
Abstract
In some internal valleys of the northern Andes, precipitation at treeline elevation is seasonal, with one or two dry periods that could exert a strong control on tree seedling survival. However, little is known about (1) the severity of the dry periods, due to a lack of climate data, or (2) the effect of drought on tree seedling demographic dynamics and how this is modulated by local plant interactions in these tropical-alpine ecosystems. We studied annual precipitation variation and seedling demography and ecophysiology in relation to microclimate and neighboring plants in a dry treeline ecotone at 3100 m a.s.l in the Venezuelan Andes. We selected seedlings of the dominant tree species Cybianthus marginatus and Clusia multiflora at three sites, each with three adjacent 10 × 10-m plots within the forest, forest border, and páramo. For both species, seedling density was zero in the páramo and did not differ between border and forest. Growth rates were low for both species, but higher for Cybianthus in the border than in the forest. Survival rates were high in both environments. Low assimilation and transpiration rates were found in both species regardless of the environment. The spatial association of seedlings with moisture-regulating elements such as mosses, ferns and basal rosettes may indicate a benefit of better hydric conditions in their neighborhood. Our study shows a strong limitation for tree establishment in the páramo, but high rates of survival at the edge of the forest. This suggests a scenario of slow potential forest expansion at this tropical treeline, tree cover advancing in a closed front by the successive formation of forest-edge vegetation allowing tree seedling establishment outside of the forest.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In alpine treeline ecotones, tree seedlings are commonly found above the closed forest mixed in with the low-stature alpine vegetation, but these seedlings rarely make it into adulthood; most die shortly after establishment. High seedling mortality can be caused by several factors, including desiccation, overheating, photodamage, pathogen attack and nightly frost (Lett and Dorrepaal 2018). Newly established seedlings are disproportionately affected by these factors, provoking mortality rates that can exceed 90% during the first year of the cohort (Maher and Germino 2006; Batllori et al. 2009). After that, mortality tends to decrease as seedlings become juveniles (Maher and Germino 2006; Brodersen et al. 2019). Such high seedling mortality rates are not uncommon in tree populations, but as viable seeds are usually available in limited numbers at treeline, seedling mortality is particularly critical for treeline dynamics (Lett and Dorrepaal 2018; Loranger et al. 2016; Brodersen et al. 2019).
Tree establishment in open areas above continuous forest usually depends not only on seed production and dispersal but also on the availability of ‘safe sites’ (sensu Harper 1977): sites with suitable conditions for seed germination, seedling development and long-term survival. Safe sites often involve sheltered conditions, including shade or improved soil chemistry, moisture and thermal regimes. Such conditions can be generated by structures like rocks, dead stems (Johnson and Yeakley 2016), microtopography (Resler et al. 2005) or by nurse plants (Germino et al. 2002; Llambí et al. 2013; Bueno and Llambí 2015).
Establishing tree seedlings are potentially limited by a wide range of environmental factors. Due to their small size, limited reserves, and lack of protective structures compared with adults, tree seedlings are more sensitive (Germino et al. 2002; Graae et al. 2011; Johnson and Yeakley 2016). However, most of the classic treeline literature has focused on the responses of adult trees, correlating their current distribution or past growth and population dynamics (based on dendrochronological techniques) to spatial and temporal climate patterns (Holtmeier and Broll 2020). Recently though, seedling establishment patterns and physiological responses are increasingly emphasized in approaches to understand treeline ecotone vegetation dynamics (see Lett and Dorrepaal 2018 for a review). Some authors have addressed specific physiological causes of establishment limitations, such as radiation damage (photoinhibition; Germino and Smith 1999), low-temperature limitation of carbon sink processes (Bansal and Germino 2010) and lack of freezing tolerance (Piper et al. 2006; Rehm and Feeley 2015). These studies focus on temperature and radiation as the main abiotic drivers in the establishment of new recruits at the treeline ecotone, while the effect of water availability on the recruitment dynamics is described as “strongly dependent on local hydrology” (Lett and Dorrepaal 2018) and as “species-specific” (Loranger et al. 2016). Decreased seedling survival under experimental warming has also been observed and attributed to water stress (Moyes et al. 2013; Conlisk et al. 2018).
Neighboring plants may affect seedling physiological responses by modifying the driving environmental factors. Spatial associations between seedlings and adults can indicate how these modifications influence seedling emergence or survival (Germino et al. 2002; Llambí et al. 2013). Both positive and negative roles of neighboring trees, shrubs, mosses and grasses on demographic processes at treeline have been documented in temperate-alpine systems (Maher and Germino 2006; Ninot et al. 2008; Batllori et al. 2009, 2010; Loranger et al. 2017; Lett et al. 2017, 2020; Sidgel et al. 2020). Some of these studies also present a mechanistic approach studying how photosynthetic efficiency is enhanced by the association (Maher et al. 2005; Germino et al. 2002) or how the reduction in sunlight exposure and positive seedling response, as indicated by its pigmentation, may explain the emergence of spatial associations (Akhalkatsi et al. 2006). A high density of shrubs or graminoids has also been associated with low occurrence of tree seedlings, indicating competition (Ninot et al. 2008; Batllori et al. 2009), and has been experimentally shown to reduce seedling survival (Loranger et al. 2017). The direction and magnitude of the effect of neighboring vegetation on the performance and survival of tree seedlings in alpine treeline ecotones thus appears to vary among sites, likely due to differences in the species and climate involved.
Tropical alpine environments (referred to as páramo in the northern Andes) have unique characteristics, and the conditions that prevail in these environments can cause different kinds of physiological stress for plants: daily thermal oscillations are more pronounced than seasonal ones and can generate thermal stress by high and low temperatures; incoming radiation is generally very high, particularly on clear days, and can cause light stress; at high elevations, soils are rocky and shallow, resulting in a low water-retention capacity which, in combination with seasonality in precipitation, can result in drought stress. Moreover, these undeveloped soils (entisols, inceptisols) may provoke mineral-nutrition stress (Azócar and Rada 2006; Körner 2003; Monasterio 1980; León et al. 2015; Rada et al. 2019). Precipitation patterns are extremely variable in tropical alpine environments because they not only depend on elevation but are modified by wind patterns unique to the geography of each mountain system. As a general rule, the amount of rainfall increases up to a maximum at middle elevations (around 2000–2500 m) and decrease again at higher elevations (Sarmiento 1986). Depending on the geographical location, the precipitation seasonality can be either unimodal (one rainy season and one dry season) or bimodal (two rainy seasons and two dry seasons) (Sarmiento 1986; Rundel 1994). Dry seasons can also show ample variability in their intensity, some páramos are permanently wet, while others experience real dry seasons. During such dry seasons, páramo plants face high evaporative demands generated by high radiation input and low relative humidity, combined with low soil water contents (see Sarmiento 2000; Llambí et al. 2003), which can lead to a constraint in productivity and probably seedling establishment (Rada et al. 2019; García-Nuñez et al. 2004; Cavieres et al. 2000).
The lower boundary of tropical alpine ecosystems is defined by the treeline ecotone, the transition between the upper-montane forest and the alpine vegetation. This ecotone constitutes an important ecological belt in tropical mountains due to its high biodiversity, including a mixture of alpine and forest species but also species characteristic of the treeline, including many tall shrubs and small trees (Ramírez et al. 2009; Hofstede 2003; Llambí 2015). Globally, treeline elevations depend on growth temperatures (Körner 1998, 2021), while locally other environmental factors such as wind, herbivory or water availability can also play important roles (Holtmeier and Broll 2020; Moyes et al. 2013; Malanson et al. 2019). Due to the strong link to climate, treeline ecotones may be sensitive to environmental change and may act as an indicator for such change (Arzac et al. 2019; Chacón-Moreno et al. 2021). Such a biotic indicator would be particularly useful in tropical mountains, since climatic records are generally lacking at such high elevations (but see Bendix et al. 2017). However, the comprehension of treelines as indicators is severely hampered by our limited understanding of the processes that determine high-mountain vegetation dynamics, especially in the tropics.
Regarding seedling establishment in tropical treeline ecotones, Bader et al. (2007a) found that seedling densities were highest just above the forest edge and decreased in the páramo and that only radiation-tolerant species could successfully establish outside the forest canopy. This suggests that high solar radiation at tropical high elevations can limit tree establishment and regeneration above this tropical treeline in Ecuador. Additionally, in that study area the recurrent páramo fires probably prevent succession via more radiation-tolerant shrub and tree species. In Colombia, Ramos and García (2008) described survival patterns in seedlings of two tree species from the treeline ecotone, showing that the highest mortality occurred during the dry season. In Peru, Rehm and Feeley (2015) conducted an experiment to evaluate the effect of temperature (night-time warming) and radiation (shading) on seedlings of three dominant tree species transplanted along an elevational gradient. They report contrasting and species-specific responses in the survival of the seedlings. Apparently, the response to climate drivers in these open alpine ecosystems differs strongly within the diverse sets of tree species present at tropical cloud-forest treelines.
Treeline ecotone vegetation in the Venezuelan Andes is relatively well studied (see review by Llambí 2015), with a focus on the Cordillera de Mérida, mainly on the humid slopes in the Sierra Nevada National Park (Yanez 1998; Tirado 1997; Cáceres and Rada 2011; Puentes 2010; Ely et al. 2011; Ramírez et al. 2009; Llambí et al. 2013; Arzac et al. 2019). However, there have been only few floristic and ecological studies on seasonally dry slopes (Abele 2000; Sodja 2001; Dulhoste 2010; Rodríguez et al. 2011). The existing body of research in the Venezuelan Andes indicates that: (i) the ecotone position varies between 2800 and 3650 m a.s.l, with lower treelines on dry slopes than on humid slopes, suggesting that precipitation amounts and/or seasonality could be important determinants of tree establishment processes (Suárez del Moral and Chacón-Moreno 2011); (ii) diversity and turnover of species and life forms along the ecotone are high, and so are floristic differences between slopes, indicating high alpha, beta and gamma diversity (Rodríguez et al. 2011; Llambí et al. 2014; Arzac et al. 2019); (iii) strong fluctuations in temperature (Rada et al. 2011) and high incoming radiation (Puentes 2010) seem to be important abiotic constraints for the establishment of tree species above the forest; (iv) páramo shrubs can act as nurse plants, being associated with increased local densities of pioneer trees in the ecotone, which may, in turn, facilitate the establishment of other tree species, both in undisturbed páramos (Llambí et al. 2013) and in successional old fields (Bueno and Llambí 2015).
All these previous studies indicate that, on humid slopes, radiation and thermal stress could be important in preventing tree seedling establishment in open areas in the páramo, and that local plant-plant interactions could significantly promote it. However, there is no information about survival, growth rates, or physiological performance of tree seedlings in treeline ecotones in dry Andean enclaves, or about how biotic interactions modify the establishment patterns there. As environmental conditions are rather different on dry slopes for at least part of the year, the limitations for tree establishment may be stronger than on humid slopes, water stress likely playing a more important role. Moreover, due to the harsher conditions, we also expect positive local associations with neighboring nurse plants to be more pronounced than on the wetter slopes.
In this paper we analyze the relationship of seedling demography with microclimate and neighboring plants of the tree species Cybianthus marginatus and Clusia multiflora in a dry tropical treeline ecotone in the Venezuelan Andes. These two species have been previously described as dominant in the study area. C. marginatus is characterized as “fairly abundant” in the interior of the forest tongues and in borders, and even some juveniles were registered in the páramo. In contrast, C. multiflora individuals are reported only in the interior of the forest tongues and in the border (Rodriguez et al. 2011). Nevertheless, adult individuals of both species can be found in forest islands in the páramo (personal observation). We address the following questions: (1) How do the density, survival and growth rates of the seedlings of these two species vary along the treeline ecotone? (2) Are there any local association patterns of the seedlings with vegetation elements that could indicate positive or negative plant-plant interactions? And (3) Do seedlings established in different environments along the treeline ecotone exhibit differences in their photosynthesis rates, transpiration, and stomatal conductance? What do these potential differences in physiological performance indicate about the role of water stress as a limiting factor for seedlings across the treeline ecotone? In addition, we started to characterize the annual precipitation patterns in our study area to quantify the stress that the seedlings may be exposed to, and as a baseline for detecting future changes.
Available climate records and future climate projections for many regions across the tropical Andes (including western Venezuela), indicate significant temperature increases and changes in rainfall patterns, including lower total annual rainfall in some cases and longer dry seasons in most areas (Urrutia and Vuille 2009; Buytaert et al. 2011; Braun and Bezada 2013; Chacón-Moreno et al. 2021). Hence, a better understanding of ecological processes in seasonally dry mountain ranges is fundamental to model and predict future vegetation development, not only in dry Andean enclaves, but also in currently more humid mountain regions (Moyes et al. 2015). More generally, understanding tree establishment at these tropical treelines will also improve our understanding of the variation in treeline controlling processes and dynamics in different climates around the world.
Methods
Study area
This study was carried out in the San José páramo (3120 m a.s.l. 08° 19′ N; 71° 19′ W), in the southern portion of the Sierra Nevada de Mérida in the Venezuelan Andes (see Fig. 1). It was selected because of its climate and relatively easy accessibility. As a consequence of its inter-Andean position, the site lies in a rain-shadow area (the Nuestra Señora semi-arid enclave) and has been described as a dry páramo (Aranguren et al. 2012). At the closest climatic station (Tostós. 2400 m a.s.l. ca. 10 km from our study site), mean annual precipitation is 547 mm and the mean annual temperature is 13.7 °C. That station reports a marked seasonality in rainfall across the year, with two dry periods: the main dry season between November and March and a second, shorter one, between June and August (Costa Talens et al. 2007; Anairamiz et al. 2013). On these dry slopes of the Venezuelan Andes, in spite of some fog occurrence (see Fig. 3), cloud forests are replaced by dry evergreen forests (Ataroff and Sarmiento 2004). The treeline ecotone occurs between 2800 and 3100 m a.s.l, one of the lowest elevations for natural forest-páramo transition zones in the Northern Andes (Suárez del Moral and Chacón-Moreno 2011). Despite the lack of evidence of intensive land use in recent decades in this area, it is possible that past fire events or other anthropogenic activities have influenced the current treeline ecotone position. However, cattle grazing is present but very scarce in the area, fire is not commonly used as a land management tool, and there is no evidence of other land use activities (e.g. agriculture).

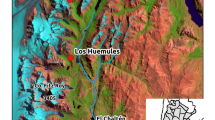
Distribution of the treeline ecotone in the Venezuelan Andes and location of the study area (San José), the city of Mérida and the more humid and well-studied treeline site La Aguada. Map by E. Chacón-Moreno
The treeline vegetation in the study area shows a heterogeneous pattern with forest tongues ascending into a páramo matrix (Appendix 1 in ESM). Rodríguez et al. (2011) described the tree community structure for the forest tongues. They reported 30 woody species, with Cybianthus marginatus as the dominant tree in the large categories (DBH > 2.5 cm) and Clusia multiflora as the dominant species in the small categories (DBH < 2.5 cm), though this species can also grow into large trees. Other tree species include Miconia tinifolia, Macleania rupestris, and Libanothamnus neriifolius, and tall shrubs include Chaetolepis lindeniana and Diplosthephium venezuelense, which occur in low densities in the forest but become more dominant in forest borders and páramo areas. The lower stratum shows a high relative abundance of herb species (different genera: Piper, Peperomia, Lachemilla, Rubus, Psychotria), epiphytic Bromeliaceae and Orchideae, and some páramo shrubs in low proportion. The bottom stratum is strongly dominated by mosses and ferns.
The páramo vegetation above the continuous forest line is dominated by tussock grasses (Calamagrostis), but other growth forms are also important including shrub species (C. lindeniana, Valeriana parviflora, Hypericum laricifolium, H. stenopetalum), stem and basal rosettes from the Espeletinae tribe like Espeletia schultzii and Ruilopezia artropurpurea, and non-graminoid herbs. At the contact edge between forest and páramo, the forest border consists of a mix of growth forms (1–2 m tall) dominated by tall shrubs (e.g. C. lindeniana. D. venezuelense), below which basal rosettes and prostrate shrubs are found, mixed with Chusquea spencei, a shrub-bamboo associated with undisturbed treeline forests and shrublands in the páramo (Ely et al. 2011). The bottom stratum consists of a mix of prostrate herbs, ferns and mosses. The border is structurally quite heterogeneous: densely covered patches are interspersed with more open microsites (referred to in this study as border-gaps).
Study species
Given their importance as a dominant tree species in the forest tongues, we focused our study on the following two species:
Cybianthus marginatus (Benth.) Pipoly (Primulaceae) (Cybianthus hereafter) is a small tree around 6 to 10 m tall with coriaceous, symmetrical and lanceolate leaves, distributed in páramos and upper Andean forests throughout the Andes from Venezuela to Peru (Hokche et al. 2008). Seeds are dispersed by animals (Correa-Gómez et al. 2013).
Clusia multiflora Kunth (Clusiaceae) (Clusia hereafter) is an evergreen tree species reaching heights up to 20 m, with oval and succulent leaves distributed from lowland forest to upper Andean forest (80–3500 m) from Mexico to Bolivia (Hokche et al. 2008; Popp et al. 1987). Seeds are dispersed by animals (Ortis and Umba 2010; Manrique and Morales 2016). Some species of this genus are known to use the Crassulacean Acid Metabolism (CAM) as an alternative photosynthesis pathway, but C. multiflora has been reported as a C3 plant (Popp et al. 1987).
Sampling design and data analysis
Three replicate rectangular vegetation plots of 600 m2 (30 × 20 m), separated from one another by around 500 m, were selected in the contact zone between forest and páramo. Plots were divided into three subplots (10 × 20 m each) in three environments: páramo, border and forest (Fig. 2a). In each of these plots we monitored the microclimate and evaluated seedling abundance and ecophysiological responses (see below).

Sampling design: a Dimensions of the vegetation plot, divided into three environments (expected differences in seedling densities are drawn for reference). b Dimensions and relative positions of the microplots used for characterizing the local neighborhood of seedlings. The paired random plots were placed 1 m from the seedling-centered plots, with the exact location chosen randomly
Climate monitoring
A climate station was installed to describe the general climate in the study area, as the nearest climate station was around 10 km away and located 720 m lower, at 2400 m, and there are no climatic records since January 1991. Due to strong topographic effects on local climates in the northern Andes, it was highly uncertain to what extent this previously existing station could adequately describe the conditions in our study area. Moreover, climate data from similar dry treelines in the northern Andes do not exist. We registered precipitation (Davis 0.2 mm Rain Gauge Smart Sensor S-RGF-M002), solar radiation (HOBO S-LIB-M003), air temperature (Temp) and relative humidity (RH; HOBO U23 Pro v2) every hour from January 2019 to February 2020. All the sensors were installed at 1.5 m from the ground in an open area at 3150 m a.s.l. about 500 m uphill from the forest edge.
Additionally, in order to characterize the microclimate for seedlings, Temp/RH sensors (HOBO U23) were installed as follows, replicated in the three vegetation plots: one sensor in the páramo zone (open space), two sensors in the border zone (under vegetation cover (“border-cover”) and in small open microsites (“border-gaps”)) and one in the forest zone (understory). All sensors were installed at 50 cm above the ground. Those in cover situations were protected from direct radiation by the shrubs or trees, and those in open situations in páramo and border-gaps with a white plastic cover. Each logger recorded data every hour from January 2019 to February 2020, but due to the loss of data in some of the sensors, we compared the climate data from the different microhabitats only for the period from August 2019 to February 2020, which includes periods of both the wet and dry season.
Precipitation and temperature data were used to construct a climate diagram (after Walter and Lieth 1960), which, although representing only one year of climate data, provides valuable information about the climate in this type of environment for which information is very scarce. Additionally, we graphically compared temperature and relative humidity in the different microenvironments (páramo, border-gaps, border-cover and forest).
Seedling distribution
To evaluate seedling survival rates, we searched for seedlings of the two selected species in all subplots, including individuals with sizes from 5 to 50 cm height. For Clusia no seedlings < 5 cm were found, for Cybianthus we excluded these very small seedlings because of the difficulty in identifying them to the species level and in tagging them. We thus follow the definition of seedlings as the “established seedlings” in Brodersen et al. (2019), because all of our individuals had already more than 4 leaves (and probably had lost their cotyledons), which allows them to support themselves by their photosynthetic activity and to develop a root system. Even if this bypasses the very first, sensitive life stage (Brodersen et al. 2019), this slightly later stage is still critical in the life of a tree, especially in environments with drought conditions (Moles and Westoby 2004).
We labeled each seedling with a permanent tag and recorded the size, health status and number of leaves of all labeled seedlings during January/February 2019 to establish a baseline. Resurveys were done in August 2019 and January 2020 to determine seedling survival, height growth, and changes in the health status (registering signs of herbivory, chlorosis or pigmentation in the leaves). In the case of Cybianthus, the seedling numbers were rather high, so that in subplots with over 30 seedlings, only a random subset of 30 seedlings (see Appendix 2 in ESM) was labeled and registered. For Clusia all seedlings in the plots were labeled.
To test for differences in growth rates between environments (forest and border) for Clusia and Cybianthus, we used t tests. We calculated the relative growth rate (RGR) using the height of each individual measured in January 2019 and the year after. For this, we used the equation:
where t1 is 2019 and t2 is 2020. RGR is thus expressed in cm cm−1 y−1.
We evaluated differences in survival rates and health status of the seedlings between environments graphically based on the proportion of seedlings alive, dead, with sign of herbivory, chlorosis and with different pigmentations (brown, red or purple leaves) for the two resurveys (August 2019 and January 2020). Only in a few cases (three individuals of Clusia, and four of Cybianthus) the seedling was classified in two of the categories, i.e. with signs of herbivory and other pigmentation, and these are presented in both categories. These differences were not tested statistically, since the values in some subplots were based on very few individuals, thus not allowing a fair subplot-based mean.
To register spatial associations between the target species and vegetation elements in their immediate neighborhood, we visually estimated the cover of all plant species belonging to the lower stratum surrounding target seedlings, using circular microplots 30 cm in diameter (700 cm2 in area). We established paired microplots in nearby random locations to register plant cover, providing a random control or “null model” for the expected cover around the seedlings. We selected each seedling randomly from the previously labeled set, and the paired random microplot was located between 1 and 2 m away from the seedling in a random direction (see Fig. 2b). As vegetation patterns are fine-grained and as our interest was to analyze the potential local effect of neighbors and not of abiotic heterogeneity in the environment, this distance was considered sufficient to assure independence of the vegetation cover from that in the seedling microplot. In each microplot we visually estimated the cover of each vascular plant species and of the different ground covers found (mosses, lichens and litter). The total number of pairs of microplots were: for Clusia, 21 in border and 24 in forest; and for Cybianthus, 42 in border and 43 in forest.
To establish whether there were spatial associations between the seedlings and other vegetation elements from the immediate neighborhood, we collapsed our community matrix (species cover per microplot) into a life form matrix, classifying and summing the cover of each species according to the following categories: graminoids (like Chusquea sp, Neurolepis sp, Festuca sp), herbs (Geranium sp, Peperomia sp, Piper sp and others), basal rosettes (Ruilopezia atropurpurea), ferns and woody plants (including shrubs like Gaultheria myrsinoides, Chaetolepis lindeniana and small trees found in the understory like Miconia sp). Then, for each seedling of both target species (separately) and its random microplot pair we calculated the Relative Interaction Index (modified from Armas et al. 2004) using the following equation:
VC (sc) is the cover of a given life form in a seedling-centered microplot and VC (rp) is the cover of the same life form in the corresponding random microplot. Spatial associations were considered significant when the confidence interval of the RII did not overlap the zero line, with values > 0 indicating positive and < 0 negative associations between seedlings and the life form in question. We followed the same approach to detect spatial associations with the different ground cover types (mosses, lichens and litter). To avoid misreading the results, i.e. false positives or negatives associations produced by low replication, we excluded from the analysis those life forms categories that were present in less than 25% of the total samples (i.e. < 6 replicates for Clusia and < 11 replicates for Cybianthus).
Clusia seedling density was based on the total count of seedlings in each subplot. For Cybianthus, where densities were higher and not all seedlings were recorded in the subplots, densities were determined based on the occurrences in the microplots in each environment.
To detect whether there are differences in seedling densities between environments (border vs forest) we performed a paired t-test for each species (based on densities per subplot, for Clusia as total densities, and for Cybianthus calculated based on densities in the microplots). Additionally, to detect intraspecific seedling clustering at small scales, we compared densities in paired random microplots and those centered around seedlings for each species (not counting the central seedlings) using paired t tests.
Ecophysiological measurements
We selected seedlings of each target tree species established in the understory of the forest and in the forest border in each of the three replicate study sites (no seedlings were found in the páramo). In the forest border we studied two local situations: under cover (shrubs or grass, low radiation levels) and in gaps, with higher radiation levels. The sample size was selected based on seedling availability. For Cybianthus, six individuals were randomly selected in each situation: forest, border-gaps and border-cover. For Clusia, five individuals were used inside forests and in the border under cover, and two individuals in the gaps in the border (no more individuals were found in this situation).
For each seedling we determined: leaf temperature, photosynthetic photon flux density (PPFD) reaching the leaves, transpiration (E), stomatal conductance (Gs), carbon exchange rate (assimilation: A), ratio between internal and atmospheric carbon concentration (Ci/Ca), and water use efficiency (WUE). Additionally, as an indicator of the strength of the moisture gradient between the leaf and the surrounding air, we calculated leaf-to-air vapor pressure difference (leaf-air VPD) for each seedling (after Lambers et al. 2008). For E, Gs, A and Ci/Ca, we used an infrared gas-exchange system (Lci. ADC Ltd. Hoddesdon. UK), setting cuvette conditions to ambient values. WUE was defined as the ratio between carbon gain in photosynthesis and water loss in transpiration (A/E) (Lambers et al. 2008). For vapor pressure difference calculations, we measured leaf temperature (Tl) and air temperature (Ta), placing a thermocouple on the abaxial surface of the leaf (for Tl) and 10 cm away from the leaf (for Ta) (tele-thermometer 04B0618 409 YSI. Yellow Springs, OH, USA). We also measured air relative humidity near leaves with a thermo-hygrometer (OMEGA RH650). Measurements were repeated three times for each seedling at two moments during the day (11:00 and 13:00) during the dry season of 2019. Since no differences between the points in time were detected the values presented are based on the means of these six measurements per seedling (i.e. only one value per parameter is used per seedling). We intended to also measure plant water potentials as an indicator of drought stress, but due to technical difficulties we could not complete the minimum number of replicates needed for analysis (part of the equipment was damaged during travel to the site, and additional visits were not possible due to the current challenges with logistics in Venezuela).
To assess whether there were differences in the physiological responses between seedlings in the different environments (border-gaps, border-cover and forest), we compared the values of each response variable (leaf temperature; leaf-air VPD; PPFD; E; Gs; A; WUE and Ci/Ca) in each situation using ANOVAS (separately for each species) and in cases where the normality test failed, we used a Kruskal–Wallis test.
We used SigmaPlot v11 for all the graphs and statistical analysis.
Results
Climate
Total precipitation at the San José Páramo (3120 m a.s.l) during 2019 was 1012 mm, distributed following a bimodal pattern: there was a dry season during January and February and a humid season with two peaks of high rainfall, a small one in April and a larger one around August (Fig. 3a). During the dry season, four periods with more than 10 consecutive days without any precipitation were recorded (with the longest period counting 13 days). The driest month was February, with 4.6 mm of rainfall, and the wettest month was August, with 161.8 mm. Mean annual air temperature (at 1.5 m from the ground) was 9.4 °C (Appendix 3 in ESM). As expected, mean monthly air temperatures were rather constant through the year (Fig. 3b), while daily fluctuations (of about 10 °C) were much larger. Maximum and minimum temperature values showed some annual variation, with a broader thermal amplitude during the dry season months (Fig. 3b upper panel). No freezing air temperatures were recorded at any time of year (Appendix 3 in ESM, but see Fig. 4 for freezing records at 50 cm in some microsites). Relative humidity showed high mean and maximum values throughout the year, but the mean daily minimum values dropped markedly, to about 60%, during the dry season (Fig. 3b middle panel) and then even reached absolute minimum values near 20% (Appendix 3 in ESM). Mean radiation values were also relatively constant through the year, but the mean daily maxima tended to be lower during the humid months (between August and November) (Fig. 3b bottom panel). Monthly radiation maxima were very high (Appendix 3 in ESM).

Climate of the treeline ecotone in San José páramo in the Venezuelan Andes recorded during the period February 2019–January 2020: a Climate diagram. Upper line: location, elevation, mean annual temperature, total annual precipitation. Yellow area: dry months. Red line: monthly average temperature. Bars: total precipitation per month. Note that above 100 mm monthly precipitation, i.e. for very humid months, the scale of the precipitation axis changes. b Monthly temperature (°C, upper panel), relative humidity (%, middle panel) and radiation (W m−2, lower panel) recorded by the climate station. Black circles: absolute maximum, white circles: mean daily maximum, black triangles: mean, white triangles: mean daily minimum, black square: absolute minimum

Microclimatic conditions (at 50 cm from the soil) in forest, border (under cover (border_cover) or in small open microsites (border_gaps)), and páramo plots in the treeline ecotone in San José páramo (3120 m a.s.l.) in the Venezuelan Andes: temperature (°C) (panel a) and relative humidity (%) (panel b). From top to bottom: páramo, border-gaps, border under cover and forest understory. Red lines: daily maxima, blue lines: daily means, gray lines: daily minima. P1: plot 1, P2: plot 2, P3: plot 3. Data were recorded every 10 min between August 2019 and February 2020. The humid season lies in the period between August and December; the dry season corresponds to the period between January and February
The three replicate temperature measurements at 50 cm from the ground were very similar within each environment (páramo, border-gaps, border-cover and forest) but differed clearly between environments (Fig. 4a). Although mean temperatures were similar (ca. 8.7 °C) among the four environments, daily fluctuations were larger in the páramo and border-gaps than in the understory of the forest and in covered areas of the forest border (Fig. 4a). This tendency was accentuated during the dry season, during which the minimum temperatures in border-gaps reached − 1 °C while they remained above 0 °C within the forest canopy. Surprisingly, the minimum temperatures in the páramo never reached values below 0 °C. The maximum temperature value was also recorded in border-gaps (see Appendix 3 in ESM). In general, the highest mean daily thermal amplitude was registered in páramo and border-gaps and was reached during the dry season in January.
Maximum relative humidity reached 100% almost every day in all environments. The daily means were also generally high while daily minima showed dry spikes most of the period, except in the very wet month of August (Fig. 4b). Although during the dry season low values were more frequent in all four microenvironments, fluctuations were more drastic in páramo and in border-gaps and most dampened in the forest during almost all of the measurement period (Fig. 4b).
Seedling density, growth and survival
No tree seedlings of our two-target species were found in the páramo, either in our plots or the surroundings. The only tall woody species found in the páramo matrix was Diplostephium venezuelense (ca. 20% cover), besides other smaller páramo shrubs (Chaetolepis lindeniana, Hypericum stenopetalum). Clusia seedling densities were very low both in the border and in the forest, with values of 0.05 (± 0.04) ind m−2 and 0.06 (± 0.02) ind m −2, respectively, and there were no statistical differences between these environments (paired t-test, n = 3, p = 0.62). Seedling densities of Cybianthus were similar between environments, with values of 15.7 (± 7.1) ind m−2 in the border and 12.9 (± 4.0) ind m−2 in the forest, also with no significant differences between environments (paired t test, n = 3, p = 0.51). We detected no intraspecific seedling clustering for either species; densities in seedling centered microplots (excluding the central seedling) were similar to those in the paired randomly placed microplots (paired t test: Cybianthus: border (p = 0.35), forest (p = 0.09); Clusia: border (p = 0.94), forest (p = 0.44)).
Relative growth rates of Cybianthus seedlings were higher in the border than in the forest, with values of 0.4 and 0.2 cm cm−1 yr−1 respectively (t = 2.43; p < 0.05). Clusia seedlings grew at similar rates with values of 0.18 and 0.22 cm cm−1 yr−1 in border and forest, respectively (t = − 0.72, p = 0.52, Fig. 5).

Average relative growth rates (± SD) of tree seedlings in two environments (border and forest) at a seasonally dry treeline in the Venezuelan Andes a of Cybianthus marginatus (n = 82 seedlings in border and 67 in forest) and b Clusia multiflora (n = 29 seedlings in border and 39 in forest). Values sharing the same letter are not statistically different (p ≥ 0.05) (Kruskal–Wallis test)
Survival rates after one year were very high (Fig. 6). Clusia seedlings showed 0% mortality in the border and only 8% in the forest. Nevertheless, the proportion of healthy seedlings decreased in time and signs of herbivory and the presence of chlorotic leaves and other pigmentation were more common in the last survey (January 2020), mainly in the forest understory. Similar tendencies were found for Cybianthus, for which we recorded mortalities < 5% in both environments.

Survival rates and health status of the seedlings of Cybianthus marginatus (a) and Clusia multiflora (b) at a seasonally dry treeline in the Venezuelan Andes, followed from February 2019, during two resurveys, in August 2019 (Aug19) and January 2020 (Jan20) in páramo-forest border (left half) and forest (right half) environments
Local spatial associations of the tree seedlings
Seedlings of Cybianthus in the border were positively associated with basal-rosette plants and with mosses, while those established in the forest understory were positively associated with ferns and lichens (Fig. 7a). Seedlings of Clusia were positively associated with ferns and mosses in forest environments (Fig. 7b). No significant negative associations were found.

Average relative interaction Index (RII) (± 95% confidence interval) for Cybianthus marginatus (a) and Clusia multiflora (b), based on the changes in cover of the different life forms and ground covers sampled in seedling-centered and paired random microplots located in border (black symbols) and forest (white symbols) environments at a seasonally dry treeline in the Venezuelan Andes. Number of pairs of microplots: Clusia: 21 in border, 24 in forest. Cybianthus: 42 in border, 43 in forest (rosettes were excluded in some situations because they appeared in < 25% of the microplots). The RII is statistically significant (*) (p < 0.05) when the confidence interval does not overlap with the zero (0, 0) line
Seedling ecophysiological responses
Both species showed low assimilation and transpiration rates in all three environments (forest, border-gaps and border under cover). Even though during the physiological measurements on Cybianthus seedlings, PPFD was significantly higher in the gaps in the border (389 µmol m−2 s−1) than in the border under cover or in the forest (ca. 29 µmol m−2 s−1), this did not result in a higher photosynthesis rates or other changes in the measured physiological parameters (Gs, E, WUE). For Clusia, none of the parameters, not even the measured local light levels, differed significantly between environments (Table 1).
Discussion
The studied treeline experienced a marked dry season, but seedling survival in the border and the interior of the forest was still very high. However, the fact that no tree seedlings of our focal species were observed beyond the forest border, neither in our plots nor in the surroundings, indicates that the páramo conditions could be preventing tree seedling establishment. Even in the sheltered conditions in border and forest, local positive interactions, especially with mosses and ferns, seem to play a role in favoring seedling establishment. Nevertheless, there seem to be strong limitations for forest expansion into the páramo at this dry treeline, produced either by filters in dispersal or establishment processes, and these could increase with more extreme dry seasons, as future climate projections predict (Sage 2020).
General climate and microclimate at the plot level
We registered higher annual precipitation values than those reported previously at a lower elevation close to the study area (> 1000 mm at our site vs. 546 mm per year at 2400 m. Aranguren et al. 2012, Appendix 4 in ESM). Even though we have only one year of data, this difference is so large that we can conclude that our site is wetter than that weather-station location, where the wettest year on record (records available for 1970–1990 with some gaps) still had only 712 mm. However, during the dry season (January and February) the total amount of precipitation at our site was only 15 mm in two months. This is an extremely low value for rainfall in the tropical alpine zone in the northern Andes, which is generally described as “wet and cold” (Sarmiento 1986; Buytaert et al. 2011). As this section of the Venezuelan Andes is part of a system of inter-Andean valleys that lie in the rain shadow of the Sierra Nevada mountain range, it is characterized by complex patterns in precipitation seasonality and relatively low amounts of annual precipitation (Buytaert et al. 2011; Suarez del Moral and Chacón-Moreno 2011). The same phenomenon is common in other inter-Andean regions in the northern tropical Andes (Sarmiento 1986; Rada et al. 2019). Even though our data represent just one year of measurements, they indicate that San Jose páramo can be characterized as a dry treeline, very different from, for example, La Aguada, the location for most studies about treeline vegetation in the Venezuelan Andes in the past three decades (see Llambí 2015 for a review) and from other humid treelines studied along the northern Andes (Wille et al. 2002; Bader et al. 2007b; Hofstede et al. 2014).
Globally, the alpine treeline appears at locations with mean air temperatures for the growing season between 5.5 and 7.5 °C (Körner and Paulsen 2004). Climate records at tropical treelines are very rare and most reported temperatures are based on soil temperatures, as a proxy for air temperatures (Körner and Paulsen 2004). Temperature data, with time spans between weeks and years, are available from some cloud-forest treelines, with relatively high temperatures (Bader et al. 2007b; Körner and Paulsen 2004; Ramírez et al. 2009; Rehm and Feeley 2013) or Polylepis patches, with relatively low temperatures (Körner and Paulsen 2004; Kessler et al. 2014). Average air temperature (recorded at 1.50 m from the soil) at our location is 9.4 °C, which fits in the range reported previously for tropical cloud-forest treelines (Bader et al. 2007b) but which is relatively warm compared to global values (Körner and Paulsen 2004). Interestingly, although globally treelines tend to occur at higher elevations in drier mountains, in the tropical Andes the reverse pattern seems to be common (Agudelo 2016). Whether this is due to direct climatic constraints or land-use patterns (an avoidance of the inhospitable wet páramo areas) remains to be determined.
At 50 cm above the ground, the average air temperature was very similar in páramo, border and forest. However, daily temperature fluctuations were larger in gaps in the forest border and in páramo than within forest and covered sites in the border. These differences could be linked to differences in radiative energy exchange. In the forest understory, the canopy reduces solar energy inputs during the day and radiative heat loss during the night, resulting in a dampening of temperature fluctuations compared to the more exposed páramo. Similar differences between forest and páramo have been found in other studies in the Northern Andes (Bader et al. 2007b; Cáceres and Rada 2011; Rada et al. 2011) although climatic characterizations of the forest border are scarce (but see Bueno and Llambí 2015 and Rehm and Feeley 2013). Our data indicate that the microclimate in the border is spatially very variable and may locally be more extreme than that of the páramo. In gaps the microclimate is similar to the páramo, but temperature extremes may be accentuated. These more extreme conditions, particularly relevant in the occurrence of frost, are probably due to reduced wind speeds in the gaps compared to the páramo. The resulting thicker boundary layer could lead to a stronger role of radiative heat exchange relative to convective heat exchange. In other words, in the gaps the cooling winds during the day and warming winds during the night are probably reduced, leading to stronger heating and cooling in these situations. However, seedlings were more abundant in these gaps than in the open páramo, suggesting that temperature could interact with other factors such as direct radiation to influence seedling establishment. Other possible mechanisms that may play a role are strong competition of the seedlings with the dense grass cover in the páramo (particularly by tall tussocks of Calamagrostis), and lower seed rain outside the border. However, seeds do reach the páramo and can overcome competition at least temporarily, as indicated by earlier observations on Cybianthus seedlings in the páramo at our study site (Rodriguez et al. 2011).
Seedling density, growth and survival
The complete absence of tree seedlings of our focal species in the páramo, in spite of earlier observations of Cybianthus seedlings in the same area (Rodríguez et al. 2011) suggests a high seedling mortality as the main cause of their current absence. At other treelines in the tropical Andes, tree seedlings were also observed to decline strongly with distance from the forest (Bader et al. 2007a; Ramos 2008; Puentes 2010; Rehm and Feeley 2013) although in some cases a few seedlings of some radiation-tolerant species were found outside the forest (Bader et al. 2007a; Puentes 2010). Low temperature preventing tree growth has been pointed out as the main reason for the existence of alpine treelines (Tranquillini 1979; Wardle 1993; Körner 1998). However, other factors such as biomass loss (dieback) and seedling mortality (including establishment failure) are additional mechanisms that can influence the position and spatial pattern (or “form”) of the ecotone (Harsch and Bader 2011; Bader et al. 2021). In sites where seedling mortality prevails as the main mechanism driving treeline structure and dynamics, an abrupt treeline form is expected, i.e. a sharp decline in tree height, as trees would not be strongly limited in growth once successfully established (Bader et al. 2021). In our study site, the treeline form could be characterized as an abrupt tree-island treeline, which would agree with the idea that low establishment probabilities are an important mechanism maintaining the position of this treeline. The island formation and the discrete boundary between forest and páramo also suggest that a positive-feedback switch (sensu Wilson and Agnew 1992) likely occurs.
The fact that adults of both species can grow as successful individuals as part of small islands above the upper continuous forest line in the study area (personal observation) indicates that, even in small proportions, some seedlings overcome the filters at this stage. Cybianthus has been reported as the dominant species in forest islands at more humid treeline sites in Venezuela (Ramírez et al. 2009). However, Clusia was absent from these islands on humid slopes. Since in the forest islands of our study area, both of our target species are mixed with the tree Diplostephium venezuelense and other páramo shrubs (Hypericum stenopetalum, Chaetolepis lindeniana), their successful establishment above continuous forests may be driven by a combination of several mild dry seasons and facilitative processes mediated by shrubs (Llambí et al. 2013; Bueno and Llambí 2015). Local topographic features such as large boulders or small depressions generating water accumulation also offer a feasible explanation for the initiation of these islands (Ramírez et al. 2009; Llambí et al. 2013). The species composition of these islands, described previously for the humid slopes, is more similar to border than to continuous forest (Ramírez et al. 2009). Although it is uncertain whether these islands are remnants of formerly more extensive forests or whether they established in a páramo matrix, the border-like composition and absence of typical closed-forest trees seems to suggest the latter.
The survival rates of the seedlings of both target species were high, but the growth rates were low, suggesting the presence of a seedling bank in the understory of the forest and in the border. Higher relative growth rates of Cybianthus in the border compared with the understory of the forest suggest that the border may be a more favorable environment for these seedlings than the forest interior (where light could become limiting). This could explain the tendency for the better health status of the seedlings established in the border compared with those in the forest. For Clusia, growth did not differ among environments, but for this species there was a higher proportion of chlorotic leaves and/or herbivory in the forest than in the border, again suggesting the border is a more favorable environment. Supporting this conclusion, Bader et al. (2007a) found in a 2.5-year experiment with transplanted seedlings in treeline forests and páramos in Ecuador, that the lowest growth rates (with mostly negative rates) occurred in Clusia flaviflora and other tree seedlings inside the forest, while higher growth rates were found in the páramo. However, in that study Clusia showed high mortality rates in the páramo when no shade was provided.
Local spatial associations of the tree seedlings
Seedlings were not randomly distributed within the forest or border environments. The most consistent pattern was a positive local spatial association of seedlings of both focal species with mosses. The role of bryophytes as “nurse species” in tree seedling establishment has also been observed at boreal alpine treelines (Lett et al. 2017, 2020). In humid tropical alpine treelines, mosses have been observed to overgrow seedlings in wet treeline forest (Bader et al. 2007a) and to have negative spatial association with seedlings of D. venezuelensis from the treeline (Llambí et al. 2013). However, Ramos (2008) found positive association of C. multiflora with mosses at a treeline in Colombia. Since mosses tend to prefer moist conditions and also can locally increase moisture by holding high amounts of water, the positive association seems reasonable in this seasonally dry treeline. However, Cybianthus seedlings in the forest were also positively associated with lichens. As lichens tend to indicate dryer and brighter conditions than mosses, this association might indicate a preference for relatively high-light microsites inside the forest. Besides mosses and lichens, ferns also seemed to be associated with favorable microsites for seedlings of our target species, at least inside the forest. Similarly, Ramos (2008) found that ferns enhanced seedling survival at a Colombian treeline, and Llambí et al. (2013) reported positive spatial association of tree seedlings with a fern from the Lycopodiaceae family in open páramo environments at a wet treeline in the Venezuelan Andes.
Rosette plants have been described previously as nurse plants for herbs, cushions and small shrubs in high tropical alpine environments (Mora et al. 2018). In our study, the basal rosette Ruilopezia artropurpurea appears to provide shelter to seedlings of Cybianthus in border environments. Interestingly, tussock grasses do not seem to act as nurse plants for seedlings of tree species in the alpine tropics, as has been described for some temperate-zone treelines (Germino et al. 2002). The fact that the Cybianthus seedlings are positively associated with mosses in the border, and that this life form is replaced as the dominant life form in the lower stratum by tussock grasses in the páramo, suggests that there could be strong negative effects of tussock grasses that inhibited the establishment of new tree seedlings, either by direct competition with the seedlings or by competition with mosses that could act as facilitators for the seedlings.
Seedling ecophysiological responses
All the seedlings showed a similar physiological status, independent of the environment where they were established. We found low assimilation rates for both target species compared to what has been previously reported for adults of these species on humid slopes in the Venezuelan Andes (Rada et al. 2009). As the measured radiation intensities of 300 µmol m−2 s−1 in the border correspond to only a fraction of the 1300 µmol m−2 s−1 sometimes measured at the site (in the páramo), it is possible that all seedlings were operating below their light saturation point, with seedlings in the forest and under cover being more acclimatized to shade and seedlings in the gaps to higher light conditions. Apart from a different light acclimation, the relatively low assimilation rates in the gaps could have been due to stomatal closure to prevent water loss, but this is not supported by the data, as Gs, E, Ci/Ca and WUE did not differ significantly between environments. Another explanation would be photoinhibition due to excess radiation in the gaps, but this is also not supported by the tendencies found in the relevant parameters. Nevertheless, Cybianthus seedlings, especially those individuals in the gaps, developed pigmented leaves (red and orange) between the first and the last survey, suggesting the production of protective pigmentation to prevent damage to the photosystem. Contrary to our findings, Olivares (1997) described that C. multiflora is present only in gaps and borders of the cloud-forest, but never inside the forest canopy, therefore no shade tolerance is excepted in this species (Olivares 1997). However, our results indicate that seedlings of this species perform well under the canopy of the forest, even if their health status seemed slightly better in the border.
Conclusion
We showed that: (i) there is a strong dry season in this tropical treeline; (ii) there is a drastic decrease in the density of tree seedlings, including our two-target species, from the forest to the páramo; (iii) the established seedlings in the forest and forest border survive well but grow very slowly, which indicates the presence of a persistent seedling bank in these shady environments; (iv) local spatial associations suggest facilitative neighbor effects, which could be linked with increased moisture; and (v) photosynthetic rates were similarly low in all the environments where seedlings were found.
The lack of seedling establishment in the páramo indicates that any upward shift of the forest will be very slow and will probably occur in a closed front around the existing forest (Llambí et al. 2013). This suggests that both forest recovery after past land use and forest expansion due to climate change could be a very slow processes (see González et al. 2011; Llambí et al. 2013; Bueno and Llambí 2015). Like in other parts of the Andes, the exact status of the treeline ecotone and páramo, whether natural or influenced by land use and fire, is not completely clear here. Currently, land use is very extensive and appears to have little effect on the ecotone. However, the relatively dry conditions of the area might have favored a more intensive land-use in the past, which may also help to explain the low ecotone elevation. If this was the case, it happened before known historical records, so the legacy is long-lasting (as reported in other semi-arid enclaves where wheat cultivation resulted in frozen succession, see Sarmiento et al. 2015).
One important question that emerged from our results is why we did not find established seedlings of our target species in the páramo, even when the microenvironment is very similar between the páramo and the gaps in the border (in terms of temperature and humidity). The answer may lie in other abiotic factors (e.g. radiation or soil moisture), seed availability, or biotic effects (e.g. local competition with the dense tussock grass cover) that could all restrict establishment in open páramo environments. The fact that the only seedlings from woody species found in open páramo areas belong the radiation-tolerant D. venezuelense (Puentes 2010) may indicate that radiation is a main factor preventing the establishment of the other tree species. This would suggest a scenario of high potential stability for this treelines, with expansion depending on the formation of a relatively closed canopy at the edge of the forest. However, longer-term monitoring of vegetation (permanent plots) and climate (to detect possible sporadic frost events), as well as experimental manipulation (e.g. removals or transplants) are required to provide a more mechanistic understanding of the observed dynamics.
Availability of data and materials
The datasets generated during and/or analyzed during the current study will be available in Dryad repository.
References
Abele A (2000) Estudio florístico de un relicto de bosque montano alto ubicado al sur de la Sierra Nevada de Mérida Venezuela. Dissertation, Universidad de Los Andes
Agudelo C (2016) Modelo espacialmente explícito de la zona de transición bosque-páramo en Colombia. Dissertation, Universidad Distrital Francisco José de Caldas
Akhalkatsi M, Abdaladze O, Nakhutsrishvili G, Smith WK (2006) Facilitation of seedling microsites by Rhododendron caucasicum extends the Betula litwinowii alpine treeline, Caucasus Mountains, Republic of Georgia. Arctic Antarct Alp Res 38:481–488. https://doi.org/10.1657/1523-0430(2006)38[481:FOSMBR]2.0.CO;2
Aranguren A, Andressen R, Henao A (2012) El clima estacional del cinturón montano en el estado Mérida-Venezuela. Rev Geogr Venez 53:187–212
Armas C, Ordiales R, Pugnaire FI (2004) Measuring plant interactions: a new comparative index. Ecology 85:2682–2686. https://doi.org/10.1890/03-0650
Arzac A, Llambí LD, Dulhoste R, Olano JM, Chacón-Moreno E (2019) Modelling the effect of temperature changes on plant life-form distribution across a treeline ecotone in the tropical Andes. Plant Ecol Divers. https://doi.org/10.1080/17550874.2019.1655108
Ataroff M, Sarmiento L (2004) Las unidades ecológicas de los Andes Venezolanos. In: La Marca E, Soriano P (eds) Reptiles de los Andes de Venezuela. Catálogo Zoológico de Venezuela vol 2. Mérida, pp 9–26
Azócar A, Rada F (2006) Ecofisiología de plantas de páramo. ICAE, Mérida, p 182
Bader MY, Van Geloof I, Rietkerk M (2007a) High solar radiation hinders tree regeneration above the alpine treeline in northern Ecuador. Plant Ecol 191:33–45. https://doi.org/10.1007/s11258-006-9212-6
Bader MY, Rietkerk M, Bregt AK (2007b) Vegetation structure and temperature regimes of tropical alpine treelines. Arctic Antarct Alp Res 39(3):353–364. https://doi.org/10.1657/1523-0430(06-055)[BADER]2.0.CO;2
Bader MY, Llambí LD, Case BS, Buckley HL, Toivonen JM, Camarero JJ, Cairns DM, Brown CD, Wiegand T, Resler LM (2021) A global framework for linking alpine-treeline ecotone patterns to underlying processes. Ecography 44(2):265–292. https://doi.org/10.1111/ecog.05285
Bansal S, Germino MJ (2010) Unique responses of respiration, growth, and non-structural carbohydrate storage in sink tissue of conifer seedlings to an elevation gradient at timberline. Environ Exp Bot 69:313–319. https://doi.org/10.1016/j.envexpbot.2010.05.002
Batllori E, Camarero JJ, Gutiérrez E (2010) Current regeneration patterns at the tree line in the Pyrenees indicate similar recruitment processes irrespective of the past disturbance regime. J Biogeogr 37:1938–1950. https://doi.org/10.1111/j.1365-2699.2010.02348.x
Batllori E, Camarero JJ, Ninot JM, Gutiérrez E (2009) Seedling recruitment, survival and facilitation in alpine Pinus uncinata tree line ecotones. Implications and potential responses to climate warming. Glob Ecol Biogeogr 18:460–472. https://doi.org/10.1111/j.1466-8238.2009.00464.x
Bendix J, Fries A, Zárate J, Trachte K, Rollenbeck R, Pucha-Cofrfrep F, Paladines R, Palacios I, Orellana J, Oñate-Valdivieso F, Naranjo C, Mendoza L, Mejia D, Guallpa M, Gordillo F, Gonzalez-Jaramillo V, Dobbermann M, Célleri R, Carrillo C, Araque A, Achilles S (2017) RadarNet-Sur first weather radar network in tropical high mountains. Bull Am Meteorol Soc 98:1235–1254. https://doi.org/10.1175/BAMS-D-15-00178.1
Braun C, Bezada M (2013) The history and disappearance of glaciers in Venezuela. J Lat Am Geogr 12:85–124. https://doi.org/10.1353/lag.2013.0016
Brodersen CR, Germino MJ, Johnson DM, Reinhardt K, Smith WK, Resler LM, Bader MY, Sala A, Kueppers LM, Broll G, Cairns DM, Holtmeier F-K, Wieser G (2019) Seedling survival at timberline is critical to conifer mountain forest elevation and extent. Front Glob Chang 2:9. https://doi.org/10.3389/ffgc.2019.00009
Bueno A, Llambí LD (2015) Facilitation and edge effects influence vegetation regeneration in old-fields at the tropical Andean forest line. Appl Veg Sci 18:613–623.https://doi.org/10.1111/avsc.12186
Buytaert W, Cuesta-Camacho F, Tobón C (2011) Potential impacts of climate change on the environmental services of humid tropical alpine regions. Glob Ecol Biogeogr 20:19–33. https://doi.org/10.1111/j.1466-8238.2010.00585.x
Cáceres Y, Rada F (2011) ¿Cómo responde la especie leñosa Vaccinum meridionale a la temperatura en su límite altitudinal de distribución en los andes tropicales? Ecotrópicos 24:80–91
Cavieres LA, Rada F, Azócar A, García-Núñez C, Cabrera HM (2000) Gas exchange and low temperature resistance in two tropical high mountain tree species from the Venezuelan Andes. Acta Oecologica 21(3):203–211. https://doi.org/10.1016/S1146-609X(00)01077-8
Chacón-Moreno E, Rodríguez-Morales M, Paredes D (2021) Impacts of global change on the spatial dynamics of treeline in Venezuelan Andes. Front Ecol Evol 9:74. https://doi.org/10.3389/fevo.2021.615223
Conlisk E, Castanha C, Germino MJ, Veblen TT, Smith JM, Moyes AB, Kueppers LM (2018) Seed origin and warming constrain lodgepole pine recruitment, slowing the pace of population range shifts. Glob Chang Biol 24:197–211. https://doi.org/10.1111/gcb.13840
Correa-Gómez DF, Stevenson PR, Álvarez E, Aldana AM, Umaña M, Cano Á et al (2013) Patrones de frecuencia y abundancia de sistemas de dispersión de plantas en bosques colombianos y su relación con las regiones geográficas del país. Colombia Forestal 16(1):33–51
Costa Talens M, Cegarra JA, Lugo Salinas L, Lozada JR, Guevara JR, Soriano P (2007) The bioclimatic belts of the Venezuelan Andes in the State of Mérida. Phytocoenologia 37:711–738. https://doi.org/10.1127/0340-269X/2007/0037-0711
Dulhoste R (2010) Respuestas ecofisiológicas de plantas del límite arbóreo (selva nublada-páramo) al estrés térmico, hídrico y lumínico en los Andes Venezolanos. Dissertation, Universidad de los Andes
Ely F, Rada F, Gutiérrez N (2011) Análisis morfofuncional de tres bambúes leñosos del ecotono selva nublada-páramo en los Andes Venezolanos. Ecotopricos 24:92–112
García-Nuñez C, Rada F, Boero C, Gallardo M, Hilal M, Prado F (2004) Leaf gas exchange and water relations in Polylepis tarapacana at extreme altitudes in the Bolivian Andes. Photosynthetica 42:133–138. https://doi.org/10.1023/B:PHOT.0000040581.94641.ed
Germino MJ, Smith WK (1999) Sky exposure. crown architecture, and low-temperature photoinhibition in conifer seedlings at alpine treeline. Plant Cell Environ 22:407–415. https://doi.org/10.1046/j.1365-3040.1999.00426.x
Germino MJ, Smith WK, Resor AC (2002) Conifer seedling distribution and survival in an alpine-treeline ecotone. Plant Ecol 162(2):157–168
González WY, Llambí LD, Smith JK, Gámez LE (2011) Dinámica sucesional del componente arbóreo en la zona de transición bosque-Páramo en Los Andes Tropicales. Ecotrópicos 24(1):60–79
Graae BJ, Ejrnæs R, Lang SI, Meineri E, Ibarra PT, Bruun HH (2011) Strong microsite control of seedling recruitment in tundra. Oecologia 166:565–576. https://doi.org/10.1007/s00442-010-1878-8
Harper JL (1977) Population biology of plants. Academic Press, London
Harsch MA, Bader MY (2011) Treeline form—a potential key to understanding treeline dynamics. Glob Ecol Biogeogr 20(4):582–596
Hofstede RGM, Dickinson KJM, Mark AF, Narváez E (2014) A broad transition from cloud forest to páramo characterizes an undisturbed treeline in Parque Nacional Llanganates. Ecuador Arctic Antarct Alp Res 46:975–986. https://doi.org/10.1657/1938-4246-46.4.975
Hofstede RGM (2003) Los páramos en el mundo: su diversidad y sus habitantes. In: Hofstede R, Segarra P, Mena Vasconez P (eds) Los páramos del mundo. Proyecto Atlas Mundial de los Páramos. UICN/EcoCiencia, Quito, pp 15–38
Hokche O, Berry P, Huber YO (2008) Nuevo catálogo de la flora vascular de Venezuela. Fundación Instituto Botánico de Venezuela “Dr. Tobías Lasser”. Caracas
Holtmeier FK, Broll G (2020) Treeline research-from the roots of the past to present time. a review. Forests. https://doi.org/10.3390/f11010038
Johnson AC, Yeakley JA (2016) Seedling regeneration in the alpine treeline ecotone: comparison of wood microsites and adjacent soil substrates. Mt Res Dev 36:443–451. https://doi.org/10.1659/mrd-journal-d-16-00024r.1
Kessler M, Toivonen JM, Sylvester SP, Kluge J, Hertel D (2014) Elevational patterns of Polylepis tree height (Rosaceae) in the high Andes of Peru: role of human impact and climatic conditions. Front Plant Sci. https://doi.org/10.3389/fpls.2014.00194
Körner C (1998) A re-assessment of high elevation treeline positions and their explanation. Oecologia 115(4):445–459. https://doi.org/10.1007/s004420050540
Körner C (2021) The cold range limit of trees. Trends Ecol Evol 36(11):979–989. https://doi.org/10.1016/j.tree.2021.06.011
Körner C, Paulsen J (2004) A world-wide study of high altitude treeline temperatures. J Biogeogr 31(5):713–732. https://doi.org/10.1111/j.1365-2699.2003.01043.x
Körner C (2003) The alpine life zone. In: Alpine plant life. Springer, Berlin. https://doi.org/10.1007/978-3-642-18970-8_2
Lambers H, Chapin FS, Pons TL (2008) Plant physiological ecology. Springer, New York, p 604p
León O, Jiménez D, Marín C (2015) Marco conceptual para la identificación de la zona de transición entre el bosque altoandino y páramo. In: Sarmiento C, León O (eds) Transición bosque– páramo. Bases conceptuales y métodos para su identificación en los Andes colombianos. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá
Lett S, Dorrepaal E (2018) Global drivers of tree seedling establishment at alpine treelines in a changing climate. Funct Ecol 32:1666–1680. https://doi.org/10.1111/1365-2435.13137
Lett S, Nilsson MC, Wardle DA, Dorrepaal E (2017) Bryophyte traits explain climate-warming effects on tree seedling establishment. J Ecol 105:496–506. https://doi.org/10.1111/1365-2745.12688
Lett S, Krab E, Teuber L (2020) Mosses modify effects of warmer and wetter conditions on tree seedlings at the alpine treeline. Glo Chag Biol 26(10):5754–5766. https://doi.org/10.1111/gcb.15256
Llambí LD, Fontaine M, Rada F, Saugier B, Sarmiento L (2003) Ecophysiology of dominant plant species during old-field succession in a High Tropical Andean ecosystem. Arctic Antarct Alp Res 35:447–453. https://doi.org/10.1657/1523-0430(2003)035[0447:EODPSD]2.0.CO;2
Llambí LD, Puentes J, García-Núñez C (2013) Spatial relations and population structure of a dominant tree along a treeline ecotone in the Tropical Andes: interactions at gradient and plant-neighbourhood scales. Plant Ecol Divers 6:343–353. https://doi.org/10.1080/17550874.2013.810312
Llambí LD, Ramírez L, Schwarzkopf T (2014) Patrones de distribución de plantas leñosas en el ecotono bosque- páramo de la Sierra Nevada de Mérida: ¿Qué nos sugieren sobre la dinámica del límite del bosque? In: Cuesta-Camacho F, Sevink J, Llambí LD, De Bièvre B, Posner J (eds) Avances en investigación para la conservación de los páramos andinos. CONDESAN, Quito, pp 488–502
Llambí LD (2015) Estructura, diversidad y dinámica de la vegetación en el ecotono bosque-páramo: Revisión de la evidencia en la cordillera de Mérida. Acta Biol Colomb 20:5–19. https://doi.org/10.15446/abc.v20n3.46721
Loranger H, Zotz G, Bader MY (2016) Early establishment of trees at the alpine treeline: Idiosyncratic species responses to temperature-moisture interactions. AoB Plants. https://doi.org/10.1093/aobpla/plw053
Loranger H, Zotz G, Bader MY (2017) Competitor or facilitator? The ambiguous role of alpine grassland for the early establishment of tree seedlings at treeline. Oikos 126:1625–1636. https://doi.org/10.1111/oik.04377
Maher EL, Germino MJ (2006) Microsite differentiation among conifer species during seedling establishment at alpine treeline1. Ecoscience 13:334–341. https://doi.org/10.2980/i1195-6860-13-3-334.1
Maher EL, Germino MJ, Hasselquist NJ (2005) Interactive effects of tree and herb cover on survivorship, physiology, and microclimate of conifer seedlings at the alpine tree-line ecotone. Can J for Res 35:567–574. https://doi.org/10.1139/x04-201
Malanson GP, Resler LM, Butler DR, Fagre DB (2019) Mountain plant communities: uncertain sentinels? Prog Phys Geogr 43(4):521–543. https://doi.org/10.1177/0309133319843873
Moles AT, Westoby M (2004) What do seedlings die from and what are the implications for evolution of seed size? Oikos 106(1):193–199
Monasterio M (1980) Los Páramos andinos como región natural. Características biogeográficas generales y afinidad con otras regiones andinas. In: Monasterio M (ed) Estudios Ecológicos en los Páramos andinos. Editorial Universidad de los Andes, Mérida, pp 15–27
Mora MA, Llambí LD, Ramírez L (2018) Giant stem rosettes have strong facilitation effects on alpine plant communities in the tropical Andes. Plant Ecol Divers 12:593–606. https://doi.org/10.1080/17550874.2018.1507055
Moyes AB, Castanha C, Germino MJ, Kueppers LM (2013) Warming and the dependence of limber pine (Pinus flexilis) establishment on summer soil moisture within and above its current elevation range. Oecologia 171:271–282. https://doi.org/10.1007/s00442-012-2410-0
Moyes AB, Germino MJ, Kueppers LM (2015) Moisture rivals temperature in limiting photosynthesis by trees establishing beyond their cold-edge range limit under ambient and warmed conditions. New Phytol 207:1005–1014. https://doi.org/10.1111/nph.13422
Ninot JM, Batllori E, Carrillo E, Carreras J, Ferré A, Gutiérrez E (2008) Timberline structure and limited tree recruitment in the Catalan Pyrenees. Plant Ecol Divers 1:47–57. https://doi.org/10.1080/17550870802260764
Manrique NT, Morales ME (2016) Frutos y semillas en remanentes de bosque altoandino del páramo de Rabanal (Boyacá, Colombia). Bistua Rev de la Fac de Cienc Básic 14(2):141–168
Olivares E (1997) Prolonged leaf senescence in Clusia multiflora. Trees 11(6):370–377
Ortis CL, Umba AC (2010) Dinámica anual de un ensamble de aves frugívoras y su relación con la dispersión de semillas. In: Estudios ecológicos en el Parque Natural Municipal Ranchería, un aporte para su conservación. Universidad Pedagógica y Tecnológica de Colombia
Piper FI, Cavieres LA, Reyes-Díaz M, Corcuera LJ (2006) Carbon sink limitation and frost tolerance control performance of the tree Kageneckia angustifolia D. Don (Rosaceae) at the treeline in central Chile. Plant Ecol 185:29–39. https://doi.org/10.1007/s11258-005-9081-4
Popp M, Kramer D, Lee H, Diaz M, Ziegler H, Lüttge U (1987) Crassulacean acid metabolism in tropical dicotyledonous trees of the genus Clusia. Trees 1:238–247. https://doi.org/10.1007/BF01816822
Puentes J (2010) Patrones y mecanismos de establecimiento de dos especies leñosas en la transicion entre el bosque paramero y el páramo en los Andes Tropicales. Dissertation, Universidad de los Andes
Rada F, García-Núñez C, Ataroff M (2009) Leaf gas exchange in canopy species of a Venezuelan cloud forest. Biotropica 41(6):659–664. https://doi.org/10.1111/j.1744-7429.2009.00537.x
Rada F, García-Núñez C, Rangel S (2011) Microclimate and regeneration patterns of Polylepis sericea in a treeline forest of the Venezuelan Andes. Ecotrópicos 24(1):113–122
Rada F, Azócar A, García-Núñez C (2019) Plant functional diversity in tropical Andean páramos. Plant Ecol Divers 00:1–15. https://doi.org/10.1080/17550874.2019.1674396
Ramírez L, Llambí LD, Schwarzkopf T, Gamez LE, Marquez N (2009) Vegetation structure along the forest – páramo transition belt in the Sierra Nevada de Mérida: implications for understanding treeline dynamics. Ecotrópicos 22:83–98
Ramos C, García MR (2008) Dinámica del daño foliar en plántulas de Drimys granadensis (Winteraceae) y Clusia multiflora (Clusiaceae) en el bosque altoandino de la Cordillera Oriental colombiana. Rev Biol Trop 56(3):1087–1100
Ramos C (2008) Estrategias regenerativas de Clusia multiflora, Drimys granadensis y Weinmannia tomentosa en el bosque altoandino. Dissertation, Universidad Nacional de Colombia
Rehm EM, Feeley KJ (2013) Forest patches and the upward migration of timberline in the southern Peruvian Andes. For Ecol Manage 305:204–211. https://doi.org/10.1016/j.foreco.2013.05.041
Rehm EM, Feeley KJ (2015) Freezing temperatures as a limit to forest recruitment above tropical Andean treelines. Ecology 96:1856–1865. https://doi.org/10.1890/14-1992.1
Resler LM, Butler DR, Malanson GP (2005) Topographic shelter and conifer establishment and mortality in an alpine environment. Glacier National Park Montana Phys Geogr 26:112–125. https://doi.org/10.2747/0272-3646.26.2.112
Rodríguez E, Schwarzkopf T, Gamez LE, Dugarte W, Dulhoste R (2011) Canopy structure and woody species distribution at the upper treeline in the Venezuelan Andes. Ecotrópicos 24(1):47–59
Rundel PW (1994) Tropical alpine climates. In: Rundel PW, Smith AP, Meinzer FC (eds) Tropical alpine environments: plant form and function. Cambridge University Press, Cambridge, pp 21–44
Sage RF (2020) Global change biology: a primer. Glob Chang Biol 26:3–30. https://doi.org/10.1111/gcb.14893
Sarmiento G (1986) Ecological features of climate in high tropical mountains. In: Vuilleumier F, Monasterio M (eds) High altitude tropical biogeography. Oxford University Press, Oxford, pp 11–45
Sarmiento L (2000) Water balance and soil loss under long fallow agriculture in the Venezuelan Andes. Mt Res Dev 20:246–253
Sarmiento L, Smith JK, Márquez N, Escalona A, Erazo MC (2015) Constraints for the restoration of tropical alpine vegetation on degraded slopes of the Venezuelan Andes. Plant Ecol Divers 8(2):277–291
Sidgel S, Eryuan S, Camarero JJ, Wang Y, Dawadi B (2020) Tree-to-tree interactions slow down, Himalayan treeline shifts as inferred from tree spatial patterns. J Biogeog 47(8):1816–1826. https://doi.org/10.1111/jbi.13840
Sodja I (2001) Estudio florístico del cerro Pozo Negro, páramo de San José (Mérida-Venezuela). Dissertation, Universidad de Los Andes
Suárez del Moral P, Chacón-Moreno E (2011) Modelo espacial de distribución del ecotono bosque-páramo en y escenarios de cambio climático. Ecotrópicos 24:3–25
Tirado C (1997) Estudio florístico de una comunidad Andina escandente. Dissertation, Universidad de Los Andes
Tranquillini W (1979) Physiological ecology of the alpine timberline. Springer, Berlin
Urrutia R, Vuille M (2009) Climate change projections for the tropical Andes using a regional climate model: temperature and precipitation simulations for the end of the 21st century. J Geophys Res Atmos 114:1–15. https://doi.org/10.1029/2008JD011021
Wardle P (1993) Causes of alpine timberline: a review of the hypotheses. In: Alden JN, Mastrantonio JL, Ødum S (eds) Forest development in cold climates. Springer, Boston, pp 89–103. https://doi.org/10.1007/978-1-4899-1600-6_7
Wille M, Hooghiemstra H, Hofstede R, Fehse J, Sevink J (2002) Upper forest line reconstruction in a deforested area in northern Ecuador based on pollen and vegetation analysis. J Trop Ecol 18:409–440
Wilson JB, Agnew AD (1992) Positive-feedback switches in plant communities. Adv Ecol Res 23:263–336
Walter DH, Lieth H(1960) Klimatogramm. Welt Atlas. Fisher, Jena. Germany
Yanez P (1998) Caracterización florística en un sector de cambio páramo- selva nublada en el parque nacional Sierra Nevada. Venezuela Revista Forestal Venezolana 42(1):51–62
Acknowledgements
The authors thank the ICAE and its researchers for the help and assistance during field work and to sr. Martin Sosa for being our host in the páramo de San José. LA Ramírez wants to thank the working group Ecological Plant Geography (University of Marburg) for all the support and the discussion of ideas during the analysis and writing process, to Luis E. Gámez for the taxonomic identification and to Dr. Eulogio Chacón-Moreno for the elaboration of the map in Fig. 1. The authors also thank the anonymous reviewers for their constructive comments. This work was supported by a PhD scholarship from the German Academic Exchange Service (DAAD) to LA Ramírez (Grant No. 57381412).
Funding
Open Access funding enabled and organized by Projekt DEAL. This work was supported by PhD scholarship from the German Academic Exchange Service (DAAD) to L.A Ramírez (Grant No. 57381412).
Author information
Authors and Affiliations
Contributions
LAR and MYB designed the study; LAR, LDLL, CJA, MF and JET collected the data; LAR performed the analyses with support from MYB and LDLL; LAR led the writing of the manuscript. All the authors discussed results, revised manuscript versions and approved of publication of the final version.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Karen Harper.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ramírez, L.A., Llambí, L.D., Azocar, C.J. et al. Patterns in climate and seedling establishment at a dry tropical treeline. Plant Ecol 223, 1047–1068 (2022). https://doi.org/10.1007/s11258-022-01257-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-022-01257-2