
Abstract
Over the last decades, since the discovery of ATP as a transmitter, accumulating evidence has been reported about the role of this nucleotide and purinergic receptors, in particular P2X7 receptors, in the modulation of synaptic strength and plasticity. Purinergic signaling has emerged as a crucial player in orchestrating the molecular interaction between the components of the tripartite synapse, and much progress has been made in how this neuron-glia interaction impacts neuronal physiology under basal and pathological conditions. On the other hand, pannexin1 hemichannels, which are functionally linked to P2X7 receptors, have appeared more recently as important modulators of excitatory synaptic function and plasticity under diverse contexts. In this review, we will discuss the contribution of ATP, P2X7 receptors, and pannexin hemichannels to the modulation of presynaptic strength and its impact on motor function, sensory processing, synaptic plasticity, and neuroglial communication, with special focus on the P2X7 receptor/pannexin hemichannel interplay. We also address major hypotheses about the role of this interaction in physiological and pathological circumstances.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Chemical synapses are specialized junctions through which cells of the nervous system (NS) communicate with each other. At chemical synapses, the transfer of information between two neurons requires the release of neurotransmitter molecules, which act on the postsynaptic neuron by binding to the specific receptor localized at the plasma membrane, in a process known as neurotransmission.
Adenosine triphosphate (ATP) is abundantly present in the cytosol and is classically known to fuel power-consuming reactions. Burnstock and colleagues first highlighted the role of ATP as a physiological modulator of synaptic transmission in the autonomic system, and subsequently, it was identified as a co-transmitter in parasympathetic and sympathetic nerves [1, 2]. At central nervous system (CNS) synapses, it is well-accepted that ATP acts as a transmitter or co-transmitter by being released with other neurotransmitters, such as glutamate or GABA [3]. Interestingly, both spontaneous and evoked ATP release have been described in pyramidal neurons of the mouse cerebral cortex and fast synaptic currents and long-lasting enhancement of the population spikes are induced by ATP release in the hippocampus [4]. The extracellular concentration of ATP in the NS is determined by the balance between its release and enzymatic degradation, and different studies indicate that this nucleotide is released by both neurons and glia [5, 6]. For instance, physiological and high-frequency stimulations modify that balance by increasing ATP release and thus its extracellular availability [7, 8]. In addition to exocytosis and considering that ATP molecules are not able to diffuse across the plasma membrane, the passage of ATP from the intracellular to the extracellular compartment is also mediated by mechanisms involving ATP-permeable channels as connexin or pannexin hemichannels (CxHCs or PanxHCs, respectively) [9, 10].
There are two fundamental classes of receptors for extracellular ATP that work on different time scales. The metabotropic P2Y receptors (P2Y1,2,4,6,11) are G-protein-coupled receptors involved in responses that can last several seconds through changes in gene expression [11,12,13]. The second class, the ionotropic P2X receptors (P2X1-7), comprises ligand-gated ion channels that mediate fast responses lasting milliseconds. These receptors can assemble as homo- or heterodimers; thus, the different combinations of subunits result in receptors with a wide variety of structural and biophysical properties [11,12,13].
In this review, we discuss the role of ATP, P2X7 receptor (P2X7Rs) and PanxHC interaction in the modulation of presynaptic strength with a special focus on the participation of the neuron-glia crosstalk in this process. For a detailed view of the structure, expression and pharmacology of P2X7Rs, the following reviews are recommended [14, 1516].
The basic functioning of P2X7Rs
The P2X7Rs are ligand-gated ion channels permeable to Na+, K+, and, more significantly, Ca2+, as all P2X receptors. However, they display distinctive properties compared to the other members of the family. Pioneer studies showed that they are the only ones that do not heterodimerize with other P2X subunits, being usually assembled with three to six P2X7 subunits [5, 17]. However, more recent evidence suggests a close association between P2X7 and P2X4 subunits [18, 19]. On the other hand, they have low affinity to ATP, requiring high levels of extracellular ATP (> 100 μm) for activation [17]. Interestingly, P2X7Rs show plasticity, which leads to selective or non-selective currents. In this sense, milliseconds of ATP-dependent activation results in the opening of small cation-selective channels, while a sustained activation—with a high concentration of ATP—increases the permeability to non-selective conductance, including organic cations and small peptides, in a process known as pore formation, which represents an intrinsic property of the channel itself [5, 17, 20,21,22]. Moreover, repeated stimulation with an agonist results in relevant changes in the amplitude and time course of the current evoked by the following applications [17, 23]. Notably, the C-terminal domain of the P2X7 subunit is suggested to be required for the pore-forming process [24]. However, the molecular mechanism underlying this process remains still unclear and controversial and two main hypotheses, the “pore-dilation” and the “pore-formation,” have been raised (reviewed in [21, 25,26,27]). The first supports that upon sustained or repetitive stimulation, the ion channel intrinsically dilates producing a pore that allows the passage of large molecules (up to ~ 900 Da) [24, 28]. A proposed alternative to this hypothesis is that the channel dilation may involve the successive incorporation of new P2X7 subunits [28, 29]. Notably, a study performed by Karasawa and colleagues suggested that the pore dilation and thus channel permeability could be modulated by the lipid microenvironment in the vicinity of P2X7Rs [27, 30]. Together, these data indicate that the “pore dilation” might be controlled by different cellular pathways. On the other hand, the “pore formation” is supported by different electrophysiology studies evidencing that the increased membrane permeability could be mediated by the interaction of P2X7R with the ATP-permeable channel, Panx1, followed by cytoskeleton rearrangements [24, 31]. We will discuss the interaction of P2X7Rs and Panx1HCs further above in the text. Electrophysiology and dye uptake studies performed in heterologous expression systems reveal that the property of increasing membrane permeability to large molecules is also observed in P2X2 and P2X4 receptors [5, 17, 20,21,22]. Interestingly, Kim and colleagues showed that P2X7Rs are dephosphorylated upon activation, leading to a negative feedback control of the channel currents [32]. As other members of the family, P2X7Rs have cytoplasmic N- and C-terminal domains and two transmembrane regions separated by an extracellular domain. The C-terminal domain is the longest of the P2X family and mediates several signaling cascades by interacting with the actin cytoskeleton and different intracellular proteins, such as the heat shock proteins (Hsp90, Hsc71, Hsp70), phosphatidylinositol 4-kinase (PI4K), and membrane-associated guanylate kinase (P55) [32, 33]. Interestingly, dephosphorylation of P2X7Rs also reduces the ability to mediate cytoskeletal rearrangements [32]. Thus, activation of P2X7Rs not only opens a cation-permeable channel, but it can participate in different signaling pathways by interacting with downstream components and promoting cytoskeleton rearrangements. In this sense, Armstrong and colleagues showed that bath application of the P2X7R agonist BzATP prevents neurotransmission at the mossy fiber–CA3 synapses. Interestingly, the synaptic depression is blocked by applying 4-(4-fluorophenyl)2-(4-methylsulfinylphenyl) 5-(4-pyridyl) imidazole, a strong antagonist of p38/MAP kinase activation. Altogether, these suggest that P2X7Rs modulate mossy fiber synaptic depression through the activation of p38/MAPK in rat hippocampus [34].
Basic functioning of Panx1HCs
The Panx1 protein forms large pore membrane channels permeable to ions and relative large molecules with physiological relevance supporting an autocrine/paracrine signaling. Panx1 is part of a family of membrane glycoproteins comprising three members (pannexins 1, 2, and 3), being Panx1 the most ubiquitously expressed in the nervous system of mammals [35,36,37]. The protein is mostly detected in neurons but is also found in oligodendrocytes, astrocytes, and microglia [36, 38,39,40,41]. Interestingly, neuronal Panx1 could co-localize with the postsynaptic density protein, PSD95, suggesting a role in synaptic transmission [42]. Structurally, pannexins consist of four α-helical transmembrane domains with cytoplasmic amino- and carboxy-termini; two extracellular and one intracellular loops link transmembrane domains [43,44,45]. Recent studies using single-particle cryo-electron microscopy (cryo-EM) at near-atomic resolution in different heterologous expression systems determined that Panx1 oligomerizes into heptamers in contrast to the hexameric conformation previously reported, forming plasma membrane channels known as pannexons or PanxHCs [46,47,48,49,50].
Despite pannexons sharing some topological and permeability properties with CxHCs or connexons, Cxs and Panxs do not show sequence homologies [44]. PanxHCs, like connexons, exhibit a large central pore allowing interchange of ions and large molecules (up to 1.2 kDa) between the cytosol and the exterior [10]. Classically, Panx1HCs have been known as non-selective high conductance (~ 500 pS) and permeable channels permitting the passage of transmitters such as ATP and glutamate under diverse physiological and pathological conditions [43, 51,52,53,54]. However, current literature associates Panx1HCs with divergent unitary properties [55]. In addition to the high conductance/permeability conformation, recent studies reported an anion-permeable low conductance (~ 68–74 pS) conformation with no ATP permeability; this channel state shows selectivity for chloride ions and is activated uniquely by voltage [56, 57]. Likewise, dual voltage clamp recordings resolved in HeLa cells expressing human Panx1-YFP and rodent spinal astrocytes described low mean unitary conductance of ~ 42 and ~ 48 pS [58]. Discrepancies between unitary conductance and ATP permeation properties may be ascribed to different modes of Panx1 activation; actually, smaller-conductance Panx1HCs (up to ~ 100 pS) may co-exist with permeation of large molecules [55, 59]. In this line, caspase cleavage of the auto-inhibitory cytoplasmic carboxy-terminus (C-tail) of Panx1 associates with broadening of the central pore; progressive removal of C-tails has been associated with stepwise graded channel activation of multiple discrete open states, each contributed by individual subunits resulting in permeation of ions and large molecules [59]. Combining cryo-EM analysis, ATP release measurements, and electrophysiology, novel studies confirmed the passage of ATP through a heptameric Panx1 channel presenting small single-channel conductance [45,46,47,48]. In accordance, a gating mechanism involving two ionic-conducting pathways, a big main pore permeable to large molecules, and seven narrow tunnels located intracellularly and permeable to small anions has been reported. In normal conditions, the main pore is physically blocked by the C-terminal tail and thus anions—but not ATP—can solely pass through the side tunnels. Under pathological conditions, the C-terminal tail is cleaved by caspase, and ATP is released through the main pore [48]. According to cryo-EM reconstructions, the Panx1 permeation pathway is exceptionally wide in the transmembrane and cytoplasmic portions thus supporting permeation of large molecule ATP or ethidium. Constriction sites would take place in the extracellular domains where the conserved tryptophan ring (W74) in the first extracellular loop may act as a putative selective filter [45,46,47,48]. Hence, Panx1 seems to have multiple open pore configurations. Finally, the Panx1HC conductance/permeability and kinetics are still a matter of investigation; stimulatory conditions may determine channel properties and conformational states [55, 60].
Panx1HCs as dynamic routes for ATP release
Pannexons are typically known as paths for ATP release [10, 51, 61]. Released ATP triggers purinergic signaling either by itself or through its metabolites (adenosine diphosphate or ADP, monophosphate or AMP, and adenosine). This unconventional Panx1-dependent efflux of ATP constitutes a paradigmatic mechanism for non-vesicular transmitter release [43]. In the NS, the release of glio- and neurotransmitters (glutamate, D-serine, and ATP) by neural cells via Panx1HCs has been implicated in synaptic plasticity in health and disease being the subject of an increasing number of publications [41, 62, 63]. Importantly, pannexons open under physiological extracellular Ca2+ concentrations and negative resting membrane potentials and show poor voltage dependence [36, 52, 64]. Pannexons can be activated by diverse mechanisms (reviewed in [55, 65,66,67]) including purinergic (P2X4/7, P2Y1/2/6), alpha1-adrenergic, and glutamatergic (N-Methyl-D-aspartate) receptors [24, 68,69,70]. When activated by purinergic receptors, Panx1HCs behave as physiological regulators of ATP release; the outcome of this association is the so-called “ATP liberation induced by ATP.” Accordingly, ATP released through pannexons opens new pannexons by stimulating purinergic receptors resulting in a positive feedback that reinforces the initial ATP signal [66, 71]. If maintained, this situation is potentially dangerous since channels might be kept in a permanent open state and ensuing exacerbated pannexon activity would be deleterious [72]. Recent reports suggested that physiological activation of Panx1HCs by ATP might be transient in the presence of purinergic P2Y/P2X7 receptors due to the existence of an inhibitory mechanism or internalization of Panx1 [72,73,74,75,76]. While the most studied roles of Panx1HCs rely on their capability to allow the release of ATP and the effects of purinergic signaling, other alternative roles might be ascribed to the non-channel properties of Panx1 [77].
Pannexin1 as subunits of gap junction channels: the debate is open
Pannexins are considered the mammalian orthologs of innexins, the gap junctions of invertebrates, but in contrast to them (and to connexons), whether PanxHCs from opposing cells come together to generate gap junction channels remains controversial. Initial electrophysiological studies carried out by Bruzzone and colleagues in paired oocytes of Xenopus indicated that exogenously expressed Panx1 assembles as intercellular channels forming either homomeric or heteromeric channels with Panx2, having both channel differential properties [36]. Dye coupling assays in C6 glioma cells loaded with sulforhodamine SR101 revealed enhanced dye coupling in Panx1-expressing cells supporting these primary observations [78]. Likewise, overexpression of Panx1 in human cell lines may induce the formation of gap junction channels permeable to Ca2+ underlying the propagation of intercellular Ca2+ waves [79]. Subsequent investigations questioned the capability of Panx1 to form cell–cell channels in appositional membranes under physiological and in vivo conditions. A main argument is the presence of N-glycosylated asparagine residues on Panx1 second extracellular loop that would prevent docking [73, 80,81,82]. In agreement with this, electrophysiological and functional studies performed in cell lines carrying different glycosylation patterns of expressed Panx1 showed that lower levels of Panx1 glycosylation associate with higher probabilities to find functional Panx1-formed gap junction channels [83]. Also, combining single-particle cryo-electron microscopy and patch-clamp electrophysiology in a glycosylation-deficient mutant of Panx1 allowed identifying a gap-junction-like structure [48]. Yet, experimental conditions promoting Panx1 glycosylation have been recently discussed [84]. Additional arguments against Panx1-forming gap junctions are that recordings of Panx1-based junction currents require special experimental conditions and may be contaminated with junction currents from connexin-based gap junction channels [81]. A more recent study elegantly performed by Sáez Laboratory using functional/electrophysiological/pharmacological/genetic and in silico assays characterized Panx1-based cell–cell channels in transfected and endogenous human lines [84]. Thus, electrophysiological recordings of exogenous human pannexin1 (hPanx1) cell–cell channels expressed in HeLa cells knocked out for Cx45 showed two states of hPanx1 intercellular channels: O-state with single-channel conductance of ∼ 175 pS, a substate of ∼ 35 pS and low transjunctional voltage (Vj) sensitivity, and S-state, with single-channel conductance of ∼ 35 pS, asymmetry in Vj dependence and permeability to DAPI. Remarkably, S-state hPanx1 intercellular channels were also recognized between TC620 cells, a human oligodendroglioma cell line that endogenously expresses hPanx1 [84]. Importantly, in silico approaches suggested that some arginine residues at the channel pore may be neutralized by hydrophobic interactions permitting DAPI permeability. These novel findings reopen the debate about the possibility that Panx1HCs assemble as functional intercellular channels and, essentially, raise the question about the eventual role of Panx1-gap junction channels as morpho-functional substrates for electrical synapses under defined physiological circumstances [84].
Panx1HC association with P2X7Rs
Panx1HCs functionally associate with ionotropic purinergic P2X7Rs. In addition to operating as a cationic channel, prolonged stimulation (up to 20 min) of the ligand-gated ion P2X7R leads to the permeation of large molecules (up to 1 kDa) such as ATP [5, 85]. As we commented above, the mechanisms underlying the permeation of P2X7Rs to large molecules are subject of intense debate. In one possible scenario, pioneering studies done in macrophages suggested that Panx1HCs could function as the large pore of the P2X7R under prolonged activation [24]. In these cells, ATP gates P2X7Rs and opens a large transmembrane route allowing ethidium uptake and the release of the pro-inflammatory cytokine interleukin-1beta (IL-1β), being these effects prevented by Panx1 block. By using small interference RNA (siRNA) directed to Panx1 and a Panx1-mimetic inhibitory peptide, the authors inhibited the dye uptake elicited by P2X7Rs without affecting the ionic current and Ca2+ entry associated with P2X7R stimulation [24]. Some studies then reported that large plasma membrane pore opening upon P2X7R activation could occur in the absence of Panx1 [86, 87]. The biphasic permeation properties of P2X7Rs were reported in systems not expressing Panx1 and under high agonist concentrations; intracellular residues of the channel pore seemed to be required for the pore dilatation under those circumstances [86].
What mechanisms link P2X7Rs and Panx1HCs? Increases in cytosolic Ca2+ activate pannexons and ATP release; thus, opening of Panx1HCs would be triggered by the increase in intracellular Ca2+ downstream P2X7R stimulation [9, 73, 88, 89]. Conversely, it has been reported that Panx1HC opening might be independent from extracellular Ca2+ and direct protein–protein interaction could underlie Panx1HC activation as suggested by the fact that Panx1 co-immunoprecipitates with the P2X7R [24, 72]. A Src tyrosine kinase-dependent process might couple Panx1HC and P2X7R activation, while the carboxy-terminal region of P2X7Rs would be required to open Panx1HCs [53, 90]. To add more intricacy to the P2X7R/Panx1 interaction, a recent study performed in heterologous expression systems revealed that Panx1 attenuates the P2X7R-dependent Ca2+ entry. This P2X7R inhibition appears to be mediated by the C-terminal domain of Panx1 [91]. Interestingly, whether Panx1 attenuates or potentiates P2X7R-dependent signaling pathways seems to depend on the degree of P2X7R activation [92]. Hence, solid evidence supports the idea that Panx1 and P2X7R exert a modulatory influence on each other; however, the accurate mechanisms underlying the physical interaction between them remain to be clarified.
In this review, we will focus on pannexons expressed in neural cells and their acting in conjunction with P2X7Rs to contribute to the shaping of presynaptic neurotransmission.
P2X7Rs and Panx1HCs as modulators of presynaptic strength
Different studies described the presence of P2X7Rs in presynaptic terminals and functional data support a pivotal role of these receptors in presynaptic neuronal function [34, 93,94,95,96,97,98,99]. Presynaptic P2X7Rs promote a wide range of effects on synapse functionality, including facilitation and/or inhibition. For instance, activation of P2X7Rs with the receptor agonist BzATP promotes neurotransmitter release in glutamatergic synapses in the cortex, hippocampus, cerebellum, spinal cord, and neuromuscular junction [93, 94, 100,101,102]. In isolated presynaptic terminals, pulses of ATP or BzATP induce an increase in presynaptic Ca2+ availability, which is sensitive to P2X7R antagonists, suggesting that functional P2X7Rs are present in presynaptic terminals [100, 101, 103]. Interestingly, the P2X7R permeability to Ca2+ is lower at depolarized membrane potentials compared to negative potentials. In this sense, it has been suggested that neurotransmitter release could be initiated through P2X7R activation, without presynaptic depolarization and voltage-gated Ca2+ channel opening, thus, also contributing to neurotransmitter release at resting membrane potentials [94, 104]. Furthermore, the P2X7R- mediated Ca2+ currents are not desensitized and potentiated when the extracellular concentration of Ca2+ or Mg2+ is decreased. The high conductivity to Ca2+ ions under physiological conditions provides P2X7Rs the alternative to act as presynaptic sources of Ca2+ [100, 105, 106]. Either presynaptic P2X7R-mediated Ca2+ signaling or P2X7R-activated intraterminal Ca2+-dependent signaling messengers modulate neurotransmitter release in central and peripheral synapses [100, 102, 107, 108]. During the last years, Panx1 has emerged as an important modulator of synaptic function. In the adult brain, the absence of Panx1 leads to changes in the dendritic arbor and excitability of hippocampal neurons, preserving spontaneous activity [77]. Moreover, Panx1HCs, which are reported to open under resting membrane potential, are also essential regulators of synaptic plasticity in hippocampal synapses. For instance, the absence or blockade of Panx1HCs in adult mice favors LTP by modifying the threshold of synaptic plasticity induction and increases excitatory synaptic transmission [109, 110]. Accordingly, a recent report showed that blocking Panx1HCs in hippocampal slices increases the synaptic level of endocannabinoids and the activation of CB1 receptors; as a consequence, GABAergic efficacy decreases. This effect shifts the excitatory/inhibitory balance toward excitation and facilitates the induction of LTP in CA3-CA1 synapses [111]. The functional significance of Panx1HCs in hippocampal synapses is suggested to be mediated by both pre- and postsynaptic mechanisms [109]. More recently, evidence suggests that neurotransmitter release probability (pr) and the size of the readily releasable pool (RRP) are affected in Panx1-knockout (Panx1KO) mice and thus glutamate release [77]. In this line, we have recently reported that, in hippocampal dissociated cultures, neuronal Panx1HCs and P2X7Rs are required for the compensatory adjustment of presynaptic strength upon chronic inactivity [99]. In the ensuing paragraphs, we review recent advances regarding the role of ATP, P2X7Rs, and Panx1HCs in the modulation of presynaptic strength at central and peripheral synapses and address major research gaps that require more study.
ATP, P2X7Rs, and Panx1HCs regulate the presynaptic release of acetylcholine (ACh) at the neuromuscular synapse
During neuromuscular transmission, in addition to released ACh, ATP originating from both nerve terminals and muscle fibers accumulates at the synaptic cleft [112, 113]. Endogenous ATP and its hydrolysis products regulate the presynaptic release of ACh [107, 108, 113,114,115]. Recent reports propose Panx1HCs as sources of synaptic ATP involved in purinergic regulation of neuromuscular junctions (NMJs). According to these studies, stimulation of P2X7Rs by the specific agonist BzATP in isolated hemidiaphragm neuromuscular preparations of Panx1KO mice enhances evoked ACh release through presynaptic activation of calmodulin, Ca2+/calmodulin-dependent kinase II, and L-type of voltage-dependent Ca2+ channels. Effects induced by P2X7R stimulation, i.e., increased quantal content of evoked endplate potentials (EPPs) and increased size of the readily releasable ACh pool, were not observed in wild-type mice yielding to the suggestion that Panx1 normally inhibits the tonic enhancement of P2X7-evoked release of ACh [107]. This is counterintuitive since activated Panx1HCs boost neurosecretion in other systems through a process involving P2X7R and amplification of cytosolic Ca2+ signals [116, 117]. However, other purinergic receptors would come into play; at the frog neuromuscular junction, ATP co-released with ACh inhibits quantal ACh release from motor nerve terminals via metabotropic P2Y receptors [118]. At mouse NMJs, the inhibitory actions of presynaptic purinergic P2Y13 and adenosine A1 receptors activated by Panx1HC-released ATP and adenosine would override the stimulatory effects of P2X7R, resulting in net inhibition of ACh release by Panx1HCs [107, 113]. Noteworthily, the global Panx1KO mice used in mammalian NMJs studies do not discriminate the cellular origin of the Panx1HCs involved in the control of presynaptic ACh release. Interestingly, at the postsynapse, Panx1 activation enhances muscle contraction; in adult skeletal muscles, Panx1HC opening mediates ATP efflux and influx of Ca2+ and glucose, necessary for potentiation of contraction [119, 120]. Hence, the final effects of the P2X7Rs/Panx1HC interplay on muscle function (stimulatory or inhibitory) will depend on the cellular localization of Panx1HCs as well as on the actions of purinergic receptors other than P2X7Rs, also activated during the Panx1HC-mediated ATP discharge. What would be the physiological role of the tonic inhibition of ACh release by Panx1HCs in mouse NMJs? As proposed by others, Panx1HCs might be part of a feedback control mechanism through which transmitters could adjust the gain of synaptic strength [63, 121]. We speculate that, at mouse motor synapses, inhibition of ACh release by the Panx1HCs during high neuronal activity might avoid synaptic fatigue. According to this idea, the resultant effects on presynaptic ACh release, due to the stimulatory (P2X7R) and inhibitory (P2YR) actions of Panx1HC-released ATP, would contribute setting the moment-to-moment dynamic range of ACh synaptic levels, within the NMJ which would efficiently perform in physiological conditions. However, further studies will be required to resolve this issue.
The neuroglial crosstalk in the modulation of synaptic strength
Different sources of evidence showed that the fine-tuned modulation of neurotransmission, that has been previously thought to only require the signaling between two neuronal cells, involves an intimate communication between glial cells and neurons. Thus, currently it is well-recognized that glial cells are not only passive support cells for neurons but also active players in neuronal network development and information processing. At synapses, neuronal and non-neuronal cells are closely associated by the fine processes of both astrocytes and microglia that modulate synapse formation and function [122,123,124]. Added to their proximity to synapses, astrocytes secrete a wide variety of chemical signals that control synaptic transmission. This led to the concept of “tripartite synapse,” a synapse composed of one presynaptic neuron, one postsynaptic neuron, and an astrocyte, shaping together a functional unit [124,125,126,127,128]. A tripartite synapse displays an intimate communication between the different cellular players through a bidirectional signaling pathway; neurons release neurotransmitters that bind to postsynaptic receptors and receptors on the adjacent astrocyte process, activating signaling pathways in the astrocytes which finally fine-tune synaptic function [127, 128]. Thus, brain function depends on the communication between neurons and glial cells that can activate a wide variety of molecular interactions leading to the modulation of synaptic strength. In this scenario, we next discuss recent findings that shed light on the role of P2X7Rs and their interaction with pannexons in the modulation of presynaptic strength in both central and peripheral nervous systems.
The P2X7R/Panx1 complex mediates astroglial D-serine release: possible impact on presynaptic strength
Astrocytes synthesize and release D-serine, an endogenous neurotransmitter that acts as a co-agonist with glutamate at N-methyl-D-aspartate receptors (NMDARs) [129, 130]. In the brain, astrocyte D-serine controls NMDAR-mediated synaptic activity, long-term potentiation (LTP), long-term depression (LTD), and cognition [131,132,133,134]. Studies reveal that the rate of supply of D-serine to neurons controls the firing rate of these cells. Neurological (i.e., Alzheimer, epilepsy, stroke) and psychiatric (i.e., schizophrenia) disorders typically exhibit alterations of extracellular D-serine. Then, the adequate function of NMDAR requires the fine adjustment of extracellular levels of this molecule [135]. Several molecular and cellular mechanisms mediate the D-serine efflux [136]. In particular, astrocytes secrete this neurotransmitter through a Ca2+-independent activation of P2X7Rs that involves Panx1; additional releasing pathways include Ca2+/SNARE-dependent exocytosis and Cx43HCs [137,138,139]. In cultured cortical astrocytes, stimulation of glial P2X7Rs by ATP elicits D-serine discharge via Panx1HCs through a Ca2+-independent PKC isozyme [137]. Formation of the P2X7R-Panx1 complex is essential for this effect, as pharmacological inhibition of the Panx1HCs and downregulation of Panx1 expression by shRNAs prevent the release of astrocyte D-serine triggered by the P2X7R agonist BzATP [137]. Whereas Ca2+-dependent exocytosis of astrocyte D-serine controls synaptic plasticity in the hippocampus and Cx43HC-mediated astrocyte-released D-serine potentiates NMDAR-synaptic currents at the prefrontal cortex, the contribution of glial D-serine and P2X7R/Panx1HCs in neurotransmission and particularly in the presynapse is unknown [134, 138]. An eventual effect of glial D-serine on presynaptic NMDARs could be a possibility. In other brain regions, electrophysiological studies have shown that D-serine promotes glutamate release from nerve terminals acting through presynaptic NMDARs [140, 141]. Besides, cortical astrocytes may provide D-serine to activate presynaptic NMDARs facilitating glutamate release [142]. Currently, future studies are required to unravel the role for astroglial D-serine and P2X7R/Panx1HC in neurotransmission, as well as to discriminate whether the different molecular mechanisms involving its release (Cx43HCs, Panx1HCs, P2X7Rs, or exocytosis) operate under specific circumstances (i.e., health or disease) and trigger precise modulatory mechanisms (i.e., pre or postsynaptically), alone or concurrently.
Paracrine signaling by Panx1HCs and P2X7Rs sustains bidirectional interactions between neurons and satellite glial cells (SGCs) in peripheral sensory ganglia: implications for nociceptive afferent transmission
Ganglia primary neurons are the first relay in the sensory pathways transmitting afferent information from the periphery (skin, muscles, organs, etc.) to the CNS (spinal cord and brainstem). The soma of these sensory neurons localizes in the peripheral dorsal root ganglion (DRG) or ganglia of sensory cranial nerves. The sensory neurons are pseudounipolar and their cell body gives origin to a short axon, which divides into two branches yielding a t-shaped bifurcation. One branch innervates the periphery while the other enters the CNS where it synapses with central neurons (second relay) localized in the posterior horn (for spinal nerve) or the medulla oblongata (for cranial nerves) [143]. Although action potential conduction from the periphery to axon terminals innervating the postsynaptic second relay might bypass the perikaryon without distortion of the sensory information, the ganglia neuronal soma provides the unique possibility for regulating the afferent sensory transmission to the CNS [144]. Moreover, notably, there is no current morphological or functional evidence indicating that neurons within the peripheral ganglia chemically synapse between each other, and sensory neurons principally rely on satellite glial cells (SGCs), the chief ganglia glial cells, to modulate their excitability through gliotransmission. Actually, cell bodies of sensory ganglia neurons are typically enwrapped by surrounding SGCs forming a “neuroglial unit” [145, 146]. Recent reports indicate that interaction mediated by purinergic signaling and Panx1HCs sustains paracrine communication between sensory neurons and SGCs contributing to regulate nociception [147,148,149,150,151,152,153]. In murine visceral nodose-petrosal-jugular (NPJ) complexes, the SGCs Cx43HCs act together with P2X7Rs and Panx1HCs to mediate a paracrine neuroglial communication [149]. In this case, the pharmacological opening of glial CxHCs increases the frequency discharge in vagal axons projecting from the NPJ sensory neurons to the medulla, being this effect inhibited in Panx1KO mice. Single application of BzATP, a P2X7R agonist, to the NPJ complexes, also increases the electrical activity in sensory neurons. The oxidized ATP (oATP), an irreversible P2X7R antagonist, and probenecid, a Panx1 blocker, reduce the increased sensory discharge elicited by pharmacological CxHC opening. Panx1 is found in both sensory neurons and SGCs [149, 154,155,156] whereas P2X7Rs are selectively expressed by satellite glial cells [147, 148, 153, 157, 158]. Since Panx1HC activation associates with P2X7R function, authors proposed that interactions between neuronal Panx1HCs, glial Cx43HCs, and P2X7Rs orchestrate the increase of NPJ sensory neuron activity [149]. In the dorsal root ganglia (DRG), stimulation of P2X7Rs by BzATP triggers cytosolic Ca2+ increases in SGCs and neurons in vivo being these responses blocked by antagonists of P2X7Rs, P2X3Rs, and Panx1 [153]. Then, in DRG, activated P2X7Rs could induce the paracrine release of glial ATP through opening Panx1HCs, which increases neuron excitability in vivo through P2X3Rs that are mainly expressed by sensory neurons [5, 157]. In turn, released ATP may result in the opening of neuronal Panx1HCs that would contribute to further ATP release [153]. Interestingly, the study by Chen and colleagues also evaluated the impact of the SGC–neuron interaction in the modulation of afferent transmission [153]. In this regard, ganglionic application of BzATP increases the number of activated DRG neurons that respond to brush stimulation of the ipsilateral hind paw. In addition, the response of C-fibers to the electrical stimulation of the sciatic nerve increases after BzATP application to the L4 DRG and the ganglionic application of carbenoxolone (CBX), a Panx1 and Cx general blocker, decreases the number of DRG neurons activated by peripheral high-intensity electrical stimulation. Hence, this work provides clear evidence that DRG neuron activation triggered by P2X7R stimulation and Panx1HCs sensitizes sensory neurons to subsequent peripheral stimulation enhancing the nociceptive transmission from the periphery to central pathways [153]. In chronic pathological pain, the initial injury induces hyperexcitability of primary afferent neurons from the ganglia (peripheral sensitization), leading to central hypersensitivity that persists after the original injury. In pathological pain, Panx1 is up-regulated as reported in murine trigeminal ganglia in a chronic orofacial pain model and in DRG after nerve injury [151, 156]. The Panx1 contribution to orofacial pain depends on the cell-type expressing the protein; targeted deletion of Panx1 in satellite glia completely blunts hypersensitivity, while silencing Panx1 expression in sensory neurons reduces baseline sensitivity and hypersensitivity duration. The role of Panx1HCs in pathological orofacial pain might involve the increased release of the algogenic mediator ATP and cytokines [151]. Since as discussed, the interaction between P2X7Rs and Panx1HCs amplifies the nociceptive entry, we might speculate that this could represent a defense mechanism to enhance alert about the presence of a noxa. Finally, the contribution of Panx1HCs in nociceptive processing and pathological chronic pain points at this protein as a promising target for new therapeutic strategies [152].
Homeostatic role of the purinergic ATP/P2X7R pathway and Panx1HCs: modulation of presynaptic strength under chronic inactivity
Homeostatic synaptic plasticity (HSP) is a form of plasticity displayed with the aim to stabilize the activity of neural circuits in response to chronic perturbations that can lead to irreversible damage. Compensatory mechanisms involve changes in pre- and postsynaptic strength and/or intrinsic neuronal excitability [159,160,161]. Consistent with a role of gliotransmission in modulating basal synaptic strength, molecular mechanisms underlying neuron-glia bidirectional communication in PSH have been also described. Indeed, studies in hippocampal dissociated and entorhino-hippocampal slice cultures showed that tumor necrosis factor alpha (TNFα) is required for a postsynaptic form of HSP known as synaptic scaling [162,163,164,165]. Moreover, the role of TNFα in postsynaptic homeostatic plasticity has been described for different neural functions including developmental plasticity, diverse addiction models, and response to psychiatric drugs [166].
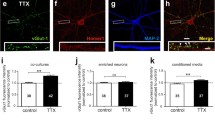
A recent study using hippocampal dissociated cultures has identified a pivotal role for gliotransmission in the compensatory adjustment of presynaptic strength upon chronic TTX exposure [99]. Evidence showed that the compensatory increase in the abundance of synaptic vGlut1 requires glial-derived ATP, astrocytic Cx43HCs, neuronal Panx1HCs, and presynaptic P2X7Rs, as suggested by blocking experiments using specific antagonists. How may this homeostatic modulation occur? The precise relationship between these molecular players is the aim of future work, but one possible scenario is that, upon chronic activity blockade, neurons reduce the release of glutamate, which is sensed by astrocytes, leading to a sustained opening of Cx43HCs and thus enhancing ATP release. Acting as a gliotransmitter, ATP activates presynaptic P2X7Rs, which are Ca2+-permeable channels in a voltage-independent manner [101, 167, 168]. Interestingly, by using a Ca2+ reporter targeted to presynaptic terminals of hippocampal dissociated neurons, Zhao and colleagues showed that adaptation to chronic inactivity involves an increase in presynaptic Ca2+ in response to an action potential [169]. In this sense, another study performed in cortical dissociated neurons showed an extensive molecular remodeling of the presynapse upon chronic inactivity, with significant changes in the amount of different presynaptic proteins, such as Cav2.1 and P/Q-type Ca2+ channels [170], suggesting that the increase in presynaptic Ca2+ upon inactivity could be due to the rise in the abundance of these channels. Interestingly, a more recent study found that low voltage–threshold T-type Ca2+ channels participate in the homeostatic modulation of excitability and integrative properties in hippocampal dissociated and organotypic slice cultures upon chronic TTX application [171]. On the other hand, the data shown by Rafael and colleagues opens up the possibility that the increase in presynaptic Ca2+ upon chronic inactivity could be, at least in part, mediated by P2X7Rs. This seems to be due more to an enhanced activation and prolonged P2X7 channel opening, than to an increase in the presynaptic amount of these receptors upon chronic inactivity [99]. This is suggested by immunocytochemistry studies where the distribution and abundance of presynaptic P2X7Rs were not affected upon chronic TTX treatment, suggesting that the availability of these channels is not modified due to HSP [99]. Interestingly, neuronal Panx1HCs are suggested to cooperate with presynaptic P2X7Rs in the compensatory adjustment of presynaptic strength [99]. Again, the abundance of Panx1HCs seems not to be affected upon chronic inactivity. In agreement with this and taking into account the documented interaction between P2X7Rs and Panx1HCs, it is tempting to hypothesize that chronic inactivity triggers the release of ATP by astrocytes, activating presynaptic P2X7Rs, increasing the entrance of Ca2+ to the presynaptic terminal, and activating presynaptic Panx1HCs. These channels may cooperate with P2X7Rs by increasing both the release of ATP (and potentiated P2X7R activation) and the entrance of Ca2+ to the presynaptic terminal by a positive feed-forward loop (Fig. 1). The precise functional crosstalk between these molecular components upon chronic inactivity awaits further studies. Thus, understanding the cell-specific mechanisms that control presynaptic P2X7Rs and Panx1HC function and interactions will enrich our knowledge about synaptic transmission.

P2X7Rs and Panx1HCs modulate presynaptic homeostatic plasticity: hypothetical pathway. Upon chronic inactivity, astrocytes sense the reduction of glutamate concentration at the synaptic cleft and, consequently, increase the amount of ATP released by Cx43HCs. Glial-released ATP activates presynaptic P2X7Rs, enhancing the amount of Ca2+ entering to the terminal and neurotransmitter release probability. Concomitantly, the increase in presynaptic Ca2+ activates neuronal Panx1HCs and thus ATP release, strengthening the activation of P2X7Rs by a positive feedback loop and promoting the compensatory increase in presynaptic function upon synaptic activity blockade
Concluding remarks
Since the discovery of ATP as a transmitter, our knowledge about the role of this nucleotide and the purinergic receptors, particularly P2X7Rs, in the modulation of synaptic strength and plasticity has increased significantly. Purinergic signaling has a crucial role in coordinating the molecular interaction between the components of the tripartite synapse and thus modulating presynaptic strength, and much progress has been made in how this interaction impacts neuronal physiology under basal and pathological conditions. On the other hand, Panx1HCs have emerged as important modulators of synaptic function under basal and activity-dependent conditions. Thus, we may speculate that the strong influence of P2X7Rs and pannexons in presynaptic function could rely on their functional interaction, which could be based on specific features that both channels share. For instance, P2X7Rs and Panx1HCs are both active under resting membrane potentials and after presynaptic depolarization. Also, the extracellular concentration of ATP fine-tunes their activation and thus membrane permeability. However, the precise mechanisms underlying their interaction need to be elucidated. Hence, deciphering the cell- and tissue-specific mechanisms sustaining the linking between P2X7Rs and Panx1HCs under certain conditions will be relevant to go deep in understanding synapse physiology in both health and disease.
Data availability
Not applicable.
References
Burnstock G (1976) Purinergic receptors. J Theor Biol 62:491–503. https://doi.org/10.1016/0022-5193(76)90133-8
Burnstock G, Satchell DG, Smythe A (1972) A comparison of the excitatory and inhibitory effects of non-adrenergic, non-cholinergic nerve stimulation and exogenously applied ATP on a variety of smooth muscle preparations from different vertebrate species. Br J Pharmacol 46:234–242. https://doi.org/10.1111/j.1476-5381.1972.tb06868.x
Burnstock G (2007) Physiology and pathophysiology of purinergic neurotransmission. Physiol Rev 87:659–797. https://doi.org/10.1152/physrev.00043.2006
Mori M, Heuss C, Gähwiler BH, Gerber U (2001) Fast synaptic transmission mediated by P2X receptors in CA3 pyramidal cells of rat hippocampal slice cultures. J Physiol 535:115–123
North R, a, (2002) Molecular physiology of P2X receptors. Physiol Rev 82:1013–1067. https://doi.org/10.1152/physrev.00015.2002
North RA, Verkhratsky A (2006) Purinergic transmission in the central nervous system. Eur J Physiol 452:479–485. https://doi.org/10.1007/s00424-006-0060-y
Cunha RA, Vizi ES, Ribeiro JA, Sebastião AM (1996) Preferential release of ATP and its extracellular catabolism as a source of adenosine upon high- but not low-frequency stimulation of rat hippocampal Slices. J Neurochem 67:2180–2187. https://doi.org/10.1046/j.1471-4159.1996.67052180.x
Wieraszko A, Goldsmith G, Seyfried TN (1989) Stimulation-dependent release of adenosine triphosphate from hippocampal slices. Brain Res 485:244–250. https://doi.org/10.1016/0006-8993(89)90567-2
Kang J, Kang N, Lovatt D et al (2008) Connexin 43 hemichannels are permeable to ATP. J Neurosci 28:4702–4711. https://doi.org/10.1523/jneurosci.5048-07.2008
Dahl G (2015) ATP release through pannexon channels. Phil Trans R Soc B 370. https://doi.org/10.1098/rstb.2014.0191
Burnstock G, Cocks T, Crowe R (1978) Evidence for purinergic innervation of the anococcygeus muscle. Br J Pharmacol 64:13–20. https://doi.org/10.1111/j.1476-5381.1978.tb08635.x
North RA, Barnard EA (1997) Nucleotide receptors. Curr Opin Neurobiol 7:346–357. https://doi.org/10.1016/s0959-4388(97)80062-1
Jarvis MF, Khakh BS (2009) Neuropharmacology ATP-gated P2X cation-channels. Neuropharmacology 56:208–215. https://doi.org/10.1016/j.neuropharm.2008.06.067
Martínez-cuesta MÁ, Blanch-ruiz MA, Ortega-luna R et al (2020) Structural and functional basis for understanding the biological significance of P2X7 receptor. Int J Mol Sci 21:1–23. https://doi.org/10.3390/ijms21228454
Sperlágh B, Vizi ES, Wirkner K, Illes P (2006) P2X7 receptors in the nervous system. Prog Neurobiol 78:327–346. https://doi.org/10.1016/j.pneurobio.2006.03.007
Sluyter R (2017) The P2X7 Receptor. Adv Exp Med Biol 1051:17–53. https://doi.org/10.1007/5584_2017_59
Surprenant A, Kawashima E, Rassendren F et al (1996) The cytolytic P2z receptor for extracellular ATP identified as a P2x receptor (P2X7). Science 272(80):735–738. https://doi.org/10.1126/science.272.5262.735
Sperlágh B, Illes P (2014) P2X7 receptor : an emerging target in central nervous system diseases. Trends Pharmacol Sci 35:537–547. https://doi.org/10.1016/j.tips.2014.08.002
Tewari M, Seth P (2015) Emerging role of P2X7 receptors in CNS health and disease. Ageing Res Rev 24:328–342. https://doi.org/10.1016/j.arr.2015.10.001
Khakh BS, Lester HA (1999) Dynamic selectivity filters in ion channels. Neuron 23:653–658. https://doi.org/10.1016/S0896-6273(01)80025-8
Virginio C, Mackenzie A, Rassendren FA et al (1999) Pore dilation of neuronal P2X receptor channels. Nat Neurosci 2:315–321. https://doi.org/10.1038/7225
Virginio C, Mackenzie A, North RA et al (1999) Kinetics of cell lysis, dye uptake and permeability changes in cells expressing the rat P2XÝ receptor. J Physiol 519:335–346. https://doi.org/10.1111/j.1469-7793.1999.0335m.x
Hibell AD, Kidd EJ, Chessell IP et al (2000) Apparent species differences in the kinetic properties of P2X 7 receptors. Br J Pharmacol 130:167–173. https://doi.org/10.1038/sj.bjp.0703302
Pelegrin P, Surprenant A (2006) Pannexin-1 mediates large pore formation and interleukin-1 release by the ATP-gated P2X7 receptor. EMBO J 25:5071–5082. https://doi.org/10.1038/sj.emboj.7601378
Di Virgilio F, Schmalzing G, Markwardt F (2018) The elusive P2X7 macropore. Trends Cell Biol 28:392–404. https://doi.org/10.1016/j.tcb.2018.01.005
Ugur M, Ugur Ö (2019) A mechanism-based approach to P2X7 receptor action. Mol Pharmacol 95:442–450. https://doi.org/10.1124/mol.118.115022
Di Virgilio F, Giuliani AL, Vultaggio-Poma V et al (2018) Non-nucleotide agonists triggering P2X7 receptor activation and pore formation. Front Pharmacol 9:1–10. https://doi.org/10.3389/fphar.2018.00039
Di Virgilio F, Dal Ben D, Sarti AC et al (2017) The P2X7 receptor in infection and inflammation. Immunity 47:15–31. https://doi.org/10.1016/j.immuni.2017.06.020
Ferrari D, Pizzirani C, Adinolfi E et al (2004) The antibiotic polymyxin B modulates P2X7 receptor function. J Immunol 173:4652–4660. https://doi.org/10.4049/jimmunol.173.7.4652
Karasawa A, Michalski K, Mikhelzon P, Kawate T (2017) The P2X7 receptor forms a dye-permeable pore independent of its intracellular domain but dependent on membrane lipid composition. Elife 6:1–22. https://doi.org/10.7554/eLife.31186
Duan S, Neary J (2006) P2X7 receptors: properties and relevance to CNS function. Glia 54:738–746. https://doi.org/10.1002/glia
Kim M, Jiang L, Wilson HL et al (2001) Proteomic and functional evidence for a P2X7 receptor signalling complex. EMBO J 20:6347–6358. https://doi.org/10.1093/emboj/20.22.6347
Denlinger LC, Fisette PL, Sommer JA et al (2001) Cutting Edge: The nucleotide receptor P2X7 contains multiple protein- and lipid-interaction motifs including a potential binding site for bacterial lipopolysaccharide. J Immunol 164:1871–1876. https://doi.org/10.4049/jimmunol.167.4.1871
Armstrong JN, Brust TB, Lewis RG, MacVicar B a (2002) Activation of presynaptic P2X7-like receptors depresses mossy fiber-CA3 synaptic transmission through p38 mitogen-activated protein kinase. J Neurosci 22:5938–5945 20026618
Panchin Y, Kelmanson I, Matz M et al (2000) A ubiquitous family of putative gap junction molecules. Curr Biol 10:473–474. https://doi.org/10.1016/s0960-9822(00)00576-5
Bruzzone R, Hormuzdi SG, Barbe MT et al (2003) Pannexins, a family of gap junction proteins expressed in brain. Proc Natl Acad Sci 100:13644–13649. https://doi.org/10.1073/pnas.2233464100
Panchin YV (2005) Evolution of gap junction proteins – the pannexin alternative. J Exp Biol 208:1415–1419. https://doi.org/10.1242/jeb.01547
Baranova A, Ivanov D, Petrash N et al (2004) The mammalian pannexin family is homologous to the invertebrate innexin gap junction proteins. Genomics 83:706–716. https://doi.org/10.1016/j.ygeno.2003.09.025
Ray A, Zoidl G, Weickert S et al (2005) Site-specific and developmental expression of pannexin1 in the mouse nervous system. Eur J Neurosci 21:3277–3290. https://doi.org/10.1111/j.1460-9568.2005.04139.x
Vogt A, Hormuzdi SG, Monyer H (2005) Pannexin1 and pannexin2 expression in the developing and mature rat brain. Mol Brain Res 141:113–120. https://doi.org/10.1016/j.molbrainres.2005.08.002
Yeung AK, Patil CS, Jackson MF (2020) Pannexin-1 in the CNS: emerging concepts in health and disease. J Neurochem 154:468–485. https://doi.org/10.1111/jnc.15004
Zoidl G, Petrasch-Parwez E, Ray A et al (2007) Localization of the pannexin1 protein at postsynaptic sites in the cerebral cortex and hippocampus. Neuroscience 146:9–16. https://doi.org/10.1016/j.neuroscience.2007.01.061
Locovei S, Bao L, Dahl G (2006) Pannexin 1 in erythrocytes: function without a gap. Proc Natl Acad Sci U S A 103:7655–7659. https://doi.org/10.1073/pnas.0601037103
Ambrosi C, Gassmann O, Pranskevich JN et al (2010) Pannexin1 and pannexin2 channels show quaternary similarities to connexons and different oligomerization numbers from each other. J Biol Chem 285:24420–24431. https://doi.org/10.1074/jbc.M110.115444
Michalski K, Syrjanen JL, Henze E et al (2020) The Cryo-EM structure of a pannexin 1 reveals unique motifs for ion selection and inhibition. Elife 9:1–14. https://doi.org/10.7554/eLife.54670
Deng Z, He Z, Maksaev G et al (2020) Cryo-EM structures of the ATP release channel pannexin 1. Nat Struct Mol Biol 27:373–381. https://doi.org/10.1038/s41594-020-0401-0
Qu R, Dong L, Zhang J et al (2020) Cryo-EM structure of human heptameric pannexin 1 channel. Cell Res 30:446–448. https://doi.org/10.1038/s41422-020-0298-5
Ruan Z, Orozco IJ, Du J (2020) Structures of human pannexin 1 reveal ion pathways and mechanism of gating. Nature 584:646–651. https://doi.org/10.1038/s41586-020-2357-y
Dahl G, Locovei S (2006) Pannexin: to gap or not to gap, is that a question? IUBMB Life 58:409–419. https://doi.org/10.1080/15216540600794526
Chiu Y, Ravichandran KS, Bayliss DA (2015) Intrinsic properties and regulation of pannexin 1 channel. Channels 8:1–7. https://doi.org/10.4161/chan.27545
Bao L, Locovei S, Dahl G (2004) Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS Lett 572:65–68. https://doi.org/10.1016/j.febslet.2004.07.009
Thompson RJ, Zhou N, Mac Vicar BA (2006) Ischemia opens neuronal gap junction hemichannels. Science (80- ) 312:924–927. https://doi.org/10.1126/science.1126241
Iglesias R, Dahl G, Qiu F et al (2009) Pannexin 1: the molecular substrate of astrocyte “hemichannels.” J Neurosci 29:7092–7097. https://doi.org/10.1523/jneurosci.6062-08.2009
Suadicani SO, Iglesias R, Wang J et al (2012) ATP signaling is deficient in cultured pannexin1-null mouse astrocytes. Glia 60:1106–1116. https://doi.org/10.1002/glia.22338
Chiu YH, Schappe MS, Desai BN, Bayliss DA (2018) Revisiting multimodal activation and channel properties of pannexin 1. J Gen Physiol 150:19–39. https://doi.org/10.1085/jgp.201711888
Ma W, Compan V, Zheng W et al (2012) Pannexin 1 forms an anion-selective channel. Pflugers Arch Eur J Physiol 463:585–592. https://doi.org/10.1007/s00424-012-1077-z
Romanov RA, Bystrova MF, Rogachevskaya OA et al (2012) The ATP permeability of pannexin 1 channels in a heterologous system and in mammalian taste cells is dispensable. J Cell Sci 125:5514–5523. https://doi.org/10.1242/jcs.111062
Garré JM, Bukauskas FF, Bennett MVL (2022) Single channel properties of pannexin-1 and connexin-43 hemichannels and P2X7 receptors in astrocytes cultured from rodent spinal cords. Glia 70:2260–2275. https://doi.org/10.1002/glia.24250
Chiu Y, Jin X, Medina CB et al (2017) A quantized mechanism for activation of pannexin channels. Nat Commun. https://doi.org/10.1038/ncomms14324
Dahl G (2018) The Pannexin1 membrane channel: distinct conformations and functions. FEBS Lett 592:3201–3209. https://doi.org/10.1002/1873-3468.13115
Dahl G, Keane RW (2012) Pannexin : from discovery to bedside in 11 4 years ? Brain Res 1487:150–159. https://doi.org/10.1016/j.brainres.2012.04.058
Cheung G, Chever O, Rouach N (2014) Connexons and pannexons: newcomers in neurophysiology. Front Cell Neurosci 8:1–19. https://doi.org/10.3389/fncel.2014.00348
Abudara V, Retamal MA, Del Rio R, Orellana JA (2018) Synaptic functions of hemichannels and pannexons: a double-edged sword. Front Mol Neurosci 11:1–24. https://doi.org/10.3389/fnmol.2018.00435
Barbe MT, Monyer H, Bruzzone R (2006) Cell-cell communication beyond connexins: the pannexin channels. Physiology 21:103–114. https://doi.org/10.1152/physiol.00048.2005
Sandilos JK, Bayliss DA (2012) Physiological mechanisms for the modulation of pannexin 1 channel activity. J Physiol 590:6257–6266. https://doi.org/10.1113/jphysiol.2012.240911
Isakson BE, Thompson RJ (2014) Pannexin-1 as a potentiator of ligand-gated receptor signaling. Channels 8:118–123. https://doi.org/10.4161/chan.27978
Whyte-fagundes P, Zoidl G (2017) Mechanisms of pannexin1 channel gating and regulation. BBA - Biomembr 1860:65–71. https://doi.org/10.1016/j.bbamem.2017.07.009
Thompson RJ, Jackson MF, Olah ME et al (2008) Activation of pannexin-1 hemichannels augments aberrant bursting in the hippocampus. Science 322:1555–1559. https://doi.org/10.1126/science.1165209
Garré JM, Retamal MA, Cassina P et al (2010) FGF-1 induces ATP release from spinal astrocytes in culture and opens pannexin and connexin hemichannels. Proc Natl Acad Sci U S A 107:22659–22664. https://doi.org/10.1073/pnas.1013793107
Billaud M, Lohman AW, Straub AC et al (2011) Pannexin1 regulates alpha1-adrenergic receptor– mediated vasoconstriction. Circ Res 109:80–85. https://doi.org/10.1161/CIRCRESAHA.110.237594
Orellana JA, Figueroa XF, Sanchez HA et al (2011) Hemichannels in the neurovascular unit and white matter under normal and inflamed conditions. CNS Neurol Disord - Drug Targets 10:404–414. https://doi.org/10.2174/187152711794653869
Locovei S, Scemes E, Qiu F et al (2007) Pannexin1 is part of the pore forming unit of the P2X7 receptor death complex. FEBS Lett 581:483–488. https://doi.org/10.1016/j.biotechadv.2011.08.021.Secreted
Locovei S, Wang J, Dahl G (2006) Activation of pannexin 1 channels by ATP through P2Y receptors and by cytoplasmic calcium. FEBS Lett 580:239–244. https://doi.org/10.1016/j.febslet.2005.12.004
Boyce AKJ, Kim MS, Wicki-Stordeur LE, Swayne LA (2015) ATP stimulates pannexin 1 internalization to endosomal compartments. Biochem J 470:319–330. https://doi.org/10.1042/BJ20141551
Boyce AKJ, Swayne LA (2017) P2X7 receptor crosstalk regulates ATP-induced pannexin 1 internalization Andrew. Biochem J 474:2133–2144. https://doi.org/10.1042/BCJ20170257
Qiu F, Dahl G (2009) A permeant regulating its permeation pore: inhibition of pannexin 1 channels by ATP. Am J Physiol - Cell Physiol 296:250–255. https://doi.org/10.1152/ajpcell.00433.2008
Flores-Muñoz C, García-Rojas F, Pérez MA et al (2022) The long-term pannexin 1 ablation produces structural and functional modifications in hippocampal neurons. Cells 11:1–31. https://doi.org/10.3390/cells11223646
Lai CPK, Bechberger JF, Thompson RJ et al (2007) Tumor-suppressive effects of pannexin 1 in C6 glioma cells. Cancer Res 67:1545–1554. https://doi.org/10.1158/0008-5472.CAN-06-1396
Vanden Abeele F, Bidaux G, Gordienko D et al (2006) Functional implications of calcium permeability of the channel formed by pannexin 1. J Cell Biol 174:535–546. https://doi.org/10.1083/jcb.200601115
Boassa D, Ambrosi C, Qiu F et al (2007) Pannexin1 channels contain a glycosylation site that targets the hexamer to the plasma membrane. J Biol Chem 282:31733–31743. https://doi.org/10.1074/jbc.M702422200
Sosinsky GE, Boassa D, Dermietzel R et al (2011) Pannexin channels are not gap junction hemichannels. Channels 5:193–197. https://doi.org/10.4161/chan.5.3.15765
Penuela S, Simek J, Thompson RJ (2014) Regulation of pannexin channels by post-translational modifications. In: FEBS Letters 1411–1415. https://doi.org/10.1016/j.febslet.2014.01.028
Sahu G, Sukumaran S, Bera AK (2014) Pannexins form gap junctions with electrophysiological and pharmacological properties distinct from connexins. Sci Rep 4:1–9. https://doi.org/10.1038/srep04955
Palacios-Prado N, Soto PA, Lopez X et al (2022) Endogenous pannexin1 channels form functional intercellular cell-cell channels with characteristic voltage-dependent properties. Proc Natl Acad Sci U S A 119:1–11. https://doi.org/10.1073/pnas.2202104119
Suadicani SO, Brosnan CF, Scemes E (2006) P2X7 Receptors mediate ATP release and amplification of astrocytic intercellular Ca2+ signaling. J Neurosci 26:1378–1385. https://doi.org/10.1523/JNEUROSCI.3902-05.2006
Yan Z, Li S, Liang Z et al (2008) The P2X 7 receptor channel pore dilates under physiological ion conditions. J Gen Physiol 132:563–573. https://doi.org/10.1085/jgp.200810059
Rigato C, Swinnen N, Buckinx R et al (2012) Microglia proliferation is controlled by P2X7 receptors in a pannexin-1-independent manner during early embryonic spinal cord invasion. J Neurosci 32:11559–11573. https://doi.org/10.1523/JNEUROSCI.1042-12.2012
De Vuyst E, Decrock E, De Bock M et al (2007) Connexin hemichannels and gap junction channels are differentially influenced by lipopolysaccharide and basic fibroblast growth factor. Mol Biol Cell 18:34–36. https://doi.org/10.1091/mbc.E06
Anselmi F, Hernandez VH, Crispino G et al (2008) ATP release through connexin hemichannels and gap junction transfer of second messengers propagate Ca2+ signals across the inner ear. Proc Natl Acad Sci 105:18770–18775. https://doi.org/10.1073/pnas.0800793105
Sorge RE, Trang T, Dorfman R et al (2012) Genetically determined P2X7 receptor pore formation regulates variability in chronic pain sensitivity. Nat Med 18:595–599. https://doi.org/10.1038/nm.2710
Purohit R, Bera AK (2021) Pannexin 1 plays a pro-survival role by attenuating P2X7 receptor-mediated Ca2+ influx. Cell Calcium 99:102458. https://doi.org/10.1016/j.ceca.2021.102458
Purohit R, Bera AK (2023) Carboxyl terminus of pannexin-1 plays a crucial role in P2X7 receptor-mediated signaling. Biochem Biophys Res Commun 664:20–26. https://doi.org/10.1016/j.bbrc.2023.04.081
Deuchars SA, Atkinson L, Brooke RE, et al (2001) Neuronal P2X7 receptors are targeted to presynaptic terminals in the central and peripheral nervous systems. J Neurosci 21:7143–7152 21/18/7143 [pii]
Sperlagh B, Kofalvi A, Deuchars J et al (2002) Involvement of P2X7 receptors in the regulation of neurotransmitter release in the rat hippocampus. J Neurochem 81:1196–1211. https://doi.org/10.1046/j.1471-4159.2002.00920.x
Atkinson L, Batten TFC, Moores TS et al (2004) Differential co-localisation of the P2X7 receptor subunit with vesicular glutamate transporters VGLUT1 and VGLUT2 in rat CNS. Neuroscience 123:761–768. https://doi.org/10.1016/j.neuroscience.2003.08.065
Cho J, Choi I, Jang I (2010) P2X7 receptors enhance glutamate release in hippocampal hilar neurons. Cell Mol Dev Neurosci 21:865–870. https://doi.org/10.1097/WNR.0b013e32833d9142
Zhang PA, Xu QY, Xue L et al (2017) Neonatal maternal deprivation enhances presynaptic p2x7 receptor transmission in insular cortex in an adult rat model of visceral hypersensitivity. CNS Neurosci Ther 23:145–154. https://doi.org/10.1111/cns.12663
Miras-Portugal MT, Ortega F, Gómez-Villafuertes R, et al (2021) P2X7 receptors in the central nervous system. Biochem Pharmacol 187. https://doi.org/10.1016/j.bcp.2021.114472
Rafael A, Cairus A, Tizzoni M et al (2020) Glial ATP and large pore channels modulate synaptic strength in response to chronic inactivity. Mol Neurobiol 57:2856–2869. https://doi.org/10.1007/s12035-020-01919-0
Alloisio S, Cervetto C, Passalacqua M et al (2008) Functional evidence for presynaptic P2X7 receptors in adult rat cerebrocortical nerve terminals. FEBS Lett 582:3948–3953. https://doi.org/10.1016/j.febslet.2008.10.041
Marcoli M, Cervetto C, Paluzzi P et al (2008) P2X7 pre-synaptic receptors in adult rat cerebrocortical nerve terminals: a role in ATP-induced glutamate release. J Neurochem 105:2330–2342. https://doi.org/10.1111/j.1471-4159.2008.05322.x
León D, Sánchez-Nogueiro J, Marín-García P, Miras-Portugal MT (2008) Glutamate release and synapsin-I phosphorylation induced by P2X7receptors activation in cerebellar granule neurons. Neurochem Int 52:1148–1159. https://doi.org/10.1016/j.neuint.2007.12.004
Miras-Portugal MT, Díaz-Hernández M, Giráldez L et al (2003) P2X7 receptors in rat brain: presence in synaptic terminals and granule cells. Neurochem Res 28:1597–1605. https://doi.org/10.1023/A:1025690913206
Egan TM, Samways DSK, Li Z (2006) Biophysics of P2X receptors. Pflugers Arch Eur J Physiol 452:501–512. https://doi.org/10.1007/s00424-006-0078-1
Lundy PM, Hamilton MG, Mi L et al (2002) Stimulation of Ca 2+ influx through ATP receptors on rat brain synaptosomes: identification of functional P2X 7 receptor subtypes. Br J Pharmacol 135:1616–1626. https://doi.org/10.1038/sj.bjp.0704624
Ireland MF, Noakes PG, Bellingham MC (2004) P2X 7-like receptor subunits enhance excitatory synaptic transmission at central synapses by presynaptic mechanisms. Neuroscience 128:269–280. https://doi.org/10.1016/j.neuroscience.2004.06.014
Miteva AS, Gaydukov AE, Shestopalov VI, Balezina OP (2018) Mechanism of P2X7 receptor-dependent enhancement of neuromuscular transmission in pannexin 1 knockout mice. Purinergic Signal 14:459–469. https://doi.org/10.1007/s11302-018-9630-7
Miteva A, Gaydukov A, Balezina O (2020) Interaction between calcium chelators and the activity of P2X7 receptors in mouse motor synapses. Int J Mol Sci 21. https://doi.org/10.3390/ijms21062034
Ardiles AO, Flores-Muñoz C, Toro-Ayala G et al (2014) Pannexin 1 regulates bidirectional hippocampal synaptic plasticity in adult mice. Front Cell Neurosci 8:1–11. https://doi.org/10.3389/fncel.2014.00326
Kurtenbach S, Prochnow N, Kurtenbach S et al (2013) Pannexin1 channel proteins in the zebrafish retina Have shared and unique properties. PLoS One 8:e77722. https://doi.org/10.1371/journal.pone.0077722
García-Rojas F, Flores-Muñoz C, Santander O et al (2023) Pannexin-1 modulates inhibitory transmission and hippocampal synaptic plasticity. Biomolecules 13:1–17. https://doi.org/10.3390/biom13060887
Santos DA, Salgado AI, Cunha RA (2003) ATP is released from nerve terminals and from activated muscle fibres on stimulation of the rat phrenic nerve. Neurosci Lett 338:225–228. https://doi.org/10.1016/S0304-3940(02)01419-2
Miteva AS, Gaydukov AE, Shestopalov VI, Balezina OP (2017) The role of pannexin 1 in the purinergic regulation of synaptic transmission in mouse motor synapses. Biochem Suppl Ser A Membr Cell Biol 11:311–320. https://doi.org/10.1134/S1990747817040067
Cunha RA, Ribeiro JA (2000) ATP as a presynaptic modulator. Life Sci 68:119–137. https://doi.org/10.1016/S0024-3205(00)00923-1
Ribeiro JA, Walker J (1975) The effects of adenosine triphosphate and adenosine diphosphate on transmission at the rat and frog neuromuscular junctions. Br J Pharmacol 54:213–218. https://doi.org/10.1111/j.1476-5381.1975.tb06931.x
Momboisse F, Olivares MJ, Báez-Matus X et al (2014) Pannexin 1 channels: new actors in the regulation of catecholamine release from adrenal chromaffin cells. Front Cell Neurosci 8:1–12. https://doi.org/10.3389/fncel.2014.00270
Maldifassi MC, Momboisse F, Guerra MJ et al (2021) The interplay between α7 nicotinic acetylcholine receptors, pannexin-1 channels and P2X7 receptors elicit exocytosis in chromaffin cells. J Neurochem 157:1789–1808. https://doi.org/10.1111/jnc.15186
Sokolova E, Nistri A, Giniatullin R (2001) Negative cross talk between anionic GABA A and cationic P2X ionotropic receptors of rat dorsal root ganglion neurons. J Neurosci 21:4958–4968. https://doi.org/10.1523/JNEUROSCI.21-14-04958.2001
Dvoriantchikova G, Ivanov D, Panchin Y, Shestopalov VI (2006) Expression of pannexin family of proteins in the retina. FEBS Lett 580:2178–2182. https://doi.org/10.1016/j.febslet.2006.03.026
Riquelme MA, Cea LA, Vega JL et al (2013) The ATP required for potentiation of skeletal muscle contraction is released via pannexin hemichannels. Neuropharmacology 75:594–603. https://doi.org/10.1016/j.neuropharm.2013.03.022
Sokolova E, Grishin S, Shakirzyanova A et al (2003) Distinct receptors and different transduction mechanisms for ATP and adenosine at the frog motor nerve endings. Eur J Neurosci 18:1254–1264. https://doi.org/10.1046/j.1460-9568.2003.02835.x
Li Q, Barres BA (2018) Microglia and macrophages in brain homeostasis and disease. Nat Publ Gr 18:225–242. https://doi.org/10.1038/nri.2017.125
Liddelow SA, Guttenplan KA, Clarke LE et al (2017) Neurotoxic reactive astrocytes are induced by activated microglia. Nature 541:481–487. https://doi.org/10.1038/nature21029
Barres BA (2008) The mystery and magic of glia: A perspective on their roles in health and disease. Neuron 60:430–440. https://doi.org/10.1016/j.neuron.2008.10.013
Araque A, Navarrete M (2010) Glial cells in neuronal network function. Philos Trans R Soc B Biol Sci 365:2375–2381. https://doi.org/10.1098/rstb.2009.0313
Durkee CA, Araque A (2019) Diversity and specificity of astrocyte–neuron communication. Neuroscience 396:73–78. https://doi.org/10.1016/j.neuroscience.2018.11.010
Araque A, Parpura V, Sanzgiri RP, Haydon PG (1999) Tripartite synapses: glia, the unacknowledged partner. Trends Neurosci 22:208–215. https://doi.org/10.1016/s0166-2236(98)01349-6
Perea G, Navarrete M, Araque A (2009) Tripartite synapses: astrocytes process and control synaptic information. Trends Neurosci 32:421–431. https://doi.org/10.1016/j.tins.2009.05.001
Stevens ER, Esguerra M, Kim PM et al (2003) D-serine and serine racemase are present in the vertebrate retina and contribute to the physiological activation of NMDA receptors. Proc Natl Acad Sci 100:6789–6794
Wolosker H, Blackshaw S, Snyder SH (1999) Serine racemase: a glial enzyme synthesizing D-serine to regulate glutamate-N-methyl-D-aspartate neurotransmission. Proc Natl Acad Sci 96:13409–13414. https://doi.org/10.1073/pnas.96.23.13409
Yang Y, Ge W, Chen Y et al (2003) Contribution of astrocytes to hippocampal long-term potentiation through release of D-serine. Proc Natl Acad Sci 100:15194–15199. https://doi.org/10.1073/pnas.2431073100
Yang S, Qiao H, Wen L et al (2005) D-Serine enhances impaired long-term potentiation in CA1 subfield of hippocampal slices from aged senescence-accelerated mouse prone/8. Neurosci Lett 379:7–12. https://doi.org/10.1016/j.neulet.2004.12.033
Panatier A, Gentles SJ, Bourque CW, Oliet SHR (2006) Activity-dependent synaptic plasticity in the supraoptic nucleus of the rat hypothalamus. J Physiol 573:711–721. https://doi.org/10.1113/jphysiol.2006.109447
Henneberger C, Papouin T, Oliet SHR, Rusakov DA (2010) Long-term potentiation depends on release of d-serine from astrocytes. Nature 463:232–236. https://doi.org/10.1038/nature08673
Martineau M, Parpura V, Mothet JP (2014) Cell-type specific mechanisms of D-serine uptake and release in the brain. Front. Synaptic Neurosci. 6:1–9. https://doi.org/10.3389/fnsyn.2014.00012
Sherwood MW, Oliet SHR, Panatier A (2021) NMDARs, coincidence detectors of astrocytic and neuronal activities. Int J Mol Sci 22:1–23. https://doi.org/10.3390/ijms22147258
Pan HC, Chou YC, Sun SH (2015) P2X7R-mediated Ca2+-independent d-serine release via pannexin-1 of the P2X7R-pannexin-1 complex in astrocytes. Glia 63:877–893. https://doi.org/10.1002/glia.22790
Bademosi AT, Lauwers E, Padmanabhan P et al (2017) In vivo single-molecule imaging of syntaxin1A reveals polyphosphoinositide- and activity-dependent trapping in presynaptic nanoclusters. Nat Commun 7:13660. https://doi.org/10.1038/ncomms13660
Mothet JP, Pollegioni L, Ouanounou G et al (2005) Glutamate receptor activation triggers a calcium-dependent and SNARE protein-dependent release of the gliotransmitter D-serine. Proc Natl Acad Sci 102:5606–5611. https://doi.org/10.1073/pnas.0408483102
Li YH, Han TZ (2007) Glycine binding sites of presynaptic NMDA receptors may tonically regulate glutamate release in the rat visual cortex. J Neurophysiol 97:817–823. https://doi.org/10.1152/jn.00980.2006
Ohi Y, Kimura S, Haji A (2015) Modulation of glutamatergic transmission by presynaptic N-methyl-d-aspartate mechanisms in second-order neurons of the rat nucleus tractus solitarius. Neurosci Lett 587:62–67. https://doi.org/10.1016/j.neulet.2014.12.031
Lench A, Massey P V, Pollegioni L, et al (2014) Neuropharmacology Astroglial D -serine is the endogenous co-agonist at the presynaptic NMDA receptor in rat entorhinal cortex. Neuropharmacology 1–9. https://doi.org/10.1016/j.neuropharm.2014.04.004
Kandel ER, Schwartz JH, Jessell TM (2000) Principles of neural science (4th ed). McGraw Hill, Health Professions Division, New York
Amir R, Devor M (2003) Electrical excitability of the soma of sensory neurons is required for spike invasion of the soma, but not for through-conduction. Biophys J 84:2181–2191. https://doi.org/10.1016/S0006-3495(03)75024-3
Hanani M (2005) Satellite glial cells in sensory ganglia: from form to function. Brain Res Rev 48:457–476. https://doi.org/10.1016/j.brainresrev.2004.09.001
Pannese E (1981) The satellite cells of the sensory ganglia. Adv Anat Embryol Cell Biol 65:1–111. https://doi.org/10.1007/978-3-642-67750-2
Zhang X, Chen Y, Wang C, Huang LYM (2007) Neuronal somatic ATP release triggers neuron-satellite glial cell communication in dorsal root ganglia. Proc Natl Acad Sci 104:9864–9869. https://doi.org/10.1073/pnas.0611048104
Gu Y, Chen Y, Zhang X et al (2010) Neuronal soma-satellite glial cell interactions in sensory ganglia and the participation of purinergic receptors. Neuron Glia Biol. 6:53–62. https://doi.org/10.1017/S1740925X10000116
Retamal MA, Alcayaga J, Verdugo CA et al (2014) Opening of pannexin- and connexin-based channels increases the excitability of nodose ganglion sensory neurons. Front Cell Neurosci 8:1–12. https://doi.org/10.3389/fncel.2014.00158
Chen Y, Zhang X, Wang C et al (2008) Activation of P2X7 receptors in glial satellite cells reduces pain through downregulation of P2X3 receptors in nociceptive neurons. Proc Natl Acad Sci 105:16773–16778. https://doi.org/10.1073/pnas.0801793105
Hanstein R, Hanani M, Scemes E, Spray DC (2016) Glial pannexin1 contributes to tactile hypersensitivity in a mouse model of orofacial pain. Sci Rep 6:1–10. https://doi.org/10.1038/srep38266
Hanani M, Spray DC (2020) Emerging importance of satellite glia in nervous system function and dysfunction. Nat Rev Neurosci 21:485–498
Chen Z, Zhang C, Song X et al (2022) Bzatp activates satellite glial cells and increases the excitability of dorsal root ganglia neurons in vivo. Cells 11:1–17. https://doi.org/10.3390/cells11152280
Hanstein R, Negoro H, Patel NK et al (2013) Promises and pitfalls of a Pannexin1 transgenic mouse line. Front Pharmacol 4:1–10. https://doi.org/10.3389/fphar.2013.00061
Feldman-Goriachnik R, Belzer V, Hanani M (2015) Systemic inflammation activates satellite glial cells in the mouse nodose ganglion and alters their functions. Glia 63:2121–2132. https://doi.org/10.1002/glia.22881
Zhang Y, Laumet G, Chen SR et al (2015) Pannexin-1 up-regulation in the dorsal root ganglion contributes to neuropathic pain development. J Biol Chem 290:14647–14655. https://doi.org/10.1074/jbc.M115.650218
Zhang XF, Han P, Faltynek CR et al (2005) Functional expression of P2X7 receptors in non-neuronal cells of rat dorsal root ganglia. Brain Res 1052:63–70. https://doi.org/10.1016/j.brainres.2005.06.022
Chen Y, Li G, Huang LYM (2012) P2X7 receptors in satellite glial cells mediate high functional expression of P2X3 receptors in immature dorsal root ganglion neurons. Mol Pain 8:1–9. https://doi.org/10.1186/1744-8069-8-9
Turrigiano GG (2008) The self-tuning neuron: synaptic scaling of excitatory synapses. Cell 135:422–435. https://doi.org/10.1016/j.cell.2008.10.008
Pozo K, Goda Y (2010) Unraveling mechanisms of homeostatic synaptic plasticity. Neuron 66:337–351. https://doi.org/10.1016/j.neuron.2010.04.028
Vitureira N, Goda Y (2013) The interplay between Hebbian and homeostatic synaptic plasticity. J Cell Biol 203:175–186. https://doi.org/10.1083/jcb.201306030
Stellwagen D, Malenka RC (2006) Synaptic scaling mediated by glial TNF-α. Nature 440:1054–1059. https://doi.org/10.1038/nature04671
Beattie EC, Stellwagen D, Morishita W et al (2002) Control of synaptic strength by glial TNF alpha. Science (80- ) 295:2282–2285. https://doi.org/10.1126/science.1067859
Vlachos A, Ikenberg B, Lenz M et al (2013) Synaptopodin regulates denervation-induced homeostatic synaptic plasticity. Proc Natl Acad Sci 110:8242–8247. https://doi.org/10.1073/pnas.121367711010.1073/pnas.1213677110
Becker D, Zahn N, Deller T, Vlachos A (2013) Tumor necrosis factor alpha maintains denervation-induced homeostatic synaptic plasticity of mouse dentate granule cells. Front Cell Neurosci 7:1–10. https://doi.org/10.3389/fncel.2013.00257
Heir R, Stellwagen D (2020) TNF-mediated homeostatic synaptic plasticity: from in vitro to in vivo models. Front Cell Neurosci 14:1–12. https://doi.org/10.3389/fncel.2020.565841
Pankratov Y, Lalo U, Krishtal OA, Verkhratsky A (2009) P2X receptors and synaptic plasticity. Neuroscience 158:137–148. https://doi.org/10.1016/j.neuroscience.2008.03.076
Khakh BS (2001) Molecular physiology of P2X receptors and ATP signalling at synapses. Nat Rev Neurosci 2:165–174. https://doi.org/10.1038/35058521
Zhao CJ, Dreosti E, Lagnado L (2011) Homeostatic synaptic plasticity through changes in presynaptic calcium influx. J Neurosci 31:7492–7496. https://doi.org/10.1523/JNEUROSCI.6636-10.2011
Lazarevic V, Schöne C, Heine M et al (2011) Extensive remodeling of the presynaptic cytomatrix upon homeostatic adaptation to network activity silencing. J Neurosci 31:10189–10200. https://doi.org/10.1523/JNEUROSCI.2088-11.2011
Rátkai A, Tárnok K, El AH et al (2021) Homeostatic plasticity and burst activity are mediated by hyperpolarization-activated cation currents and T-type calcium channels in neuronal cultures. Sci Rep 11:1–17. https://doi.org/10.1038/s41598-021-82775-3
Acknowledgements
A.R. received a PhD Fellowship from the Agencia Nacional de Investigación e Innovación (ANII-MEC).
Funding
This work was supported by grants to N.V., V.A. and AR by the Comisión Sectorial de Investigación Científica (CSIC-UdelaR, Uruguay, grant number: 22520220100095UD, 22520220100243UD and 22320200200227UD, respectively) and by the Programa de Desarrollo de las Ciencias Básicas (PEDECIBA, Uruguay).
Author information
Authors and Affiliations
Contributions
All authors contributed to the manuscript conception and design. The manuscript was written by both NV and VA and all authors reviewed the manuscript. AR prepared the figure. All authors read and approved the manuscript.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Vitureira, N., Rafael, A. & Abudara, V. P2X7 receptors and pannexin1 hemichannels shape presynaptic transmission. Purinergic Signalling 20, 223–236 (2024). https://doi.org/10.1007/s11302-023-09965-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11302-023-09965-8