
Abstract
Cementation of salt-containing soils can be achieved by salt-tolerant or halophilic calcite precipitation bacteria. Therefore, the isolation of calcite-producing bacteria in the presence of salt is the first step in the microbial cementation of saline soils. Urease producing bacteria can cause calcite nano-crystals to precipitate by producing urease in the presence of urea and calcium. The purpose of this study was to isolate urease producing halophilic bacteria in order to make calcite precipitate in saline soil. The calcite and the properties of the strains were further analyzed by X-ray diffraction (XRD) and scanning electron microscope equipped with an energy dispersive X-ray detector. In this study, a total of 110 halophilic strains were isolated, from which 58 isolates proved to have the ability of urease production. Four strains were identified to produce nano-calcite using urease activity in the precipitation medium. The XRD studies showed that the size of these particles was in the range of 40–60 nm. Strain H3 revealed that calcite is mostly produced in the precipitation medium containing 5% salt in comparison with other strains. This strain also produced calcite precipitates in the precipitation medium containing 15% salt. Phylogenetic analysis indicated that these isolates are about 99–100% similar to Staphylococcus saprophyticus.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
A variety of microbial metabolic pathways including photosynthesis, ammonification, denitrification, urea hydrolysis and sulfate reduction can lead to the precipitation of carbonates in different natural environments (Hammes et al. 2003). The potential of microorganisms to synthesize calcium carbonate is called microbially induced calcite precipitation (MICP), which has many uses in civil engineering and environmental science. Urea hydrolyzing bacteria can make calcium carbonate precipitate due to bacterial hydrolysis of urea in the presence of calcium ions. Microbial precipitation of calcium carbonate results in the production of different polymorphs such as calcite, aragonite, vaterite and two hydrated crystalline components. Among these, calcite is the most common and stable bacterial calcium carbonate polymorph (Rodriguez Navarro et al. 2012). The production of these polymorphs of calcium carbonate depends both on their growing environments and bacterial strains.
MICP has emerged as a novel technique and successful alternative for remediation and restoration of different building materials in recent years (Dhami et al. 2012, 2016). From the biological point of view, the MICP process is regulated mainly by four key factors: (1) concentration of calcium ion; (2) concentration of dissolved inorganic carbon; (3) pH; and (4) availability of nucleation sites (Hammes and Verstraete 2002). Several genera of halophilic bacteria such as Halomonas, Deleya, Flavobacterium, Acinetobacter and Salinivibrio and others have been reported to make carbonates precipitate in natural marine habitats (Ferrer et al. 1988a, b; Rivadeneyra et al. 1993, 2006). To date, a lot of researchers have studied carbonate precipitation in the saline aquatic environments, freshwater environments, and saline soils (Baskar et al. 2005; Novitsky 1981; Rivadeneyra et al. 2004).
In the MICP process, the hydrolysis of urea by urease can increase the pH of the medium during the MICP process. The carbon dioxide produced by urease activity or respiration is quickly transformed into bicarbonate (HCO3−) in the presence of water. This bicarbonate reacts with the hydroxyl ions to form carbonate (CO32−) ions, and in the presence of calcium ions, the calcite is precipitated (Castanier et al. 2000). As, urease producing bacteria used for bio-cementation are active in the presence of high salt concentrations, halophilic or halotolerant, and alkaliphilic urease producing bacteria are thus preferred choices for bio-cementation (Mortensen and DeJong 2011).
Halophilic microorganisms use different strategies to overcome the stress of high salt concentrations. Halophilic archaea maintain their cytoplasmic osmotic balance by accumulating high concentrations of inorganic ions in the cytoplasm. In contrast, halophilic or halotolerant bacteria adapt themselves by accumulating high concentrations of various organic compatible solutes. These mechanisms are called “salt in cytoplasm strategy” and “salt out cytoplasm strategy”, respectively (Galinski and Trüper 1994). Some researchers such as Rivadeneyra showed that both Na+ and Cl− ions exert a weak or no influence on the microbial precipitation of carbonates (Rivadeneyra et al. 1991). Deleya halophila is a moderately halophilic microorganism, whose salt requirements and its ability to precipitate carbonates make it a useful tool in carbonate precipitation studies. Studies on this bacterium have shown that high salinity has a negative effect on calcite production (Ferrer et al. 1988b). Also, the crystals that are formed are calcite and vaterite, and the ratio of calcite to vaterite is dependent on the amount of salt and type of medium (Rivadeneyra et al. 1991). Further, aragonite is not detected even at high magnesium ion concentration in the medium (Ferrer et al. 1988b).
The purpose of this study was to isolate a potent bacterium with calcite precipitation ability in the presence of high salt concentration. To that end, this study investigated the effect of salinity on the type of crystal formation by using isolated bacteria in the precipitation media. The effect of salt amount on the type of crystalline calcium carbonate was studied by X-ray diffraction (XRD).
Materials and methods
Isolation of halophilic bacteria
About 5 g of soil sample or 5 ml of water sample were suspended in 45 ml of sterilized physiology serum and then mixed by shaking on a shaker for several hours at 120 rpm. The flasks were shaken thoroughly and the suspension was let clarified by settling. Fifty microliters of diluted samples were spread on solid tryptic soy broth (TSB) (Biolife, Italy) and Luria–Bertani media. Plates were incubated at 30 °C for 5 days. In order to isolate halophilic bacteria, salt was added to TSB medium at different concentrations (up to 20%) (Carpa et al. 2014; Vahed et al. 2011).
Isolation of urease producing bacteria
The bacteria isolated in the previous step were selected for urease testing. For this purpose, each of the purified isolates was cultured on a solid medium containing 30 g/l TSB, 0.012 g/l MnSO4·H2O, 0.024 g/l NiCl2·6H2O, 0.01 g/l phenol red, 20 g/l urea, and 15 g/l agar (pH 6.8–7.3). In addition, due to the fact that halophilic or halotolerant bacteria need salt, 0–20 percent NaCl was added to the medium. Urease producing isolates brought about the alkalization of the culture medium and thus changed the color to pink by releasing ammonium due to the urease activity (Stabnikov et al. 2013).
Isolation of calcite-producing bacteria
Purified bacteria were cultured on precipitation medium at 30 °C for 2 weeks. Colonies were screened based on the observation of calcite precipitation by a light microscope. Precipitation medium consisted of 20 g/l urea, 2.12 g/l NaHCO3, 10 g/l NH4Cl, 3 g/l nutrient broth (Merck, Germany), 25 g/l CaCl2·2H2O, 0–200 g/l NaCl, and 15 g/l agar. The pH of medium was adjusted to 8 with the addition of 1 N NaOH (Chahal et al. 2011).
Specific urea degradation rate
Each isolate was cultured overnight on a urea agar medium containing 5% salt and incubated at 30 °C. A loop of the active bacterium was transferred to the broth medium containing urea and 5% salt, and then 1% of it was inoculated into the culture medium when OD600 nm reached 0.6–0.8. Culture medium was incubated at 30 °C on a shaking incubator at 120 rpm, and every 8 h optical density (OD) and total ammonium nitrogen of each isolate were measured using a spectrophotometer at 600 nm and Nessler method (Thompson and Morrison 1951), respectively. Specific urea degradation (SUD) rate was defined as the ratio of ammonium production (AP) per unit of bacterial growth in a given time (t), which is obtained by the following formula (Dick et al. 2006):
Analysis of nanocrystals
The colonies that produced nanocrystals were cultured on precipitation medium and then were collected and dried at 37 °C for XRD analysis. XRD measurements were done by a one-circle µ = µ diffractometer (XRD, D8 ADVANCE, Germany). The work condition of XRD was CuKα radiation via a rotating anode at 40 kV and 40 mA. The data were collected at a step of 0.05° 2−1 s and range of scattering angles (2θ) from 5° to 80°. The crystalline phases were identified using the ICDD database. Crystallite particle size was determined by XRD using Scherrer formula (Yena et al. 2007). Dried samples were ground into fine powder using agate mortar and then coated with carbon, and these samples were used for size characteristics of nanocrystals by scanning electron microscope (SEM) (MIRA3 TESCAN, Czech Republic). The elementary composition of the nanocrystals was determined qualitatively by energy dispersive X-ray spectrometry (EDS) with an In Beam-SE detector.
Identification of calcifying bacterial isolates
Nanocalcite producing isolates were identified using morphological, biochemical and molecular analysis. Biochemical tests were performed according to Bergey’s manual of systematic bacteriology (Brenner et al. 2005). Molecular phylogenies were determined by sequencing of 16S rRNA using universal primers 27F-YM (Frank et al. 2008) and 1492R-Y (Gökçekus et al. 2011). Bacterial DNA was extracted using boiling method. The 1.5 ml of bacterial culture was centrifuged for 10 min at 6000×g and the pellet was suspended in biology grade water and centrifuged at 6000×g for 10 min. The cells were washed three times using this water, subjected to two freeze–thaw cycles, suspended in 1 ml of molecular biology grade water, boiled at 100 °C in a water bath for 15 min, and centrifuged at 6000×g for 5 min (Yang et al. 2008). The supernatant was used directly for PCR or stored at − 20 °C.
Amplification reactions were conducted in 25 µl volume containing 1 µl of each primer (10 ρmol/µl), 0.5 µl of dNTPs (10 mM), 2.5 µl of 10× PCR buffer, 0.75 µl of MgCl2 (50 Mm), 18 µl of PCR H2O, template DNA extracted from 1 µl of bacterium, and 0.25 µl of Taq polymerase (5 U/µl) (Sina Colon, Iran). The PCR was performed in a Thermal Cycler (T100, Bio-Rad, USA) using appropriate programs optimized for these primers. After denaturation of DNA through heating for 4 min at 94 °C, the PCR program was run for 35 cycles; each cycle consisted of these steps: denaturation at 94 °C for 1 min, annealing at 55 °C for 1 min, and extension at 72 °C for 2 min (Gökçekus et al. 2011). This was followed by a final elongation step for 5 min at 72 °C. After amplification, purified PCR product was sequenced in both directions using an automated sequencer by Macrogen (Seoul, Korea). Sequences were edited by Fintch TV version 1.4.0. (URL: http://www.geospiza.com/finchtv.html) and assembled with BioEdit version 7.1.3.0 (URL: http://www.mbio.ncsu.edu/BioEdit/bioedit.html). All the assembled sequences were aligned with the published sequence using BLASTN program, and sequences were submitted to NCBI database. Phylogenetic tree of bacterial 16S rRNA was studied based on Bayesian inference (MrBayes) (Huelsenbeck and Ronquist 2001). For Bayesian phylogenetic inference, we used MrBayes v3.2.6 (MCMC ngen 6 million generations, samplefreq 1000, nchains 4 and the first 25% of the trees sampled as burn-in) (Ronquist and Huelsenbeck 2003). The analyses were stopped when the standard deviation of split frequencies was well below 0.01. Trees were visualized using TreeGraph2 (Stöver and Müller 2010).
Results
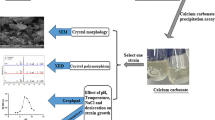
Isolation of calcite precipitating bacteria
A total of 110 halophilic strains were isolated, from which 58 isolates had the ability of urease production. The microscopic studies of colonies showed that only four isolates were able to produce crystals on precipitation medium. Strains were isolated from sediments of Kish Island (H1), water of Kish Island (H3, H4) (Kish Island, Iran) and tap water from Ahvaz (H5) (Ahvaz, Iran). All the four isolates produced crystals on precipitation medium. As seen in Fig. 1, these isolates produced different types of calcite morphology.

Morphology of monocrystal and polycrystal produced by isolates H1 (a), H3 (b), H4 (c) and H5 (d) strains cultured on precipitating medium containing 5% salt at × 10 magnification. Crystals are visible outside and inside colonies
Specific urea degradation rate
According to SUD rate, it was shown that strain H5 has the maximum urease activity. This strain produces ammonium 2.55 mmol/l in an hour. The SUD rate of some strains is illustrated in Fig. 2. H3 produces 1.66 mmol of ammonium per liter in an hour.

The SUD rate for H1 (dashed square), H3 (dotted square), H4 (dashed triangle) and H5 (dashed diamond)
XRD analysis
On the basis of Fig. 3, strain H1, in precipitation medium without additional salt, deposited only calcite (Fig. 3a), but in precipitation medium with 5% salt (Fig. 3b), in addition to calcite, halite and sal ammoniac were also produced. No crystals were formed in precipitation medium containing 10% salt by this strain.

XRD patterns of precipitates collected from isolates 1, 3, 4, and 5, cultured on precipitating medium with different NaCl concentrations: a H1, without added NaCl, b H1, 5% NaCl, c H3, 5% NaCl, d H3, 15% NaCl, e H4, without salt, f H4, 5% NaCl, g H5, without added NaCl, h H5, 5% NaCl
Strain H3 showed the most amount of calcite production in precipitation medium containing 5% salt in comparison with other strains. In addition to calcite, halite and sal ammoniac were detected in the dried colony powder of this bacterium (Fig. 3c). This strain was also able to produce calcite precipitates in precipitation medium containing 15% salt (Fig. 3d). The amount of calcite in the medium containing 5% NaCl was more than in the other medium with 15% NaCl.
Studies on strain H4 showed that this strain could also produce calcite precipitates in a precipitation medium with or without 5% salt (Fig. 3e, f). In precipitation medium with 10% salt, only halite and hydroxyapatite were produced, and calcite was not detected (not shown here). Strains H1, H4 and H5 produced more calcites in a medium containing 5% NaCl compared to a salt free medium. In strain H5, XRD analysis showed calcite and halite in the precipitation media with or without salt. In this strain, hydroxyapatite was produced in the precipitation medium without salt, and sal ammoniac was produced in the precipitation medium of 5% salt. But in the precipitation medium with 10% salt, no crystals were produced by strain H5 (not shown here). Table 1 summarizes the types of crystals produced by the strains examined in different salt concentrations.
SEM and EDX analysis
Figure 4 shows the image of the SEM of the crystals formed by isolate H1. SEM showed that calcite nanocrystals produced by strain H1 are homogeneous in shape and size. According to SEM analysis, it seems that calcite nano-particles have been integrated by the bacteria trapped inside them. The XRD studies showed that the size of these particles was in the range of 40–60 nm. Figure 5 shows energy dispersive X-ray (EDX) spectrum of precipitates of H1, which proves the presence of its constituent elements of calcite.

SEM and EDX images of aggregation of nanocrystals produced by strain H1 on bacterial surfaces in precipitating medium with different degrees of magnification. The scale bar is 5, 2, 1 micron and 200 nm, respectively

SEM–EDX analyses of three areas from a calcinated precipitate produced by strain H1. The scale bar is 5, 2, 1 micron and 200 nm, respectively
Bacteria identification
These strains were identified by biochemical and molecular analysis. The results of the biochemical tests of the isolates are presented in Table 2. All the strains can use maltose, sucrose, fructose and glucose as their carbon and energy sources. These isolates can grow in the presence of 15% of salt and therefore can tolerate and grow in the highly salty soils. The 16S rRNA gene sequence of strains 1, 3, 4 and 5 showed to be similar, respectively, by 99.85%, 99.92%, 99.92%, and 99.92% to Staphylococcus saprophyticus. Their 16S rRNA sequences were deposited in GenBank under the accession numbers of MG655151, MG655152, MG655153 and MG655154, respectively. Figure 6 shows phylogenetic tree of isolates according to MrBayes. The amplified PCR product was approximately 1500 bp in length, and the alignment length was 1391 characters (including indels). H5 and H3 strains, respectively, were placed in separate clads of I and IV with 0.926 the posterior probability, while H1 and H4 strains were branched together in one cluster.

Phylogenetic tree of isolated bacterial 16S rRNA studied based on Bayesian inference (MrBayes). S.: Staphylococcus (Ronquist and Huelsenbeck 2003)
Discussion
Many studies have suggested that urease-producing bacteria can produce calcite through urease enzyme activity. Our data showed that urease activity was present in four isolates when tested in the urease activity assay media. Studies of Hammes et al. (2003) and Wei et al. (2015) showed a good relationship between urease activity and calcite production. Four isolates of S. saprophyticus, Sporosarcina globispora, Bacillus lentus, and Sporosarcina sp. with a high level of urease activity were isolated from calcareous soils and sands. S. saprophyticus had a higher SUD rate than other strains, but there was not a strong relationship between the calcite precipitation and urease activity (Kim and Youn 2016). Similarly, we did not observe a strong relationship between urease activity and calcite production in these strains. H5 strain had a good urease activity in comparison with other strains. However, H3 strain produced more calcites than other strains. The main crystal produced by Deleya halophila in the presence of salt, which is a moderately halophilic microorganism, was aragonite (Rivadeneyra et al. 1996). In the present study, isolates H1, H3, H4 and H5 produced calcite in the precipitation medium with or without salt. In contrast to the above research, there was not any aragonite produced by these strains as determined by XRD analysis of crystals. Additionally, in the presence of strains such as H4 and H5, more calcites were produced in the precipitation medium containing 5% NaCl in comparison with salt-free medium. In the precipitation medium containing 5% salt, calcite is mostly produced by isolate H3 in comparison with other strains. Unlike other strains, this strain also produced calcite precipitates when the salt concentration reached 10%. In addition, this strain produced calcite in the 15% salt containing precipitation medium. Lots of research have shown that an increase in salinity decreases calcite precipitation. In the medium of halophilic bacteria Exiguobacterium mexicanum, calcium carbonate precipitation decreased significantly by 10% salt stress compared with 3.6–7.5% salt stress (Bansal et al. 2016). All strains in this study produced calcite in salt free precipitation medium, but by increasing the salt concentration the amount of deposited calcite decreased and then disappeared in some strains (e.g. H1, H4, and H5).
The SUD profile in Fig. 2 suggests that urease activity behavior of each strain is specific to itself and there is a specific point for optimum activity. H3 strain produces calcite precipitation in 5% salt containing medium more than in 15% salt containing and salt-free precipitation media. Ferrer et al. (1988b) showed that optimal calcium carbonate crystal formation occurs at a salt concentration of 2.5% in Deleya halophila. In another study by Del Moral et al. (1987), calcite precipitation was tested with various species of moderately halophilic bacteria and it was found that these bacteria produced calcite and vaterite at 2% total salt, pure calcite at 10% salt, and mainly aragonite at 20% total salt.
The very good production of calcite by isolate 3 in the 5% salt containing medium suggested that this strain may be a good industrial strain for cementation of saline soils. The most notable result of the study is application of halophilic urease producing bacteria in cementation of saline soil. This study demonstrated there are a lot of bacteria in saline soil that can cause precipitation in biocementation with stimulation. Use of indigenous halophilic bacteria of salty soil is a new important method for cementation of salty soil.
The presence of other ions, such as magnesium, could interfere with calcite formation (Sánchez-Román et al. 2007). High concentration of magnesium ion could also result in deposition of magnesium carbonate trihydrate (MgCO3·3H2O) besides the formation of calcite in MICP process (Cheng et al. 2014). In this study, like those conducted by Sánchez-Román et al. (2007) and Cheng et al. (2014), in addition to calcite, other crystals such as halite, sal ammoniac, and hydroxyapatite were deposited. In the present study, the XRD analysis showed the presence of calcite in culture media, and EDX analysis demonstrated that chemical composition of materials would be CaCO3. In studies by Bansal, SEM analysis of the mortar specimens subjected to bacterial treatment revealed the presence of dense matrix of crystals with visible rod shaped bacterial cells in close association with crystals (Bansal et al. 2016). A similar arrangement was seen in H4 strain between nanocrystals and bacteria. The reason for this phenomenon is that bacteria act as nucleation sites in calcite precipitation.
References
Bansal R, Dhami NK, Mukherjee A, Reddy MS (2016) Biocalcification by halophilic bacteria for remediation of concrete structures in marine environment. J Ind Microbiol Biotechnol 43:1497–1505
Baskar S, Baskar R, Mauclaire L, Mckenzie JA (2005) Role of microbial community in stalactite formation, Sahastradhara caves, Dehradun, India. Curr Sci 88:1305–1308
Brenner DJ, Krieg NR, Staley JT, Garrity GM (2005) Bergey’s manual of systematic bacteriology, vol 2. Springer, New York, pp 392–420
Carpa R, Keul A, Muntean V, Dobrotă C (2014) Characterization of halophilic bacterial communities in Turda Salt Mine (Romania). Orig Life Evol Biosph 44:223–230
Castanier S, Le Metayer-Levrel G, Perthuisot JP (2000) Bacterial roles in the precipitation of carbonate minerals. In: Riding RE, Awramik SM (eds) Microbial sediments. Springer, Heidelberg, pp 32–39
Chahal N, Rajor A, Siddique R (2011) Calcium carbonate precipitation by different bacterial strains. Afr J Biotechnol 10:8359–8372
Cheng L, Shahin M, Cord-Ruwisch R (2014) Bio-cementation of sandy soil using microbially induced carbonate precipitation for marine environments. Géotechnique 64:1010–1013
Del Moral A, Roldan E, Navarro J, Monteoliva-Sanchez M, Ramos-Cormenzana A (1987) Formation of calcium carbonate crystals by moderately halophilic bacteria. Geomicrobiol J 5:79–87
Dhami NK, Reddy M, Mukherjee A (2012) Biofilm and microbial applications in biomineralized concrete. In: Seto J (ed) Advanced topics in biomineralization. InTech, Rijeka, pp 137–164
Dhami NK, Reddy MS, Mukherjee A (2016) Significant indicators for biomineralisation in sand of varying grain sizes. Constr Build Mater 104:198–207
Dick J, De Windt W, De Graef B, Saveyn H, Van der Meeren P, De Belie N, Verstraete W (2006) Bio-deposition of a calcium carbonate layer on degraded limestone by Bacillus species. Biodegradation 17:357–367
Ferrer MR, Quevedo-Sarmiento J, Rivadeneyra MA, Bejar V, Delgado R, Ramos-Cormenzana A (1988a) Calcium carbonate precipitation by two groups of moderately halophilic microorganisms at different temperatures and salt concentrations. Curr Microbiol 17:221–227
Ferrer MR, Quevedo-Sarmiento J, Bejar V, Delgado R, Ramos-Cormenzana A, Rivadeneyra MA (1988b) Calcium carbonate formation by Deleya halophila: effect on salt concentration and incubation temperature. Geomicrobiol J 6:49–57
Frank JA, Reich CI, Sharma S, Weisbaum JS, Wilson BA, Olsen GJ (2008) Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl Environ Microbiol 74:2461–2470
Galinski EA, Trüper HG (1994) Microbial behavior in salt stressed ecosystems. FEMS Microbiol Rev 15:95–108
Gökçekus H, Türker U, LaMoreaux JW (2011) Survival and sustainability: environmental concerns in the 21st century. Springer, Berlin
Hammes F, Verstraete W (2002) Key roles of pH and calcium metabolism in microbial carbonate precipitation. Rev Environ Sci Biotechnol 1:3–7
Hammes F, Boon N, Villiers JD, de Villiers J, Verstraete W, Douglas Siciliano S (2003) Strain-specific ureolytic microbial calcium carbonate precipitation. Appl Environ Microbiol 69:4901–4909
Huelsenbeck JP, Ronquist F (2001) MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17:754–755
Kim G, Youn H (2016) Microbially induced calcite precipitation employing environmental isolates. Materials 9:468
Mortensen BM, DeJong JT (2011) Strength and stiffness of MICP treated sand subjected to various stress paths. In: Han J, Alzamora DA (eds) Geo-Frontiers 2011: advances in geotechnical engineering, vol 211. ASCE Geotechnical Special Publication, Dallas, pp 4012–4020
Novitsky JA (1981) Calcium carbonate precipitation by marine bacteria. Geomicrobiol J 2:375–388
Rivadeneyra MA, Delgado R, Quesada E, Ramos-Cormenzana A (1991) Precipitation of calcium carbonate by Deleya halophila in media containing NaCl as sole salt. Curr Microbiol 22:185–190
Rivadeneyra MA, Delgado R, Delgado G, Moral A, Ferrer MR, Ramos-Cormenzana A (1993) Precipitation of carbonate by Bacillus sp. isolated from saline soils. Geomicrobiol J 11:175–184
Rivadeneyra MA, Ramos-Cormenzana A, Delgado G, Delgado R (1996) Process of carbonate precipitation by Deleya halophila. Curr Microbiol 32:308–313
Rivadeneyra MA, Parraga J, Delgado R, Ramos-Cormenzana A, Delgado G (2004) Biomineralization of carbonates by Halobacillus trueperi in solid and in liquid media with different salinities. FEMS Microbiol Ecol 48:39–46
Rivadeneyra MA, Delgado R, Párraga J, Ramos-Cormenzana A, Delgado R (2006) Precipitation of minerals by 22 species of moderately halophilic bacteria in artificial marine salts media: influence of salt concentration. Folia Microbiol 51:445–453
Rodriguez Navarro C, Jroundi F, Schiro M, Ruiz-Agudo E, Gonza ´lezMun ˜oz MT (2012) Influence of substrate mineralogy on bacterial mineralization of calcium carbonate: implications in stone conservation. Appl Environ Microbiol 78:4017–4029
Ronquist F, Huelsenbeck JP (2003) MrBayes: Bayesian phylogenetic inference under mixed models. Bioinformatics 19:1572–1574
Sánchez-Román M, Rivadeneyra MA, Vasconcelos C, McKenzie JA (2007) Biomineralization of carbonate and phosphate by moderately halophilic bacteria. FEMS Microbiol Ecol 61:273–284
Stabnikov V, Jian C, Ivanov V, Li Y (2013) Halotolerant, alkaliphilic urease-producing bacteria from different climate zones and their application for biocementation of sand. World J Microbiol Biotechnol 29:1453–1460
Stöver BC, Müller KF (2010) TreeGraph 2: combining and visualizing evidence from different phylogenetic analyses. BMC Bioinformatics 11:7
Thompson J, Morrison G (1951) Determination of organic nitrogen control of variables in the use of Nessler’s reagent. Anal Chem 23:1153–1157
Vahed SZ, Forouhandeh H, Hassanzadeh S, Klenk HP, Hejazi MA, Hejazi MS (2011) Isolation and characterization of halophilic bacteria from Urmia Lake in Iran. Microbiol 80:834–841
Wei S, Cui H, Jiang Z, Liu H, He H, Fang N (2015) Biomineralization processes of calcite induced by bacteria isolated from marine sediments. Braz J Microbiol 46:455–464
Yang JL, Wang MS, Cheng AC, Pan KC, Li CF, Deng SX (2008) A simple and rapid method for extracting bacterial DNA from intestinal microflora for ERIC-PCR detection. World J Gastroenterol 14:2872–2876
Yena FS, Chang PL, Yu PC, Yang RJ (2007) Characterization on microstructure homogeneity of θ-Al2O3 powder systems during phase transformation. Key Eng Mater 351:81–87
Acknowledgements
The current study was supported by the Grant from Shahid Chamran University of Ahvaz to Maryam Haddadi for obtaining M.Sc. degree. The authors would like to thank Sara Ghashghaei and Masoud Mohammadi Farsani for their help.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ghezelbash, G.R., Haddadi, M. Production of nanocalcite crystal by a urease producing halophilic strain of Staphylococcus saprophyticus and analysis of its properties by XRD and SEM. World J Microbiol Biotechnol 34, 174 (2018). https://doi.org/10.1007/s11274-018-2544-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11274-018-2544-2