
Abstract
In the Prairie Pothole Region of North America, disturbances to wetlands that disrupt water-level fluctuations in response to wet–dry climatic conditions have the potential to alter natural vegetative communities in favor of species that proliferate in stable environments, such as cattail (Typha spp.). We evaluated the effect of water-level dynamics during a recent fluctuation in wet–dry conditions on cattail coverage within semipermanently and permanently ponded wetlands situated in watersheds with different land use and amounts of wetland drainage. We found that ponded water depth increase was significantly greater in wetlands where water levels were not near the spill point of the topographic basin, where banks were steeper, and in larger wetlands where past dry conditions had less influence on change in pond area. Proportion of the wetland covered by cattail was negatively correlated with increased water depth, bank slope and pond area. Our observations provide evidence that cattail coverage in prairie wetlands is regulated by water-level fluctuations and that land use surrounding the wetland might have an indirect effect on cattail coverage by altering water-level response to wet–dry climate conditions. For example, drainage of smaller wetlands into larger wetlands that are characterized by more permanent hydroperiods, leads to stabilized water levels near their spill point and is therefore a potential mechanism for increased cattail abundance in the northern prairie region.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
It has been suggested that the prevalence of monotypic stands of cattail (Typha spp.) has increased during the past century within wetlands in the Prairie Pothole Region of North America (Kantrud 1992; Swanson 1992; Anteau and Afton 2008). Such an increase has implications for abundance and quality of food resources and foraging habitat for waterbirds, particularly wading birds and dabbling waterfowl. When shallow-water zones become dominated by dense emergent vegetation, wetlands have less shallow-water foraging habitat, a reduction in or absence of submerged aquatic vegetation, and a reduction in the variety of decomposer organisms—including macroinvertebrates that are important food resources for waterbirds (Voigts 1976; Kantrud 1986). Dense stands of emergent vegetation within wetlands may eliminate foraging and roosting habitat needed by migrating and breeding wading birds and waterfowl (Weller and Spatcher 1965; Voigts 1976; Kantrud 1986; Anteau 2012). Landscape modifications may alter natural wetland hydrology, which may impact water-level fluctuations, sedimentation and nutrification, or community composition within wetlands (Galatowitsch et al. 2000; Gleason et al. 2003; Anteau 2012; Wiltermuth 2014; McCauley et al. 2015). Such changes might be responsible for greater cattail abundance and resultant degradation of many wetland ecosystems across North America (Galatowitsch et al. 1999). Therefore, understanding how landscape modifications have influenced the abundance of cattails—and the concomitant change in value of those wetlands to provide habitat for waterbirds—can inform ongoing conservation efforts aimed toward reversing the trend of increasing cattail encroachment in wetlands of the Prairie Pothole Region.
Water-level fluctuations in response to climate variability help shape vegetation community structure in prairie wetlands. Because the plant community composition shifts in response to wet–dry climate periods, the overall community structure is dominated by species adapted to dynamic hydrologic conditions (Stewart and Kantrud 1972; Kantrud et al. 1989; Grace and Wetzel 1981; LaBaugh et al. 1987; van der Valk 2005). But, if water-level fluctuations are reduced due to landscape modifications, then this mechanism for dynamics in vegetation community composition is eliminated and species adapted to more-stable water environments will proliferate (van der Valk and Davis 1980; Swanson 1992). Higher and more-stable water levels may alter vegetation communities, likely toward cattail-dominated systems where other conditions are suitable (Swanson 1992; Shay et al. 1999; Boers and Zedler 2008).
Disturbances within wetlands and their surrounding catchment that increase sedimentation and nutrient loading into wetlands or stabilize water levels may create conditions more favorable to cattail proliferation in prairie pothole wetlands (Wang et al. 1994; Gleason and Euliss 1998; Shay et al. 1999). Consolidation drainage—drainage of smaller wetlands into a larger wetland having a more-permanent hydroperiod (i.e., permanence of ponded water) within the same watershed—is a common practice in the Prairie Pothole Region to improve efficiency of agricultural production. Evidence from other studies has indicated consolidation drainage leads to higher and more stable water levels in wetlands receiving drainage water (Wiltermuth 2014; McCauley et al. 2015). Increased sedimentation rates in wetlands surrounded by cropland can decrease wetland depth through addition of nutrient-rich soil while reducing or eliminating competing wetland plant species by burial of seed banks (Jurik et al. 1994; Gleason and Euliss 1998). These conditions favor development of monotypic stands of cattail (Gleason and Euliss 1998; Galatowitsch et al. 1999; Shay et al. 1999). Further, additional nitrogen inputs from uplands can promote growth and expansion of cattail, especially where nitrogen is limiting (Shay et al. 1999). Moreover, stabilization of water levels can increase soil release of phosphorus and increase uptake by cattail resulting in enhanced growth (Boers and Zedler 2008). In addition to changes in vegetation structure, nutrient enrichment in wetlands can lead to changes in the macroinvertebrate community (Voigts 1976; McCormick et al. 2004); species that are best adapted to exploit available nutrients will gain a competitive advantage over those adapted for a more oligotrophic environment.
In the Prairie Pothole Region of North Dakota, cattail was previously found during 2002 to be present in 28 % of a sample of wetlands and averaged 37 % areal coverage within those wetlands (Ralston et al. 2007). It has been hypothesized that monotypic stands of cattail will increase in occurrence and coverage within wetlands where landscape modifications have altered wetland hydrology and upland land cover (Swanson 1992); therefore, there is a need to quantitatively evaluate factors that may be contributing to wetland conditions that are favorable to increased cattail coverage. Accordingly, we evaluated effects of watershed characteristics, wet and dry climate conditions, and land use on water depth in prairie wetlands having semipermanent and permanent hydroperiods. We then evaluated how water depth change, climate conditions, and land use influenced coverage of cattails in these wetlands.
Methods
Study area
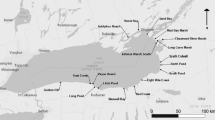
The Prairie Pothole Region in North America (Fig. 1) was shaped by the last glacial retreat (ca. 13,000 years ago) which created millions of depressional wetlands (Bluemle 1977). Hydrology of these wetlands is influenced by fluctuating wet–dry climatic conditions through a network of atmospheric, groundwater, and surface water connections. Thus, prairie pothole wetlands are dynamic ecosystems, and these dynamics drive high-levels of biological productivity that enables wetlands to provide habitat to various wildlife species (Kantrud et al. 1989).

North Dakota study area showing wetlands surveyed as dots within the shaded Prairie Pothole Region. Inset map of North America highlights North Dakota with a solid black outline and the entire Prairie Pothole Region is shaded
During April and May of 2011, we returned to 126 wetlands within the Prairie Pothole Region of North Dakota that were previously selected and surveyed once in 2004 or 2005 (hereafter, 2004/2005) by Anteau and Afton 2008). These wetlands were lacustrine semipermanently ponded and shallow-water permanently ponded wetlands (Cowardin et al. 1979). Wetlands were randomly selected within a hierarchal sampling design of three regions, three sub-regions, three clusters of three townships (one large region had an additional three clusters per sub-region), and three wetlands within each township (see Anteau and Afton 2008). For wetlands to be included in the 2004/2005 survey they must have had an open-water area larger than 120 m in diameter and a minimum ponded water depth of 0.25 m. In 2004/2005 as part of Anteau and Afton (2008), if reselection in the field was necessary (e.g., due to lack of open water), the nearest suitable wetland having a semipermanent hydroperiod was surveyed. Based on National Wetlands Inventory (NWI) classification (U.S. Fish and Wildlife Service 2003), 67 % of wetlands surveyed were semipermanently ponded, 3 % seasonally ponded, and the remainder were permanently ponded wetlands or shallow-water lakes. Those wetlands classified by NWI as seasonally ponded were wetlands reselected in the field during 2004/2005 that were more characteristic of semipermanently ponded wetlands at time of field assessment.
Because of the open water requirement of our initial wetland selection (Anteau and Afton 2008), our observations likely under represent cattail coverage across the region and should not be used for such estimation. However, our data set should provide a good basis for understanding cattail dynamics in the context of landscape modifications, with the exception of not containing information for wetlands with complete cattail coverage.
Spring wetland surveys and data preparation
During wetland surveys, we measured water depth, estimated emergent vegetation coverage and taxa composition, and classified land use surrounding each wetland. We used aerial imagery and digital elevation models to delineate watershed boundaries and to calculate pond area, topographic basin area, slope of the wetland bank, and area of drained wetlands. We used a time series of spatially modeled temperature and precipitation data from 1985 to 2010 to calculate a wetland-specific climate index.
Water depth
To estimate change in ponded water depth between surveys of 2004/2005 and surveys of 2011, we measured water depth to the nearest 0.1 m at 10 randomly selected survey locations estimated to be 10 m into open water from either the shoreline or the emergent vegetation to open-water interface during 2004/2005. When waves were present, we recorded the average depth between wave crests and troughs. As an estimate of the shoreline or emergent vegetation to open-water interface of each wetland in 2004/2005, we delineated these features using imagery (National Agricultural Imagery Program, U.S. Department of Agriculture; hereafter NAIP) acquired in the summer (July) prior those spring surveys. We used the imagery from the previous year because it better represents the location of the emergent vegetation to open-water interface used to locate their sampling locations. We digitized the interface by using a geographic information system (GIS) at a scale of 1:5000, and offset (i.e., buffered) the digitized open-water interface by 10 m in the direction of the wetland center to approximate locations of 2004/2005 sampling. We derived our 10 survey locations from the 10-m buffer, but constrained locations so there was a minimum distance of 40 m between locations. We navigated to survey locations that were pre-loaded onto a GIS data logger (Trimble GeoXT, Trimble Navigation, Sunnyvale, CA).
Emergent vegetation
We estimated the width of emergent vegetation coverage (0, >0–4, 5–10, 11–30, 31–60, or >60 m) and the proportion (recorded to the nearest 0.1) of emergent vegetation that was cattail at ten randomly selected transects that extended perpendicular from the shoreline through the survey locations where water depth measurements were taken to the center of the wetland. We calculated an average width of cattail coverage for each wetland using the midpoint value (0, 2, 7.5, 20.5, 45, 90 m respectively) from each of the 10 categorical estimates of emergent vegetation coverage multiplied by the proportion that was cattail. We selected the midpoint of the >60 m class to be at least twice as large as the preceding class. Finally, we calculated the area of cattail coverage within each wetland by using a GIS to create a polygon from the ponded water perimeter measured using 2010 imagery (see "Pond area" section) extending inward the distance of the average width of cattail coverage.
Land use
We classified current land use and land cover immediately surrounding each wetland using ground observations in 2011. We used GIS software in the field to attribute empty polygon outlines of land-cover data (Habitat and Population Evaluation Team 1996) based on current observations with one of the following categories: native grassland, restored grassland, unknown grassland, hayland, cropland, shrubs, trees, developed, barren, or wetland. We calculated the percent of cropland for the upland area that was both within the watershed (see "Watershed boundaries" section) and within 400 m of the focal wetland. We also estimated the width of any upland vegetation buffers (0, 1–4, 5–10, 11–50, >50 m) between any development or cropland use and the wetland. In our analyses we used the midpoint of categorical estimates, with maximum value of 75 m.
Pond area
We used pond area delineated for each surveyed wetland by McCauley et al. (2015) using photointerpretation of NAIP aerial imagery for 2003–2006 and 2009–2010. Photointerpretation was performed while images were viewed as panchromatic instead of true-color, because they were done as part of a larger study that involved some panchromatic imagery (see McCauley et al. 2015). Where the waterline was obscured by emergent vegetation, the waterline was approximated to be halfway between the emergent-vegetation to open-water interface and clearly identifiable upland. NAIP imagery was not available for 2007 and 2008. For those years we used pond area delineated once in either 2007 or 2008 using high-resolution digital elevation model (DEM; source data: 1 m pixel light detection and ranging [lidar]; Data available from the U.S. Geological Survey) or 1.25 m pixel interferometric synthetic aperture radar (InSAR) orthorectified image (Intermap Technologies, Inc., Englewood, Colorado).
Bank slope
We calculated the bank slope of each wetland because we expected the habitable area for emergent vegetation would be greater with less slope at the pond perimeter. We calculated bank slope using the change in elevation of ponded water divided by the change in average radius between wet and dry years (McCauley et al. 2015). We used 2007 or 2008 as the dry years and 2010 as the wet year; we substituted years if water levels were not higher in 2010 (≈5 %) due to reasons other than general wet conditions in the region, such as the wetland being part of a drainage network.
Watershed boundaries
We delineated boundaries for watersheds as the portion of the landscape where surface water flows into a focal wetland, and included intermittently-ponded wetlands that make up a watershed derived wetland complex (McCauley and Anteau 2014; McCauley et al. 2015). Wetland complexes were derived with our focal wetland at the terminus of the watershed. Watershed boundaries were delineated for wetlands using high resolution DEMs (3 m pixel lidar or 5 m pixel InSAR) and surface hydrology modeling tools (McCauley and Anteau 2014; McCauley et al. 2015). Some watersheds (46 %) that were large or highly irregular in shape were truncated to a 2.5 km maximum radius from the focal wetland for logistical reasons associated with assembling landscape variables, such as amount of wetland area drained. The 2.5-km radius encompassed >90 % of the total watershed area for 68 % of watersheds. Amount of landscape modification within these 2.5-km truncated watersheds should represent the total amount of modification in the full watershed, and the conditions nearest the wetland likely influence the wetland most. Therefore, we assumed the 2.5-km-truncated watershed provided a reasonable area to evaluate impacts of land use in wetlands that we surveyed and are an improvement over simply buffering around a wetland a set distance, a common practice (McCauley and Anteau 2014).
Wetland drainage
We used estimates of the percent watershed area that was previously drained wetland using multiple data sources, including: aerial photographs, DEM, NWI, and spatially explicit soil data (McCauley et al. 2015). Wetlands were identified as drained if they were present in historical photographs (dating back to 1937) but not present in current photographs, or if the wetlands were identified as part of a drainage network. Significant drainage occurred prior to 2003 in our study area; however, there was negligible evidence of additional drainage after 2003.
We also recorded presence of drainage networks and connections between wetlands while in the field in 2011. Surface-water connections included: inflow and outflow streams, ditches, culverts, and temporary connections between wetland basins due to flooding conditions.
Basin area
We defined a wetland basin as the topographic depression where ponded water is isolated from basins of other wetlands of an equal or more permanent hydroperiod. We measured basin area to determine the maximum pond area before the water would flow out of the basin (spill point). We used high-resolution DEMs (3 m pixel lidar or 5 m pixel InSAR) to find the spill point of the basin and then delineated the area at that elevation. We calculated the percent of the basin that was filled in 2004/2005 based on pond area.
Wetland-specific climate index
We used a fine-scale climate-based index (2.5 arc-minute) to estimate the spatially-explicit effect of wet–dry conditions on increase in water depth in wetlands. To create this index, we calculated a standardized precipitation-evapotranspiration index (SPEI; Vicente-Serrano et al. 2010) using monthly averaged PRISM (Parameter-elevation Regression on Independent Slopes Model) Climate Group data (PRISM Climate Group 2002) from 1895 to 2010. In SPEI, positive values represent wet conditions and negative values represent dry conditions standardized to the entire range of data. We then calculated wetland-specific climate index values by averaging SPEI values from the past 72 months, starting from the month imagery used for pond area delineation was acquired. We used the 72-month time period because in preliminary analyses this duration best explained changes in pond area during 2003–2010 (M. Post van der Burg personal communication). Using the annual averages of wetland-specific climate index values, we generalized 2003–2005 as wet conditions, 2006–2008 as dry conditions and 2009–2010 as wet conditions (Fig. 2). In our analysis, we calculated the difference in wetland-specific climate index values from the wet conditions during 2003–2005 and wet conditions in 2009–2010.

Wetland-specific climate index averaged (95 % CI) among all wetlands surveyed for each year 2003–2010. The index was calculated using spatially modeled temperature and precipitation from 1895 to 2010 that was standardized and then averaged at each wetland over the past 72 months. Positive values indicated wet conditions and negative values indicate dry conditions.eps
Statistical analysis
Water depth change
We examined the potential influence of climate, upland disturbance and wetland connectivity on change in ponded water depth using mixed-effects regression analysis (Package lme4 in R Statistical Computing Environment; R Development Core Team 2010; Bates et al. 2011). Our dependent variable was water depth change from 2004/2005 to 2011 in meters. We created an a priori model that included independent variables accounting for change in wetland-specific climate index, proportion watershed area that was drained wetland, presence or absence of wetland surface connections, proportion surrounding upland that was cropland, basin full proportion, bank slope (natural logarithm transformed, hereafter “log”), pond area (log) in 2010, and proportional change in pond area from wet conditions (2003–2005) to dry conditions (2006–2008). We found no evidence of strong correlations among independent variables (\(|r| \le 0.24\)). We also included interaction terms for climate-index-difference-by-pond-area because climate variability likely has a greater influence on water depth in terminal wetlands that were smaller rather than larger in pond area, for wetland-drainage-by-proportion-basin-full because water depth within ponds that already filled their basins would be less influenced by drainage within the watershed than would water depth in ponds that have potential to increase, and for pond-area-change-by-pond-area to account for differences in effect of proportional change in pond area that vary with pond area. We specified township surveyed as a random error term to account for residual error attributable to spatial clustering within our sampling design. We evaluated significance of variables within the model using \(\alpha = 0.05\) and the variance explained by the model using marginal and conditional coefficients (Nakagawa and Schielzeth 2013). Variance in fixed effects only is indicated in the marginal coefficient, while variance in both the fixed and random effects is indicated in conditional coefficient. There was insufficient evidence in our data to support the interactions of climate-index-by-pond-area (t-value = 1.344, df = 116, P-value = 0.182) and drainage-by-proportion-basin-full (t-value = 0.243, df = 116, P-value = 0.778); therefore, we removed those interactions to interpret their main effects and we kept the pond-area-change-by-pond-area interaction in our final model.
Cattail coverage
We evaluated how coverage of cattail within wetlands during 2011 may have been influenced by the change in water depth from 2004/2005 to 2011, rate of change in pond area during 2003–2005 to 2006–2008, wetland drainage within the watershed, and proportion of upland that is cropland using a linear model (R Development Core Team 2010). We used an arc-sine square root transformation of the proportion wetland area covered by cattail as the response variable in our model. In our a priori model we also included the effects of pond area (log), proportion percent of the topographic basin filled by ponded water, and bank slope (log) as independent variables, and we included a cropland-by-upland-buffer-width (log+1 of width) interaction term because an upland vegetation buffer can mitigate the effects of surrounding land use on cattail coverage (Castelle et al. 1994; Anteau and Afton 2008; Anteau et al. 2011). We found no evidence of strong correlations among independent variables (\(|r| \le 0.26\)), with the exception of amount of surrounding cropland and width of the upland buffer (\(r = -0.65\)) for which the interaction term was included. We did not include the random error term for township surveyed in this model because we found negligible evidence for its influence (1 % of residual error) in a preliminary analysis of this model (using Package lme4 in R Statistical Computing Environment; (R Development Core Team 2010; Bates et al. 2011). We used \(\alpha = 0.05\) to evaluate significance of variables within the model and \(R^2\) to evaluate variance explained by the model.
There was insufficient evidence in our data to support the interaction of cropland-by-upland-buffer-width in our model (t-value = 1.259, df = 116, P-value = 0.211); therefore, we removed the interaction term from our model to evaluate each main effect formerly included in the interaction. We evaluated cropland and upland vegetation buffer width (log+1) in two separate models that included all other independent variables, but not together in a single model due to their correlation, and selected to interpret the model with greater variation explained. We plotted back-transformed model predicted effects and 95 % confidence intervals across the observed range of variable of interest and held other variables at median levels.
Results
Within the 126 wetlands we surveyed, the average pond area was 68 ha (median \(=\) 26 ha, range \(=\) 1–965 ha, SD \(=\) 132) as measured from 2010 mid-summer imagery. During 2006–2008 under dry conditions, average pond area proportional change was −0.22 (median \(=\) −0.17, range \(=\) −0.98 to 0.50, SD \(=\) 0.23). During spring 2011, maximum water depth within wetlands averaged 2.0 m (SD \(=\) 0.7), and on average, water depth increased 1.1 m (SE \(=\) 0.1) from 2004/2005 to 2011 (Fig. 3). We detected cattail on at least one transect in 64 wetlands (i.e., 51 % of wetlands surveyed). Mean coverage percent of wetland area was 18 % (SD \(=\) 2 %) and coverage by other species of emergent vegetation was 2 % (SD \(=\) 7 %).

Histogram showing change in water depth from 2004 or 2005 to 2011 within 126 wetlands with more-permanent hydroperiods in North Dakota
Our model explained 29 % (marginal) and 50 % (conditional) of the variation in water depth change from 2004/2005 to 2011. Water depth increased more where ponds filled less of the topographic wetland basin, where ponds were larger and decreased in area less during dry conditions, had steeper bank slopes, and where conditions were wetter indicated by the wetland-specific climate index (Table 1). Model predictions indicated mean water depth increases were 1.5 to 1.0 m in wetland basins that were 5 and 100 % full, respectively (Table 2)—ponded water in full basins can merge with adjacent ponds to become one larger, deeper pond. There was insufficient evidence in our data to indicate a direct relationship of increased water depth to identified surface connections, surrounding cropland, and wetland drainage in upper-portions of the watershed. Spatial clustering represented by the surveyed township random error term accounted for 30 % residual error remaining after fixed effects were fitted in the model.
Our model explained 37 % (\(R^2\)) of the variation observed in cattail coverage. Proportion of wetlands covered by cattail was strong-negatively correlated with increased water depth (\(\hat{\beta }= -0.151 \pm 0.042\) SE; Fig. 4). Cattail coverage was also negatively correlated with bank slope and pond area (Table 3). However, there was insufficient evidence in our data to indicate a direct relationship of cattail coverage in wetlands to wetland drainage in upper-portions of the watershed, width of upland vegetation buffers, fullness of the topographic basin, and drying phase pond area dynamics.

Model estimated effect (95 % CI) of change in water depth on cattail coverage within more-permanent hydroperiod wetlands in North Dakota. Estimates are plotted back-transformed from the arc-sine square root transformation of proportion cattail coverage in wetlands and all other model variables were held at median values
Discussion
Our results show that water depth in northern prairie wetlands was greater during the recent wetting conditions in 2011 than at the end of the previous wet conditions in 2005. However, increases in water depth were less where ponds had already filled, or nearly filled the topographic wetland basin, indicating water-level stabilization. Additionally, we observed a strong correlation between increased water depth and less cattail coverage, suggesting cattail was eliminated where water depth increased. This process of cattail elimination by increased water levels during wet periods is consistent with observations from other studies that suggest >1 m increase in water depth for as little as two years may be sufficient to eliminate most emergent vegetation (van der Valk 1994). Evidence from other research suggests that drainage of wetlands in upper-portions of a watershed causes water levels in wetlands positioned lower in a watershed to progressively increase with each fluctuation of wet–dry conditions until those ponds fill their topographic wetland basin and then water levels stabilize (Wiltermuth 2014). It is then possible, in a wetland under those hydrologic conditions, that cattail coverage is minimized by increases in water level resulting from consolidation drainage; but, ultimately cattail coverage can potentially increase as water levels stabilize at their basin spill elevation.
Wetlands that cycle between increased ponding and drawdown conditions of generally equal amplitude should exhibit natural vegetative cycles with seed germination during drawdown, growth during ponding, and die-off during high-water periods (Weller and Spatcher 1965; van der Valk and Davis 1978). However, where consolidation drainage has resulted in higher and more-stable water levels in remaining wetlands, cattail can become established at the newly formed shallow water fringes of the wetland while high water levels persist. Cattail is not likely to be eliminated by drowning within wetlands where ponds have filled their topographic basin because these wetlands are already at their spill-point, and any additional inflow will contribute to a larger watershed. However, expansion of cattail toward the center of the wetland can occur by extension of the habitable zone as a result of sedimentation and accretion of organic debris (Waters and Shay 1992; Gleason and Euliss 1998; Shay et al. 1999). Further, there is evidence that persistent high-water conditions could cause soil slumping on back slope topography when soils along the wetland fringe become saturated. Accumulation of sediment and organic debris increases the rate of habitable zone expansion, ultimately reducing the water depth required to reach the basin spill point elevation. Wetlands in landscapes with high agricultural cropland use are likely to have greater rates of sedimentation and nutrient input (Gleason and Euliss 1998), than those surrounded by grasslands, making those wetlands prime environments for cattail.
Based on evidence from other studies (Wiltermuth 2014; McCauley et al. 2015), consolidation drainage appears to have more of an indirect effect on increased cattail coverage in wetlands through stabilization of water levels that promote cattail habitat, rather than a direct effect, such as increase in nutrients. Consolidation drainage, however, may actually temporarily suppress cattail coverage in wetlands with ponds that have not yet filled their topographic basin, because during each climatic wetting phase water levels progressively increase (Wiltermuth 2014) to overtop (i.e., inundate) existing cattail stands—essentially drowning them out. However, this could only be a temporary response, until a pond fills the topographic wetland basin and water levels subsequently stabilize. Once water-levels stabilized, cattail coverage is likely to increase within wetlands where consolidation drainage has occurred.
Growing evidence suggests that landscape modifications may be resulting in an elimination of wetlands with functional semipermanent hydroperiods. For example, consolidation drainage that alters hydrology within wetlands receiving water is resulting in wetlands that are larger and fluctuate less in response to climate variability (Wiltermuth 2014; McCauley et al. 2015). We suggest that there are two likely functional fates for semipermanently ponded wetlands in landscapes that contain consolidation drainage and the fate realized is determined by the topographic shape of the basin (i.e., potential maximum water depth). Consolidating water into wetlands with deep basins will result in deep water bodies that are more-stable and function more like lakes. In some landscapes, these water bodies will increase in size and merge with neighboring basins to have open-water areas where water depth is too great for emergent and submerged vegetation to grow (>3 m). Littoral communities will be limited to the periphery of the water body, thus reducing the biological productivity to wetland area ratio compared to that of the former semipermanently ponded wetland—because secondary productivity is generally greater in littoral zones predominately containing submerged vegetation compared to limnetic zones. The alternative fate occurs in shallow wetland basins when water levels fill to the spill elevation and stabilize, thereby creating a stable shallow-water environment conducive to cattail encroachment. In this situation, sediment and cattail leaf litter accumulation can further facilitate cattail coverage across the wetland until the wetland is completely covered; facilitation occurs by means of reducing depth and increasing sediment nutrient availability. Artificially lowering the spill-point elevation of a wetland via a ditch that drains water to reduce the extent of the wetland can also create stable shallow-water environments that are conducive to becoming completely covered by cattail. Invertebrate productivity within zones of dense cattail is less compared to zones predominately containing submerged aquatic vegetation (see Voigts 1976; Rose and Crumpton 1996). Therefore, when stable shallow-water zones result from consolidation drainage and cattail cover correspondingly increases, there is a reduction in secondary productivity to wetland area. The potential for consolidation drainage and other landscape modifications to eliminate functional semipermanently ponded wetlands can have consequences on biological productivity and diversity, and available habitat for waterbirds in wetlands throughout the Prairie Pothole Region (Wiltermuth 2014).
In this study we have focused on landscape modifications and surface water because we were not able to incorporate groundwater levels into our study. We suspect that groundwater interactions could account for variability that we could not (Sloan 1994; Winter and Rosenberry 1995). Specifically how prairie wetland hydrology responds temporally to wet–dry climate dynamics of various magnitudes because antecedent groundwater levels may attenuate or delay surface water responses to short term precipitation and evaporation differences. Ultimately better accounting for water-level dynamics will contribute to further understanding proliferation of cattail in the northern prairie region.
Our observations provide evidence that cattail cover decreased with higher water levels and that water-level changes were influenced by wet–dry conditions and basin characteristics. Landscape modifications that have altered hydrologic processes within wetlands may have the potential to alter natural vegetative cycles, resulting in increased coverage of cattail in monotypic stands where water-levels are stabilized rather than more diverse plant communities interspersed within wetlands. This change can negatively impact invertebrate forage and waterbird populations that feed on aquatic invertebrates. Restoration of more natural water-level fluctuations in response to wet–dry climate periods, perhaps through restoration of complexes of wetlands having various hydroperiods, may restore vegetative cycles and transition monotypic stands of cattail back into vegetative communities that vary in wetland coverage according to hydrologic conditions.
References
Anteau MJ (2012) Do interactions of land use and climate affect productivity of waterbirds and prairie-pothole wetlands? Wetlands 32(1):1–9. doi:10.1007/s13157-011-0206-3
Anteau MJ, Afton AD (2008) Amphipod densities and indices of wetland quality across the upper-Midwest, USA. Wetlands 28(1):184–196. doi:10.1672/07-53.1
Anteau MJ, Afton AD, Anteau ACE, Moser EB (2011) Fish and land use influence Gammarus lacustris and Hyalella azteca (Amphipoda) densities in large wetlands across the upper Midwest. Hydrobiologia 664(1):69–80. doi:10.1007/s10750-010-0583-2
Bates D, Maechler M, Bolker B (2011) lme4: linear mixed-effects models using S4 classes. R package version 0.999375-42. http://CRAN.R-project.org/package=lme4/
Bluemle JP (1977) The face of North Dakota: the geologic story. Educational Series 11, North Dakota Geological Survey
Boers AM, Zedler JB (2008) Stabilized water levels and Typha invasiveness. Wetlands 28(3):676–685. doi:10.1672/07-223.1
Castelle AJ, Johnson AW, Conolly C (1994) Wetland and stream buffer size requirements: a review. J Environ Qual 23(5):878–882. doi:10.2134/jeq1994.00472425002300050004x
Cowardin LM, Carter V, Golet FC, LaRoe ET (1979) Classification of wetlands and deepwater habitats of the United States. FWS/OBS 79/31, U.S. Fish and Wildlife Service, Washington, DC. http://pubs.er.usgs.gov/publication/2000109
Galatowitsch S, Whited D, Lehtinen R, Husveth J, Schik K (2000) The vegetation of wet meadows in relation to their land-use. Environ Monit Assess 60(2):121–144. doi:10.1023/a:1006159028274
Galatowitsch SM, Anderson NO, Ascher PD (1999) Invasiveness in wetland plants in temperate North America. Wetlands 19(4):733–755. doi:10.1007/BF03161781
Gleason RA, Euliss NH Jr (1998) Sedimentation of prairie wetlands. Great Plains Res 8:97–112. http://digitalcommons.unl.edu/greatplainsresearch/363/
Gleason RA, Euliss NH Jr, Hubbard DE, Duffy WG (2003) Effects of sediment load on emergence of aquatic invertebrates and plants from wetland soil egg and seed banks. Wetlands 23(1):26–34. doi:10.1672/0277-5212(2003)023[0026:eosloe]2.0.co;2
Grace JB, Wetzel RG (1981) Habitat partitioning and competitive displacement in cattails (Typha): experimental field studies. Am Nat 118(4):463–474. doi:10.2307/2460778
Habitat and Population Evaluation Team (1996) U.S. Fish and Wildlife Service 1996 landcover: U.S. Fish and Wildlife Service, Bismarck
Jurik TW, Wang SC, van der Valk AG (1994) Effects of sediment load on seedling emergence from wetland seed banks. Wetlands 14(3):159–165. doi:10.1007/BF03160652
Kantrud HA (1986) Effects of vegetation manipulation on breeding waterfowl in prairie wetlandsa literature review. Fish and Wildlife Technical Report 3, U.S. Fish and Wildlife Service, Washington, DC. http://pubs.er.usgs.gov/publication/2000113
Kantrud HA (1992) History of cattails on the prairies: wildlife impacts. In: Linz GM (ed) Proceedings of the cattail management symposium. U.S. Department of Agriculture Denver Wildlife Research Center and U.S. Fish and Wildlife Service, North Dakota State University, Fargo, pp 9–12
Kantrud HA, Krapu GL, Swanson GA (1989) Prairie basin wetlands of the Dakotas: a community profile. Biological Report 85(7.28), U.S. Fish and Wildlife Service, Washington, DC. http://pubs.er.usgs.gov/publication/2000127
LaBaugh JW, Winter TC, Adomaitis VA, Swanson GA (1987) Hydrology and chemistry of selected prairie wetlands in the Cottonwood Lake area, Stutsman County, North Dakota, 197982. Professional Paper 1431, U.S. Geological Survey, Washington, DC. http://pubs.er.usgs.gov/publication/pp1431
McCauley LA, Anteau MJ (2014) Generating nested wetland catchments with readily-available digital elevation data may improve evaluations of land-use change on wetlands. Wetlands 34(6):1123–1132. doi:10.1007/s13157-014-0571-9
McCauley LA, Anteau MJ, Post van der Burg M, Wiltermuth MT (2015) Land use and wetland draiange affect water levels and dynamics of remaining wetlands. Ecosphere 6(6):art92. doi: 10.1890/ES14-00494.1
McCormick PV, Shuford RBE, Rawlik PS (2004) Changes in macroinvertebrate community structure and function along a phosphorus gradient in the Florida Everglades. Hydrobiologia 529(1):113–132. doi:10.1007/S10750-004-5737-7
Nakagawa S, Schielzeth H (2013) A general and simple method for obtaining \(R^2\) from generalized linear mixed-effects models. Methods Ecol Evol 4(2):133–142. doi:10.1111/j.2041-210x.2012.00261.x
PRISM Climate Group (2002) Parameter-elevation regressions on independent slopes model. Oregon State University. http://prism.oregonstate.edu/
R Development Core Team (2010) R: a language and environment for statistical computing. http://www.R-project.org/
Ralston ST, Linz GM, Bleier WJ, Homan HJ (2007) Cattail distribution and abundance in North Dakota. J Aquat Plant Manag 45(1):21–24. http://digitalcommons.unl.edu/icwdm_usdanwrc/708
Rose C, Crumpton WG (1996) Effects of emergent macrophytes on dissolved oxygen dynamics in a prairie pothole wetland. Wetlands 16(4):495–502. doi:10.1007/BF03161339
Shay JM, de Geus PMJ, Kapinga MRM (1999) Changes in shoreline vegetation over a 50-year period in the Delta Marsh, Manitoba in response to water levels. Wetlands 19(2):413–425. doi:10.1007/bf03161773
Sloan CE (1994) Ground-water hydrology of prairie potholes in North Dakota. US Government Printing Office, Washington, DC
Stewart RE, Kantrud HA (1972) Vegetation of prairie potholes in North Dakota, in relation to quality of water and other environmental factors. Professional Paper 585-D, U.S. Geological Survey, Washington, DC. http://pubs.er.usgs.gov/publication/pp585D
Swanson GA (1992) Cycles of cattails in individual wetlands: environmental influences. In: Linz GM (ed) Proceedings of the cattail management symposium. Denver Wildlife Research Center and U.S. Fish and Wildlife, Fargo, pp 13–19
US Fish and Wildlife Service (2003) National wetlands inventory. U.S. Department of the Interior, Fish and Wildlife Service, Washington, DC. http://www.fws.gov/wetlands/
van der Valk AG (1994) Effects of prolonged flooding on the distribution and biomass of emergent species along a freshwater wetland coenocline. Vegetatio 110(2):185–196. doi:10.1007/bf00033398
van der Valk AG (2005) Water-level fluctuations in North American prairie wetlands. Hydrobiologia 539(1):171–188. doi:10.1007/s10750-004-4866-3
van der Valk AG (1978) The role of seed bands in the vegetation dynamics of prairie glacial marshes. Ecology 59(2):322–335. doi:10.2307/1936377
van der Valk AG, Davis CB (1980) The impact of a natural drawdown on the growth of four emergent species in a prairie glacial marsh. Aquat Bot 9:301–322. doi:10.1016/0304-3770(80)90033-9
Vicente-Serrano SM, Beguera S, Lpez-Moreno JI (2010) A multiscalar drought index sensitive to global warming: the standardized precipitation evapotranspiration index. J Clim 23(7):1696–1718. doi:10.1175/2009jcli2909.1
Voigts DK (1976) Aquatic invertebrate abundance in relation to changing marsh vegetation. Am Midl Nat 95(2):313–322. doi:10.2307/2424396
Wang SC, Jurik TW, van der Valk AG (1994) Effects of sediment load on various stages in the life and death of cattail (Typha glauca). Wetlands 14(3):166–173. doi:10.1007/BF03160653
Waters I, Shay JM (1992) Effect of water depth on population parameters of a Typha glauca stand. Can J Bot 70(2):349–351. doi:10.1139/b92-046
Weller MW, Spatcher CS (1965) Role of habitat in the distribution and abundance of marsh birds. Special Report 43, Iowa Agriculture and Home Economics Experiment Station, Ames
Wiltermuth MT (2014) Influences of climate variability and landscape modifications on water dynamics, community structure, and amphipod populations in large prairie wetlands: Implications for waterbird conservation. Ph.D. thesis, North Dakota State University. http://search.proquest.com/docview/1650252117
Winter TC, Rosenberry DO (1995) The interaction of ground water with prairie pothole wetlands in the Cottonwood Lake area, east-central North Dakota, 1979–1990. Wetlands 15(3):193–211. doi:10.1007/bf03160700
Acknowledgments
Funding for this research was provided by: North Dakota Department of Game and Fish—through the State Wildlife Grant; Plains and Prairie Pothole Landscape Cooperative, Ducks Unlimited-Great Plains Regional Office, Dr. Bruce D. J. Batt Fellowship in Waterfowl Conservation granted by the Institute for Wetland and Waterfowl Research of Ducks Unlimited Canada, North Dakota State University, and the U.S. Geological Survey. We thank J. Bivens, J. Coulter, A. Lawton, L. McCauley, J. McClinton, P. Mockus, S. Paycer, J. H. Pridgen, A. Smith, N. Smith, and M. M. Weegman for assisting with wetland surveys, or GIS work. We also thank U.S. Fish and Wildlife service Refuge system and Water Management Districts in North Dakota for logistical support, as well as numerous private landowners. We appreciate the helpful comments provided on previous versions of this manuscript provided by M.G. Butler, M.E. Clark, E.S. DeKeyser, R.G. Finocchiaro, and two anonymous reviewers. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Wiltermuth, M.T., Anteau, M.J. Is consolidation drainage an indirect mechanism for increased abundance of cattail in northern prairie wetlands?. Wetlands Ecol Manage 24, 533–544 (2016). https://doi.org/10.1007/s11273-016-9485-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-016-9485-z