
Abstract
Classical swine fever (CSF) is an economically important and highly contagious disease of pigs caused by CSF virus, genus Pestivirus. Serological diagnosis of the disease is highly valuable for surveillance and thereby containment of spread of the disease. In this study, we have demonstrated the development of CSFV envelope glycoprotein E2-based indirect ELISA (E2-iELISA) for the detection of CSFV specific antibodies. The full-length E2 protein was expressed in E. coli and the purified protein was used as a coating antigen in indirect ELISA for detecting CSFV specific antibodies in pigs. A panel of 506 pig sera samples was used to validate the ELISA and the results were highly comparable to the results obtained with the commercial antibody detection kit (PrioCHECK CSFV Ab kit). The in-house E2-iELISA demonstrated high diagnostic sensitivity (95.4%) and specificity (95.5%), highlighting its potential application for sero-surveillance or monitoring of the disease in the swine population.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Classical swine fever (CSF), formerly called hog cholera (HC), is an economically important, highly contagious disease in pigs. The disease is a multi-systemic, often fatal infection in swine including domestic, wild boars and feral pigs (Lamothe-Reyes et al. 2023). It is one of the World Organization for Animal Health (WOAH, formerly OIE) notifiable diseases. The disease shows worldwide distribution whereas it is not present in Australia, New Zealand and North America. The outbreak in Japan after being CSF-free previously, showed the re-emergence of the disease in 2018. CSF is an endemic disease in many developing countries posing a constant threat to the pig population (Ganges et al. 2020) and jeopardizing food security worldwide. A large number of outbreaks have been reported in India since the first outbreak in 1944, thereby acquiring an endemic status (Dong and Chen 2007; Patil et al. 2010).
The clinical symptoms and severity of the disease vary from acute to chronic form depending on the strain of the virus, virulence of the strain, age and the immune status of the animal. CSF is characterized by several clinicopathological signs such as high fever (> 40℃), conjunctivitis, respiratory signs, diarrhea, skin hemorrhages, lethargy, convulsions, clumsy movements, and staggering gait of infected animals with an incubation period of 03 to 10 days (Postel et al. 2018). The disease causes abortions, stillbirths, mummified fetuses, malformations, and high mortalities in pregnant populations. Direct or indirect contact between pigs, contaminated feed and water, through infected boar semen, artificial insemination, farm equipment, transport vehicles, and visitors are the main possible means of the disease spread (Malik et al. 2020).
Classical swine fever virus (CSFV) is the etiological agent of CSF belonging to the Flaviviridae family of the Pestivirus genus. Bovine viral diarrhea virus (BVDV) 1 and 2, border disease virus (BDV) are the other members of the genus. The International Committee on Taxonomy of Viruses (ICTV) has recently re-designated Pestivirus genus, CSFV as Pestivirus C (Wei et al. 2021). CSFV is an enveloped, single-stranded, positive-sense RNA virus with the size of 40–60 nm in diameter. The genome of CSFV is 12.3 kb enclosing a 5′ untranslated region (5′ UTR), a single large open reading frame (ORF) encoding a polyprotein of about 3900 amino acids, and a 3′ untranslated region (3′ UTR). CSFV poly protein is post translationally processed into four structural proteins namely, Capsid protein (C), envelope glycoproteins (Erns, E1 and E2and eight non-structural proteins (Npro, p7, NS2, NS3, NS4A, NS4B, NS5A and NS5B) (Meyers and Thiel 1996; Wang et al. 2020). Antibodies have been detected so far against Erns, E2, and NS3 in the pigs that recover from CSFV infection. E2 is the most immunogenic protein and has the ability to produce neutralizing antibodies while, Erns has low or no neutralizing activity. NS3 is not suitable for diagnostics due to its high genetic stability and presence of non-CSFV pestiviruses. The Erns and E2 glycoproteins are the two major targets for developing vaccine candidates as well as for diagnostics (Beer et al. 2007).
Laboratory confirmation is imperative for the early and specific detection of the infection as a wide range of clinical signs can be observed in the affected animals. The virus neutralization test (VNT) is the gold standard test for the serological diagnosis of CSFV infections (Cao et al. 2022). However, the requirement of cell culture, live virus, expertise and a dedicated facility makes the test expensive and difficult to deploy. Rapid tests such as immunochromatographic strip tests have been developed for detection of CSFV antibodies (Bai et al. 2022; Huynh et al. 2024). While strip tests are faster and simpler to perform, they are not suitable for high-throughput screening. Alternatively, serological diagnosis by enzyme-linked immunosorbent assay (ELISA) (Ji et al. 2018; Cao et al. 2023) is advantageous over VNT as a large number of samples can be screened at a time, less time consuming, no requirement of live virus or cell culture facility. ELISA is thus a valuable tool for surveillance of CSF in apparently disease-free areas or for monitoring in a CSF eradication program in the vaccinated population (Yi et al. 2022). Despite recent efforts to develop new and safer marker vaccines against CSF, along with improved diagnostic tools, there is still a need for further improvements. Here, we describe the development of an in-house full-length E2-protein based indirect ELISA (E2-iELISA) for detection of CSFV E2 antibodies in the swine population.
Materials and methods
Serum samples
Serum samples collected randomly and during the phase of disease outbreaks were maintained at the laboratory. The samples were collected from different regions of the nine Indian states (Telangana, Tamil Nadu, Kerala, Maharashtra, Sikkim, Tripura, Punjab, Odisha and Mizoram). A total of 506 samples were used in this study including the standard positive and negative control serum. The samples included vaccinated (n = 198), non-vaccinated (n = 245) and few randomly collected (n = 63).
For western blot, the polyclonal serum available in the laboratory (raised in rabbits against the CSFV, vaccine strain) was used for detection of the expressed protein.
Expression and purification of full-length E2 protein
The nucleotide sequence of the entire E2 gene of an Indian isolate of CSFV was obtained from the GenBank database at the National Center for Biotechnology Information website (accession no. QAV56547.1) and synthetic gene was commercially sourced. The E2 ORF (1017 bp) was cloned into the expression plasmid pET32a(+) and the orientation of the E2 gene was confirmed by sequencing. The construct designated as pET32a(+)-E2 plasmid (supplementary data) was transformed into E. coli BL21(DE3) pLysS strain and individual colony was grown at 37ºC in Luria Bertani Broth containing ampicillin (50 µg/mL) and chloramphenicol (35 µg/mL). Once the bacterial culture reached an optical density (OD at 600 nm) of 0.6, Isopropyl-β-D-1-thiogalactopyranoside (IPTG) was added at a final concentration of 1 mM and the cells were further grown for 8 h at 30ºC. The bacterial culture without the addition of IPTG was also maintained as an un-induced culture for the analysis of protein expression. The cells were harvested by centrifugation at 4000 rpm for 10 min and were purified by Ni-NTA affinity chromatography. Briefly, the cell pellet was resuspended in a denaturation buffer (6 M Urea, 50 mM Tris, 300 mM NaCl pH 8.0), sonicated and centrifuged. The cell lysate supernatant containing the protein was allowed to bind for 1 h at room temperature with Ni-NTA resin (Invitrogen) equilibrated with denaturation buffer in a shaker. Alternatively, inclusion bodies were first isolated (cells were resuspended in 10mM Tris-HCl pH 8.0, sonicated, centrifuged at 12,000xg to get pellet containing inclusion bodies) and subsequently suspended in the denaturation buffer and then allowed to bind with the resin equilibrated with same buffer. The resin was then washed with the denaturation buffer containing 25 mM imidazole. The protein was eluted with the denaturation buffer containing 250 mM imidazole in 1 mL fractions. The eluted fractions were checked in SDS-PAGE and the fractions containing the target protein were pooled, dialyzed at 4 °C overnight against phosphate buffered saline (PBS) pH 7.4 for renaturation of the protein. The protein obtained in soluble form was quantified using a Qubit 4 Fluorometer (Invitrogen).
Characterization of the protein
The purified protein was subjected to SDS-PAGE analysis and western blotting. Briefly, 12% polyacrylamide gel was used to resolve the proteins and the proteins were either stained with Coomassie brilliant blue R-250, or transferred onto the nitrocellulose membrane for western blot analysis. Rabbit anti-CSFV polyclonal antibody, raised against the virus, was used as the primary antibody (1:100 dilution) and incubated for 1 h, followed by washing three times with tris buffered saline containing 0.1% tween 20. Goat anti-rabbit HRP conjugate (Dako, Denmark) (at 1:2000 dilution) was added as a secondary antibody, and incubated for 1 h. The protein reactivity was visualized by developing the membrane using the western ECL substrate (Bio Rad, USA).
Development of E2 based indirect ELISA (E2-iELISA)
A checkerboard titration method was performed to evaluate the optimal concentrations of coating antigen and sera dilutions. Briefly, the 96-well ELISA plates (Maxisorp, Nunc) were coated with purified E2 antigen at concentrations of 25, 50, 100 and 200 ng/well in the carbonate bicarbonate buffer (pH 9.6) and incubated at 37ºC for 1 h. The plates were washed with PBST (PBS containing 0.05% Tween-20) and added with block buffer (wash buffer containing 2% skim milk powder, and 0.5% Bovine serum albumin fraction V) for 30 min at 37ºC. A standard positive serum sample was added to the washed ELISA plate at dilutions of 1:50, 1:100, 1:200 and 1:400 in the block buffer and incubated at 37ºC for 1 h followed by washing. The secondary antibody, anti-pig conjugate (Bethyl lab) was added to the ELISA plate at 1:20,000 dilution. The reactivity of antibodies was detected using O-Phenylenediamine dihydrochloride (OPD) as the substrate and the reaction was stopped using 1 M H2SO4. The optical density (OD) was measured at 492 nm using a microplate reader (Infinite F50, TECAN). Percent positivity (PP) value was calculated using the following formula:
Where, OD sample, OD negative and OD positive represent OD values obtained with test serum sample, negative control serum and positive control serum, respectively.
The PrioCHECK CSFV Ab test
The PrioCHECK CSFV Ab Plate kit is a commercial test (Thermo Fisher Scientific, USA) for the detection of antibodies against CSFV in clinical samples such as serum and plasma of suspected pigs. The test was performed as per the manufacturer’s protocol. The Percent inhibition (PI) values were calculated as per the given formula with a PI ≥ of 50% being considered positive while, the PI ≤ of 30% being considered negative. Percent inhibition (PI) value was calculated using the following formula where reference serum 4 indicates standard negative control serum:
Analytical sensitivity and specificity
The analytical sensitivity of the E2-iELISA was determined using standard positive and negative control sera. Different dilutions of the sera (1:10, 1:25, 1:50, 1:100, 1:200, 1:400, 1:800, and 1:1600) were tested in the E2-iELISA for determining the endpoint detection limits. The specificity of the test was determined by testing other disease serum samples specific for Bovine viral diarrhoea (BVD) (n = 8), African swine fever (ASF) (n = 4), Foot-and mouth disease (FMD) (n = 22) and Haemorrhagic septicemia (HS) (n = 4) in the E2-iELISA.
Diagnostic sensitivity and specificity of the E2-iELISA
The diagnostic sensitivity and specificity of the developed E2-iELISA were tested using a panel of 506 random swine sera samples which included standard positive and negative control sera. The test was compared with the commercial kit using the same panel of serum samples and the cut-off was calculated by receiver operator characteristic (ROC) analysis for maximum diagnostic sensitivity and specificity using the software MedCalc version 22.023 (MedCalc Software Ltd). Samples with PP values greater than or equal to the cut-off value were considered positive and samples with PP less than the cut-off were considered to be negative.
Results
Expression and purification of full length E2 protein
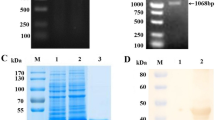
The recombinant CSFV E2 protein expressed in E. coli was analyzed using pellet treated with sample buffer in SDS PAGE. The IPTG-induced E. coli cell lysate showed an expression of protein at an expected size of 45 kDa which was not observed in the un-induced cell lysate confirming the expression of the target protein (Fig. 1). The protein was expressed in insoluble form. From the lysate prepared from induced culture of E. coli by sonication of pellet in the presence of protease inhibitor cocktail (1x) and phenyl methyl sulfonyl fluoride (1mM), the protein was purified by Ni-NTA affinity chromatography (Thermo fisher, USA) under denaturing conditions. The eluted fractions containing the proteins were dialysed against the PBS (0.1 M) and stored. The protein was quantified using the Qubit 4 fluorometer (Invitrogen) and the concentration was found to be on an average, 5 ± 1.8 mg from 1 L broth culture.

Expression of E2 protein. SDS-PAGE analysis of E2 protein on staining with Coomassie brilliant blue R-250. Protein marker (lane 1), uninduced cell lysate (lane 2), IPTG induced cell lysate expressing E2 protein (indicated with an arrow, lane 3)
Characterization of E2 protein by western blot analysis
The purified protein was confirmed by SDS-PAGE and western blot analysis. A distinct band with a molecular weight of approximately 45 kDa was observed in the SDS-PAGE (Fig. 2A). The purified protein also showed specific reactivity with the anti-CSFV polyclonal antibody in western blot (Fig. 2B).

Characterisation of E2 protein. SDS-PAGE (A) and Western blot analysis (B) of purified E2 protein. A Protein marker (lane 1), purified E2 protein (lane 2), B Reactivity of E2 protein with CSF convalescent serum (lane 1), Protein marker (lane 2)
Optimization of E2-iELISA
Based on the checkerboard titration data using standard control serum samples, the optimized parameters for the indirect ELISA were determined. The optimal concentration of the coating antigen was found to be 50 ng/well and the dilution of the swine serum samples of 1:100 was found to be adequate to allow maximum discrepancy between the standard positive and negative control samples (Fig. 3).

Optimization of CSFV E2-iELISA procedure. Optimization of concentration of coating antigen and dilution of swine sera using checkerboard titration method. P/N, positive control/negative control; ✵, the optimized condition for ELISA test
Diagnostic performance of E2-iELISA in comparison with PrioCHECK CSFV Ab test
A total of 506 swine serum samples were tested using the optimized parameters in the developed E2-iELISA and PrioCHECK commercial kit. Samples representing different profiles with respect to CSFV infection were tested, and the results are summarized in Table 1. The cut-off, diagnostic sensitivity and specificity of the developed ELISA were determined by receiver operator characteristic (ROC) analysis using the MedCalc software. Among multiple cut-off values suggested by Youden index, the optimal cut-off Percent positivity (PP) value was selected at 35% for maximum diagnostic sensitivity of 95.4% (95% CI: 89.6–98.5) and specificity of 95.5% (95% CI: 92.9–97.3) as shown in Table 2. Samples with a PP value ≥ of 35% were considered positive while samples with a PP value less than 35% were considered as negative. The results of the E2-iELISA highly correlated with the results obtained in PrioCHECK CSFV Ab kit as evidenced by the Table 1 and scatter plot (Fig. 4B). The ROC analysis revealed an area under the curve (AUC) value of 0.97 which is considered an excellent performance for the assay on the given set of samples. Based on the above set parameters, prevalence rate of 21.54% (109/506) was found when randomly collected serum samples were screened using the E2-iELISA.

Evaluation of a panel of positive and negative CSF sera using E2-iELISA. A Receiver operating characteristic (ROC) analysis of serum samples (n = 506). B Scatter Plot analysis of results between the E2-iELISA and the PrioCHECK CSFV Antibody ELISA test
Analytical sensitivity
The analytical sensitivity of the test was analyzed using three CSFV standard positive sera and a standard negative serum at different dilutions. The E2-iELISA displayed a higher sensitivity with an endpoint detection limit of 1:200 dilution of the serum (Fig. 5).

Limit of detection of the E2-iELISA. Samples (n = 3) positive in the ELISA were serially diluted and tested in E2-iELISA to determine their end point detection limits at the indicated dilutions
Analytical specificity
To rule out the non-specific reactivity of the E2-iELISA, serum samples positive for BVD, ASF, FMD and HS were screened in the E2-iELISA. The test showed no cross-reactivity with these serum samples. All the serum samples showed PPV lower than the cutoff value set in the E2-iELISA revealing specific reactivity.
Discussion
Classical swine fever has a severe socio-economic impact on the global pig industry (Luo et al. 2014). Specific and quick diagnosis is very crucial for the control and sero-surveillance of the disease. The two major strategies for control of the disease are, systematic vaccination and stamping out policy (under non-vaccination conditions), both of which require rapid diagnosis. Systematic vaccination in CSF-endemic countries is the most effective strategy as stamping out involves high costs. Eradication and control programs have been implemented by many countries for the control of the disease (Moennig and Becher 2015). Two types of vaccines are available for CSF, the live attenuated vaccine and E2 subunit vaccines. The E2 subunit vaccines are of great advantage as they allow differentiation of infected pigs from vaccinated pigs through companion diagnostics Erns ELISA (Yang et al. 2012). The conventional live vaccine is highly safe and efficacious but no DIVA (differentiating infected from vaccinated animals) strategy can be applied as vaccinated animals elicit similar antibody responses as observed in naturally infected animals (Nath et al. 2016; Yuan et al. 2023). Many DIVA vaccines through several approaches have been described but none of them are making it to the market making it difficult to develop new diagnostics for sero-diagnosis of the disease.
The virus neutralization test (VNT) is the gold standard for detection of CSFV-specific antibodies (WOAH). However, VNT has several drawbacks such as time required for testing, necessity for specialized expertise and dedicated cell culture facility, apart from challenges associated with handling of live viruses. Hence it is unsuitable for large-scale screening purposes. Therefore, simpler and faster diagnosis is needed for specific detection of the disease such as ELISA which detects antibodies to structural proteins of the virus. Upon infection, three proteins, namely E2, Erns, and NS3 induce detectable antibodies but antibodies against E2 and Erns confer virus neutralizing potential and protective immunity (Wong et al. 1998). Since E2 is a major glycoprotein that induces neutralizing antibodies and provides protective immunity, it is widely used as a marker for measuring vaccine efficacy and antibody titer associated with infection. Previously reported ELISAs are indirect ELISA and competitive ELISA for specific detection of antibodies against CSFV based on the truncated E2 of CSFV expressed in mammalian cells and E. coli cells (Wensvoort et al. 1986; Moser et al. 1996; Colijn et al. 1997).
The Prokaryotic expression system has been a powerful tool for the cost-effective production of large amounts of immunologically active recombinant proteins. Expression of full-length protein of E2 in E. coli has been challenging and probably requires fusion tags to promote proper folding and improve protein solubility (Lin et al. 2008; Kumar et al. 2016; Bachan et al. 2019). A number of laboratories have expressed truncated proteins in E. coli to overcome this problem for the application in sero-diagnostics (Clavijo et al. 2001; Lin et al. 2008). In this paper, we expressed the full-length CSFV E2 protein in E. coli with only 6xHis-tag. Protein was of expected size (~ 45 kDa) in western blot or SDS-PAGE as a distinct band unlike apparent cleavage of protein observed in an earlier report (Wong et al. 1998). The apparent molecular weight of the protein was slightly higher than the predicted Mw, however such observation is reported earlier (Ma et al., 2023). The recombinant protein showed antigenicity with CSFV positive sera in ELISA or western blot. The ELISA optimized with the recombinant protein showed excellent reactivity with positive sera and did not react with the negative sera or sera positive for antibodies to FMDV, HS, ASFV and BVDV, a closely related virus to CSFV. Further, the purified E2 protein showed specific reactivity with anti-CSFV polyclonal antibody in the western blot and thus was employed as a diagnostic antigen in the E2-iELISA.
The Optimized E2-iELISA showed that 50 ng/well of E2 antigen for coating and 1:100 dilution of the swine serum samples was found to be adequate for discriminating between positive and negative samples as compared to a previous study which required more amount of E2 as coating antigen (600 ng/well) and lower dilution of serum (1:80) (Lin et al. 2008).
A total of 506 swine serum samples were tested using the optimized parameters in the E2-iELISA in comparison with a commercial kit (PrioCHECK CSFV Ab kit). The E2-iELISA demonstrated a diagnostic sensitivity (Dsn) of 95.4% and diagnostic specificity (Dsp) 95.5%. These values are marginally lower than the ELISA using the E2 protein expressed in insect cells based on larger panels of sera (n-2710) by a study which showed a Dsn of 98.3% and Dsp of 94.2% (Moser et al. 1996). A solid-phase blocking ELISA based on E2 protein developed for detection of antibodies against CSFV exhibited a positive rate of 74.68% (Cao et al. 2022). A few other studies on E2 based indirect ELISA development showed a Dsn and Dsp of 97.5% and 96.7% (Lin et al. 2010), 96% and 95.3% respectively (Yang et al. 2012) and Dsp of 94.5% (Lin et al. 2005). Results of the E2 based blocking and indirect ELISAs exhibited an agreement rate of 94.7% (Cao et al. 2023), 92.2% (Ji et al. 2018) and 80.3% (Li et al. 2013) in comparison with the commercial kit (IDEXX, USA). Several studies based on immunochromatographic test strip for detecting CSFV antibodies showed sensitivity and specificity values nearly comparable to commercial ELISAs (Li et al. 2012; Sastre et al. 2016; Bai et al. 2022). Although the test strips are rapid and easy to perform, it is suitable only for herd level testing and not suitable for population surveillance while ELISA has higher throughput. In this study, the results of the E2-iELISA however showed high correlation with the results obtained in the PrioCHECK CSFV antibody kit on the same panel of sera. The E2-iELISA displayed a higher absolute sensitivity with an endpoint detection limit of 1:200 dilution of the serum.
The problem associated with the CSFV E2 is its sequence similarity with the BVDV E2 and thus may affect the accurate diagnosis of CSF, hence a large number of serum samples positive for BVDV antibodies need to be tested to substantiate the results (Kassimi et al. 1995; Huang et al. 2021). Notably, studies using immunochromatographic test strip (Bai et al. 2022; Huynh et al. 2024) with full-length CSFV E2 showed no cross-reactivity with BVDV E2. The lack of cross-reactivity of the E2-iELISA with the sera positive for FMDV, BVDV, ASFV and HS was also demonstrated, though assay could not be tested with sera positive for other swine diseases presenting one or more similar clinical signs.
In conclusion, the newly developed E2-iELISA has a diagnostic sensitivity and specificity which is comparable to the commercially available kit. Hence E2-iELISA is an alternative, inexpensive, and rapid serological diagnostic tool for sero-surveillance of CSF.
Data availability
No datasets were generated or analysed during the current study.
References
Bachan R, Bhattacharya S, Saini M, Rama T, Rana M, Gupta PK (2019) Expression of immunodominant regions of E2 from an Indian isolate of classical swine fever virus. J Anim Res 9(3):387–395. https://doi.org/10.30954/2277-940X.03.2019.1
Bai Y, Jia R, Wei Q, Wang L, Sun Y, Li Y, Luo J, Zhang G (2022) Development and application of a high-sensitivity immunochromatographic test strip for detecting classical swine fever virus antibodies. Transbound Emerg Dis 69(4):e788–e798. https://doi.org/10.1111/tbed.14367
Beer M, Reimann I, Hoffmann B, Depner K (2007) Novel marker vaccines against classical swine fever. Vaccine 25:5665–5670. https://doi.org/10.1016/j.vaccine.2006.12.036
Cao Y, Yuan L, Yang S, Shang Y, Yang B, Jing Z, Guo H, Yin S (2022) Establishment and application of a solid-phase blocking ELISA method for detection of antibodies against classical swine fever virus. J Vet Sci 23(5):e32. https://doi.org/10.4142/jvs.21262
Cao Z, Yin D, Zhang L, Ma S, Zhang K, Yang R, Shan H, Qin Z (2023) A novel blocking enzyme-linked immunosorbent assay based on a biotinylated nanobody for the rapid and sensitive clinical detection of classical swine fever virus antibodies. Microbiol Spectr 11(1):e02996–e02922. https://doi.org/10.1128/spectrum.02996-22
Clavijo A, Lin M, Riva J, Mallory M, Lin F, Zhou EM (2001) Development of a competitive ELISA using a truncated E2 recombinant protein as antigen for detection of antibodies to classical swine fever virus. Res Vet Sci 70(1):1–7. https://doi.org/10.1053/rvsc.2000.0434
Colijn E, Bloemraad M, Wensvoort G (1997) An improved ELISA for the detection of serum antibodies directed against classical swine fever virus. Vet Microbiol 59:15–25. https://doi.org/10.1016/S0378-1135(97)00178-8
Dong XN, Chen YH (2007) Marker vaccine strategies and candidate CSFV marker vaccines. Vaccine 25(2):205–230. https://doi.org/10.1016/j.vaccine.2006.07.033
Ganges L, Crooke HR, Bohórquez JA, Postel A, Sakoda Y, Becher P, Ruggli N (2020) Classical swine fever virus: the past, present and future. Virus Res 289:198151. https://doi.org/10.1016/j.virusres.2020.198151
Huang YL, Meyer D, Postel A, Tsai KJ, Liu HM, Yang CH, Huang YC, Berkley N, Deng MC, Wang FI, Becher P, Crooke H, Chang CY (2021) Identification of a common conformational epitope on the glycoprotein E2 of classical swine fever virus and Border Disease Virus. Viruses 13(8):1655. https://doi.org/10.3390/v13081655
Huynh LT, Sohn E-J, Park Y, Kim J, Shimoda T, Hiono T, Isoda N, Hong SH, Lee HN, Sakoda Y (2024) Development of dual immunochromatographic test strip to detect E2 and Erns antibodies against classical swine fever. Front Microbiol 15:1383976. https://doi.org/10.3389/fmicb.2024.1383976
Ji S, Luo Y, Zhang T, Shao L, Meng XY, Wang Y, Gao Y, Li Y, Li S, Sun Y, Jin X, Qiu HJ (2018) An improved indirect ELISA for specific detection of anti- bodies against classical swine fever virus based on structurally designed E2 protein expressed in suspension mammalian cells. Arch Virol 163:1831–1839. https://doi.org/10.1007/s00705-018-3809-7
Kassimi BL, Gonzague M, Ermel G, Plateau E, Crère C (1995) Partial sequencing of hog cholera virus Alfort strain genome and its comparison with other pestivirus strains. Vet Res 26:300–309
Kumar R, Barman NN, Khatoon E, Kumar S (2016) Development of single dilution immunoassay to detect E2 protein specific classical swine fever virus antibody. Vet Immunol Immunopathol 172:50–54. https://doi.org/10.1016/j.vetimm.2016.03.004
Lamothe-Reyes Y, Figueroa M, Sánchez O (2023) Host cell factors involved in classical swine fever virus entry. Vet Res 54(1):115. https://doi.org/10.1186/s13567-023-01238-x
Li X, Wang L, Shi X, Zhao D, Yang J, Yang S, Zhang G (2012) Development of an immunochromatographic strip for rapid detection of antibodies against classical swine fever virus. J Virol Methods 180(1–2):32–37. https://doi.org/10.1016/j.jviromet.2011.12.006
Li W, Mao L, Yang L, Zhou B, Jiang J (2013) Development and partial validation of a recombinant E2-based indirect ELISA for detection of specific IgM antibody responses against classical swine fever virus. J Virol Methods 191:63–68. https://doi.org/10.1016/j.jviromet.2013.03.003
Lin M, Trottier E, Mallory M (2005) Enzyme-linked immunosorbent assay based on a chimeric antigen bearing antigenic regions of structural proteins erns and E2 for serodiagnosis of classical swine fever virus infection. Clin Vaccine Immunol 12(7):877–881. https://doi.org/10.1128/CDLI.12.7.877-881.2005
Lin GZ, Qiu CQ, Zheng FY, Zhou JZ, Cao XA (2008) Secretory expression of E2 main antigen domain of CSFV C strain and the establishment of indirect ELISA assay. Virol Sin 23:363–368. https://doi.org/10.1007/s12250-008-2970-7
Lin GZ, Zheng FY, Zhou JZ, Cao XA, Gong XW, Wang GH, Qiu CQ (2010) An indirect ELISA of classical swine fever virus based on quadruple antigenic epitope peptide expressed in E. Coli. Virol Sin 25:71–76. https://doi.org/10.1007/s12250-010-3086-4
Luo Y, Li S, Sun Y, Qiu HJ (2014) Classical swine fever in China: a minireview. Vet Microbiol 172:1–6. https://doi.org/10.1016/j.vetmic.2014.04.004
Ma Z, Zhao Y, Lv J, Pan L (2023) Development and application of classical swine fever virus monoclonal antibodies derived from single B cells. Vet Res 54:90. https://doi.org/10.1186/s13567-023-01229-y
Malik YS, Bhat S, Kumar OV, Yadav AK, Sircar S, Ansari MI, Dhama K (2020) Classical swine fever virus biology, clinicopathology, diagnosis, vaccines and a meta-analysis of prevalence: a review from the Indian perspective. Pathogens 9(6):500. https://doi.org/10.3390/pathogens9060500
Meyers G, Thiel HJ (1996) Molecular characterization of pestiviruses. Adv Virus Res 47:58–118. https://doi.org/10.1016/s0065-3527(08)60734-4
Moennig V, Becher P (2015) Pestivirus control programs: how far have we come and where are we going? Anim Health Res Rev 16:83–87. https://doi.org/10.1017/s1466252315000092
Moser C, Ruggli N, Tratschin DJ, Hofmann MA (1996) Detection of antibodies against classical swine fever virus in swine sera by indirect ELISA using recombinant envelope glycoprotein E2. Vet Microbiol 51:41–53. https://doi.org/10.1016/0378-1135(96)00019-3
Nath MK, Sarma DK, Das BC, Deka P, Kalita D, Dutta JB, Mahato G, Sarma S, Roychoudhury P (2016) Evaluation of specific humoral immune response in pigs vaccinated with cell culture adapted classical swine fever vaccine. Vet World 9(3):308. https://doi.org/10.14202/vetworld.2016.308-312
Patil SS, Hemadri D, Shankar BP, Raghavendra A, Veeresh H, Sindhoora B, Chandan S, Sreekala K, Gajendragad MR, Prabhudas K (2010) Genetic typing of recent classical swine fever isolates from India. Vet Microbiol 141:367–373. https://doi.org/10.1016/j.vetmic.2009.09.021
Postel A, Austermann-Busch S, Petrov A, Moennig V, Becher P (2018) Epidemiology, diagnosis and control of classical swine fever: recent developments and future challenges. Transbound Emerg Dis 65:248–261. https://doi.org/10.1111/tbed.12676
Sastre P, Pérez T, Costa S, Yang X, Räber A, Blome S, Goller KV, Gallardo C, Tapia I, García J, Sanz A (2016) Development of a duplex lateral flow assay for simultaneous detection of antibodies against African and classical swine fever viruses. J Vet Diagn Invest 28(5):543–549. https://doi.org/10.1177/1040638716654942
Wang L, Madera R, Li Y, McVey DS, Drolet BS, Shi J (2020) Recent advances in the diagnosis of classical swine fever and future perspectives. Pathogens 9(8):658. https://doi.org/10.3390/pathogens9080658
Wei Q, Liu Y, Zhang G (2021) Research progress and challenges in vaccine development against classical swine fever virus. Viruses 13:445. https://doi.org/10.3390/v13030445
Wensvoort G, Terpstra C, Boonstra J, Bloemraad M, Van-Zaane D (1986) Production of monoclonal antibodies against swine fever virus and their use in laboratory diagnosis. Vet Microbiol 12:101–108. https://doi.org/10.1016/0378-1135(86)90072-6
Wong ML, Liu JJ, Chang YS, Chang TJ (1998) Expression of the glycoprotein E2 of the classical swine fever virus in Escherichia coli. J Vet Med Sci 60:541–544. https://doi.org/10.1292/jvms.60.541
Yang ZH, Li L, Pan ZS (2012) Development of multiple ELISAs for the detection of antibodies against classical swine fever virus in pig sera. Virol Sin 27:48–56. https://doi.org/10.1007/s12250-012-3227-z
Yi W, Zhu H, Wu Y, Li Q, Lou W, Zhao H, Pan Z (2022) The recombinant erns and truncated E2-based indirect enzyme-linked immunosorbent assays to distinguishably test specific antibodies against classical swine fever virus and bovine viral diarrhea virus. Virol J 19(1):121. https://doi.org/10.1186/s12985-022-01851-w
Yuan M, Yang X, Zhang X, Zhao X, Abid M, Qiu HJ, Li Y (2023) Different types of vaccines against pestiviral infections: “barriers” for “pestis.” Viruses 15:2. https://doi.org/10.3390/v15010002
Acknowledgements
The authors acknowledge the Director, ICAR-NIVEDI, Bengaluru, for facilitating this study.
Funding
The research was funded by Indian Council for Agricultural Research (ICAR).
Author information
Authors and Affiliations
Contributions
SSP and MH contributed to the research study conception and design. SG, MH and SSP performed the laboratory studies and analysis. SG, MH, SSP, BVJ drafted the manuscript. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval
The animal study was reviewed and approved by the Institutional Animal Ethics Committee (NIVEDI/IAEC/2022/2021-01E).
Consent to participate
Not applicable.
Consent to publish
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Gopinath, S., Hosamani, M., Joseph, B.V. et al. Development of classical swine fever virus E2-protein based indirect ELISA for detection of antibodies against the virus in pigs. Vet Res Commun (2024). https://doi.org/10.1007/s11259-024-10482-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11259-024-10482-1