
Abstract
Gibberellins (GAs) have been widely used for many years to induce seedless grapevine and increase fruit set in production. However, the role of GAs and how they crosstalk with auxin and cytokinin during fruit set in grapevine remains unclarified. To investigate their role, GA3, 4-chlorophenoxyacetic acid (4-CPA), 6-benzylaminopurine (6-BA), and different hormone combinations were applied to unpollinated ovaries of the ‘Fenghou’ grape (Vitis vinifera × Vitis labrusca) at anthesis. The results showed that the application of 4-CPA, 6-BA, GA3, 4-CPA+GA3, 6-BA+GA3, and 4-CPA+6-BA induced parthenocarpy. Among them, 4-CPA-, 6-BA-, and 4-CPA+6-BA-induced fruits were smaller than those induced by GA3, 4-CPA+GA3, and 6-BA+GA3. Application of the GA biosynthesis inhibitor paclobutrazol reduced both the fresh weight and fruit set of 4-CPA- and 6-BA-induced fruits, but the effect of the inhibitor was reversed by the application of GA3. In contrast to unpollinated ovaries, the ovaries induced by 4-CPA and 6-BA showed enhanced accumulation of active GAs, but at a lower level than in pollinated ovaries between 7 and 21 days after anthesis (DAA). Gene analysis showed that the increase in the GA content in 4-CPA- and 6-BA-treated ovaries was the result of the upregulation of GA biosynthesis genes, such as VvGA3ox1, and suppression of GA catabolism genes, such as VvGA2ox3 and VvGA2ox4. In addition, the concentration of active IAA was enhanced in the ovaries induced by 6-BA, which was accompanied by the elevated expression of the IAA biosynthesis genes VvYUC2 and VvYUC6. These results suggested that 4-CPA- and 6-BA-induced fruit set in grapevine requires downstream gibberellin biosynthesis.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Fruit set (defined as the changeover from the ovary in the mature flower to functional fruit) is not only very important for plant reproduction because of its close relationship to the normal development of seeds but also determines the fruit quality and yield potential (Gillaspy et al. 1993). In general, the initiation of fruit formation depends on the successful completion of pollination and fertilization because signals derived from pollination and young embryos are required for fruit initiation and early fruit development (Wang et al. 2009). The signals that trigger fruit set and growth after pollination and fertilization are considered to be phytohormones (Vivian-Smith et al. 2001), and seeds are the sources of these phytohormones (Archbold and Dennis 1985; Picken 1984). Moreover, in practice, plant growth regulators are used to improve fruit set under suboptimal growth conditions, which limits pollination and fertilization in the production of horticultural crops (Fos et al. 2001; Gillaspy et al. 1993; Martinelli et al. 2009). In addition, unpollinated pistils can develop into seedless fruit by the exogenous application of various plant growth regulators, which is known as artificial parthenocarpy (Gorguet et al. 2005; Serrani et al. 2007a; Vivian-Smith and Koltunow 1999). An increase of the endogenous hormone levels has also been observed in parthenocarpic fruits during the stage of fruit set (Aloni et al. 2006; Talon et al. 1992), suggesting that the stage of fruit formation is controlled by plant hormones.
Auxin and GAs are the key regulators in the early development stage of fruits following fertilization (Dorcey et al. 2009; Hu et al. 2008; Ozga and Reinecke 2003). This view is supported by the fact that the exogenous application or transgenic elevation of these two hormones leads to the development of parthenocarpic fruit in the absence of fertilization (Costantini et al. 2007; Gorguet et al. 2005; Lu et al. 1997). Recently, a synergistic effect of auxin and gibberellin on fruit set has been observed in model plants. The crosstalk of these two plant hormones was thought to be necessary in regulating fruit formation and growth (Ozga and Reinecke 1999; Serrani et al. 2008). Auxin produced in the fertilized ovule induces the biosynthesis of downstream gibberellin (Dorcey et al. 2009; Gallego-Giraldo et al. 2014; Pattison and Catalá 2012), which relieves the restraint for the ovary and promotes fruit development (Fuentes et al. 2012; Martí et al. 2007). On the other hand, these two hormones have different effects on the morphological and histological development of fruit tissues in tomato (Serrani et al. 2007a). Auxin-induced fruits have greater cell numbers, while fruits induced by gibberellin have larger mesocarp cells. However, the simultaneous application of both of these hormones induces parthenocarpic fruits that are similar to pollinated fruits (Ruan et al. 2012). In grapevine, auxin and gibberellin could also be a substitute for pollination and fertilization because they can increase the number and size of the berries per bunch (Lavee 1960). Unpollinated grape ovaries, which were treated with auxin or gibberellin at anthesis, could develop parthenocarpically (Weaver and Mccune 1960). In addition, auxin is present at a high level in the ovary at anthesis and fruit set, while the concentration of gibberellin increases within the cell enlargement period (Böttcher et al. 2010; Pérez et al. 2000). In agreement with this finding, there is an increase in the transcripts of auxin and gibberellin biosynthesis genes in the grape pistil following pollination and fertilization (Kühn and Arce-Johnson 2012), suggesting that both auxin and gibberellin are required for fruit set in grapevine. Although the mechanism of auxin and gibberellin underlying fruit set and growth has been well clarified in model plants, this mechanism is still not clear in grapevine.
In addition to GAs and auxin, cytokinin has also been shown to play an essential role in fruit set and growth (Mapelli 1981; Weaver et al. 1962). In tomato, the cytokinin biosynthesis genes in the ovary were induced by pollination and fertilization and associated with an increase of the cytokinin content (Mariotti et al. 2011). Cytokinin application may induce parthenocarpy in tomato partially through a modulation of GAs and IAA metabolism (Ding et al. 2013). Similarly, the concentration of cytokinins are also high at anthesis and fruit set in grapevine, and the size of seedless berries can be increased by applying cytokinins at the cell enlargement period (Zhang et al. 2003). Generally, cytokinin is thought to participate in stimulating cell division during fruit development (Dauelsberg et al. 2011). In fact, the mechanism of cytokinin action during the set stage and its relationship with other hormones is still unknown in the grapevine.
The application of these hormones before or after flowering has been widely used to improve the fruit quality in grapevine (Coombe 1960; Lu et al. 1997), even though the molecular mechanisms under these processes are mostly unknown. Moreover, grapevine is a woody perennial plant with fruits having particularities that differentiate them from those of commonly studied organisms. To improve our understanding of the hormonal control of fruit set and development in grapevine, it is necessary to determine the role of these hormones and how they crosstalk with each other during grape fruit set. In this work, we investigated the effect of the application of GA, auxin, and cytokinin on the parthenocarpic development of unpollinated ovaries. Next, we investigated whether the induction of fruit set and early fruit development by auxin and cytokinin is mediated by GAs using paclobutrazol (PAC), a GA biosynthesis inhibitor. In addition, we also conducted an analysis of the contents of active GAs and of GA metabolism-related transcripts in the grape ovary and tested the relationship between auxin and cytokinin.
Materials and methods
Plant materials
The grape cultivar ‘Fenghou’ (Vitis vinifera × Vitis labrusca) was grown in a Beijing hot spring nursery. In 2013–2014, 146 inflorescences at the same junction of grapevine branches from 30 plants were emasculated and bagged 10 days before anthesis (DBA) to avoid self-pollination (Supplemental Fig. S1). Ovaries from each inflorescence were chosen from the first 1/3 segments at 1, 4, 7, 14, and 21 days after anthesis (DAA) for microscopy, hormone content, and gene expression analysis.
Hormone treatments and sampling
On the day equivalent to anthesis, unpollinated ovaries were sprayed with different hormone combinations, namely, 4-chlorophenoxyacetic acid (4-CPA, 15 ppm), 6-benzylaminopurine (6-BA, 500 ppm), gibberellic acid (GA3, 5 ppm), 4-CPA (15 ppm) + GA3 (5 ppm), 6-BA (500 ppm) + GA3 (5 ppm), and 4-CPA (15 ppm) + 6-BA (500 ppm). The hormone concentrations used in this work were based on a previous study (Weaver and McCune 1960; Weaver et al. 1965), with some modifications. At the same time, PAC (50 ppm), a GA biosynthesis inhibitor, was applied to inhibit GA biosynthesis in pollinated and 4-CPA- and 6-BA-treated ovaries. Up to 30 inflorescences were used for 4-CPA or 6-BA treatment, and 8 inflorescences were used for every remaining treatment. PAC, GA3, 4-CPA, and 6-BA (Sigma-Aldrich, USA) were dissolved in a solution containing 5 % ethanol and 0.1 % Tween 80. Unpollinated ovaries that were treated with solvent alone were used as the controls.
Fruit morphological structure observations and statistical analysis
The ovaries or fruits sampled at 4, 7, 14, and 21 DAA were fixed in FAA (50 % ethanol, 5 % glacial acetic acid, and 5 % formaldehyde) and then passed through an ethanol series for tissue dehydration. These samples were then embedded in paraffin and cut into 8-μm sections, which were stained with Safranin and Fast Green and photographed using an Olympus CX31 microscope. The cell diameters and pericarp thickness of the ovaries from different treatment groups at 21 DAA were measured using ImageJ software.
Determination of the plant hormone content
The contents of GA1 + 3, GA4 + 7, and IAA were analyzed by using GA1 + 3, GA4 + 7, and IAA ELISA kits (China Agricultural University, Beijing, China). Up to 0.5 g of ovaries or fruits collected from different development stages was used for the extraction, purification, and immunoassay, as described previously (Teng et al. 2006).
RNA extraction and real-time PCR analysis
Total RNA was extracted from the ovaries/fruits in the grapevine using the revised CTAB method (Murray and Thompson 1980). After the products were purified with RNase-free DNase I (Takara, Japan), the RNA concentration was determined using an atom UV spectrophotometer (OD260/280 = 1.8–2.0). Up to 2 μg of total RNA was used to synthesize the first-strand cDNA using reverse transcriptase (Promega,USA) and an oligo dT primer. The reaction conditions for RT were as follows: the samples were incubated at 70 °C for 5 min, followed by 42 °C for 1 h, and then immediately placed on ice. These cDNAs were used as templates for quantitative real-time PCR (qRT-PCR), which was conducted on an ABIPRISM 7500. A 20-μl total reaction volume was used, including 2 μl of sample cDNA (total RNA, 0.025 μg), 10 μl of 2× Bestar SybrGreen qPCR Mastermix (DBI, Germany), 0.8 μl of primers, 0.4 μl of 50× ROX Reference Dye, and 6.8 μl of double-distilled water. All experiments were performed with three biological replicates and two technical replicates. The primers that were used in previous studies (Giacomelli et al. 2013; Jung et al. 2014) were used for the GA biosynthesis genes (VvGA20ox1 [gibberellin 20-oxidase-1], VvGA20ox2 [gibberellin 20-oxidase-2], VvGA20ox3 [gibberellin 20-oxidase-3], VvGA3ox1 [3b-hydroxylase (3OH-1)], VvGA3ox2 [3b-hydroxylase (3OH-1)], VvGA3ox3 [3b-hydroxylase (3OH-1)]), GA catabolism genes (VvGA2ox1 [gibberellin 2-oxidase-1], VvGA2ox2 [gibberellin 2-oxidase-2], VvGA2ox3 [gibberellin 2-oxidase-3], VvGA2ox4 [gibberellin 2-oxidase-4], and VvGA2ox7 [gibberellin 2-oxidase-7]), and the auxin biosynthesis genes (VvYUC2 and VvYUC6). VvActin1 was used as an endogenous control for the normalization of gene expression. All of these primers are listed in Supplemental Table S1.
Statistical analysis
One-way analysis of variance and Tukey’s test were conducted to assess statistical significance among different treatments. Significant differences between the means were based on the least significant method when the F values were significant (p < 0.05).
Results
Gibberellin, auxin, and cytokinin induce parthenocarpy and early fruit development in grapevine
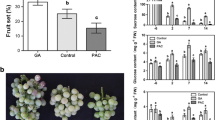
To determine whether gibberellin (GA), auxin, and cytokinin can induce parthenocarpy in grapevine, 4-CPA, 6-BA, GA3, 4-CPA+GA3, 6-BA+GA3, and 4-CPA+6-BA were applied to unpollinated ovaries at anthesis. The unpollinated ovaries that were treated with solvent solution (control) did not grow (3.63 mm at 14 DAA compared to approximately 1 mm at 0 DAA) and had already abscised at 21 DAA, while the longitudinal diameter of the pollinated ovaries was 16.04 mm at 14 DAA (Fig. 1a). The application of GA3, 4-CPA, or 6-BA alone was able to induce parthenocarpy and fruit growth. Among them, the fruit size of GA3-induced fruits was larger than that of 4-CPA- and 6-BA-induced fruits. Two hormone combinations were also used in our work; the results show that the fruit size of the 4-CPA+6-BA treatment was smaller than that induced by 4-CPA+GA3 and 6-BA+GA3 and was similar to that of the fruits treated with 4-CPA and 6-BA alone. However, the fruit size of the 4-CPA+GA3-, 6-BA+GA3-, and GA3-treated fruits was comparable to that of the pollinated fruits (Fig. 1b, c). There was no difference in fruit set among the different treatments (Fig. 1d). In addition, the internal morphology of GA3 and 4-CPA fruits was quite different. Ovules had limited growth and degenerated in GA3-induced fruits, whereas the development of pseudoembryos was observed in 4-CPA-induced fruits (Fig. 1b).

Responses of unpollinated ovaries to different hormones. Pollinated and unpollinated ovaries at 14 DAA (a). Effects of different hormones on the development of unpollinated fruits at 21 DAA (b). Responses of fruit weight (c) and fruit set (d) to different hormones. e Microscopic analysis of unpollinated ovaries treated with different hormones at 21 DAA. Scale bar, 100 μm. P pollination. Values of fruit set percentage are the means of eight inflorescences (±SE) and fruit weight represents the mean ± SE of three biological replicates with 10 fruits as a replicate. Significant differences (p < 0.05) between treatments are indicated using different letters according to Tukey’s test
Due to the difference in the size among different hormone-treated fruits, we compared the microstructure of these fruits at 21 DAA. The results showed that there were no significant differences among all of the treatments in the cell layers and the diameters of the exocarp and endocarp. Nevertheless, the mesocarp had comparable cell layers in the 4-CPA- (23.2 ± 0.4), 6-BA- (23.8 ± 1.6), and 4-CPA+6-BA (23.8 ± 1.3)-induced fruits compared to the pollinated fruits (26.0 ± 0.7) and significantly more cell layers than the fruits treated with GA3 (16.8 ± 0.8). In the case of the mesocarp cell diameter, the GA3-induced fruits (146.2 ± 13.1 μm) were larger than the fruits obtained by pollination (126.3 ± 12.5 μm). However, the mesocarp cell diameter in the 4-CPA- (72.0 ± 18.9 μm), 6-BA- (72.6 ± 12.9 μm), and 4-CPA+6-BA (75.0 ± 13.3 μm)-induced fruits was much smaller than that in the GA3 treatment. In addition, the fruits that were treated simultaneously with GA3 and 4-CPA or 6-BA had similar mesocarp cell layers and diameters compared to the pollinated fruits (Fig. 1e and Table 1).
PAC reduces auxin- and cytokinin-induced parthenocarpy and fruit growth
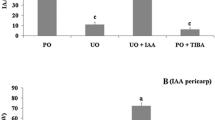
To investigate the relationship between GA and auxin or cytokinin in grapevine, PAC, a GA biosynthesis inhibitor, was applied to pollinated and unpollinated ovaries with 4-CPA or 6-BA. The application of PAC totally inhibited the fruit set of pollinated ovaries; however, grape fruit set was restored by the co-application of GA3 (Fig. 2). In unpollinated ovaries treated with 4-CPA and 6-BA, PAC also significantly reduced the fresh weight (Fig. 2a) and berry set percentage (Fig. 2b), but this inhibition was reversed by the co-application of GA3. These results showed that 4-CPA- and 6-BA-induced parthenocarpy and fruit growth requires the involvement of GA3, which may act downstream of auxin and cytokinin during grape fruit set.

Effect of PAC on parthenocarpic fruit induction by 4-CPA and 6-BA. PAC inhibited the growth (a) and fruit set (b) of the 4-CPA- and 6-BA-induced fruits, while the effect was reversed by GA3 (a and b). Values of fruit set percentage are the means of eight inflorescences (±SE) and fruit weight represents the means ± SE of three biological replicates with 10 fruits as a replicate. Significant differences (p < 0.05) between treatments are indicated using different letters according to Tukey’s test
High levels of bioactive gibberellin accumulated in the parthenocarpy induced by 4-CPA and 6-BA
To further analyze the role of GAs in the 4-CPA- and 6-BA-induced parthenocarpy, the content of bioactive gibberellin was quantified in pollinated and unpollinated ovaries treated with auxin and cytokinin or not (as a control) at 1, 4, 7, 14, and 21 DAA. Non-treated unpollinated ovaries contained lower levels of GA1 + 3 and GA4 + 7 than the ovaries induced by pollination, 4-CPA, and 6-BA. On the contrary, the concentrations of two active GAs in pollinated ovaries increased at 7 DAA and peaked at 14 DAA. Ovaries treated with 4-CPA and 6-BA also showed high contents of GA1+3 and GA4+7, but the contents were lower than those in pollinated ovaries between 7 and 21 DAA (Fig. 3a, b). These findings suggested that increased endogenous bioactive GA content plays an important role in inducing parthenocarpic fruits via 4-CPA and 6-BA.

GA1 + 3 (a) and GA4 + 7 (b) contents of pollinated and 4-CPA- and 6-BA-induced ovaries after anthesis. P pollination. Values are the mean ± SE of three biological replicates. Significant differences (p < 0.05) between treatments are indicated using different letters according to Tukey’s test
Auxin and cytokinin increase the bioactive GA content by regulating the expression of GA metabolism genes
To determine whether the high content of bioactive GA that accumulated in auxin- and cytokinin-induced fruits was the result of an alteration of the transcriptional regulation of GA metabolism genes, the transcripts of GA biosynthesis genes (VvGA20ox1, 2, and 3 and VvGA3ox1, 2, and 3) were analyzed by qRT-PCR.
The expression levels of VvGA20ox1, VvGA20ox2, and VvGA20ox3 in 4-CPA- and 6-BA-treated ovaries were higher than those in unpollinated ovaries between 1 and 14 DAA and were even higher than those in pollinated ovaries at the same time (Fig. 4a–c).

Changes in the expression of GA biosynthesis genes in different treatments between 1 and 21 DAA. VvGA20ox1, VvGA20ox2, and VvGA20ox3 (a–c). VvGA3ox1, VvGA3ox2, and VvGA3ox3 (d–f). Values are the mean ± SE of three biological replicates. Significant differences (p < 0.05) between treatments are indicated using different letters according to Tukey’s test
The expression of VvGA3ox1 was much higher in pollinated 4-CPA- and 6-BA-treated ovaries than in unpollinated ovaries at 7 DAA. However, 4-CPA- and 6-BA-induced fruits had a lower transcript level compared to pollinated fruits at 7 DAA and then quickly declined during 14–21 DAA. Expression of the VvGA3ox2 gene decreased from 1 to 21 DAA in all treatments after anthesis. In the case of VvGA3ox3, an increased transcript level was detected in pollinated and 4-CPA-induced fruits at 21 DAA (Fig. 4d–f).
Next, the expression of GA catabolism genes (VvGA2ox1, 2, 3, 4, and 7) was analyzed. Generally, the transcript content of all GA2ox genes in unpollinated ovaries remained high after anthesis (Fig. 5). The VvGA2ox3 and VvGA2ox4 transcript levels were significantly elevated at 7 DAA (Fig. 5c, d). In contrast, the low expression of VvGA2ox genes was observed in pollinated 4-CPA- and 6-BA-induced fruits between 1 and 14 DAA (Fig. 5).

Changes in the expression of GA catabolism genes for different treatments between 1 and 21 DAA. a–e VvGA2ox1, VvGA2ox2, VvGA2ox3, VvGA2ox4, and VvGA2ox7. Values are the mean ± SE of three biological replicates. Significant differences (p < 0.05) between treatments are indicated using different letters according to Tukey’s test
Cytokinin increases the endogenous IAA content by upregulating the expression of auxin biosynthesis genes
The IAA content in pollinated, unpollinated, and 6-BA-treated ovaries at different developmental stages was also analyzed. There was no difference in the IAA content among these treatments between 1 and 4 DAA. However, the IAA content in pollinated and 6-BA-induced fruits was higher than that in unpollinated ovaries between 7 and 21 DAA. A decrease in the IAA content was observed in unpollinated ovaries at 7 DAA (Fig. 6a). The transcript levels of VvYUC2 and VvYUC6 in pollinated and 6-BA-treated ovaries were higher than those in unpollinated ovaries (Fig. 6b, c), which was consistent with the IAA content changes after anthesis (Fig. 6a).

Effect of 6-BA on the induction of endogenous IAA. Changes in the IAA content (a) and the expression of IAA biosynthesis genes (VvYUC2 and VvYUC6) in unpollinated, pollinated, and 6-BA-treated ovaries (b and c). Values are the mean ± SE of three biological replicates. Significant differences (p < 0.05) between treatments are indicated using different letters according to Tukey’s test
Effect of plant growth regulator on the development of grape ovules
To clarify the different effects of plant growth regulators on the development of grape ovules (Fig. 1b), we compared the histology of seeds in pollinated and hormone-induced fruits at 4, 7, 14, and 21 DAA. Normal seed was evident in pollinated fruits, where zygote and endosperm cells were clearly observed (Fig. 7a–d). In the case of 4-CPA-induced fruits, the integument still developed and formed some vascular-like cells at 21 DAA, even though the egg cell and polar nucleus had already degenerated at 4 DAA (Fig. 7e–h). Regarding the 6-BA-induced fruits, the degeneration of the egg apparatus began at 4 DAA (Fig. 7i–l), but the ovule had developed compared to the GA-induced fruits (Supplemental Fig. S2 and Supplemental Table S2).

Ovule development in pollinated and 4-CPA- and 6-BA-treated seeds between 4 and 21 DAA. Normal zygote and endosperm nuclei from pollinated seeds at 4 DAA (a and b) and 7 DAA (c). Normal embryo development in pollinated seeds at 21 DAA (d). Abnormal embryo sac without an egg cell and polar nucleus in 4-CPA-treated seeds at 4 DAA (e), 7 DAA (f), and 14 DAA (g). Integument of the ovule still develops in 4-CPA-treated seeds at 21 DAA (h). Degradation of the egg apparatus from 6-BA-treated seeds at 4 DAA (i) and 7 DAA (j). Ovules of 6-BA-treated seeds show some level of development at 14 DAA (k) and 21 DAA (l). Scale bar, 100 μm. z zygote, en endosperm nuclei, s synergid
Discussion
The effects of different plant growth regulators on inducing parthenocarpy fruit growth vary with plant species. In tomato and sweet pepper, the parthenocarpic fruit that was induced with auxin was larger than the GA-induced fruits (Serrani et al. 2007b; Tiwari et al. 2012). Interestingly, GAs and auxin have an opposite effect on the fruit growth in Arabidopsis (Vivian-Smith and Koltunow 1999). For some crops, such as kiwifruit and bottle gourd, cytokinin has more potential to induce parthenocarpic fruit growth than auxin and GAs (Lewis et al. 1996; Yu 1999). In this work, the unpollinated grapevine ovaries treated with gibberellin, auxin, or cytokinin alone at anthesis develop parthenocarpically, as reported by Weaver and others (Weaver and Mccune 1960; Weaver et al. 1965). However, the fruits treated with auxin or cytokinin alone were smaller than the GA-induced fruits, the fruit size of which was approximate to pollinated fruits (Fig. 1). In contrast to 4-CPA and 6-BA application separately or simultaneously, which had the same effect on fruit development, simultaneous application of 4-CPA+GA3 or 6-BA+GA3 could generate fruits similar to pollinated fruits. Because there was a significant difference in fresh weight among these parthenocarpic fruits (Fig. 1), we inspected the microscopic sections of berries treated with different hormones at 21 DAA. The results showed that the fruits treated with GA3 alone had fewer cell layers but larger cell diameters in the mesocarp compared to the pollinated fruits. However, the cell layers of the mesocarp in 4-CPA-, 6-BA-, and 4-CPA+6-BA-treated fruits were similar to those of the pollinated fruits, and the cell diameter of the mesocarp was smaller than that of the pollinated and GA3-treated fruits. The cell layers and diameter of the mesocarp in parthenocarpic fruits were comparable to those in pollinated fruits when GA3 was applied along with 4-CPA or 6-BA (Fig. 1e and Table 1). These results suggested that all three hormones are required for grape fruit set and have a distinct role in this process, that is, GA stimulates mesocarp cell expansion and auxin and cytokinin stimulate mesocarp cell division. The effect of these hormones on cell division and cell expansion in grapevine is similar to the observation in tomato (Serrani et al. 2007b), but different from that in Arabidopsis (Vivian-Smith and Koltunow 1999). The coordinated action of these three hormones is also required for normal fruit development in Arabidopsis (Vivian-Smith and Koltunow 1999), tomato (Bünger-Kibler and Bangerth 1983), citrus (Guardiola et al. 1993), and pea (Vercher and Carbonell 1991).
PAC is an inhibitor of P450-dependent monoxygenases (Hedden and Graebe 1985) that has been used to reduce the GA content in pollinated ovaries of tomato (Fos et al. 2000, 2001). In this study, the application of PAC that reduced fruit set and fresh weight in pollinated fruits supports the hypothesis that grapevine fruit set and growth depend on the synthesized GAs after pollination and fertilization. Similarly, the fruit set and fresh weight of the unpollinated ovaries induced by 4-CPA or 6-BA were significantly reduced in the presence of PAC, and the effect of PAC was reversed by the application of GA3 (Fig. 2). These results are consistent with previous reports in tomato, Arabidopsis, and sweet pepper (Dorcey et al. 2009; Serrani et al. 2008; Tiwari et al. 2012), in which parthenocarpic growth induced by the application of auxin or cytokinin is mediated by GAs.
Endogenous bioactive GAs play a critical role in fruit set and growth in many plants (Csukasi et al. 2011; Fos et al. 2001; Zhang et al. 2010). The tomato mutations pat, pat2, and pat3/pat4 could induce a natural facultative parthenocarpic capacity by increasing the concentrations of bioactive GAs in the ovaries before pollination (Fos et al. 2001; Olimpieri et al. 2007). In the seedless mandarin Satuma, high levels of endogenous GAs are associated with an elevated parthenocarpic ability, while in another seedless mandarin Clementine, low levels of GAs are associated with a low ability of parthenocarpy and fruit set, suggesting that endogenous GA content is a limiting factor for parthenocarpic development (Talon et al. 1992). In this work, the content of active gibberellin in unpollinated ovaries was low and constant after anthesis. However, the GA1 + 3 and GA4 + 7 contents in pollinated fruits were relatively low between 1 and 4 DAA, increased at 7 DAA, and peaked at 14 DAA, in agreement with the idea that a high endogenous bioactive GA content is required in pollinated fruits during berry set and early berry cell enlargement. The changing patterns of GA1 + 3 and GA4 + 7 in 4-CPA- and 6-BA-induced fruits were similar to those in pollinated fruits, which suggested that the induction of parthenocarpy by auxin and cytokinin in grapevine is dependent on the accumulation of GAs (Fig. 3). On the other hand, the content of bioactive GAs, especially GA1 + 3 in the ovaries induced by 4-CPA and 6-BA, was lower than that in pollinate ovaries during 14–21 DAA (Fig. 3a), which was the key period for fruit set and enlargement (Coombe 1962; Ojeda et al. 1999). Combined with the results presented above, 4-CPA, 6-BA, and 4-CPA+6-BA induced smaller fruits compared to pollinated fruits, whereas the co-application of 4-CPA+GA3 or 6-BA+GA3 could acquire fruits that were similar to that by pollination at 21 DAA (Fig. 1). These results suggested that fruits treated with 4-CPA and 6-BA could develop parthenocarpically, although the development of parthenocarpic fruits was limited due to the lower endogenous GA content during the fruit enlargement period. In addition, the GA1 and GA4 contents also decreased after flowering in plum (El-Sharkawy et al. 2014), similar to what we observed in grapevine (Fig. 3), suggesting that the changes in the GA content in perennial plants were different from those in annual plants (Ozga et al. 2009; Serrani et al. 2007a), where there was a significant increase in the GA levels in the ovaries immediately after pollination.
GA metabolism genes are differentially expressed according to the developmental stage in different plants (Rieu et al. 2008; Serrani et al. 2007a; Zhu et al. 2006), and the expression levels of these genes have been reported to correlate with the endogenous GA content (Ozga and Reinecke 2003; Ozga et al. 2009; Rieu et al. 2008; Serrani et al. 2007b). After pollination, bioactive GAs accumulated in fertilized ovaries, accompanied by a high expression of GA20oxs genes in tomato (Dorcey et al. 2009; Olimpieri et al. 2007) or by transient upregulation of GA20oxs and GA3oxs genes in Arabidopsis (Dorcey et al. 2009). Here, based on the above experiment, we inferred that auxin and cytokinin may participate in the regulation of GA metabolism genes to promote the accumulation of bioactive gibberellin in the ovaries. Gene expression analysis showed that the transcript levels of VvGA20ox1, 2, and 3 and VvGA3ox1, 2, and 3 genes in pollinated and 4-CPA- and 6-BA-treated ovaries were higher than those in unpollinated ovaries after anthesis (Fig. 4). Similarly, 4-Cl-IAA application in pea promoted GA biosynthesis by increasing the transcripts of both GA20ox (van Huizen et al. 1997) and GA3ox (Ozga and Reinecke 2003) in deseeded pods. The content of GAs was also induced by cytokinin by upregulating GA biosynthesis genes in tomato (Ding et al. 2013). Furthermore, notably, VvGA3ox1 expression was significantly induced by 4-CPA and 6-BA treatment at 7 DAA (Fig. 4d). In the wine grape cultivar ‘Pinot Noir’ (V. vinifera L.), the VvGA3ox1 transcript was significantly upregulated at fruit set, even though its transcript was much less abundant than that of VvGA3ox2 and VvGA3ox3 (Giacomelli et al. 2013). However, the transcript abundance of VvGA3ox1 was higher than that of VvGA3ox2 and VvGA3ox3 in our work. VvGA3ox1 transcripts were lower in 4-CPA and 6-BA fruits than in pollinated fruits at 7 and 21 DAA (Fig. 4d), which may be associated with the difference in the GA content among these fruits (Fig. 3). These results suggest that VvGA3ox1 may play a main role during fruit set and enlargement period in grapevine.
In contrast to the GA biosynthesis genes, most catabolism-related VvGA2ox genes showed low expression in the fruits induced by pollination and 4-CPA and 6-BA treatment after anthesis (Fig. 5), which was especially true for the VvGA2ox3 and VvGA2ox4 transcript levels, which were much lower in pollinated and 4-CPA- and 6-BA-induced fruits than in unpollinated ovaries at 7 DAA (Fig. 5c, d). Generally, most VvGA2oxs transcripts showed a common trend, peaking at anthesis and then decreasing in grapevine (Dauelsberg et al. 2011; Giacomelli et al. 2013). These results were consistent with our hypothesis that the effect of 4-CPA and 6-BA on grape fruits set is mediated by active gibberellin through the enhancement of GA biosynthesis and suppression of GA catabolism.
In tomato, the application of CPPU can increase the concentration of endogenous GAs and IAA. It seems that cytokinin-induced fruits are dependent on the enhancement of the GA and the auxin contents (Ding et al. 2013). CPPU application also can induce the IAA content in kiwi fruit (Ohara et al. 1997). In agreement with these reports, we also observed increased IAA levels in pollinated and 6-BA-induced fruits. Furthermore, histology analysis of seeds showed that most of the unfertilized ovules degenerated in the GA3-induced fruits (Supplemental Fig. S2 and Supplemental Table S2), whereas they developed to some extent in the case of 4-CPA application at 21 DAA (Figs. 1 and 7). Induction of pseudoembryos by auxin application has also been reported in tomato (Ding et al. 2013), but not in grapevine. Considering that a certain degree of development was observed in the ovule of 6-BA-induced fruits and a similar transcript profile of GA metabolism genes was found in 4-CPA- and 6-BA-treated ovaries, we hypothesized that cytokinin may induce auxin biosynthesis in grape. The IAA content and gene expression analysis confirmed our speculation that cytokinin may enhance endogenous IAA content in grape ovary by regulating the expression of auxin biosynthesis genes VvYUC2 and VvYUC6 (Fig. 6).
In summary, the effects of auxin, cytokinin, and GA on fruit set in grapevine were investigated in this study. The results showed that the application of exogenous auxin and cytokinin induced parthenocarpy in grapevine, although the development of parthenocarpic fruits was limited due to less endogenous GA content during the fruit enlargement period. Auxin and cytokinin increased the active GA content in the fruit by upregulating GA biosynthesis genes (e.g., VvGA3ox1 and VvGA20oxs) and by inhibiting the expression of GA catabolism genes (e.g., VvGA2ox3 and VvGA2ox4), thus inducing fruit set. Nevertheless, non-treated unpollinated ovaries have a low GA content, which is associated with a low level of GA biosynthesis transcripts and a high level of GA catabolism transcripts after pollination, which then decreases at 21 DAA (Fig. 8). In addition, elevated expression of the IAA biosynthesis genes VvYUC2 and VvYUC6 was associated with the enhanced concentration of the cytokinin-induced active IAA in the ovaries. These results indicated that auxin- and cytokinin-induced fruit set in grapevine requires downstream gibberellin biosynthesis, which plays an essential role in the fruit set and enlargement period.

Scheme of the parthenocarpy fruit induced by auxin and cytokinin in grapevine. Pollination or auxin and cytokinin application increases the content of active GA1+3 and GA4+7 in the grape ovary by upregulating the transcription of genes encoding enzymes for GA biosynthesis (e.g., VvGA20oxs and VvGA3ox1) and by inhibiting the transcription of the genes encoding enzymes for GA inactivation (e.g., VvGA2ox3 and VvGA2ox4). In contrast, emasculated ovaries abscised at 21 DAA resulted from a low GA content
References
Aloni R, Aloni E, Langhans M, Ullrich CI (2006) Role of auxin in regulating Arabidopsis flower development. Planta 223:315–328. doi:10.1007/s00425-005-0088-9
Archbold DD, Dennis FG (1985) Strawberry receptacle growth and endogenous IAA content as affected by growth regulator application and achene removal. J Am Soc Hortic Sci 110:816–820
Böttcher C, Keyzers RA, Boss PK, Davies C (2010) Sequestration of auxin by the indole-3-acetic acid-amido synthetase GH3-1 in grape berry (Vitis vinifera L.) and the proposed role of auxin conjugation during ripening. J Exp Bot 61:3615–3625
Bünger-Kibler S, Bangerth F (1983) Relationship between cell number, cell size and fruit size of seeded fruits of tomato (Lycopersicon esculentum Mill.), and those induced parthenocarpically by the application of plant growth regulators. Plant Growth Regul 1:143–154
Coombe BG (1960) Relationship of growth and development to changes in sugars, auxins, and gibberellins in fruit of seeded and seedless varieties of Vitis vinifera. Plant Physiol 35:241
Coombe BG (1962) The effect of removing leaves, flowers and shoot tips on fruit-set in Vitis vinifera L. J Hortic Sci 37:1–15
Costantini E, Landi L, Silvestroni O, Pandolfini T, Spena A, Mezzetti B (2007) Auxin synthesis-encoding transgene enhances grape fecundity. Plant Physiol 143:1689–1694
Csukasi F, Osorio S, Gutierrez JR, Kitamura J, Giavalisco P, Nakajima M, Fernie AR, Rathjen JP, Botella MA, Valpuesta V, Medina-Escobar N (2011) Gibberellin biosynthesis and signalling during development of the strawberry receptacle. New Phytol 191:376–390
Dauelsberg P, Matus JT, Poupin MJ, Leiva-Ampuero A, Godoy F, Vega A, Arce-Johnson P (2011) Effect of pollination and fertilization on the expression of genes related to floral transition, hormone synthesis and berry development in grapevine. J Plant Physiol 168:1667–1674
Ding J, Chen B, Xia X, Mao W, Shi K, Zhou Y, Yu J (2013) Cytokinin-induced parthenocarpic fruit development in tomato is partly dependent on enhanced gibberellin and auxin biosynthesis. PLoS One 8:e70080
Dorcey E, Urbez C, Blázquez MA, Carbonell J, Perez-Amador MA (2009) Fertilization-dependent auxin response in ovules triggers fruit development through the modulation of gibberellin metabolism in Arabidopsis. Plant J Cell Mol Biol 58:318–332 (315)
El-Sharkawy I, Sherif S, Kayal WE, Mahboob A, Abubaker K, Ravindran P, Jyothi-Prakash PA, Kumar PP, Jayasankar S (2014) Characterization of gibberellin-signalling elements during plum fruit ontogeny defines the essentiality of gibberellin in fruit development. Plant Mol Biol 84:399–413
Fos M, Nuez F, García-Martínez JL (2000) The gene pat-2, which induces natural parthenocarpy, alters the gibberellin content in unpollinated tomato ovaries. Plant Physiol 122:471–479
Fos M, Proaño K, Nuez F (2001) Role of gibberellins in parthenocarpic fruit development induced by the genetic system pat-3/pat-4 in tomato. Physiol Plant 111:545–550
Fuentes S, Ljung K, Sorefan K, Alvey E, Harberd NP, Ostergaard L (2012) Fruit growth in Arabidopsis occurs via DELLA-dependent and DELLA-independent gibberellin responses. Plant Cell 24:3982–3996. doi:10.1105/tpc.112.103192
Gallego-Giraldo C, Hu J, Urbez C, Gomez MD, Sun TP, Perez-Amador MA (2014) Role of the gibberellin receptors GID1 during fruit-set in Arabidopsis. Plant J 79:1020–1032
Giacomelli L, Rota-Stabelli O, Masuero D, Acheampong AK, Moretto M, Caputi L, Vrhovsek U, Moser C (2013) Gibberellin metabolism in Vitis vinifera L. during bloom and fruit-set: functional characterization and evolution of grapevine gibberellin oxidases. J Exp Bot 64:4403–4419
Gillaspy G, Ben-David H, Gruissem W (1993) Fruits: a developmental perspective. Plant Cell 5:1439
Gorguet B, Heusden AV, Lindhout P (2005) Parthenocarpic fruit development in tomato. Plant Biol 7:131–139
Guardiola JL, Barrés MT, Albert C, García-Louis A (1993) Effects of exogenous growth regulators on fruit development in Citrus unshiu. Ann Bot 71:169–176 (168)
Hedden P, Graebe JE (1985) Inhibition of gibberellin biosynthesis by paclobutrazol in cell-free homogenates of Cucurbita maxima endosperm and Malus pumila embryos. J Plant Growth Regul 4:111–122
Hu JH, Mitchum MG, Neel B, Ayele BT, Ogawa M, Nam E, Lai WC, Hanada A, Alonso JM, Ecker JR, Swain SM, Yamaguchi S, Kamiya Y, Sun TP (2008) Potential sites of bioactive gibberellin production during reproductive growth in Arabidopsis. Plant Cell Online 20:320–336
Jung CJ, Hur YY, Yu HJ, Noh JH, Park KS, Lee HJ (2014) Gibberellin application at pre-bloom in grapevines down-regulates the expressions of VvIAA9 and VvARF7, negative regulators of fruit set initiation, during parthenocarpic fruit development. PLoS One 9:e95634
Kühn N, Arce-Johnson P (2012) Pollination: a key event controlling the expression of genes related to phytohormone biosynthesis during grapevine berry formation. Plant Signal Behav 7:7–11. doi:10.4161/psb.7.1.18353
Lavee S (1960) Effect of gibberellic acid on seeded grapes. Nature 185:395
Lewis DH, Burge GK, Hopping ME, Jameson PE (1996) Cytokinins and fruit development in the kiwifruit (Actinidia deliciosa). II. Effects of reduced pollination and CPPU application. Physiol Plant 98:187–195
Lu J, Lamikanra O, Leong S (1997) Induction of seedlessness in ‘Triumph’ muscadine grape (Vitis rotundifolia Michx.) by applying gibberellic acid. HortScience 32:89–90
Mapelli S (1981) Changes in cytokinin in the fruits of parthenocarpic and normal tomatoes. Plant Sci Lett 22:227–233
Mariotti L, Picciarelli P, Lombardi L, Ceccarelli N (2011) Fruit-set and early fruit growth in tomato are associated with increases in indoleacetic acid, cytokinin, and bioactive gibberellin contents. J Plant Growth Regul 30:405–415. doi:10.1007/s00344-011-9204-1
Martí C, Orzáez D, Ellul P, Moreno V, Carbonell J, Granell A (2007) Silencing of DELLA induces facultative parthenocarpy in tomato fruits. Plant J 52:865–876
Martinelli F, Uratsu SL, Reagan RL, Chen Y, Tricoli D, Fiehn O, Rocke DM, Gasser CS, Dandekar AM (2009) Gene regulation in parthenocarpic tomato fruit. J Exp Bot 60:3873–3890 (3818)
Murray MG, Thompson WF (1980) Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res 8:4321–4325
Ohara HKM, Matsui H, Hirata N, Takahashi E (1997) Comparison of the levels of endogenous plant growth substances in CPPU-treated and -untreated kiwi fruit. J Jpn Soc Hort Sci 65:692–705
Ojeda H, Deloire A, Carbonneau A, Ageorges A, Romieu C (1999) Berry development of grapevines: relations between the growth of berries and their DNA content indicate cell multiplication and enlargement. Vitis 38:145–150
Olimpieri I, Siligato F, Caccia R, Soressi GP, Mazzucato A, Mariotti L, Ceccarelli N (2007) Tomato fruit set driven by pollination or by the parthenocarpic fruit allele are mediated by transcriptionally regulated gibberellin biosynthesis. Planta 226:877–888
Ozga JA, Reinecke DM (1999) Interaction of 4-chloroindole-3-acetic acid and gibberellins in early pea fruit development. Plant Growth Regul 27:33–38
Ozga JA, Reinecke DM (2003) Hormonal interactions in fruit development. J Plant Growth Regul 22:73–81
Ozga JA, Reinecke DM, Ayele BT, Ngo P, Nadeau C, Wickramarathna AD (2009) Developmental and hormonal regulation of gibberellin biosynthesis and catabolism in pea fruit. Plant Physiol 150:448–462
Pattison RJ, Catalá C (2012) Evaluating auxin distribution in tomato (Solanum lycopersicum) through an analysis of the PIN and AUX/LAX gene families. Plant J 70:585–598
Pérez FJ, Viani C, Retamales J (2000) Bioactive gibberellins in seeded and seedless grapes: identification and changes in content during berry development. Am J Enol Vitic 51:315–318
Picken AJF (1984) A review of pollination and fruit set in the tomato (Lycopersicon esculentum Mill.). J Hortic Sci 59:1–13
Rieu I, Ruiz-Rivero O, Fernandez-Garcia N, Griffiths J, Powers SJ, Gong F, Linhartova T, Eriksson S, Nilsson O, Thomas SG, Phillips A, Hedden P (2008) The gibberellin biosynthetic genes AtGA20ox1 and AtGA20ox2 act, partially redundantly, to promote growth and development throughout the Arabidopsis life cycle. Plant J 53:488–504
Ruan Y-L, Patrick JW, Bouzayen M, Osorio S, Fernie AR (2012) Molecular regulation of seed and fruit set. Trends Plant Sci 17:656–665. doi:10.1016/j.tplants.2012.06.005
Serrani JC, Sanjuán R, Ruiz-Rivero O, Fos M, García-Martínez JL (2007a) Gibberellin regulation of fruit set and growth in tomato. Plant Physiol 145:246–257
Serrani JC, Fos M, Atarés A, García-Martínez JL (2007b) Effect of gibberellin and auxin on parthenocarpic fruit growth induction in the cv Micro-Tom of tomato. J Plant Growth Regul 26:211–221
Serrani JC, Ruiz-Rivero O, Fos M, García-Martínez JL (2008) Auxin-induced fruit-set in tomato is mediated in part by gibberellins. Plant J 56:922–934
Talon M, Zacarias L, Primo-Millo E (1992) Gibberellins and parthenocarpic ability in developing ovaries of seedless mandarins. Plant Physiol 99:1575–1581
Teng N, Wang J, Chen T, Wu X, Wang Y, Lin J (2006) Elevated CO2 induces physiological, biochemical and structural changes in leaves of Arabidopsis thaliana. New Phytol 172:92–103
Tiwari A, Offringa R, Heuvelink E (2012) Auxin-induced fruit set in Capsicum annuum L. requires downstream gibberellin biosynthesis. J Plant Growth Regul 31:570–578
Van Huizen R, Ozga JA, Reinecke DM (1997) Seed and hormonal regulation of gibberellin 20-oxidase expression in pea pericarp. Plant Physiol 115:123–128
Vercher Y, Carbonell J (1991) Changes in the structure of ovary tissues and in the ultrastructure of mesocarp cells during ovary senescence or fruit development induced by plant growth substances in Pisum sativum. Physiol Plant 81:518–526
Vivian-Smith A, Koltunow AM (1999) Genetic analysis of growth-regulator-induced parthenocarpy in Arabidopsis. Plant Physiol 121:437–452
Vivian-Smith A, Luo M, Chaudhury A, Koltunow A (2001) Fruit development is actively restricted in the absence of fertilization in Arabidopsis. Development 128:2321–2331
Wang H, Schauer N, Usadel B, Frasse P, Zouine M, Hernould M, Latché A, Pech JC, Fernie AR, Bouzayen M (2009) Regulatory features underlying pollination-dependent and -independent tomato fruit set revealed by transcript and primary metabolite profiling. Plant Cell 21:1428–1452
Weaver RJ, Mccune SB (1960) Further studies with gibberellin on Vitis vinifera grapes. Bot Gaz 121:155–162
Weaver R, Mccune S, Hale C (1962) Effect of plant regulators on set and berry development in certain seedless and seeded varieties of Vitis vinifera L. Vitis 3:84–96
Weaver RJ, van Overbeek J, Pool RM (1965) Induction of fruit set in Vitis vinifera L. by a kinin. Nature 206:952–953
Yu JQ (1999) Parthenocarpy induced by N-(2-chloro-4-pyridyl)-N′-phenylurea (CPPU) prevents flower abortion in Chinese white-flowered gourd (Lagenaria leucantha). Environ Exp Bot 42:121–128 (128)
Zhang X, Luo G, Wang R, Wang J, Himelrick DG (2003) Growth and developmental responses of seeded and seedless grape berries to shoot girdling. J Am Soc Hortic Sci Am Soc Hortic Sci 128:316–323
Zhang C, Tateishi N, Tanabe K (2010) Pollen density on the stigma affects endogenous gibberellin metabolism, seed and fruit set, and fruit quality in Pyrus pyrifolia. J Exp Bot 61:4291–4302
Zhu YY, Takahito N, Xu YH, Zhang YY, Peng Y, Mao BZ, Atsushi H, Zhou HC, Wang RX, Li P, Zhu XD, Mander LN, Yuji K, Shinjiro Y, He ZH (2006) Elongated uppermost internode encodes a cytochrome p450 monooxygenase that epoxidizes gibberellins in a novel deactivation reaction in rice. Plant Cell 18:442–456
Acknowledgments
The National Natural Science Foundation of China (31471842) supported this research.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The sequences of the VvGA20oxs, VvGA3oxs, VvGA2oxs, and VvYUCs reported here are available in GenBank (http://www.ncbi.nlm.nih.gov/): VvGA20ox1 [GenBank accession no. KC898188]; VvGA20ox2 [GenBank accession no. KC898186]; VvGA20ox3 [GenBank accession no. KC KC898189]; VvGA3ox1 [GenBank accession no. KC898176]; VvGA3ox2 [GenBank accession no. KC898175]; VvGA3ox3 [GenBank accession no. KC898177]; VvGA2ox1 [GenBank accession no. KC898179]; VvGA2ox2 [GenBank accession no. KC898180]; VvGA2ox3 [GenBank accession no. KC898181]; VvGA2ox4 [GenBank accession no. KC898182]; VvGA2ox7 [GenBank accession no. KC898184]; VvYUC2 [GenBank accession no. XM_002280979]; VvYUC6 [GenBank accession no. XM_010650228].
Additional information
Communicated by M. Wirthensohn
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplemental Table S1
Primers used for the qRT-PCR performed in this work. (DOCX 18 kb)
Supplemental Table S2
The size of ovule from different treatment at 21 DAA. Values are the means of eight sections (±SE), one section per fruit, and eight fruits. Significant differences (p < 0.05) between treatments are indicated using different letters according to a Tukey’s test. (DOCX 16 kb)
Supplemental Fig. S1
Intact inflorescence (a) and emasculated ovaries (b) at 10 DBA (Days before anthesis). (JPG 3767 kb)
Supplemental Fig. S2
Ovule with degrading embryo sac from GA3-treated unpollinated ovaries at 14 (a) and 21 DAA (b). Scale bar=100 μm. (JPG 3776 kb)
Rights and permissions
About this article

Cite this article
Lu, L., Liang, J., Zhu, X. et al. Auxin- and cytokinin-induced berries set in grapevine partly rely on enhanced gibberellin biosynthesis. Tree Genetics & Genomes 12, 41 (2016). https://doi.org/10.1007/s11295-016-0980-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11295-016-0980-4