Jarzynska G, Falandysz J (2012) Metallic elements profile of Hazel (Hard) Bolete (Leccinum griseum) mushroom and associated upper soil horizon. Afr J Biotechnol 11:4588–4594
Jarzynska G, Gucia M, Kojta A, Rezulak K, Falandysz J (2011) Profile of trace elements in Parasol Mushroom (Macrolepiota procera) from Tucholskie Forest. J Environ Sci Halth Part B 46:741–751. doi:10.1080/03601234.2011.603986
Jennings DH (2007) The physiology of fungal nutrition. Cambridge University Press, pp 460. ISBN: 9780521038164
Jorhem L, Sundstrom B (1995) Levels of some trace elements in edible fungi. Z Lebensm Unters Forsch 201:311–316
Kalač P (2001) A review of edible mushroom radioactivity. Food Chem 75:29–35. doi:10.1016/S0308-8146(01)00171-6
Kalač P, Svoboda L (2000) A review of trace elements concentrations in edible mushrooms. Food Chem 69:273–281. doi:10.1016/S0308-8146(99)00264-2
Kalač P (2010) Trace element contents in European species of wild growing edible mushrooms: a review for the period 2000–2009. Food Chem 122:2–15. doi:10.1016/j.foodchem.2010.02.045
Kojta AK, Gucia M, Jarzynska G, Lewandowska M, Zakrzewska A, Falandysz J, Zhang D (2011) Phosphorus and certain metals in parasol mushrooms (Macrolepiota procera) and soils from the Augustowska forest and Elk region in north-eastern Poland. Fresenius Environ Bull 20:3044–3052
Krasinska G, Falandysz J (2015) Mercury in Hazel Bolete Leccinum griseum and soil substratum: distribution, bioconcentration and dietary exposure. J Environ Sci Halth Part A 50:1259–1264. doi:10.1080/10934529.2015.1055151
Krasinska G, Falandysz J (2016) Mercury in Orange Birch Bolete Leccinum versipelle and soil substratum: bioconcentration by mushroom and probable dietary intake by consumers. Environ Sci Pollut Res 53:86–869. doi:10.1007/s11356-015-5331-8
Kuldo E, Jarzynska G, Gucia M, Falandysz J (2014) Mineral constituents of edible parasol mushroom Macrolepiota procera (Scop. ex Fr.) Sing and soils beneath its fruiting bodies collected from a rural forest area. Chem Pap 68(4):484–492. doi:10.2478/s11696-013-0477-7
Li T, Wang Y, Zhang J, Zhao Y, Liu H (2011) Trace element content of Boletus tomentipes mushroom collected from Yunnan, China. Food Chem 127:1828–1830. doi:10.1016/j.foodchem.2011.02.012
Malinowska E, Szefer P, Falandysz J (2004) Metals bioaccumulation by bay bolete, Xerocomus badius, from selected sites in Poland. Food Chem 84:405–416. doi:10.1016/S0308-8146(03)00250-4
Manzi P, Aguzzi A, Vivanti V, Paci M, Pizzoferrato L (1999) Mushrooms as a source of functional ingredients. In Euro. Food Chem X European conference on functional foods. A new challenge for the food chemist 1: 86–93, 22–24 September, Budapest, Hungary
Melgar MJ, Alonso J, Garcia MA (2009) Mercury in edible mushrooms and underlying soil: bioconcentration factor and toxicological risks. Sci Total Environ 407:2328–2334. doi:10.1016/j.scitotenv.2009.07.001
Mendil D, Ulüozlü ÖD, Tüzen M, Hasdemir E, Çaglar A (2004) Determination of trace elements on some wild edible mushroom samples from Kastamonu, Turkey. Food Chem 88:281–285. doi:10.1016/j.foodchem.2004.01.039
Nikkarinen M, Mertanen E (2004) Impact of geological origin on trace element composition of edible mushrooms. J Food Compos Anal 17:301–310. doi:10.1016/j.jfca.2004.03.013
Official Gazette 154/08 (2008). Regulations on maximum levels of certain contaminants in foodstuffs. National Journal, Zagreb, Croatia
Ouzouni PK, Petridis D, Koller WD, Kyriakos A, Riganakos KA (2009) Nutritional value and metal content of wild edible mushrooms collected from West Macedonia and Epirus, Greece. Food Chem 115:1575–1580. doi:10.1016/j.foodchem.2009.02.014
Ouzuni PK, Veltsistas PG, Paleologos EK, Riganakos KA (2007) Determination of metal content in wild edible mushroom species from regions of Greece. J Food Compos Anal 20:480–486. doi:10.1016/j.jfca.2007.02.008
Paradis E, Claude J, Strimmer K (2004) APE: analyses of phylogenetic and evolution in R language. Bioinformatics 20:289–290
Pelkonen R, Alfthan G, Järvinen O (2006) Cadmium, lead, arsenic and nickel in wild edible mushrooms. The Finnish Environment 17/2006
Petkovšek SS, Pokorny B (2013) Lead and cadmium in mushrooms from the vicinity of two large emission sources in Slovenia. Sci Total Environ 443:944–954. doi:10.1016/j.scitotenv.2012.11.007
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-project.org/
Saba M, Falandysz J, Nnorom IC (2016) Accumulation and distribution of mercury in fruiting bodies by fungus Suillus luteus forged in Poland, Belarus and Sweden. Environ Sci Pollut Res 23:2749–2757. doi:10.1007/s11356-015-5513-4
Sarkar D (2008) Lattice: multivariate data visualization with R. Springer, New York. ISBN 978-0-387-75968-5
Sarikurkcu C, Copur M, Yildiz D, Akata I (2011) Metal concentration of wild edible mushrooms in Soguksu National Park in Turkey. Food Chem 128:731–734. doi:10.1016/j.foodchem.2011.03.097
Schuster PF, Krabbenhoft DP, Naftz DL, Cecil LD, Olson ML, Dewild JF, Susong DD, Green JR, Abbott ML (2002) Atmospheric mercury deposition during the last 270 years: a glacial ice core record of natural and anthropogenic sources. Environ Sci Technol 36:2303–2310
Smith ML, Bruhn JN, Anderson JB (1992) The fungus Armillaria bulbosa is among the largest and oldest living organisms. Nature 356:428–431. doi:10.1038/356428a0
Soylak M, Saracoglu S, Tüzen M, Mendil D (2005) Determination of trace metals in mushroom samples from Kayseri, Turkey. Food Chem 92:649–652. doi:10.1016/j.foodchem.2004.08.032
Stefanović V, Trifković J, Mutić J, Tešić Ž (2016) Metal accumulation capacity of parasol mushroom (Macrolepiota procera) from Rasina region (Serbia). Environ Sci Pollut Res 1–13. doi:10.1007/s11356-016-6486-7
Svoboda L, Havlickova B, Kalač P (2006) Contents of cadmium, mercury and lead in edible mushrooms growing in a historical silver-mining area. Food Chem 96:580–585. doi:10.1016/j.foodchem.2005.03.012
Svoboda L, Zimmermannova K, Kalač P (2000) Concentrations of mercury, cadmium, lead and copper in fruiting bodies of edible mushrooms in an emission area of copper smelter and a mercury smelter. Sci Total Environ 246:61–67
Širić I, Kos I, Bedeković D, Kaić A, Kasap A (2014) Heavy metals in edible mushrooms Boletus reticulatus Schaeff. collected from Zrin, mountain, Croatia. Period Boil 116:319–322
Thomas GW (1996) Soil pH and soil acidity. Methods of soil analysis. Part 3—chemical methods. Soil Science Society of America and American Society of Agronomy 5:457–490
Thomet U, Vogel E, Krahenbuhl U (1999) The uptake of cadmium and zinc by mycelia and their accumulation in fruiting bodies of edible mushrooms. Eur Food Res Technol 209:317–324
Turkdogan KM, Kilicel F, Kara K, Tuncer I, Uygan I (2003) Heavy metals in soil, vegetables and fruits in the endemic upper gastrointestinal cancer region of Turkey. Environ Toxicol Pharmacol 13:175–179. doi:10.1016/S1382-6689(02)00156-4
Turkekul I, Elmastas M, Tuzen M (2004) Determination of iron, copper, manganese, zinc, leads and cadmium in mushrooms samples from Tokat, Turkey. Food Chem 84:389–392. doi:10.1016/S0308-8146(03)00245-0
UNEP, 2013. Mercury—time to act. United Nations Environmental Programme (http://www.unep.org/PDF/PressReleases/Mercury_TimeToAct.pdf)
van Schöll L, Hoffland E, van Breemen N (2006) Organic anion exudation by ectomycorrhizal fungi and Pinus sylvestris in response to nutrient deficiencies. New Phytol 170:153–163. doi:10.1111/j.1469-8137.2006.01649.x
Vetter J (2004) Arsenic content of some edible mushroom species. Eur Food Res Technol 219:71–74
Wang Z, Chen J, Chai L, Yang Z, Huang S, Zheng Y (2011) Environmental impact and site-specific human health risks of chromium in the vicinity of a ferro-alloy manufactory, China. J Hazard Mater 190:80–985
Yamaç M, Yildiz D, Sarikurcu C, Celikkollu M, Solak MH (2007) Heavy metals in some edible mushrooms from the Central Anatolia, Turkey. Food Chem 103:263–267. doi:10.1016/j.foodchem.2006.07.041
Żarski TP, Żarska H, Arkuszewska E, Válka J, Sokol J, Beseda I (1999) The bioindicative role of mushrooms in the evaluation of environmental contamination with mercury compounds. Ekol Bratislava 18:223–229
Acknowledgments
The authors would like to thank Professor Romano Božac for help, organization, and support during the collection and identification of mushrooms.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Elena Maestri
Rights and permissions
About this article

Cite this article
Širić, I., Humar, M., Kasap, A. et al. Heavy metal bioaccumulation by wild edible saprophytic and ectomycorrhizal mushrooms. Environ Sci Pollut Res 23, 18239–18252 (2016). https://doi.org/10.1007/s11356-016-7027-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-016-7027-0