
Abstract
Copper (Cu) is a micronutrient essential for plant development. However, in excess, it is toxic to plants and may cause various physiological and morphological changes. The study of the growth of plants exposed to excess Cu is important for the development of phytoremediation programs and for understanding the mechanisms involved in the tolerance of this metal. In this context, the objective of this research was to evaluate the effect of excess copper on photosynthetic responses and root morphology of Hymenaea courbaril L. Biometric measurements, gas exchange, root morphology, and Cu content in tissues and indices (TI and TF) were assessed, involving metal content and biomass. Up to a concentration of 200 mg kg−1, Cu favored growth, gas exchange, and root morphology of the plants under study. At a higher concentration (800 mg kg−1) in the soil, it affected plant growth and caused a decrease in photosynthetic rate. Biochemical limitations in photosynthesis were observed, as well as lower maximum net photosynthetic rate (Amax), respiration rate in the dark (Rd), light compensation point (LCP), light saturation point (LSP), and apparent quantum yield (α), when exposed to excess Cu. Root length, surface area, mean diameter, root volume, dry biomass, and specific root length decreased with high Cu concentrations in the soil. Cu was accumulated in the roots as a mechanism of tolerance to the excess of this metal in order to preserve the most metabolically active tissues present in the leaves. At a concentration of 800 mg kg−1, copper also caused inhibition of the root system. Plants of H. courbaril showed tolerance to excess Cu in the soil and can be indicated for the recovery of areas contaminated with this metal.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Copper (Cu) is a micronutrient essential for plant development. However, in excess in the soil, it becomes phytotoxic to plants, interfering in innumerable physiological processes (Yruela 2013; Küpper and Andresen 2016). The synthesis of photosynthetic pigments, cell membrane permeability, homeostasis, and plant nutrient balance is altered at high Cu concentrations (Adrees et al. 2015; Gautam et al. 2016). Excess Cu ions in plants cause disturbances in photosynthesis, which can result in oxidative stress, with increasing reactive oxygen species (ROS), reducing carbon fixation, and reducing or inhibiting plant development (Küpper and Andresen 2016). Exposure to Cu stress leads to remodeling in root morphology (Cai et al. 2014), since the root is the first organ to come into contact with the contaminant. The use of digital images of roots is an important parameter for analysis of root development. However, it is worth mentioning that there are few studies on the root morphology of native species and this often occurs due to the difficulty in analyzing this underground organ. High copper concentrations reflect on a reduction in root biomass, as well as in shoot biomass (Feigl et al. 2013).
The knowledge of physiological, biochemical, and morphoanatomic responses of plants, when exposed to heavy metals, is of great importance for the recovery of contaminated areas. Phytoremediation and the physiological processes involved have aroused worldwide interest (Ent et al. 2013), but morphological and physiological responses are still poorly understood in forest species. The use of trees in phytoremediation programs has advantages, once the metal remains immobilized in plant tissues for a longer period and the roots present a barrier against contaminant leaching. The species Hymenaea courbaril L. (Caesalpinioideae) is native to Brazil and has qualities in the recovery of degraded areas due to the wide distribution of this species and the tolerance and adaptation to a variety of environments. However, its tolerance and mechanisms of adaptation to copper stress are not yet known. In this context, the objective of this research was to evaluate the effect of excess copper on photosynthetic responses and root morphology of Hymenaea courbaril L.
2 Material and Methods
2.1 Experimental Conditions
The experiment was conducted at the Institute of Agricultural Sciences of Universidade José do Rosário Vellano (UNIFENAS), in Alfenas, MG, Brazil, located at 21° 25′ 45″ S, 45° 56′ 50″ W and altitude of 881 m. The experimental design was in randomized blocks (DBC), consisting of five copper concentrations (CuSO4·5H2O), 0, 100, 200, 400, and 800 mg kg−1, and four replicates. The plant material consisted of Jatobá—Hymenaea courbaril L.—plants, with an approximate age of 120 days. The sowing was accomplished in tubes with capacity of 180 cm3, filled with commercial substrate Plantmax® and basic fertilization composed of 3 kg m−3 of Yorim Master® (16% P2O5, 18% Ca, 7% Mg). Three side dressing fertilizations were carried out at 45, 60, and 90 days after sowing with: 20 g of N and 15 g of K2O, respectively, with urea and potassium chloride as the source, which were dissolved in 10 l of water and applied 10 ml solution per plant.
In the experiment, subsurface soil was used, with the chemical characterization described in Table 1. The substrate used consisted of 360 dm3 soil, supplemented with 3.6 kg earthworm humus, 160 g limestone, and 230 g simple superphosphate. Plants of H. courbaril were transferred to pots (18 dm3), kept in a greenhouse with a plastic cover (150 μm), and covered on the sides with a screen (sombrite® 50%). During the experiment, soil moisture was controlled every 2 days, by weighing the pots and replacing the water, to maintain soil moisture at 70% retention capacity. Thirty days after transplanting seedlings, copper concentrations were applied to the soil surface via solution. Sixty days after copper application, nitrogen fertilization was carried out at a dose of 10 mg kg−1 N, in the form of urea. The experiment occurred from December 2014 to May 2015.
2.2 Biometric Measurements
Six measurements were made for each treatment of height (H) and stem diameter (SD). Measurements were established using a digital caliper and a tape measure at 20, 40, 60, 80, 100, and 120 days of cultivation.
2.3 Gas Exchange
Gas exchange parameters were measured using a portable photosynthesis system (IRGA, LI-6400XT, Li-Color, Lincoln, NE, USA) and a chamber with artificial light source (LI-6400-02B RedBlue) at 20, 40, 60, 80, 100, and 120 days of cultivation after copper application to the soil. Spot measurements were performed in the morning from 9.00 to 11.00 a.m. on a fully expanded leaf in the fourth node counting from the apical region. The parameters evaluated were leaf photosynthesis rate (A), stomatal conductance (gs), transpiration (E), concentration of intercellular CO2 (Ci), water use efficiency (A/gs), and carboxylation efficiency (A/Ci). The measurements were performed on a leaf area of 6 cm2, and the airflow in the chamber was at a CO2 concentration of 380 mmol mol−1. The air was collected from outside the greenhouse, transported to a gallon buffer tank, and then pumped into the chamber. A photon flux density (PPFD) of 1000 μmol m−2 s−1 was used, with blue-red LED light source, and the chamber temperature was 28 °C.
The curves of the net photosynthetic rate (A) in response to the flux of photosynthetically active radiation (PAR) were determined by the same portable photosynthesis system. All measurements were made on the same leaf used for spot measurements in three plants per treatment. The PAR used were 1400, 1000, 800, 200, 100, 50, 25, and 0 μmol m−2 s−1 for 5 min at 28 °C and at ambient CO2 conditions (approximately 380 μmol mol−1). The curves were made at four different times 30, 60, 90, and 120 days of cultivation. Data referring to the curves were adjusted to the rectangular hyperbolic function: A = a + [(Amax × PAR)/(b + PAR)], where Amax is the maximum net photosynthetic rate and a and b are coefficients of the equation. Through the curve, maximum net photosynthetic rate (Amax) was obtained, as well as respiration rate in the dark (Rd), measured in leaves adapted to the dark, light compensation point (LCP), and light saturation point (LSP). Apparent quantum yield (α) was estimated by linear regression of the initial region of the curve (0 ≤ PAR ≤ 200 μmol m−2 s−1), where A = c + (α × PAR), and c and α are adjustment coefficients.
2.4 Root Morphology
For root morphology, three roots were used per treatment at the end of the experiment. After the washing process, the plants were separated into root system and shoot, at the stem-root region. For the morphology analysis of the root system, the image analysis system WinRHIZO Pro 2007a (Regent Instruments, Sainte-Foy, QC, Canada) was used, coupled to a professional scanner (Epson, Expression 10000 XL, Epson America, Inc., Long Beach, CA, USA), equipped with an additional photon unit (TPU). The procedures for obtaining the images were made according to Souza et al. (2012a). The following characteristics were determined: root length (RL) (cm), root surface area (SA) (cm2), root mean diameter (RMD) (mm), and root volume (RV) (cm3). Root length was also analyzed through diameter classes (0 to 4.5 mm) by the same software. The roots were then stored in paper bags and transported to a forced circulation oven at 65 °C until a constant mass was obtained. Other attributes involving morphological and dry matter data were the following: ratio between root dry matter and shoot dry matter (RDM/SDM, g g−1), specific root length (SRL, cm g−1), root fineness (RF, cm cm3), and root tissue density (RMDe, g cm3).
2.5 Leaf and Root Anatomy
Two samples were collected in the region of the middle third of the first fully expanded leaf (from the apex) for leaf anatomy. Each sample consisted of a leaflet. The samples were fixed in solution of formaldehyde, acetic acid, and 90% ethanol (FAA 90); paradermal sections were cut according to Souza et al. (2012a), and cross sections of mesophyll and midrib were cut according to Souza et al. (2010).
Sections were photographed under an Olympus BX-60 light optical microscope coupled to a digital camera. The parameter measured in the paradermal sections of the leaf was stomatal index (SI): [(number of stomata/mm2)/number of epidermal cells] × 100]. The parameters evaluated for the cross section of the midrib were as follows: xylem vessel diameter and area of perivascular fibers. For the leaf limb, the following parameters were measured: cuticle thickness of adaxial and abaxial surfaces. Evaluations in the leaf blade were performed 549 μm (μm) away from the midrib. This distance was chosen based on the position of the secondary leaf ribs. For root anatomy, a root sample (with all regions) was collected, two plants/treatments/replications. Fixation and cross sections were performed according to Souza et al. (2009) and were photographed under an Olympus BX-60 light optical microscope, coupled to a digital camera. The following parameters were analyzed in the root: vascular cylinder area and cortex thickness. All these measurements were made by the image analysis program UTHSCSA ImageTool (University of Texas, San Antonio, USA) (Souza et al. 2009, 2010).
2.6 Cu Content in Tissues and Indexes
The dry matter of roots, stems, and leaves was ground in a Wiley mill and, with plant tissue samples, the determination of Cu contents by atomic absorption spectroscopy was performed, according to Carmo et al. (2000).
To evaluate the tolerance of H. courbaril plants and the ability to accumulate copper, tolerance index and translocation factor were calculated. The tolerance index (TI) is based on biomass production (Shi et al. 2011) and was used to evaluate the tolerance of this tree for each Cu concentration. The index was calculated as follows: TI = Bt/Bc, where Bt (g/plant) is the biomass of plants cultivated in soils with Cu and Bc (g/plant) is the biomass of control plants.
The translocation factor (TF) indicates the ability of the plant to translocate metals from the roots to the shoot, calculated as: TF = As/Ar × 100, where As (mg kg−1) is the total Cu accumulated in the shoot and Ar (mg kg−1) is the total Cu accumulated in the roots (Shi et al. 2011).
2.7 Data Analysis
For all analyzed parameters, the means and ± standard error (SE) were calculated. For the statistical analysis of the results, when significant, linear and non-linear regression analyses were used; the analysis of variance ANOVA and the Skott-Knott test were performed, using the software Sisvar version 4.3 (Universidade Federal de Lavras, Lavras, Brazil).
3 Results and Discussion
3.1 Biometric Measurements
In all treatments with copper (Cu), the plants of H. courbaril maintained linear growth in height (H) and stem diameter (SD) (p < 0.05) (Fig. 1a, b), with higher growth at concentrations of 0, 100, and 200 mg kg−1. However, at concentrations of 400 and 800 mg kg−1, it is observed that Cu negatively affected the height of the plants (H), as well as stem diameter (SD) at the highest concentration (800 mg kg−1) (p < 0.05) (Fig. 1a, b).

Height and stem diameter of H. courbaril exposed to different Cu concentrations. Regressions with asterisk correspond to p ≤ 0.05% probability. Statistical difference by the Scott-Knott test at 5% probability between Cu concentrations at 120 days of cultivation. Means followed by the same letter for treatments at 120 days of cultivation did not differ by the Skott-Knott test at 5% probability (p ≤ 0.05). Each value indicates treatment means ± SE
Up to a concentration of 200 mg kg−1, copper favored growth, gas exchange and root morphology of H. courbaril plants, possibly due to the fact that it is a micronutrient, within the limits favorable to plant development (Souza et al. 2014). However, in excess in the soil, Cu induced a reduction in the growth of this tree due to its toxic potential at high concentrations, which results in the limitation of plant development (DalCorso et al. 2014; Adrees et al. 2015). This result resembles that found by Marco et al. (2016), who investigated excess Cu in tree species Senna multijuga and Erythrina cristagalli, and found a decrease in their growth, probably due to excess Cu, which reduces plant growth and stabilization in the subsequent stages of their development. The reduction in growth observed in plants exposed to the highest Cu concentrations can also be attributed to the reduction in carbon assimilation (Cambrollé et al. 2015).
3.2 Effect of Copper on Gas Exchange
At a concentration of 200 mg kg−1 Cu, H. courbaril plants showed higher photosynthesis (A), stomatal conductance (gs) and carboxylation efficiency (A/Ci), up to the period of 100 days, with a decline in the same parameters at 120 days (Fig. 2 a, b, d). However, it was observed that excess Cu in the soil (800 mg kg−1) caused a decrease in parameters A, gs and A/Ci (Fig. 2a, b, d). H. courbaril showed an increase in water use efficiency (A/gs) at a concentration of 800 mg kg−1 Cu applied to the soil, with a decrease at 120 days (Fig. 2c).

Gas exchange of H. courbaril exposed to different Cu concentrations. a Photosynthesis (A). b Stomatal conductance (gs). c Water use efficiency (A/gs). d Carboxylation efficiency (A/Ci). Each value indicates treatment means ± SE
In H. courbaril plants, the maximum net photosynthetic rate (Amax) was higher in the treatment with 200 mg kg−1, followed by the concentration of 100 mg kg−1 Cu, with an increase in the measurements performed over the days (Fig. 3a). At 800 mg kg−1 Cu applied to the soil, the lowest Amax was observed (Fig. 3a). Regarding respiration rate in the dark (Rd), a greater Rd was observed at 100 mg kg−1, at 30 days of culture, followed by an apparent decline (Fig. 3b). At 800 mg kg−1, a smaller Rd occurred in relation to the other concentrations (Fig. 3b). Rd remained largely unchanged over time at this concentration. In the measurements, a lower light compensation point (LCP) was observed at a concentration of 800 mg kg−1 (Fig. 3c). After 6 days of cultivation, there was a significant increase and stabilization of this parameter for excess Cu (800 mg kg−1). On the other hand, for light saturation point (LSP), H. courbaril showed higher means at concentrations of 200 and 100 mg kg−1, and lower LSP at the highest Cu concentration (800 mg kg−1) (Fig. 3c). It is noticed that, over time, there was an increase in LSP at all concentrations, except for 0 mg kg−1. With respect to apparent quantum yield values (α) obtained for H. courbaril, it was observed that excess Cu (800 mg kg−1) caused a decrease and, with the increase in days of cultivation, an increase and stabilization in α was observed (Fig. 3d).

a Maximum net photosynthetic rate (Amax). b Respiration rate in the dark (Rd). c Light compensation point (LCP). d Light saturation point (LSP). e Apparent quantum yield (α) in H. courbaril plants exposed to different Cu concentrations. Each value indicates treatment means ± SE
Cu acts as a structural element of mitochondria, chloroplasts, and proteins, such as plastocyanin, which is involved in the electron transport chain between PS II and PS I in the photochemical phase of photosynthesis (Yruela 2013; DalCorso et al. 2014; Adrees et al. 2015). The presence of copper in the soil at concentrations up to 200 mg kg−1 positively influenced the evaluated parameters, resulting in a higher photosynthesis (A) in H. courbaril plants. Another probable factor for the high A is the high stomatal conductance (gs) at this concentration, thus favoring greater CO2 fixation by these plants.
Excess Cu ions in plants can cause competition with other metallic ions (Zn, Fe, Ni) and substitution of structural elements in enzymes, such as ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisco), causing a disturbance in photosynthesis; leading to an increase in intracellular reactive oxygen species (ROS) in leaves that are highly toxic and increasing lipid peroxidation; decreasing carbon fixation and energy production and, consequently, reducing or inhibiting plant development (Küpper and Andresen 2016). This possibly explains the lower photosynthesis (A) and the low maximum net photosynthetic rate (Amax) found in H. courbaril plants at the highest concentration (800 mg kg−1). Another likely explanation for the results found for A and Amax could be attributed to the different effects of Cu toxicity on the integrity or function of the photochemical apparatus—biochemical limitation, as well as its impact on leaf chlorophyll concentrations (Cambrollé et al. 2015). Fluorescence may occur, and it is another limitation for A, suggesting that excess Cu increases photoinhibition induced by light stress (Zhang et al. 2013; Cambrollé et al. 2015). These deleterious effects of excess Cu on this tree can be confirmed when analyzing carboxylation efficiency (A/Ci), which is reduced at a concentration of 800 mg kg−1, possibly due to toxicity that led to the inhibition of carboxylic enzymes (rubisco) (Mateos-Naranjo et al. 2013).
The maximum net photosynthetic rate (Amax) reflects the maximum photosynthetic capacity of the leaf (Zhang et al. 2014). Amax was influenced by the high Cu concentration (800 mg kg−1), indicating that the photosynthetic capacity of H. courbaril plants decreased with excess Cu. This fact can be reinforced by the light compensation point (LCP) and light saturation point (LSP), which reflect the plant ability to use the lowest and highest levels of light, respectively, showing the relationship between light and photosynthesis (Zhang et al. 2014), which was limited at 800 mg kg−1, when compared to the other treatments. These results differ from those found by Zhang et al. (2014), where LCP and LSP were higher with excess heavy metal, indicating that the metal could increase light use efficiency at high light levels and reduce it at low light levels. LCP is the light value in which CO2 fixation rate in photosynthesis is equal to the rate of CO2 release in respiration and photorespiration (Nunes et al. 2009). The high LCP reflects a higher respiration, a lower Rubisco carboxylase activity, and a greater Rubisco oxygenase activity, suggesting a possible protection mechanism to avoid photoinhibition, dissipating the excess of ATP and NADPH, producing internal CO2 (maintenance of Rubisco activity) and consuming oxidants (H2O2) through catalysis (Baccio et al. 2009). Cu is possibly at a very toxic level in the soil, reflecting a large amount of intracellular ions, preventing this protection mechanism with the increase in LCP. Respiration rate in the dark (Rd) was lower at 800 mg kg−1, when compared to the other treatments, possibly due to Cu toxicity. In apparent quantum yield (α), the negative effect of Cu excess is observed; however, over time (days), H. courbaril plants adapt to the stress condition induced by this metal, increasing α and showing tolerance.
The higher water use efficiency (A/gs) in H. courbaril plants at the highest copper concentration reflects in water savings. Plants that save water in environments with excess heavy metals are important for phytoremediation programs, once they are not damaged by additional stresses such as water, suggesting the possible use of this species in phytoremediation programs, since it tolerates more environmental variations such as drought. This fact favors a better performance (survival) of the plant, since A/gs is an indicator parameter of drought tolerance (Pinheiro et al. 2014). One of the major hurdles of phytoremediation is the influence of other environmental stresses (besides the presence of metal) on contaminant uptake by plants (Ali et al. 2013).
3.3 Root System Morphology and Dry Matter Attributes
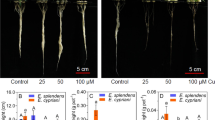
Excess Cu negatively affected all studied parameters of root system morphology, with a marked decline at 800 mg kg−1 Cu for length, surface area, mean diameter, and root volume of H. courbaril plants (Fig. 4a–d). However, it was observed that, for root length parameters up to a concentration of 172.68 mg kg−1, surface area of 169.37 mg kg−1, mean root diameter of 166.66 mg kg−1, and root volume of 193.16 mg kg−1, root development is favored and, above these concentrations, toxic and inhibitory effects are observed in the root system (Fig. 4a–d).

Root characteristics. a Length, b surface area, c mean diameter, and d volume of the species H. courbaril exposed to different Cu concentrations. *p ≤ 0.05%
Regarding the distribution of length, surface area, and volume through diameter classes for H. courbaril plants, the same behavior of the previous parameters was observed (Figs. 4 and 5), with root growth at initial concentrations above the control (0 mg kg−1) and root growth inhibition at higher copper concentrations. In general, the root system in H. courbaril is dominated by fine roots. However, it was observed that concentrations of 100, 200, and 400 mg kg−1 Cu have higher length, surface area, and volume of finer roots (0 to 1 mm), when compared to the control (0 mg kg−1). It is worth mentioning that, despite leading to an increase in these characteristics for fine roots at concentrations of 200 and 400 mg kg−1 Cu, an increase in medium roots (with a diameter of 2 mm) is simultaneously observed. At 800 mg kg−1, a decrease in length, surface area, and volume of finer roots is observed (Fig. 5a–c).

a Root length, b root surface area, and c root volume distributed through diameter classes in H. courbaril exposed to different Cu concentrations
For shoot dry matter (SDM), up to a concentration of 100 mg kg−1 and root dry matter (RDM) up to 200 mg kg−1, an increase in biomass occurs, followed by a decrease for these parameters at the other concentrations (Fig. 6a, b). The ratio between RDM/SDM increases at higher concentrations (Fig. 6c). For specific root length (SRL), the highest values were observed in the control and at 100 mg kg−1, with a decrease at 200, 400 and 800 mg kg−1 (Fig. 6d). Up to 200 mg kg−1, root fineness (RF) is favored, followed by a decrease at 400 and 800 mg kg−1, due to the toxic effect of Cu (Fig. 6e). Root tissue density (RMDe) showed an increase with high Cu concentrations applied to the soil (Fig. 6f).

Dry matter attributes and morphology of H. courbaril plants exposed to different Cu concentrations. a Shoot dry matter (SDM). b Root dry matter (RDM). c Ratio between root dry matter and shoot dry matter (RDM/SDM). d Specific root length (SRL). e Root fineness (RF). f Root tissue density (RMDe). Each value indicates treatment means ± SE
The morphological parameters of roots can be influenced by biotic and abiotic factors such as heavy metals, which may determine plant survival in a given environment (Cai et al. 2014). A well-developed root system is an important characteristic for stress tolerance. In general, exposure to stress leads to remodeling in root morphology (Cai et al. 2014). High copper concentrations affected root morphology/architecture in H. courbaril. The toxic effect of Cu resulted in a decrease in length, surface area, mean diameter, and root volume, which reflected in a reduction in both root and shoot biomass at these concentrations. Batool et al. (2015) emphasize that the evident reduction in root growth may be due to a reduction in cell division, leading to an increase in the cell wall thickness of the roots, when exposed to heavy metals.
These results were similar to those found by Cai et al. (2014) in the root morphology analysis of Elsholtzia haichowensis, emphasizing that the sharp reduction in root volume under Cu stress reflects the relevant decrease in root biomass, which depends directly on cell division and stretching. Similar effects were found by Bochicchio et al. (2015) in a combination of heavy metals Cd, Cu and Zn on the root morphology of Arabidopsis thaliana. Marco et al. (2016) reported that increased Cu reduced morphological parameters in the studied trees (Senna multijuga and Erythrina crista-galli). Feigl et al. (2013) found the inhibitory effect on the most prominent root system in Brassica juncea, compared to Brassica napus, caused by Cu stress, showing genetic variability among the species in the tolerance to this metal.
On the other hand, under micronutrient conditions, Cu favored the root development of H. courbaril and, according to Imada et al. (2008), the root surface area is the one that is most related to nutrient absorption, and an increase in root volume leads to a greater efficiency in nutrient absorption, favoring a good plant development.
Cu favored root length development, surface area, and volume of fine roots, which are the roots suitable for water absorption, up to a concentration of 400 mg kg−1. However, even at 400 mg kg−1, it was observed that the favoring of roots classified as thick (Ø > 2.0 mm) (Magalhães et al. 2012) also occurred as a sign of support, not absorption roots. Thus, up to 200 mg kg−1, concentration seems to have actually favored the root system of H. courbaril in soil exploration. However, toxic and inhibitory effects are observed for the same parameters, when at high concentrations in the soil, since fine roots are more sensitive under high Cu levels. In addition to the decrease in primary and lateral roots, root number and density are also reduced by Cu (Feigl et al. 2013). This fact can be observed when analyzing root anatomy, where the plants of this tree presented smaller cortex thickness and vascular cylinder area.
The application of the highest Cu concentration in the soil caused a decrease in dry matter attributes, with an increase in the ratio root/shoot dry matter (RDM/SDM), suggesting that there is an increase in root tissue density in this tree at the highest concentrations. It is known that a greater specific root length (SRL) reflects in a greater exploration and acquisition of water and nutrients in the soil per carbon unit (Bouma et al. 2001). This fact corroborates the result found for root fineness (RF). At 100 mg kg−1, SRL is favored, which was also observed with the increase in fine roots (RF). In reforestation areas, this Cu concentration could be indicated due to the stimulus in root increase for soil exploration.
The high root tissue density (RMDe) (roots with large amounts of dead and fibrous matter) is an example of the characteristic of species inserted in stress environments, which eventually reduce specific root length (Wahl and Ryser 2000). Cu can increase lignification in tolerant species, since it accumulates in the cell wall (Colzi et al. 2015) and may also be adhered to apoplastic barriers in the exodermis and endodermis (Caspary striae) (Freitas et al. 2015). This fact possibly explains the increase in the area of perivascular fibers (lignin) in H. courbaril leaves. Cu can also increase lignification in xylem cell walls (Lequeux et al. 2010), which is probably related to the increase in xylem diameter, that is, larger caliber vessels have a higher amount of lignin. Fibers are important in plants that are inserted in environments with metals, since lignin has adsorption sites of heavy metals (Lin et al. 2005). Fu et al. (2015) reported that the increase in Cu provided an increase in xylem vessels by 60% in E. haichowensis, which corroborates the increase in xylem vessels in the leaves of the studied tree.
3.4 Leaf and Root Anatomy
In H. courbaril leaves, it is observed that excess Cu caused a decrease in stomatal density, adaxial and abaxial cuticle thickness, and area of perivascular fibers, except for xylem diameter (Fig. 7a–e). On the other hand, for the root anatomy of these plants, there is a reduction in vascular cylinder area and in cortex thickness, with the increase in Cu concentrations in the soil (Fig. 7f, g).

Leaf and root anatomy of H. courbaril plants exposed to increasing Cu concentrations. Leaf: a stomatal density, b adaxial cuticle thickness, c abaxial cuticle thickness, d xylem diameter, e area of perivascular fibers. Root: f Vascular cylinder area and g cortex thickness. Regressions with asterisk correspond to p ≤ 0.05% probability. Each value indicates treatment means ± SE
The absorption of heavy metals by a plant can cause several anatomical changes, with possible functional consequences in the plant, besides other morphophysiological alterations (Batool et al. 2015). The abaxial cuticle in the leaves, as well as the vascular cylinder area and cortex thickness in H. courbaril roots, was affected by excess Cu. These anatomical changes in leaves and roots possibly influenced the negative effect on gas exchange and, mainly, the root morphology of this tree. However, up to 200 mg kg−1, Cu stimulates cuticle production in the leaves, which would possibly help these plants survive under stress conditions, such as drought due to decreased transpiration.This result resembles those found by Fu et al. (2015).
The stomatal density of H. courbaril plants was favored up to 200 mg kg−1 Cu, which explains the higher photosynthetic rate (A and Amax). Even with a decline in stomatal density at the highest concentrations, it was still similar to the control (0 mg kg−1), differing from the results found by Silva et al. (2012), who reported a decreasing trend for stomatal density at the highest concentrations. The same authors further emphasize that a reduction in stomatal size can occur and may lead to negative impacts on gas exchange, such as photosynthesis and transpiration, due to the presence of the metal, inducing stomatal closure.
When exposed to increasing Cu concentrations, the smallest vascular cylinder area and cortex thickness of H. courbaril plants may be related to the marked decrease in the size of root tissue cells, due to the decreased elasticity of the cell walls, either of the cortical parenchyma or the conduction system (xylem and phloem), resulting in reduced root diameter, when exposed to excess metals (Batool et al. 2015). This fact may also be related to high Cu levels, altering the photosynthetic metabolism of leaves, responsible for the energy supply (ATP and NADPH) for root development, which reflects in morphological changes in the root system.
3.5 Copper Content in Tissues and Indexes
With the increase in copper in the soil, its increase was also observed in H. courbaril tissues (p < 0.05) (Fig. 8). Cu was absorbed and mainly allocated to the roots of this species (Fig. 8).

Copper content in roots, stems, and leaves of H. courbaril plants exposed to different Cu concentrations. Total Cu bioaccumulated in plants p ≤ 0.05% (asterisk)
Metal absorption by plants depends on the physiological requirement and metal availability in the soil (DalCorso et al. 2014). Copper is absorbed and found at higher concentrations in roots, since it strongly binds to the cell wall of root cells and, therefore, it is not easily mobile in the plant, resulting in a low translocation of this metal to the shoot (Kabata-Pendias 2011). The Cu absorbed by H. courbaril plants was retained in the roots, mainly at the highest concentrations. This result resembles that found by Silva et al. (2015), with emphasis on the tree Enterolobium contortisiliquum (Vell.) Morong.; it is the main species studied for accumulating Cu in the root system. This behavior is used by many plants as a tolerance mechanism to heavy metals to reduce metal translocation to the shoot and thus avoid damage caused by the toxicity of the heavy metal to the photosynthetic apparatus present in the leaves (Marco et al. 2016). This fact corroborates the results found in this study, once they did not die and still accumulated the metal in the roots, although the plants suffered with the presence of excess copper.
For tolerance index (TI) and translocation factor (TF) in H. courbaril plants, it is observed that the values decreased with increasing Cu concentrations applied to the soil (p < 0.05) (Table 2).
Even with the toxic effect of Cu at high concentrations, H. courbaril plants presented tolerance and survival. The tolerance index (TI) confirms this fact, based on biomass production, even at the highest concentration, where this tree managed to survive. TI results close to 1.0 indicate higher stress tolerance caused by the metal (Souza et al. 2012b). Increasing Cu concentrations incorporated in the soil reduced the translocation factor (TF) of this tree, and it was observed that only 4.45% Cu was translocated from the roots to the shoot by the plants when they were exposed to the concentration of 800 mg kg−1. Cu concentrations higher in the roots and lower in the shoot suggest that there may be some protection mechanisms through which Cu translocation from shoot to root was limited (Wang et al. 2016). This fact is probably related to tolerance mechanisms against the deleterious effects that excess Cu can cause in the plant (Cambrollé et al. 2013). A study by Marco et al. (2016) with trees showed similar results with TF reduction in soils with Cu in Senna multijuga and Erythrina crista-galli.
4 Conclusion
Up to a concentration of 200 mg kg−1, copper favored and stimulated growth, gas exchange and root morphoanatomy of H. courbaril plants. However, excess Cu (800 mg kg−1) caused toxicity, with a decrease in photosynthesis and biochemical limitations, inhibition of root development, mainly of fine roots, and changes in leaf and root anatomy for this tree. Cu was accumulated in the roots of these plants as a protection mechanism to the photosynthetic apparatus present in the leaves. Plants of Hymenaea courbaril L. showed high tolerance to excess copper and can be indicated for areas contaminated by this metal and phytoremediation programs such as phytostabilization.
References
Adrees, M., Ali, S., Rizwan, M., Ibrahim, M., Abbas, F., Farid, M., Zia-ur-Rehman, M., Irshad, M. K., & Bharwana, S. A. (2015). The effect of excess copper on growth and physiology of important food crops: a review. Environmental Science and Pollution Research, 22(11), 8148–8162.
Ali, H., Khan, E., & Sajad, M. A. (2013). Phytoremediation of heavy metals—concepts and applications. Chemosphere, 91(7), 869–881.
Baccio, D., Tognetti, R., Minnocci, A., & Sebastiani, L. (2009). Responses of the Populus x euramericana clone I-214 to excess zinc: carbon assimilation, structural modifications, metal distribution and cellular localization. Environmental and Experimental Botany, 67(1), 153–163.
Batool, R., Hameed, M., Ashraf, M., Ahmad, M. S. A., & Fatima, S. (2015). Physio-anatomical responses of plants to heavy metals. In: M. Öztürk, M. Ashraf, A. Aksoy, M.S.A. Ahmad (Eds.) Phytoremediation for green energy (pp. 79–96). Netherlands: Springer.
Bochicchio, R., Sofo, A., Terzano, R., Gattullo, C. E., Amato, M., & Scopa, A. (2015). Root architecture and morphometric analysis of Arabidopsis thaliana grown in Cd/Cu/Zn-gradient agar dishes: a new screening technique for studying plant response to metals. Plant Physiology and Biochemistry, 91, 20–27.
Bouma, T. J., Nielsen, K. L., Van Hal, J., & Koutstaal, B. (2001). Root system topology and diameter distribution of species from habitats differing in inundation frequency. Functional Ecology, 15(3), 360–369.
Cai, S., Xiong, Z., Li, L., Li, M., Zhang, L., Liu, C., & Xu, Z. (2014). Differential responses of root growth, acid invertase activity and transcript level to copper stress in two contrasting populations of Elsholtzia haichowensis. Ecotoxicology, 23(1), 76–91.
Cambrollé, J., Mancilla-Leytón, J. M., Muñoz-Vallés, S., Figueroa-Luque, E., Luque, T., & Figueroa, M. E. (2013). Effects of copper sulfate on growth and physiological responses of Limoniastrum monopetalum. Environmental Science and Pollution Research, 20(12), 8839–8847.
Cambrollé, J., García, J. L., Figueroa, M. E., & Cantos, M. (2015). Evaluating wild grapevine tolerance to copper toxicity. Chemosphere, 120, 171–178.
Carmo, C. A. F. S., Araújo, W. S., Bernardi, A. C. C. & Saldanha, M. F. C. (2000) Métodos de análise de tecidos vegetais utilizados na Embrapa Solos. Embrapa Solos, Rio de Janeiro. http://www.infoteca.cnptia.embrapa.br/bitstream/doc/337672/1/Metododeanalisedetecido.pdf. (in portuguese).
Colzi, I., Pignattelli, S., Giorni, E., Papini, A., & Gonnelli, C. (2015). Linking root traits to copper exclusion mechanisms in Silene paradoxa L.(Caryophyllaceae). Plant and Soil, 390(1–2), 1–15.
DalCorso, G., Manara, A., Piasentin, S., & Furini, A. (2014). Nutrient metal elements in plants. Metallomics, 6(10), 1770–1788.
Ent, A. V. D., Baker, A. J., Reeves, R. D., Pollard, A. J., & Schat, H. (2013). Hyperaccumulators of metal and metalloid trace elements: facts and fiction. Plant and Soil, 362(1–2), 319–334.
Feigl, G., Kumar, D., Lehotai, N., Tugyi, N., Molnár, Á., Ördög, A., Szepesi, Á., Gémes, K., Laskay, G., Erdei, L., & Kolbert, Z. (2013). Physiological and morphological responses of the root system of Indian mustard (Brassica juncea L. Czern.) and rapeseed (Brassica napus L.) to copper stress. Ecotoxicology and Environmental Safety, 94, 179–189.
Freitas, T. A., França, M. G. C., de Almeida, A. A. F., de Oliveira, S. J. R., de Jesus, R. M., Souza, V. L., Silva, J. V. S., & Mangabeira, P. A. (2015). Morphology, ultrastructure and mineral uptake is affected by copper toxicity in young plants of Inga subnuda subs. luschnathiana (Benth.) TD Penn. Environmental Science and Pollution Research, 22(20), 15479–15494.
Fu, L., Chen, C., Wang, B., Zhou, X., Li, S., Guo, P., Shen, Z., Wang, G., & Chen, Y. (2015). Differences in copper absorption and accumulation between copper-exclusion and copper-enrichment plants: a comparison of structure and physiological responses. PLoS One, 10(7), e0133424.
Gautam, S., Anjani, K., & Srivastava, N. (2016). In vitro evaluation of excess copper affecting seedlings and their biochemical characteristics in Carthamus tinctorius L. (variety PBNS-12). Physiology and Molecular Biology of Plants, 22(1), 121–129.
Imada, S., Yamanaka, N., & Tamai, S. (2008). Water table depth affects Populus alba fine root growth and whole plant biomass. Functional Ecology, 22(6), 1018–1026.
Kabata-Pendias, A. (2011). Trace elements in soils and plants (4th ed.). London: CRC Press.
Küpper, H., & Andresen, E. (2016). Mechanisms of metal toxicity in plants. Metallomics, 8, 269–285.
Lequeux, H., Hermans, C., Lutts, S., & Verbruggen, N. (2010). Response to copper excess in Arabidopsis thaliana: impact on the root system architecture, hormone distribution, lignin accumulation and mineral profile. Plant Physiology and Biochemistry, 48(8), 673–682.
Lin, C. C., Chen, L. M., & Liu, Z. H. (2005). Rapid effect of copper on lignin biosynthesis in soybean roots. Plant Science, 168(3), 855–861.
Magalhães, P. C., De Souza, T. C., Cantão, F. R. D. O., & Padilha, F. A. (2012). Root morphology of maize lines with contrasting drought resistance under three toxic levels of aluminum. Brazilian Journal of Maize and Sorghum, 11(1), 35–48.
Marco, R., Silva, R. F., Andreazza, R., Ros, C. O., Scheid, D. L., & Bertollo, G. M. (2016). Copper phytoaccumulation and tolerance by seedlings of native Brazilian trees. Environmental Engineering Science, 33(3), 176–184.
Mateos-Naranjo, E., Andrades-Moreno, L., Cambrollé, J., & Perez-Martin, A. (2013). Assessing the effect of copper on growth, copper accumulation and physiological responses of grazing species Atriplex halimus: ecotoxicological implications. Ecotoxicology and Environmental Safety, 90, 136–142.
Nunes, C., Araújo, S. S., Silva, J. M., Fevereiro, P., & Silva, A. B. (2009). Photosynthesis light curves: a method for screening water deficit resistance in the model legume Medicago truncatula. Annals of Applied Biology, 155(3), 321–332.
Pinheiro, D. G., Streck, N. A., Richter, G. L., Langner, J. A., Winck, J. E. M., Uhlmann, L. O., & Zanon, A. J. (2014). Soil water threshold for transpiration and leaf growth in cassava plants in two water deficit periods. Revista Brasileira de Ciência do Solo, 38(6), 1740–1749.
Shi, X., Zhang, X., Chen, G., Chen, Y., Wang, L., & Shan, X. (2011). Seedling growth and metal accumulation of selected woody species in copper and lead/zinc mine tailings. Journal of Environmental Sciences, 23(2), 266–274.
Silva, N. D. G., Cholewa, E., & Ryser, P. (2012). Effects of combined drought and heavy metal stresses on xylem structure and hydraulic conductivity in red maple (Acer rubrum L.) Journal of Experimental Botany, 63(16), 5957–5966.
Silva, R. F. D., Ros, C. O. D., Scheid, D. L., Grolli, A. L., Marco, R. D., & Missio, E. L. (2015). Copper translocation and tolerance in seedlings of tree species grown in contaminated soil. Revista Brasileira de Engenharia Agrícola e Ambiental, 19(11), 1093–1099.
Souza, T. D., Castro, E. D., Pereira, F. J., Parentoni, S. N., & Magalhães, P. C. (2009). Morpho-anatomical characterization of root in recurrent selection cycles for flood tolerance of maize (Zea mays L.) Plant Soil and Environment, 55(11), 504–510.
Souza, T. C. D., Magalhães, P. C., Pereira, F. J., Castro, E. M. D., Silva Junior, J. M. D., & Parentoni, S. N. (2010). Leaf plasticity in successive selection cycles of ‘Saracura’ maize in response to periodic soil flooding. Pesquisa Agropecuária Brasileira, 45(1), 16–24.
Souza, T. C., De Castro, E. M., Magalhães, P. C., Alves, E. T., & Pereira, F. J. (2012a). Early characterization of maize plants in selection cycles under soil flooding. Plant Breeding, 131(4), 493–501.
Souza, S. C. R., de Andrade, S. A. L., de Souza, L. A., & Schiavinato, M. A. (2012b). Lead tolerance and phytoremediation potential of Brazilian leguminous tree species at the seedling stage. Journal of Environmental Management, 110, 299–307.
Souza, V. L., Almeida, A. A. F., Souza, J. S., Mangabeira, P. A. O., Jesus, R. M., Pirovani, C. P., Ahnert, D., Baligar, V. C., & Loguercio, L. L. (2014). Altered physiology, cell structure, and gene expression of Theobroma cacao seedlings subjected to Cu toxicity. Environmental Science and Pollution Research, 21, 1217–1230.
Wahl, S., & Ryser, P. (2000). Root tissue structure is linked to ecological strategies of grasses. New Phytologist, 148(3), 459–471.
Wang, Q. Y., Liu, J. S., & Hu, B. (2016). Integration of copper subcellular distribution and chemical forms to understand copper toxicity in apple trees. Environmental and Experimental Botany, 123, 125–131.
Yruela, I. (2013). Transition metals in plant photosynthesis. Metallomics, 5(9), 1090–1109.
Zhang, L., Zhao, J., Duan, M., Zhang, H., Jiang, J., & Yu, R. (2013). Inhibition of dsDNA-templated copper nanoparticles by pyrophosphate as a label-free fluorescent strategy for alkaline phosphatase assay. Analytical Chemistry, 85(8), 3797–3801.
Zhang, X., Zhang, X., Gao, B., Li, Z., Xia, H., Li, H., & Li, J. (2014). Effect of cadmium on growth, photosynthesis, mineral nutrition and metal accumulation of an energy crop, king grass (Pennisetum americanum× P. purpureum). Biomass and Bioenergy, 67, 179–187.
Acknowledgements
This paper is dedicated to Prof. Dr. Marcelo Polo, Federal University of Alfenas, Alfenas, MG, Brazil, for the long-term merits of his studies on plant ecophysiology.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Marques, D.M., Veroneze Júnior, V., da Silva, A.B. et al. Copper Toxicity on Photosynthetic Responses and Root Morphology of Hymenaea courbaril L. (Caesalpinioideae). Water Air Soil Pollut 229, 138 (2018). https://doi.org/10.1007/s11270-018-3769-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11270-018-3769-2