
Abstract
The success of a genetic improvement program is contingent on developing genotypes with superior performance in productivity and/or quality across varied environmental conditions. In order to provide a better insight of the dynamics of genotype-by-environment (GxE) interaction on 975 loblolly pine (Pinus taeda L.) clones, the present study evaluated a series of six trials in the Southeast USA that were measured for total stem volume and survival under two silvicultural treatments (operational and intensive). Objectives included the following: (1) estimate the magnitude of the GxE and to understand its dynamics; (2) estimate and compare type B genetic correlations and clonal genetic values based on and one- and two-stage analyses and factor analytic analysis; (3) obtain clonal rankings for each site, and across all sites and calculate potential genetic gains; and (4) explore relationships between genotypes and environments through Biplots. Results indicate similar ranking for genotypes selected in each of the two silvicultural treatments (clonal genetic correlations >0.89 for volume). Important levels of GxE interaction for volume and survival were detected for clones in the one-stage analyses (average type B genetic correlation estimates of 0.63 and 0.57 for VOL and SURV, respectively). The one- and two-stage analyses provided similar genetic correlations, rankings, and breeding values. Factor analytic error structure was appropriate to model complex GxE interaction. One-stage analyses produced higher heritabilities than two-stage approaches with these data from six sites. Biplots summarized the GxE interaction successfully and confirmed the positive correlation between all environments for volume and survival.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The success of any genetic improvement program depends on producing genotypes with superior performance in terms of productivity and quality across multiple environmental conditions. To achieve this goal, it is necessary to understand the phenotype, which is the result of genotype performance and environmental conditions (Malosetti et al. 2013). Genotypes differ in efficiency to capture and convert environmental inputs, a difference that is determined by its particular ensemble of genes. Usually, tree breeders are interested in the phenotype for a given trait at a particular age, which is a cumulative result of casual interactions between the plant genotype and its environment.
One of the main objectives in plant breeding is to match genotypes to specific environments in such a way that the response is optimized. Some genotypes can do well across most conditions; however, there are some genotypes that do better (or worse) than others exclusively under a specific set of environmental conditions or managements. This adaptation is related to an interaction denominated genotype-by-environment (GxE) (Malossetii et al. 2013). GxE interaction is a differential response of a genotype in an environment and it is important in several aspects. For most genetic improvement programs, this GxE interaction is valuable information, among others, to: (1) evaluate the stability of genotype’s response across their deployment area, (2) define breeding strategies, (3) evaluate breeding zones, (4) maximize genetic gain, and (5) increase accuracy of predictions as GxE modeling incorporates the structure of the data into the statistical model.
GxE interaction is one of the aspects that all breeders consider before making deployment decisions, as its presence may result in different ranking of genotypes for each site (or environment) tested, and therefore, it is seen as a lack of performance consistency. For this reason, presence of GxE interactions reduces selection progress, as difficulties arise on identifying the best genotype across all environments (Comstock and Moll 1963). However, the importance of this GxE interaction will depend on its specific implications for a breeding program, where, in some cases, it might be ignored (even if statistically significant).
The estimation of the magnitude of GxE interaction, and its pattern, for a given trait and population can be determined by using different statistical approaches, such as variance component estimation (Freeman 1973; Burdon 1977; Muir et al. 1992), type B correlations of genetic values of given genotypes for the same trait in different environments (Burdon 1977; Muir et al. 1992), multi-environment trials (MET) analyses in one - stage (Silva et al. 2014; Malosetti et al. 2013; Costa e Silva et al. 2006; Hardner et al. 2009; Cullis et al. 2014) or two - stages (Mohring and Piepho 2009), principal component analysis with combination of Biplots (Malosetti et al. 2013), and regression models (Freeman 1973). These approaches help to investigate GxE dynamics across all evaluated sites or environments and assist with the selection of genotypes.
Variance component estimation, by fitting linear mixed models (LMM) in MET analyses, is a common and powerful statistical tool used to partition the different sources of variability in the phenotype response, which are later used to estimate heritability and to calculate predictions of genetic values (i.e., best linear unbiased prediction, BLUP). For these analyses, presence of GxE interaction is often identified as a result of a large and significant variance component associated with the factor that models GxE; however, GxE interaction can also be identified as different genetic variances for each environment (Silva et al. 2014). Here, better environments tend to have larger genetic variances than poor environments, but the opposite can happen as well (Malosetti et al. 2013). Type B genetic correlation is also an important measure of the magnitude of the GxE interaction (Burdon 1977; Silva et al. 2014), ranging from −1 to 1, where values close to 0 indicate weak agreement.
MET analyses are commonly used in forest tree breeding programs and can be performed by using two different, but complementary, LMMs, identified as explicit and implicit. The explicit model provides with a predicted genetic value for each of the genotypes (individual or clones) across all sites evaluated; it also provides with a deviation (or interaction) of each genotype within a site. This model assumes that type B correlation is the same across any pair of sites limiting the understanding of the GxE across sites. Nevertheless, this is the most frequently fitted model, it is often easy to converge, and is preferred whenever a large number of sites is evaluated (Frensham et al. 1997; Smith et al. 2001).
In contrast, the implicit model, which provides predicted genetic values for each site, contains within the linear model a single genetic term that predicts each individual genotype within a site, together with a variance-covariance matrix of these genetic effects between pairs of sites. There are multiple forms of this matrix that can be specified, where the simplest is compound symmetry (CS). CS is completely equivalent to the explicit model, and if this matrix is expressed as a correlation matrix, it leads to be exactly the same type B correlation value. Nevertheless, more complex forms can be fitted (see for example, Cullis et al. 2006), where the most complex form estimates a different genetic variance for each site and a unique covariance for each pair of sites (often known as unstructured). However, given the large number of parameters that need to be estimated whenever many sites are analyzed simultaneously, some authors have suggested other simpler forms that approximate the unstructured matrix, such as factor analytic (Thompson et al. 2003; Meyer 2009).
The advantage of specifying a complex variance-covariance matrix is that it permits further understanding of the structure of the GxE interaction by looking at individual covariances (or correlations) between pairs of sites. In addition, other statistical tools, such as principal component analysis (PCA), can be used to decompose this matrix to facilitate the understanding of the relationship between the genotypes and/or sites. PCA can also be complemented with the use of Biplot (Gabriel 1971) to help group environments and identify specific clusters of genotypes with a similar response (Malosetti et al. 2013). Biplot is often used in the context of principal components but can also be used in statistical modeling. It is a multidimensional graphic composed by lines and dots where the lines are variables and dots are observations (Kohler and Luniak 2005).
Most statistical analyses of METs are computationally and statistically challenging, as they need to combine sites with different levels of connectivity and precision into an analysis that accommodates the varied design structures, background errors, and individual-site heritabilites. A one-stage analysis approach fits a single LMM for all data simultaneously in a single step. For situations with a limited number of sites and simple genetic variance-covariance matrices (e.g., compound symmetry), this can be done on a routine basis. However, often, it is not possible to fit this model as convergence issues arise. An alternative, particularly for large number of sites, is to perform a two-stage analysis that separates the evaluation in two steps. Both one- and two-stage analyses have practical and statistical advantages and disadvantages that depend on the connectivity among sites, number of genotypes considered, computational limitations, level of precision desired, among other things. Malosetti et al. (2013) indicated that one-stage analyses have theoretical advantages; however, two-stage analyses are computationally and logistically easier to fit. Two-stage analysis was originally proposed by Patterson (1978), and it has been extended and modified by Cullis et al. (1996) and Piepho et al. (2012). This type of analysis involves first fitting individual linear models to each of the sites and obtaining genotype mean predictions (i.e., least square means). Later, these predictions are combined, in a second step, together with their corresponding weights (or standard errors) into a MET analysis. Weights are expected to be required here as the genotype mean predictions from the first stage have different levels of precision, due to, for example, number of replications and level of relatedness with other genotypes present in the same trial.
In order to provide better insight of the statistical methods used to explore GxE interaction, the present study evaluated a series of six loblolly pine (Pinus taeda L.) clonal trials established in the Southeast USA that were measured for total stem volume and survival. This dataset provided a unique opportunity to explore both interactions between genotypes and cultural treatments and interactions between genotypes and sites. Furthermore, it represents an opportunity to rank a large set of clones over many sites with reasonable replication at each site.
The main objective of this study is to explore GxE interaction for this clonal population by fitting a range of LMMs. Some of the specific objectives are as follows: (1) estimate the magnitude and dynamics of the GxE interaction; (2) estimate and compare type B genetic correlations and clonal genetic values based on a one- and two-stage analyses, to contrast unstructured and factor analytic variance-covariance structures; (3) estimate variance-covariance matrices and clonal rankings for each site, and across all sites, and calculate potential genetic gains; and (4) use Biplot analysis to explore relationships between genotypes and environments.
Materials and methods
Genetic material and experimental layout
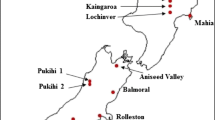
The dataset used in this study originates from a series of six genetic trials established by the Forest Biology Research Cooperative (FBRC) from the University of Florida known as the CCLONES Series 1 (Baltunis et al. 2007). The six sites considered were as follows: BF Grant, GA (BFG), Cuthbert, GA (CUT), Nassau, FL (NAS), Oakfield, GA (OAK), Palatka, FL (PAL), and Santa Rosa, FL (SNR) (Fig. 1). This study contains 61 full-sib loblolly pine families planted in single-tree plots as an incomplete block design with eight complete full block replicates per site. For all but two sites, four contiguous blocks were assigned (i.e., left and right sides) to two different silvicultural treatments (operational and intensive management). All sites have both silvicultural (cultural) treatments, except for CUT which only had the intensive and SNR which only had the operational. Here, the operational managements represent the typical cultural practices, where the intensive entails both more frequent weed control and fertilizer applications. Unfortunately, this silvicultural treatment leads to potential confounding of treatment and block effects. Each site contains ∼80 % cuttings and ∼20 % seedlings from each of the tested full-sib families. However, for this study, only measurements that correspond to cuttings were considered, and only from those clones that were present in at least three sites. Hence, a total of 61 full-sib families were considered in this analysis for a total of 975 clones, i.e., ∼16 clones per family.

Map of the geographic location of each site for the CCLONES loblolly Pine Series 1 study. The site abbreviations are as follows: BF Grant (BFG), Cuthbert (CUT), Nassau (NAS), Oakfield (OAK), Palatka (PAL), and Santa Rosa (SNR)
The variables evaluated were total stem volume (VOL, dm3) and survival (SURV, %). Total stem volume was calculated as follows:
where DBH (dm) is the diameter at breast height and Ht (dm) is the total height. SURV was recorded as 1 for alive trees and 0 for dead trees. For each site, the latest available measurement was considered corresponding to 4 (CUT, OAK), 8 (BFG, NAS, SNR), and 9 (PAL) years since planting. Additional information about these sites is presented in Table 1.
Statistical analyses
Initially, a single-site analysis was fitted considering a different heritability for each of the two silvicultural treatments (operational and intensive). Here, the objectives were to evaluate if there are statistical differences among treatment means based on an approximated t test and to determine the statistical significance of the genotype-by-silviculture interaction by using a likelihood ratio test (LRT, Gilmour et al. 2009), where the test is against the null hypothesis of the genetic correlation equaling 0. The fitted model corresponded to the following:
where y is the data vector (VOL or SURV); μ is the overall mean with the first column of ones; c is the fixed vector of silvicultural treatment effect; r is the random vector of replicate, with r ∼ MVN(0, \( {\sigma}_r^2{\boldsymbol{I}}_{\boldsymbol{r}} \)); i(r) is the random vector of incomplete-block-within-replicate effects, with i(r) ∼ MVN(0, \( {\sigma}_i^2{\boldsymbol{I}}_{\boldsymbol{i}} \)); g(c) is the random vector of nested effects of clone within culture effect, with g(c) ∼ MVN(0, I g ⊗ G c ); and e is the random vector of error effects, with e ∼ MVN(0, D e ). Here, 1 is a vector of ones, X and Z are incidence matrices of their corresponding effects, the matrix I x is an identity matrix of its proper dimension, G c is a 2 × 2 unstructured covariance matrix containing a different total genetic variance for each silvicultural treatment (\( {\sigma}_{c1}^2 \), \( {\sigma}_{c2}^2 \)), and a covariance between them (σ c1c2), and ⊗ denotes the Kronecker product. Also, D e is a diagonal matrix containing a different error variance for each of the two silvicultural treatment effects (i.e., \( {\sigma}_{e1}^2 \), \( {\sigma}_{e2}^2 \)). The fitting of the clonal effect as being nested within treatment was used to estimate separate clonal variances for each treatment and its correlation, and it does not imply that clones are confounded with treatments within sites.
A clonal genetic correlation between culture effects (\( {\hat{\rho}}_c^2 \)) and a broad-sense heritability was estimated for each site according to the following:
Note that the above expression corresponds to the mean within-treatment broad-sense heritability and should not be confused with the across-treatment heritability.
Later, both one- and two-stage analyses were performed using all information available to obtain the information required for a MET analysis. In both analyses, silvicultural treatment effects were ignored. The one-stage analysis was done by fitting an explicit and an implicit model. The one-stage explicit GxE model corresponds to the following:
where y is the data vector; μ is the overall mean; s is the fixed effect of sites; r(s) is the random effect of replicate within site, with r(s) ∼ MVN(0, D sr ); i(rs) is the random effect of incomplete blocks within a replicate-site combination, with i(rs) ∼ MVN(0, D si ); g is the random effect of the total genetic (i.e., clonal) effect, with g ∼ MVN(0, \( {\sigma}_g^2{\boldsymbol{I}}_{\boldsymbol{g}}\Big) \); gs is the random effect of genotype-by-site interaction, with gs ∼ MVN(0, \( {\sigma}_{gs}^2 \) I g ⊗ I s ); and e is the random residual error, with e ∼ MVN(0, D e ). Here, the matrices D x correspond to diagonal matrices with a different variance component for each of the sites for the corresponding term, and the other matrices were previously defined. Note that the above model estimates a single genetic and GxE variance across sites, and therefore a type B genetic correlation estimate \( \left({\hat{\rho}}_g^2\right) \) is estimated across all sites according to \( {\hat{\rho}}_g^2={\hat{\sigma}}_g^2/\left({\hat{\sigma}}_g^2+{\hat{\sigma}}_{gs}^2\right) \).
The expression considered to fit the one-stage implicit GxE model was as follows:
where all terms are identical to the ones presented in Eq. (2), except that gs now corresponds to a random effect of genotype nested within site, with gs ∼ MVN(0, I g ⊗ G s ). This factor gs corresponds to a nested effect as a result of not including the term g in the above model (as done in Eq. (1)), which is then contained within the gs model term. Here, G s is a 6 × 6 variance-covariance GxE matrix of the following form:
where diagonal terms correspond to the total genetic variances for a given site and off-diagonal terms correspond to estimated genetic covariances between any two sites. Several structures of the above variance-covariance matrix are possible, where that presented above is known as the unstructured (US) matrix and requires 21 (=6 + (6 × 5) / 2) components to be estimated, and it can also be parameterized as the heterogeneous general correlation (CORGH), which is defined in terms of correlations and not covariances (Gilmour et al. 2009).
Unique clonal-mean heritabilities (or repeatabilities) were estimated for each site \( \left({H}_{(i)}^2\right) \), due to the specification of individual-site residual variances, according to the formulae below:
where (i) identifies the i th site, k (i) is the effective mean number of ramets per clone at i th site, and all other terms were previously described.
For the two-stage MET analysis, two steps were done. The first step performed a single-site analysis where all the design features of each site are considered. The single-site model fitted corresponded to the following:
where all the model terms are identical to the ones presented in Eq. (1), with the difference that here g is now assumed to be a fixed effect of the total genetic (i.e., clonal) effects. For this step, the data were analyzed per site resulting in adjusted predicted least square means for each genotype in each environment. For the second step, these predicted means for the genotypes are used as responses to fit a simple weighted LMM of the following form:
where this model is similar to the implicit GxE model presented in Eq. (3), with y corresponding to the adjusted means for each site obtained from the single-site analysis Eq. (4), gs is the random effect of genotype nested within site, with gs ∼ MVN(0, I g ⊗ G s ), and e is the random residual errors, with e ∼ MVN(0, D s ). Here, G s is the 6 × 6 variance-covariance GxE matrix and D s corresponds to the diagonal matrix with a different fixed residual variance component obtained from the models fitted in the first step.
Three types of weights, πij for genotype i in site j, were considered for model fitting in the second step. The first weight involved the use of the inverse of the standard error of the predictions of SEij, as
The second weight, termed effective replication (ER), was proposed by Smith et al. (2001) and uses an estimated effective error variance across sites, \( {\tilde{\sigma}}_j^2 \) calculated as follows:
where r ij is the non-missing plot response for each genotype i in site j, n j is the number of non-missing genotypes on site j, and \( {\tilde{\sigma}}_j^{ii} \) is the standard error of the prediction of genotype i in site j. Hence, the weights are a function of varying level of expression of genetic variation scale and replication according with Eq. (7) as
where \( {r}_{ij}^{*} \) = \( {\tilde{\sigma}}_j^{ii}*{\tilde{\sigma}}_j^2 \); for an orthogonal analysis \( {r}_{ij}^{*} \) is the actual replication for each genotype, and for a non-orthogonal analysis, it is the effective replication.
Finally, the third type is an equal weight alternative (NONE) with π ij = 1 for all genotypes in all sites.
All previously described models were fitted using the software ASReml 3.0 (Gilmour et al. 2009) that estimates variance components using restricted maximum likelihood (REML) (Patterson and Thompson 1971).
In addition, for both of the models fitted in Eqs. (3) and (5), different variance structure forms of the matrix G s were evaluated. The unstructured (US), heterogeneous uniform correlation (CORUH), and factor analytic of order 1 (FA1) were evaluated for one- and two-stage analysis. These structures were compared and evaluated by contrasting log-likelihood (logL) values and information criteria (AIC and BIC). To compare the one-stage analysis against the two-stage analysis and one-stage against single-site analysis Pearson’s product-moment correlations (r PRED) and rank correlations (r RANK) were calculated for each site, using the predictions of the fitted one-stage implicit model using US structure as a baseline (Eq. (3)).
To evaluate the implications of GxE interaction on selection and to compare selection efficiency between one- and two-stage analysis and between one-stage and single-site analysis, clones were selected based on their overall (i.e., across all sites) genetic prediction value. This selection was contrasted with individual-site selections based on genetic predictions for each site. Selection intensities used were 2, 5, and 10 %, corresponding to the top 20, 50, and 100 genotypes, respectively.
Finally, to assist in the interpretation of the estimated G s matrix and to evaluate the relationship between sites and genotypes, a Biplot analysis was performed using the genetic correlation matrix of G s estimated from fitting the model from Eq. (3) (i.e., one-stage analysis) with the US matrix together with the clonal predictions for each of the sites. This was done for both of the response variables VOL and SURV using the statistical package GenStat 17th (Payne et al. 2011).
Results
The phenotypic means and its standard deviations for VOL and SURV for each treatment together with total genetic correlation (i.e., clonal) for these two silvicultural treatments and its p value are presented in Table 2. CUT and SNR are the two sites with only one silvicultural treatment. The phenotypic means of each silvicultural treatment do not differ considerably for either VOL or SURV with the exception of NAS for VOL, where the intensive level was 29 % higher than the operational. For SURV, almost no differences between the two silvicultural treatment means were found. The results from fitting the single-site models (Eq. (1)) show high \( {\hat{H}}^2 \) values for VOL on all sites with an average of 0.31 (Table 2). Here, the lowest value was found on site OAK (0.19) and the highest on SNR (0.46). For SURV, all sites presented low heritability values (average of 0.04) particularly site PAL with almost null genetic control. The genetic correlation estimates between silvicultural treatments were high for VOL and SURV across most of the sites, with averages of 0.95 and 0.65, respectively. The only exception was site PAL for SURV, which had a value of −0.28 and, as indicated earlier, also showed low \( {\hat{H}}^2 \). Note that statistically, inferences on the different silvilcultural treatments are limited given that these were not replicated and that the expressions used for broad-sense heritability includes the estimated variance of replicate within site, which in all cases resulted of low magnitude.
The results of the explicit model analysis (Eq. (2)) provided interesting type B genetic correlation estimates of 0.63 and 0.57 for VOL and SURV, respectively (details not shown), reflecting moderate levels of GxE interaction. Estimates of type B genetic correlations between any pair of sites were obtained simultaneously from fitting the implicit model from Eq. (3) for VOL using unstructured (US) variance and covariance matrix. These values range from 0.41 (OAK-BFG) to 0.80 (PAL-NAS) (average of 0.57, Table 3). The site with the largest interactions (i.e., lowest genetic correlations) with any other site was OAK. As seen in Table 3, the mean clonal heritability values are high for all sites (0.66 to 0.81). The same model Eq. (3) but using factor analytic 1 (FA1) showed smaller type B genetic correlation estimates than US (Table 4), the values ranging from 0.36 (OAK-CUT) to 0.80 (PAL-NAS, SNR-NAS) (average of 0.56). However, the heritability values for FA1 are almost identical to the ones estimated under an US structure (Table 3). Model comparisons indicated, as expected, better results for US based on the logL, AIC, and BIC, where for the two-stage analysis using an implicit model with US structure the logL was −9337 and for FA1 structure this changed to −9362. Also, the AIC values were −18,675 (US) and 18,725 (FA1), where the implicit model using US was significantly different than the one using FA1.
Estimated genetic correlations for VOL obtained from the two-stage analysis Eq. (5) using US for all weights are shown in Table 4. The genetic correlation estimates for ER and NONE weights were smaller than the ones obtained for the one-stage analysis (see Table 3); however, 1/SE2 weights presented the least reduction. For ER weights, the genetic correlation estimates ranged from 0.24 (PAL-OAK) to 0.60 (PAL-NAS) (average of 0.36). However, when the 1/SE2 was used as weights, the correlations were apparently overestimated, the values being higher than those found for one-stage analysis (Table 3). The estimated values ranged from 0.49 (SNR-BFG) to 0.92 (PAL-NAS) (average of 0.67).
Higher estimated values of heritability were found for one-stage when compared to the two-stage analyses, reflecting a bias in the latter estimates. For one-stage analysis, similar heritability estimates were found for US and FA variance. The heritability estimates for these analyses ranged from 0.66 to 0.81, the lowest being found for OAK and the highest for SNR. However, for the two-stage analysis, the heritability estimates were different according with the methods of weighting. Higher estimates were found for ER (0.49 to 0.74) and NONE (0.40 to 0.73). The method 1/(SE)2 showed lower heritability values, ranging from 0.28 to 0.70. OAK was the site with lowest heritability for all methods.
To compare the performance between the one- and two-stage analyses, Pearson product-moment correlations (r PRED) and rank correlations (r RANK) were obtained for two-stage analyses (Table 5). Even though heritability values and type B genetic correlation estimates showed apparent bias, the r PRED and r RANK values between one- and two-stage predictions were high when all the genotypes were selected. The best r PRED and r RANK were found for the weight 1/SE2. Surprisingly, given the moderate levels of GxE interaction present for these sites, high correlation estimates were found when comparing one-stage with single-site analysis. When selecting either the top 20, 50, and 100 clones, the rank correlation was lower for any weights compared with the one-stage analysis, with the lowest values for ER and NONE weights.
Genetic gains from clonal selection at different selection intensities provided interesting results for VOL across single-site, one-stage, and two-stage analyses (Table 6). For single site analysis, the estimated gain was 40.81 % when 2 % of the genotypes were considered; however, when the datasets were combined into a one-stage analysis, gain increased to 50.24 %. For the two-stage analysis, the estimated gain was reduced due to the presence of bias for heritability and genetic correlations resulting in underestimation of genetic gain (Fig. 1).
The graphical output from the Biplot analyses using the type B correlation matrix from the one-stage analysis with the implicit model (Eq. (3)) in a two-dimensional space for VOL and SURV is shown in Fig. 2a, b. These Biplots are presented with only the two main principal components which explained a total of 86.71 % of the variation for VOL (76.18 and 10.53 % for the first and second components, respectively) and 93.03 % for SURV (79.47 and 13.59 %, respectively). In Fig. 2, the lines represent the sites emerging from the origin, and the angle between two lines correspond to the type B correlation between two given sites. Also, individual genotypes are represented by gray circles. For VOL, all environments present positive correlations (see also Table 3) where BFG and CUT and SNR and PAL are grouped together, showing similarity in response for these sites (Fig. 2a). There are a few genotypes that were consistently outstanding in all sites (see circles on the right of panel from Fig. 2a). For SURV, most sites were highly correlated, with the exception of OAK that was uncorrelated (angle ∼90°) to all other sites. For this trait, the genotypes appear more clustered than for VOL, with a few genotypes that were consistently outstanding on all sites (Fig. 2b).

Biplot from the principal componet analysis of all clones and sites for: a volume (dm3) and b survival (%)
Discussion
The total genetic correlation estimates (i.e., clonal) between the different silvicultural treatments (Table 2) were high for VOL and SURV across most of the sites, with the exception of PAL for SURV. The approximated t tests that evaluated the mean differences between the silvicultural treatments gave significant results only for sites NAS and PAL on VOL, note that their stem volume for seedlings was larger than for cuttings. These results are expected as there are important consequences in tree size and survival in response to different silviculture treatments implemented. From this analysis, it is clear that clonal rankings do not differ significantly between the two silviculture treatments, but there are differences in the magnitude of the genetic effects.
The analyses performed in this study explored different aspects of the genetic structure of loblolly pine clones planted in the Southeast USA that are relevant for current and future breeding strategies. A wide range of values for clonal-mean repeatability in the single-site analysis was found for VOL as reported for other breeding programs (Balocchi et al. 1993). The clonal-mean heritability estimates were relatively high for SNR, and low for OAK, which could be product of the varied ages and environment effects that influenced the level of expression of the evaluated clones (Owino 1977).
According with Burdon (1977), one way to estimate and understand GxE interaction is through the genetic correlation (type B). This correlation has advantages for characterizing the roles of environments in generating GxE, and thereby helps breeding programs to establish the best strategies. The explicit GxE model provided a single type B correlation estimate across all sites and reflects a moderate to high level of GxE interaction for both VOL and SURV. Particularly, for SURV, the type B correlation was 0.57, a relatively low estimate that indicates limited consistency of rankings among sites.
Considering the implicit GxE model with US variance structure, the different levels of heritability indicate how the same trait is expressed in different environments (Table 3). The different type B correlation estimates also indicate the level of agreement on genetic values, and indirectly on ranking, between the sites. OAK was the site with the lowest genetic correlations and therefore highest GxE interaction with respect to the others. This could be due to the young age (4 years) of the trees at this site, as younger trees may not fully express their genetic merit and hence, and thereby tend to reflect environment effects more than older trees (Balocchi et al. 1993). In contrast, NAS was the site with the lowest levels of GxE interaction. Similar results have been supported by Lucero et al. (2003), where they found troublesome genotype-by-environment interaction for VOL for some combinations of sites.
Other MET studies using FA models for plant breeding have shown substantial superiority, in terms of goodness-of-fit to model variance components, when compared with simpler structures (Smith et al. 2015). Piepho and van Eeuwijk (2002) commented about computational difficulty in fitting multiple variance-covariance structures to MET. However, the FA structure showed computational advantages when the study has a large number of sites. Ogut et al. (2014) also recommend using multiplicative models with extended FA models for forest tree MET data to address the heterogeneity for more complex genetic variance-covariance structures.
The results from the two-stage analysis showed lower genetic correlations than the one-stage analysis for all weighting methods used, which is to be expected, as with the two-stage analysis, some information is lost; however, the magnitude of this loss depended on the weighting method used. According to Mohring and Phiepo (2009), the best weighting method depends more on the dataset than the evaluation criteria. The heritability values for two-stage analyses were lower than one-stage, which Ogut et al. (2014) also found when comparing heritability values between one- and two-stage analyses.
The higher Pearson product–moment correlation found for predictions and rankings can be explained due to the similar performance of the genotypes across all sites, showing lower GxE interactions between one- and two-stage analyses (Mohring and Phiepo 2009). The high correlation found for the NONE method (unweighted) for predictions and rank indicates that unweighted method showed reasonable results and the differences among weighting schemes were smaller when larger numbers of genotypes are selected. When the number of genotypes were reduced to 100, 50, and then 20, the differences among weights are more pronounced and correlation between predictions and ranks are reduced, as would be expected with increasingly truncated distributions.
For the Biplot analyses, genotypes that are physically closer to each other represent similar genetic responses across all environments (Fig. 2). For VOL, the trials BFG and CUT and SNR and PAL were grouped together and showed a similar response trend. The perpendicular projection of the genotype onto a given environment axis shows the performance of that genotype in that environment (or site) (Boer et al. 2007). For OAK, the trait SURV presented almost no correlation with all other sites (Fig. 2b); this is probably due to site conditions that produced the lowest average survival of all trials (90.6 %, see Table 1). Note also that several genotypes had low SURV values, but the majority have high SURV predictions and are located to on the right side of the panel (see Fig. 2b).
Conclusions
Relevant levels of genotype-by-environment interaction for individual-tree volume across the six sites studied were detected. Some sites present higher levels of interactions than could be explained by their ages, geographical location, site preparation, and understory competition, among other factors.
The one-stage analysis unstructured model resulted in the best analysis of genotype-by-environment interaction for these six sites. Nevertheless, for more complex multi-environmental trial analyses (e.g., >10 sites), a two-stage analysis is recommended, with an unstructured or factor analytic structure, due to its minimum loss of information and its computational advantages, as found in this study. Factor analytic structure seems more appropriate when there is a large number of sites with diverse levels of genotype-by-environment interaction.
The one- and two-stage analyses gave different type B correlation estimates, meaning that at least one incurred bias; nevertheless, the correlation for rankings and breeding values was similar. In addition, discrepancies in the genetic correlation and heritability estimates were found among all three weighting methods, which need to be taken into consideration. The weights based on the inverse of the variance of the predictions, 1/(SE)2, showed considerably better results than the other weightings with minimum discrepancies in variance-component estimates for the dataset analyzed in this study.
The use of the Biplots originating from the PCA was a useful tool to summarize genotype-by-environment and confirmed the positive correlation between all the environments detected. Also, they provide different insight on selection of genotypes, which can be complemented with stability indexes (e.g., superiority measure), which will yield better selections and higher genetic gains for breeding programs.
This present study provided an opportunity to explore both interactions between genotypes and cultural treatments and interactions between genotypes and sites for loblolly pine in the Southeastern USA. The statistical tools presented here can be easily applied to other analysis for complex tree breeding programs that evaluate a large series of trials. Here, it is recommended to implement one-stage analyses whenever possible with the use of factor analytic variance-covariance structures; however, the use of a two-stage analysis with weightings based on the inverse of the standard error of the prediction will lead to similar results, allowing one to evaluate large and messy multi-environmental datasets.
References
Balocchi CE, Bridgwater FE, Bryant R (1993) Age trends in genetic parameters for tree height in a nonselected population of loblolly pine. For Sci 39:231–251
Baltunis BS, Huber DA, White TL, Goldfarb B, Stelzer HE (2007) Genetic analysis of early field growth of loblolly pine clones and seedlings from the same full-sib families. Can J For Res 37:195–205
Boer MP, Wright D, Feng L, Podlich DW, Luo L, Cooper M, van Eeuwijk FA (2007) A mixed-model quantitative trait-loci (QTL) analysis for multiple-environment trial data using environment covariables for QTL-by-environment interactions, with an example in maize. Genetics 177:1801–1813
Burdon RD (1977) Genetic correlation as a concept for studying genotype-environment interaction in forest tree breeding. Silvae Genet 26:168–175
Comstock RW, Moll RH (1963) Genotype-environment interactions. In Statistical Genetics and Plant Breeding, NAS-NRC 982:164–196
Costa e Silva J, Potts BM, Dutkowski GW (2006) Genotype by environment interaction for growth of Eucalyptus globulus in Australia. Tree Genet Genomes 2:61–75
Cullis BR, Jefferson P, Thompson R, Smith AB (2014) Factor analytic and reduced animal models for the investigation of additive genotype-by-environment interaction in outcrossing plan species with application to a Pinus radiata breeding programme. Theor Appl Genet 127:2193–2210
Cullis BR, Smith AB, Coombes N (2006) On the design of early generation variety trials with correlated data. J Agric Biol Environ Stat 11:381–393
Cullis BR, Thompson FM, Fisher JA, Gilmour AR, Thompson R (1996) The analysis of the NSW wheat variety database. II. Variance component estimation. Theor Appl Genet 92:28–39
Freeman GH (1973) Statistical methods for the analysis of genotype-environment interactions. Heredity 31:339–354
Frensham A, Cullis B, Verbyla A (1997) Genotype by environment heterogeneity in a two-stage analysis. Biometrics 53:1373–1383
Gabriel KR (1971) The biplot graphic display of matrices with application to principal components analysis. Biometrika 58(3):453–467
Gilmour AR, Gogel BJ, Cullis BR, and Thompson R (2009) ASReml User Guide v. 3.0. VSN International Ltd, Hamel Hempstead, 398 pp.
Hardner CM, Dieters M, Dale G, DeLacy I, Basford KE (2009) Patterns of genoype-by-environment interaction in diameter at breast height at age 3 for eucalypt hybrid clones grown for reafforestation of lands affected by salinity. Tree Genet Genomes 6:833–851
Kolher U, Luniak M (2005) Data inspection using Biplot. Stata J 5:208–223
Lucero VS, Huber DA, McKeand SE, White TL, Rockwood DL (2003) Genotype-by-environment interaction and deployment considerations for family from Florida provenances of loblolly pine. For Genet 10:85–92
Malosetti M, Ribaut JM, Eeuwijk FA (2013) The statistical analysis of multi-environment data modeling genotype-by-environment interaction and its genetic basis. Frontiers in Physiology – Plant Physiol 4:1–17
Meyer K (2009) Factor-analytic models for genotype x environment type problems and structured covariance matrices. Genet Sel Evol 41:21–32
Muir W, Nyquist WE, Xu S (1992) Alternative partitioning of the genotype-by-environment interaction. Theor Appl Genet 84:193–200
Mohring J, Piepho HP (2009) Comparison of weighting in two-stage analysis of plant breeding trials. Crop Sci 49:1977–1988
Ogut F, Maltecca C, Whetten R, McKeand S, Isik F (2014) Genetic analysis of diallel progeny test data using factor analytic linear mixed models. For Sci 60:119–127
Owino F (1977) Genotype x environment interaction and genotype stability in loblolly pine. Silvae Genet 4:131–134
Patterson HD (1978) Routine least squares estimation of variety means in incomplete tables. J Nat Inst of Agric Botany 13:142–151
Patterson HD, Thompson R (1971) Recovery of inter-block information when block sizes are unequal. Biometrika 58:545–554
Payne RW, Murray DA, Harding SA, Baird DB, Soutar DM (2011) An introduction to GenStat for WINDOWS, 17th edn. VSN International, Hemel Hempstead
Piepho HP, van Eeuwijk F (2002) Stability analysis of crop performance evaluation, Chap 11. Haworth Press, New York, pp. 315–351
Piepho HP, Mohring J, Schulz-Streeck T, Ogutu J (2012) A stage-wise approach for the analysis of multi-environment trials. Biometrical J 54:844–860
Silva GAP, Gezan SA, Carvalho MP, Gouvea LRL, Verardi CK, Oliveira ALB, Goncalves PS (2014) Genetic parameters in a rubber tree population: heritabilities, genotype-by-environment interactions and multi-trait correlations. Tree Genet Genomes 10:1511–1518
Smith AB, Cullis BR, Thompson R (2001) Analyzing variety by environment data using multiplicative mixed models and adjustments for spatial field trend. Biometrics 57:1138–1147
Smith AB, Ganesalingam A, Kuchel H, Cullis BR (2015) Factor analytic mixed models for the provision of grower information from national crop variety testing programs. Theor Appl Genet 128:55–72
Thompson R, Cullis BR, Smith AB, Gilmour AR (2003) A sparse implementation of the average information algorithm for factor analytic and reduced rank variance models. Austr New Zeal J Stat 45:445–459
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R. Burdon
Data Archiving Statement.
We followed standard Tree Genetics and Genomes policy. Data used in this study came from the Forest Biology Research Cooperative (FBRC) from the University of Florida. In addition, supplementary information of original genotypes numbers/names and data are included in the Supplementary Files
Electronic supplementary material
ESM 1
(XLSX 3096 kb)
Rights and permissions
About this article

Cite this article
Gezan, S.A., de Carvalho, M.P. & Sherrill, J. Statistical methods to explore genotype-by-environment interaction for loblolly pine clonal trials. Tree Genetics & Genomes 13, 1 (2017). https://doi.org/10.1007/s11295-016-1081-0
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11295-016-1081-0