
Abstract
Urbanisation is associated with the loss and fragmentation of natural land, the disruption of ecosystem functioning and services, and the loss of biodiversity. Cape Town is situated in a global biodiversity hotspot, with high rates of endemism, but both agricultural and housing demands are increasing pressure on remaining patches of natural land and consequently the biodiversity they support. The aims of this study were to use a standardised camera trap survey to determine which native medium and large mammal species still survive in 12 City of Cape Town (CCT) municipal reserves (range 32–8400 ha), and to understand how reserve size, area-perimeter ratio, connectivity, habitat heterogeneity and presence of permanent freshwater aquatic habitat might influence medium and large mammal (>0.5 kg) community composition. Cameras were placed at 151 locations across all reserves using a stratified placement protocol that resulted in 13,360 independent trigger events by targeted taxa. Nineteen native species (11 carnivores, 7 herbivores, 1 omnivore) were recorded, which is 49% of the 39 species believed to have been present historically. Species richness varied from 1 to 12 species (mean ± SD = 7 ± 3.6) across reserves, and linear models showed that higher species richness and the presence of large carnivores was best explained by improved connectivity to large amounts of natural habitat. It is recommended that maintaining biodiversity in urban reserves will be best achieved by preserving and establishing corridors of suitable habitat that allow for the movement of animals to and from other patches.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Medium and large mammal species richness is an important indicator of ecosystem functioning and health (Augustine and McNaughton 1998; Kerley et al. 2003; Ordeñana et al. 2010). With an increase in anthropogenic activity worldwide, many mammal species are being adversely affected contributing to global declines in biodiversity (Ceballos et al. 2005; Pekin and Pijanowski 2012; Ceballos and Ehrlich 2006; Visconti et al. 2011). In urban areas, natural environments tend to be reduced to small and often isolated pockets which are broadly impacted by a range of anthropogenic activity (Kerley et al. 2003; McDonald et al. 2008; Ceballos et al. 2005; Pekin and Pijanowski 2012; De Stefano and De Graaf 2003; Visconti et al. 2011) including increases in pollution, human interactions and disease exposure (Ceballos and Ehrlich 2006; McDonald et al. 2008; Ordeñana et al. 2010; Visconti et al. 2011). To ensure healthy mammal species richness in protected areas, it is important to understand which factors influence species richness patterns (Ramesh et al. 2016; Gonçalves et al. 2018). This in turn demands effective monitoring of mammals, an understanding of their ecological role and the subsequent prioritization of management and conservation actions that seek to improve or maintain richness and community composition (Ramesh et al. 2016).
At a global scale, climatic variables such as temperature and evapotranspiration are considered the most important correlates of mammal species richness (Torres-Romero and Olalla-Tárraga 2015; Ramesh et al. 2016). Limits of temperature tolerance constrain the geographic range of most mammal species (Ramesh et al. 2016), while evapotranspiration influences primary productivity and bottom-up processes (Torres-Romero and Olalla-Tárraga 2015). At both a regional and a landscape scale, climate and geological features are the most important drivers of species richness patterns (Torres-Romero and Olalla-Tárraga 2015; Ramesh et al. 2016). On smaller scales, anthropogenic activity has influenced species richness patterns at all scales due to habitat loss and fragmentation, pollution, alien species introduction, resource depletion and human-wildlife conflict (Ceballos and Ehrlich 2006; McDonald et al. 2008; Ordeñana et al. 2010; Visconti et al. 2011).
Most large cities are located in areas of high biodiversity and endemism, and consequently urban land use has had a disproportionate impact on mammal species richness (McDonald et al. 2008; Pekin and Pijanowski 2012; Visconti et al. 2011). Despite this many species persist in small urban and peri-urban reserves with certain species thriving in human-modified habitats (De Stefano and De Graaf 2003; Garden et al. 2006; Baker and Harris 2007; Ordeñana et al. 2010; Hoffman and O’Riain 2012a; Šálek et al. 2015). Urbanisation also tends to affect trophic level dynamics within the urban environment and human activity often supplements food sources (Pickett et al. 2011; Saito and Koike 2013). This increases the abundance of certain species and can eliminate others, changing trophic level structure and community composition (De Stefano and De Graaf 2003; Picket et al. 2011).
Understanding which species survive in urban reserves and how they are impacted by varying levels of isolation and neighbouring land use is important to ensure the survival of the remaining species and sound ecosystem functioning where they persist in the few natural spaces remaining (Fischer and Lindenmayer 2007; Ramesh et al. 2016). In general the species richness of small, isolated reserves is affected by reserve size (De Stefano and De Graaf 2003; Kerley et al. 2003; Ceballos et al. 2005; Visconti et al. 2011; Matthies et al. 2017; Gonçalves et al. 2018; Zungu et al. 2019), reserve shape (Diamond 1975; Zungu et al. 2019), habitat heterogeneity within the reserve (Ramesh et al. 2016; Matthies et al. 2017), connectivity to additional suitable habitat (Diamond 1975; Stevens et al. 2006; Correa Ayram et al. 2016) and surrounding land use (McDonald et al. 2008; Visconti et al. 2011; Pekin and Pijanowski 2012; Mann et al. 2015; Torres-Romero and Olalla-Tárraga 2015; Gonçalves et al. 2018).
Garden et al. (2006) reviewed demographic data on urban mammal species, and found that population size at a landscape level was significantly affected by patch size, vegetation and habitat type, and connectivity. Patch size affects the number of individuals and species it can support and the smaller the patch the higher the likelihood of local extinction events as risk of extinction is usually related to population size (Turgeon and Kramer 2012). For this reason, one large patch is often considered preferable to many small patches over the landscape (Heegaard et al. 2007). Patch shape also plays an important role in species presence and therefore richness (Ramesh et al. 2016; Matthies et al. 2017). The shape of an area determines the amount of core habitat available and the amount of exposed edge, with a longer edge in relation to core area allowing for more exposure to anthropogenic effects in an urban context (Herse et al. 2018). Some species respond positively to increased edge and the resulting variation in habitat (Diamond 1975), but many species are sensitive to edge effects and so require a larger core area (Hardt and Forman 1989; Ramesh et al. 2016).
Connectivity refers either to the spatial relationship between patches in a landscape (structural connectivity) or the ability of the landscape to facilitate the movement of species between patches (functional connectivity), and is considered important for species population persistence within fragmented landscapes (Hurst et al. 2013; Correa Ayram et al. 2016). When patch size is small, a species’ persistence may rely solely on its ability to disperse, which depends largely on the type of habitat adjacent to and between patches (Brooker et al. 1999; Söndgerath and Schröder 2002; Turgeon and Kramer 2012). Diamond (1975) and Turgeon and Kramer (2012) suggest that the number of species a patch can hold is a balance between rates of extinction and immigration, such that if animals are able to move into and between areas, local extinction may be reduced. Connectivity is also affected by permeability of patch boundaries (Stevens et al. 2006). Physical barriers (e.g. walls, fences) are important for reducing collisions with vehicles and unregulated access by domestic animals and people but also prevent animal movement between patches (Garden et al. 2006; Ordeñana et al. 2010). There are a number of methods available to quantify both structural and functional connectivity between two potentially habitable areas (Stevens et al. 2006). These methods can be complex and depend largely on the arrangement of patches in relation to the size, shape and quality of corridors within a larger landscape matrix, as well as the size and dispersal behavioural of the species within reserves (Brooker et al. 1999; Tischendorf and Fahrig 2000; Kindlmann and Burel 2008; Crooks et al. 2011; Bateman and Fleming 2012; Correa Ayram et al. 2016).
Mammal community composition should also be considered when trying to understand mammal persistence in urban environments as certain species may be more vulnerable to, or alternatively more adaptable to the effects of urbanization (Bateman and Fleming 2012). This means that naturally-occurring species community composition might be disrupted, influencing how species interact with each other and the environment (Lowry et al. 2013). Large predatory mammals tend to have large home ranges which are invariably reduced and fragmented by urban development (Kerley et al. 2003; Ordeñana et al. 2010). The removal of large predators reduces both competition and predation for smaller predators and prey species, which may then enable mesopredators to persist at higher numbers (Fischer et al. 2012; Bateman and Fleming 2012; Lowry et al. 2013). Species which typically thrive in human transformed areas, such as red fox (Vulpes vulpes), rabbit (Leporidae spp.), deer (Cervidae spp.), chacma baboon (Papio ursinus), vervet monkey (Chlorocebus pygerythrus) and caracal (Caracal caracal) (De Stefano and De Graaf 2003; Hoffman and O’Riain 2012b; Patterson et al. 2018; Leighton et al. 2020), tend to be those which are able to generalize in terms of food sources and habitat preference, and further benefit from the absence of competition and predation (Garden et al. 2006; Ordeñana et al. 2010; Šálek et al. 2015). Certain non-native species (mostly domestic species) also respond positively (Kerley et al. 2003; Ordeñana et al. 2010; Nattrass et al. 2020), indicating that another implication of urbanization is an increase in alien invasive fauna with subsequent effects on local mammal populations (Bernardo and De Melo 2013).
Mammal conservation in large urban areas is extremely challenging and the City of Cape Town (CCT) is no exception (Hoffman and O’Riain 2012c; Holmes et al. 2012; Serieys et al. 2019). Many of the large carnivore and herbivore species such as lion (Panthera leo), black rhinoceros (Diceros bicornis bicornis) and common eland (Tragelephus oryx) were hunted to local extinction within the first 50 years of colonization (Rebelo 1992; Anderson and O’Farrell 2012), but while smaller mammal species have been able to persist in the area, rapid urbanization has become an increasing threat. Flat, fertile soils in proximity to freshwater were rapidly developed firstly for farming and subsequently for housing (Anderson and O’Farrell 2012). Most wildlife populations were thus restricted to marginal habitats including wetlands and mountain habitat where development costs were high (Anderson and O’Farrell 2012). Consequently, only small, fragmented patches were set aside as nature reserves within the lowland areas of Cape Town (Rebelo et al. 2011). Despite this fragmentation and low proportion of protected habitat (approximately 10% excluding the mountainous Table Mountain National Park), the Cape Town area still maintains an extraordinary wealth of biodiversity as highlighted in April 2019 when citizen science aided in the recording of 4157 individual fauna, flora and fungal species to win the iNaturalist City Nature Challenge (iNaturalist Network 2019).
To best conserve the medium and large mammal species still remaining within the CCT boundary and more specifically the reserves of the CCT municipality, a clear understanding of current species richness patterns and the potential drivers thereof is necessary. The aim of this study was to use camera traps to determine which medium and large mammal species assemblages still remain within the respective CCT reserves larger than 30 ha and to identify the potential drivers responsible for the expected differences in mammal community composition. It was hypothesised that species richness would be affected by reserve size, levels of connectivity with other natural land, area-perimeter ratio of the reserve, habitat het\erogeneity and the presence of permanent freshwater aquatic habitat. In accordance with the literature reviewed it was predicted that larger, more connected reserves with a greater area-perimeter ratio and high habitat heterogeneity will individually and collectively have a positive effect on species richness. It was also predicted that the presence of permanent freshwater aquatic habitat in reserves will further elevate richness estimates by attracting water associated mammals such as Cape clawless otter - Aonyx capensis (Okes and O’Riain 2017).
Methods
Study sites
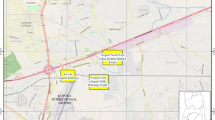
This study was conducted within 12 of the CCT municipal nature reserves. The CCT municipality covers an area of 2461 km2 and surrounds the Table Mountain National Park (Fig. 1). The CCT area forms part of the Cape Floristic Region and more specifically the Fynbos Biome (Rebelo et al. 2006). The area experiences a Mediterranean climate characterised by cool, wet winters and warm, dry and windy summers (Cowling et al. 1996; Rebelo et al. 2006). Mean annual rainfall varies with terrain and latitude from a low of 400 mm in the southern peninsula to 500–600 mm on the lower-lying central areas and 1300–2000 mm on the upper slopes of the northern peninsula inside the Table Mountain National Park (Cowling et al. 1996; Harris et al. 2010). Rebelo et al. (2006) and SANBI (2016) recognise three major vegetation complexes within the boundaries of the CCT, namely fynbos, renosterveld and strandveld, and also classify certain parts as azonal vegetation. Typical fynbos is characterised by shrubland consisting of at least 5% Restionaceae species, with the presence of Ericaceae and Proteaceae shrubs in varying proportions and a low grass component (Rebelo et al. 2006). Fynbos systems are fire-prone and occur mainly in sandy, nutrient-poor soil. Renosterveld structure can vary between shrubland and grassland, consisting of small-leaved, evergreen Asteraceae shrubs, grasses and a large proportion of geophytes. Renosterveld typically excludes Erica and Protea species, and occurs on shale- and granite-derived clay soils. It is also fire prone. Strandveld vegetation occurs along coastal areas but out of direct ocean spray (Rebelo et al. 2006). This vegetation has a medium to dense structure formed by sclerophyllous shrubs, and while Restio species may be present, no Protea and little to no Erica species occur. Strandveld relies on calcium-rich soils and has a low fire frequency (Rebelo et al. 2006). The vegetation types making up these vegetation complexes can be grouped biogeographically, and within the CCT area these are known as Southwest Fynbos, West Coast Renosterveld and West Strandveld (SANBI 2016). The azonal areas are defined as specialised, water-associated vegetation (Rebelo et al. 2006) such as wetland, coastal and riverine vegetation.

City of Cape Town (CCT) nature reserve study sites and undeveloped (i.e. not built up) areas within the CCT municipal area. Table Mountain National Park is also indicated but does not fall under the CCT municipality. “Protected areas” refers to formally protected conservation areas. “Open space” refers to the remainder of undeveloped land, some of which may be managed as conservation areas but which are not protected by any formal legislation, and may include private property. Blank areas indicate urban land use zones within the municipal boundary. Reserve numbers in the legend correspond to the reserve numbers of Table 1 (adapted from CCT 2019a, 2019b)
Approximately 60% of the CCT area has been transformed by urban, agricultural and industrial developments, and only small remnants of natural vegetation persist (Rebelo et al. 2011). At the start of this study, seventeen nature reserves had been proclaimed and are managed by CCT in an effort to conserve some of the remaining natural land and the floral and faunal biodiversity. Thirteen of these reserves are larger than 30 ha and 12 were considered sufficiently secure from a safety and theft perspective to enable a camera trap survey and form the focal areas of this study (Fig. 1).
The 12 reserves vary substantially in size and vegetation cover and for the purpose of this study the vegetation was divided into five broad habitat types. Areas covered by the Southwest Fynbos, West Coast Renosterveld and West Strandveld vegetation are further referred to as fynbos, renosterveld and strandveld respectively. The azonal vegetation associated with the wetland, coastal and riverine vegetation are together classified as ‘water-associated habitat’ and anthropogenically altered areas such as lawns, plantation and developed areas are grouped under ‘transformed habitat’. Table 1 provides a summary of the size and habitat types within each reserve. A more detailed description of each reserve can be found in the electronic supplementary information (ESM 1).
Camera trap survey
Ninety un-baited Bushnell® Trophy Cam infrared remote sensing cameras (number × model = 64 × 119437, 5 × 119678, 12 × 119736, 1 × 119774, 1 × 119837 and 7 × 119876) were used to record the presence of medium to large mammal species in each reserve. Sampling took place over 21 months from June 2017 to Feb 2019, and multiple reserves were sampled simultaneously when possible. Infrared cameras were chosen due to high human activity and risk of theft in most of the reserves.
Camera placement
Decisions on camera trap placement were made based on the reserve size, habitat heterogeneity, camera theft risk, accessibility and obstructions posed by infrastructure (e.g. parking lots, building clusters) with the aim to exhaustively trap each reserve until all medium and large mammal species present were detected at least once. Maps for each reserve were created in QGIS v2.18.23 (QGIS Development Team 2019) to indicate the five major habitat types. The fynbos, renosterveld and strandveld areas were demarcated using the bioregion maps of Rebelo et al. (2006) and SANBI (2016), and the water-associated areas followed the azonal demarcations of the same map series. Vegetation type delineations were refined using aerial imagery (CCT 2019c) and ground-truthing when walking the reserves on foot. A 1 × 1 km grid layer was projected over each reserve area within QGIS and adjusted to ensure best fit (i.e. grid alignment was adjusted to ensure each reserve area was covered with the fewest grid squares possible) and a camera placement point identified as close to the centre of each 1 × 1 km block that fitted entirely within a reserve. If this protocol under-represented one of the four major natural habitat types (i.e. fynbos, strandveld, renosterveld, water-associated) an additional camera was placed in that habitat type to ensure that potential habitat specialists were sampled (O’Brien 2008). Areas with high risk of camera theft were avoided. A minimum of five cameras were placed in each reserve, ensuring that a minimum of 1000 camera days could be achieved within a maximum period of 200 survey days. In Steenbras Nature Reserve (8400 ha), the number of cameras available was insufficient to cover the whole reserve in one sampling event. Consequently, the total area was sub-sampled ensuring that representation of all habitat types was included. The cameras were placed within the different habitats at similar densities (i.e. minimum one camera per 1 × 1 km block) to the other smaller reserves to ensure spacing was similar.
Camera placement was optimised to detect mammals following the methods of Colyn et al. (2017). This involved searching for sign of mammal presence (e.g. scat, spoor, and foraging signs) within a 120 m radius of the grid point. If no signs were found within 120 m of the grid centre then cameras were placed as close to the grid point as possible without compromising camera safety. Each camera was fixed to a wooden pole with the camera lens at 30 cm above ground level (Tobler et al. 2008) and made to face either north or south so as to prevent false triggers and/or over-exposure from direct sunlight. Cameras were set to take a burst of three photos when triggered, with a delay of 30 s between trigger events (Colyn et al. 2017). Due to prevailing wind activity and the need to adequately hide each camera to prevent theft, sensitivity was set to medium and vegetation within a one metre arc of the camera lens was cut to reduce vegetation movement triggering the camera. Cameras were serviced every 20–30 days, where SD cards were changed and any potential problems with cameras addressed.
Survey effort
Each reserve was surveyed for a minimum 1000 camera days before cameras were removed/moved to another reserve. This was to enable capture of most species (Si et al. 2014; Colyn et al. 2017). If a species accumulation curve for a specific reserve did not reach an asymptote after a 1000 days, the survey period for that reserve was extended to attempt to reach the number of days required for adequate detection of all species, as time and resources allowed. EstimateS v9.1.0 software (Colwell 2013) was used to generate sample-based species accumulation curves for the recorded native species of each reserve and to determine whether the survey effort in each reserve was sufficient (Si et al. 2014; Colyn et al. 2017). The generated curves show the cumulative number of species recorded over sampling effort and were generated using 1000 randomized runs, with number of samples represented by the number of survey days (Olwell et al. 2004; Mann et al. 2015). Non-parametric species richness estimators (incidence coverage estimator (ICE), Chao 2, Jack 1 and Jack 2) were used to estimate how many species may have been missed during sampling. Accumulation and estimator curves for each reserve were compared to determine whether sampling effort for this study was sufficient and to estimate what can be considered as sufficient survey effort for the future monitoring of each area (Chao and Chiu 2016).
A robust estimate of sampling effort required to adequately survey a reserve was considered to be the point where the richness estimates of all four estimators reach an asymptote and where the variance between the four estimators richness values was at its lowest. In this way no particular estimator was favoured over another and it reduced any particular estimator’s chance of biasing interpretation.
Species richness and community composition
Camera trap data was managed using Camera Base v1.7 software (Tobler 2015). Only photographs of non-burrowing mammals exceeding 0.5 kg in weight were used for analysis. Mole-rat species (i.e. Cape dune mole-rat - Bathyergus suillus and Cape mole-rat - Georychus capensis) were thus excluded. Non-native and reintroduced medium and large mammal species were noted but also excluded from richness analyses.
Species richness predictor variables
Aerial photography (CCT 2019c), vegetation shapefiles (SANBI 2016) and official City of Cape Town mapped boundaries (CCT 2019b) together with ground-truthing and reserve manager liaison were used to extract information on variables hypothesised to influence species richness patterns in small urban reserves. Variables included reserve size (Diamond 1975; Matthies et al. 2017), area-perimeter ratio (Lagro 1991; Helzer and Jelinski 1999; Ewers and Didham 2007; Herse et al. 2018), habitat heterogeneity (Ramesh et al. 2016; Matthies et al. 2017), connectivity (Stevens et al. 2006; Turgeon and Kramer 2012; Correa Ayram et al. 2016), and the presence of permanent freshwater aquatic habitat (Okes and O’Riain 2017).
Reserve size and area-perimeter ratio
The perimeter (km) and area (km2) of each reserve were calculated using QGIS v2.18.23 software (QGIS Development Team 2019). An area-perimeter ratio was then determined for each reserve by dividing the area of the reserve by the perimeter length. This proportion is considered to be indicative of the ratio of core habitat relative to edge (Helzer and Jelinski 1999) with high ratio values indicating a greater proportion of interior habitat relative to edge habitat.
Habitat heterogeneity
The area (ha) of each of the five habitat types within each reserve was calculated using GIS vegetation shapefiles (SANBI 2016) and aerial photography (CCT 2019c) for transformed areas. These absolute values were then used to calculate a Shannon-Wiener diversity index for each reserve to appraise large-scale habitat heterogeneity (Matthies et al. 2017) using the formula:
where H = habitat diversity index, s = number of habitat types present, and pi = proportion of the total area (ha) of the reserve (Shannon 1948). The higher the index value, the more diverse or heterogeneous the reserve is believed to be from a habitat type perspective.
Permanent freshwater aquatic habitat
If a reserve possessed at least one permanent wetland such as a perennial river/stream and/or large water body (dam, vlei, etc.) it was considered as having suitable habitat for a water-associated medium to large mammal species. Results were classified as binary indicator variables, with presence of permanent freshwater habitat indicated by a score of 1 and absence by a score of 0.
Connectivity
Connectivity is comprised of both a structural and functional component which is nearly impossible to integrate into a single continuous numerical variable that will be relevant and accurate to all mammal species (Taylor et al. 2006). This is especially true where the connectivity of multiple reserves with different mammal communities are considered. For the purpose of this study a subjective categorical grouping approach was used to classify reserves. Linear features of the landscape such as the size and shape of corridors and the size of the land to which a corridor connects were integrated with knowledge on the quality of both the corridor habitat and the land to which a corridor connects. Disturbed or transformed areas covered by non-native vegetation were considered as functional habitat if perceived to mimic the structure of native vegetation that can potentially provide a form of cover to facilitate movement (e.g. stands of invasive alien plants) or habitation.
Analyses
Only native species believed to have persisted in the reserves despite anthropogenic pressure were included in analyses, i.e. re-introduced or alien species were not included. Pairwise plots and correlation coefficients were used to determine which covariates may be correlated with species richness, as well as to identify potential covariate collinearity in R v3.5.3 (R Core Team 2019). Collinearity between covariates was tested for in the car package (Fox and Weisberg 2019) using variance inflation factors (VIF) with species richness as the response variable. Any covariates with VIF scores of >5 and correlation coefficients of >0.7 were considered to have collinearity and were modelled separately (Dormann et al. 2013).
Because covariates contained no random effects and only one value per site, simple linear regression models were used to assess drivers of species richness. Linear regression models were run using various combinations of covariates and due to small sample size (n = 11) only two covariates were included in each model. Models were ranked according to second-order Akaike information criterion (AICc) calculated using the AICcmodavg package (Mazerolle 2019). The models with the lowest ∆AICc scores (difference between the model and lowest AICc score) were considered likely to predict species richness (Burnham et al. 2004). P values and F-statistics were also compared to verify which model was most parsimonious in predicting species richness.
Non-metric multidimensional scaling (NMDS) ordination was conducted using the vegan package (Oksanen et al. 2019) in R v3.5.3 (R Core Team 2019) to provide a visual representation of similarities between reserves based on medium and large mammal community structure (using Jaccard index for species presence/absence) and non-correlated species richness predictor variables.
Results
Sampling took place between June 2017 and February 2019 and cameras were placed at 151 different locations across the 12 reserves that cover a total area of 14,429 ha. Survey effort culminated in 14,876 camera days (Table 2). Five cameras were stolen from five different reserves within the sampling period, and in these cases either a new camera was placed in another location within the reserve for the required survey days or the remaining cameras were left in situ for longer to make up for the lost camera days. A total of 13,360 trigger events by medium and large mammals was recorded.
Survey effort
For Kenilworth Racecourse Conservation Area only one native species, the Cape grysbok (Raphicerus melanotis), was recorded in 1000 camera days so no species richness curve could be generated, nor species richness results further analysed. Eight of the remaining 11 reserves’ species richness estimators converged and reached asymptotes before a survey effort of 1000 camera days were reached (Fig. 2). Steenbras nature reserve needed 1560 days to reach an asymptote, while the accumulations curves of Helderberg and Witzands did not converge, nor reached an asymptote despite more than 1800 camera days of effort. Species richness estimates for these two reserves were consequently calculated at 13 for Helderberg and 15 for Witzands respectively by averaging richness estimators.

Sample-based species accumulation curves [S(est)] and non-parametric species richness estimations for each study site. The vertical dotted line indicates the number of camera days at which variance between all curves is at its lowest
Species richness and community composition
Cape hare (Lepus capensis) and scrub hare (Lepus saxatilis) could not be reliably differentiated from infrared photographs, so for the purposes of this study, Cape hare and scrub hare were thus grouped as “Lepus spp.”. A total of 27 medium and large mammals were recorded across the 12 reserves. Of these, five were non-native species (domestic cat - Felis sylvestris catus, domestic dog - Canus lupus familiaris, domestic rabbit - Oryctolagus cuniculus, domestic horse - Equus ferus caballus, and eastern grey squirrel - Sciurus carolinensis) and three were reintroduced native species (common eland - Tragelephus oryx, red hartebeest - Alcelaphus buselaphus caama and hippopotamus - Hippopotamus amphibious). Nineteen native medium to large mammal species that are believed to have persisted in the areas despite anthropomorphic pressure were recorded across the 12 nature reserves and were used for further analysis. In the case of Kenilworth Racecourse Conservation Area, the only recorded species, the Cape grysbok, has been reintroduced after becoming locally extinct (CCT 2019d) and the site is consequently excluded from further discussion and analysis. Species richness of the 19 focal species varied across the 11 remaining study sites ranging from three (Uitkamp Wetland) to 15 species (Witzands Aquifer) (Table 3).
More carnivore species (n = 11) than herbivores (n = 7) were recorded, and only one omnivore, the chacma baboon (Papio ursinus) was detected. Mean and median estimated species richness across the reserves is eight species, with the mode at five species. Cape porcupine (Hystrix africaeaustralis) was present in all 11 reserves, with Cape grysbok and small grey mongoose (Galerella pulverulenta) being recorded in 10 and nine of the reserves respectively. Klipspringer (Oreotragus oreotragus) and Hewitt’s red rock hare (Pronolagus saundersiae) were only recorded in Steenbras and Cape fox (Vulpes chama) only in Witzands.
Calculated species richness predictor variables
Reserve sizes ranged from 0.32 to 84 km2 (mean = 13.07 ± 23.03) (Table 4). Area to perimeter proportions varied from 0.05 for Uitkamp to 1.69 for Steenbras (mean = 0.46 ± 0.48), with values for the largest reserves (Witzands and Steenbras) showing the largest area in proportion to reserve edge (>1). Blaauwberg, Witzands and Steenbras are the only three reserves with an area-perimeter ratio of more than 0.6.
Habitat heterogeneity scores varied from 1.28 in Table Bay to 0 for Wolfgat which had only one habitat type present (strandveld). The mean habitat heterogeneity score was 0.56 ± 0.35. Permanent freshwater aquatic habitat was present at seven of the 11 reserves.
Application of the connectivity criteria to the 11 reserves resulted in reserves being placed in five categories of connectivity (Table 5). Steenbras and Helderberg were exceptionally well connected to additional natural habitat, forming part of the same much larger unfragmented conserved landscape. Four reserves had very little to no connectivity due to potential corridors being narrow and surrounded by urban structures. The remaining five reserves had connectivity to agricultural land exposed to different levels of isolation, utilization and transformation that resulted in them being divided into three separate classes.
Modelling of predictor variables and non-metric multidimensional scaling (NMDS)
Strong correlations are evident between reserve size and area-perimeter ratio (correlation coefficients of 0.911), and also between connectivity and area-perimeter ratio (0.742) (Table 6). This is confirmed by the VIF scores for reserve size and area-perimeter ratio which produce values >10, indicative of strong collinearity (Dormann et al. 2013). Reserve size, area-perimeter ratio and connectivity were thus modelled separately from each other. The correlation matrix also indicates a significant positive correlation between species richness and connectivity (0.88), as well as between species richness and area-perimeter ratio (0.68).
Ten different linear regression models were run using individual variables first in order to rank model fit and thus discard the least significant collinear models, then additional models were run using combinations of the remaining covariates (Table 7).
The most parsimonious model explaining species richness was connectivity (p = 0.0003). It also yielded the highest model weight (0.860), highest F-statistic (31.34) and lowest standard error (1.957). It was a stronger fit than the collinear covariates of area-perimeter ratio (∆AICc = 9.801, model weight = 0.0.006) and reserve size (∆AICc = 14.172). The next best fitting model was the combination of connectivity and permanent freshwater aquatic habitat (∆AICc = 5.216), but is similar in predictive power to a combination of connectivity ratio and habitat heterogeneity (∆AICc = 5.230). Both of these models are different from the minimum AIC score by >4, thus there is not much support for either model (Burnham et al. 2004). Water and heterogeneity considered separately were not found to be significant predictors of species richness (∆AICc >15), and even less so when combined (∆AICc = 21.061). When richness is plotted as a function of connectivity (Fig. 3), seven of the observed values fall within the 95% confidence intervals of the model. The R2 value of 0.7521 and p = 0.0003 indicates a strong and significantly positive linear relationship between species richness and the connectivity scores.

Linear model (solid line) fit where species richness is a function of connectivity (R2 = 0.7521 and p = 0.0003). Shaded area indicates 95% confidence intervals and points indicate observed species richness values
The NMDS ordinance indicates connectivity as the strongest driver of species richness (Fig. 4), and as expected associates strongest with Steenbras and Helderberg which holds the highest connectivity scores. These two reserves have nine species in common, of which six (klipspringer, Hewitt’s red rock hare, leopard - Panthera pardus, chacma baboon, honey badger - Mellivora capensis, common duiker - Sylvicapra grimmia) have a strong relationship with connectivity.

Non-metric dimensional scaling (NMDS) plot of reserve similarity using species presence and uncorrelated species richness predictor covariates. Reserves are indicated by capital letters (red): UK (Uitkamp Wetland), TB (Table Bay), FB (False Bay), TY (Tygerberg), ZV (Zandvlei Estuary), WG (Wolfgat), ST (Steenbras), HB (Helderberg), BR (Bracken), WZ (Witzands Aquifer) and BB (Blaauwberg). Blue lines indicate the strength of predictor variables: Heterogeneity (habitat heterogeneity), Freshwater (presence of permanent freshwater aquatic habitat), and Connectivity. Species are indicated in green text with abbreviations: WMg (water mongoose), LMg (large grey mongoose), SMg (small grey mongoose), LSG (large-spotted genet), SSG (small-spotted genet), otter (Cape clawless otter), porc (Cape porcupine), grysbok (Cape grysbok), fox (Cape fox), polecat (striped polecat), hare (Cape/scrub hare), HRR (Hewitt’s red rock hare), baboon (chacma baboon), badger (honey badger) and duiker (common duiker). A “+” indicates that species overlap, i.e. have the same centre point
Uitkamp Wetland Nature Reserve, which falls into the lowest connectivity category and has an intermediate heterogeneity score, does not associate closely with any other reserves. However, both freshwater habitat and the presence of large grey mongoose (Herpestes ichneumon) make it most similar to Table Bay and False Bay nature reserves. False Bay, Zandvlei and Wolfgat cluster at the bottom of the plot and all three reserves comprise of large proportions of strandveld vegetation and have four mammal species (Cape porcupine, grysbok, small grey mongoose and large-spotted genet - Genetta tigrina) in common. Witzands and Blaauwberg group together with both having no water-associated habitat and the same high connectivity score. Bracken, Witzands and Blaauwberg reserves have no permanent freshwater aquatic habitat and share the rarer steenbok (Raphicerus campestris) and striped polecat (Ictonyx striatus). Unsurprisingly the NMDS plot shows a strong association between Cape clawless otter and reserves with permanent freshwater aquatic habitat.
Discussion
The greatest threat to biodiversity in the Cape Floristic Region, a recognized global biodiversity hotspot, is agricultural and urban expansion (Rebelo et al. 2011). Both of these human activities result in the loss and fragmentation of natural habitat, with fragments becoming increasing isolated as anthropogenic land use expands and densifies (McKinney 2002; De Stefano and De Graaf 2003; Rebelo et al. 2011; Pickett et al. 2011; Ramesh et al. 2016). In this study the influence of patch and landscape level variables on the species richness of mammals in nature reserves in the City of Cape Town was assessed. Connectivity was found to be the most important factor explaining mammal species richness in the 11 city nature reserves sampled. The metric for connectivity included both structural and functional components, viz. corridor size, matrix composition and the amount of suitable land adjacent to the reserves surveyed. Higher values of connectivity thus include larger corridors comprised of habitat more suitable for dispersal and linkages to larger amounts of preferred habitat (Table 5). Reserves with the highest connectivity scores, characterized by long permeable boundaries (>5 km) to mixed use landscapes or direct connections to extensive areas of suitable natural habitat had the highest observed and predicted levels of mammal species richness.
This finding supports previous research showing that the amount of suitable habitat adjacent to a patch is important for maintaining biodiversity (Crooks 2002; Hurst et al. 2013; Correa Ayram et al. 2016; Fahrig et al. 2019). Access to large amounts of suitable habitat improves both survival and reproduction (Jewitt et al. 2015) while large corridors allow for ease of dispersal and the re-colonisation of small reserves that are vulnerable to stochastic local extinction events (Tischendorf and Fahrig 2000; Boscolo and Metzger 2011; Fahrig et al. 2019). The NMDS plot (Fig. 3) revealed the close association between the two reserves which are well connected with large amounts of suitable habitat (i.e. Steenbras and Helderberg) and they have the only large carnivore remaining in Cape Town reserves, namely the leopard present. Leopards in particular have large home ranges (up to 900 km2) within the Western Cape mountain habitat (Martins and Harris 2013), and could not persist in small isolated reserves, but may pass through them provided they are connected to larger areas with suitable habitat. Both reserves are unusual amongst those surveyed in having rugged mountain habitat which is unsuitable for both urban and rural land uses and has been left functionally intact. Unsurprisingly therefore these were also the only reserves which between them still have rocky habitat specialists such as klipspringer and Hewitt’s rock rabbit (Matthee et al. 2016; Druce et al. 2009) and the more generalist chacma baboon which shows a preference for steep slopes as a refuge from predators (Hoffman and O’Riain 2012b).
Taylor et al. (1993) defined landscape connectivity as the degree to which the landscape facilitates or impedes movement among resource patches. Reserves in Cape Town that are surrounded by dense networks of roads and residential suburbs with either small corridors or corridors that required animals to traverse transformed habitat (e.g. agriculture and urban land use) had lower species richness. This trend was apparent for both medium sized reserves like Zandvlei and small reserves like Bracken, highlighting the importance of connectedness relative to reserve size. It is further important to note that Helderberg is a medium sized reserve relative to those included in this study, but has the second highest levels of observed and expected species richness. This is attributed to its high functional connectivity with physical parameters (i.e. corridor size) driving high structural connectivity and biotic elements (i.e. natural vegetation in corridors) promoting animal movement into and out of the reserve (Taylor et al. 1993; Tischendorf and Fahrig 2000). It is important to note that while most reserves in this study had fences demarcating their borders, none of these were designed to prevent the movement of any of the detected species presented in Table 3. Rather, fences served primarily to indicate to people that they were entering a protected area. The only exception to this was Kenilworth Racecourse which, as a horse racing venue, must prevent access to people and domestic animals. High walls surround the reserve separating it from the surrounding matrix of dense urban development and roads. This effectively makes this the most isolated of all reserves in Cape Town. It is thus illuminating that only one species of mammal, the common grysbok, manages to survive in this urban reserve and that is only after reintroduction (CCT 2019d).
It was predicted that both an increase in reserve size and area-perimeter ratio would result in higher levels of species richness. Higher species richness values for larger patches and patches with shape characteristics that maximize suitable habitat relative to edges is well supported by theory and substantiated by empirical proof for a number of taxa (Diamond 1975; Helzer and Jelinski 1999; Orrock et al. 2003; Ewers and Didham 2007; Nams 2011; Herse et al. 2018). While both variables were positively correlated with species richness (Table 6) they were also collinear to connectivity which had the strongest univariate predictive power, followed by area perimeter ratio and then reserve size (Table 6). Models which included these factors did not perform as well as that which included connectivity and hence both were excluded from the NMDS plot. Neither habitat heterogeneity nor permanent freshwater aquatic habitat were strong predictors of species richness in the study area.
Habitat heterogeneity showed a weak negative correlation (−0.24) with species richness (Table 6). This contradicts a body of literature suggesting that increased habitat heterogeneity positively influences species richness in an area (Ramesh et al. 2016; Matthies et al. 2017). While the fynbos, strandveld and renosterveld vegetation classifications used here differ in floral species composition and soil nutrient status, they may provide similar structural cover (low shrubland) for species (Rebelo et al. 2006). Separating vegetation types as fynbos, strandveld and renosterveld in this context might thus only be relevant to large obligate grazer species such as Cape mountain zebra that seeks high quality grass fodder mostly associated with the more nutrient rich renosterveld areas (Radloff et al. 2010; Watson and Chadwick 2007). Secondary consumers such as mesopredators and small bodied herbivores that selectively browse from only the most nutritious parts of shrubs and forbs (e.g. Cape grysbok, steenbok and duiker, Skinner and Chimimba 2005) might not differentiate between the three vegetation complexes in terms of functional habitat. Inclusion of additional parameters to define habitat such as vegetation structure or terrain ruggedness may therefore be more relevant when exploring species richness patterns and is something that future studies should consider including. It is also possible that in the case of mesopredators, release from larger predators (Brassine and Parker 2012) in addition to access to domestic and synanthropic food sources, are such important drivers of species richness in small reserves that the effects of vegetation type are largely negated. This combination of factors may explain why Tygerberg, with no apex predators and large edges with farmland that support abundant synanthropic, domestic and exotic prey species (Serieys et al. 2019) had the most carnivore species (n = 8).
Seasonal differences in survey time between reserves (Table 1) could have influenced richness estimates if certain species are only present in specific seasons, but it is considered unlikely. None of the detected herbivore species are known to migrate on a seasonal basis and all three detected antelope species (Cape grysbok, steenbok and duiker) hold small permanent territories (Skinner and Chimimba 2005). Porcupine might be drawn to reserves at times when geophytes sprout (Bragg et al. 2005), but this species was found in every single reserve except Kenilworth Racecourse which was surveyed from May until December and thus included the flowering spring season. The absence of easily accessible surface water for drinking during the dry season (Nov – March, Rebelo et al. 2006) might also affect some species, but the three reserves (Helderberg, Table bay and Zandvlei) predominantly surveyed during the dry period have permanent freshwater bodies present. Species reliant on the presence of freshwater aquatic habitat were accounted for in the analysis and explains the selective presence of two water specialists viz. Cape clawless otter and water mongoose. Curiously, large grey mongoose clusters close to otters in the NMDS plot (Fig. 4) but is not known to associate closely with permanent freshwater aquatic habitat (Palomares and Delibes 1990).
Wildlife winners and losers in the development of Cape Town
Historical accounts suggest that 40 medium and large mammal species (39 if scrub and Cape hare are pooled) were present in the general area at the time of European settlement in 1652 (Boshoff and Kerley 2001; Skead 2011). Of these, only 19 (49%) seem to have survived urbanization and managed to persist in the City of Cape Town reserves to this day. Larger species such as African elephant (Loxodonta africana), black rhinoceros, Cape mountain and plains zebras (Equus zebra zebra and E. quagga), lion, brown hyena (Hyena brunnea), spotted hyena (Crocuta crocuta) and wild dog (Lycaon pictus) were not expected to be recorded, as it is known that they were hunted to local extinction in the 1700s (Rebelo 1992). The absence of large mammals is also to be expected as mammals with large body sizes tend to require large home ranges and are thus more sensitive to habitat loss and fragmentation linked to urban and rural development (McCleery 2010). This trend is particularly true for large carnivores which require extensive connected ranges to complete their life history (Hansen et al. 2011). Hippopotamus, common eland and red hartebeest, while reintroduced to two reserves (hippopotamus and eland into False Bay Nature Reserve, eland and red hartebeest into Blaauwberg Nature Reserve) (CCT 2019d) were also not expected to have persisted unaided in local reserves given their large space requirements.
Small grey mongoose, Cape porcupine and Cape grysbok were recorded in the majority of reserves. Globally, species which are able to generalize in terms of food sources and habitat preference seem to thrive in urban areas due to varied combinations of increased food availability and absence of competition and predation pressure from species which are more sensitive to reduce home ranges and disturbance (De Stefano and De Graaf 2003; Baker and Harris 2007; Ordeñana et al. 2010; Newsome et al. 2010; Hoffman and O’Riain 2012b; Šálek et al. 2015). Cape porcupine, which readily raid domestic gardens, and small grey mongoose, which are able to predate on a variety of food sources including small mammals and insects which are readily available in transformed habitats (Cavallini and Nel 1990), probably fit the profile of generalists. However, Cape grysbok is endemic to the fynbos biome and is a highly selective browser (Kigozi et al. 2008) and they may be present mainly due to the availability of fynbos-specific habitat requirements rather than being adaptable generalists. Their common occurrence across reserves suggests that they can survive and even thrive within small areas with some protection against anthropogenic pressure. Their success may also be from reduced predation pressure in small urban reserves, however caracal (Caracal caracal), which are also present in many reserves, are known to predate on them (Leighton et al. 2020). Additionally, if only a lack of predators were to explain grysbok’s wide occurrence, similar results would be expected for steenbok and duiker, which are far less common.
Eight species that were presumed to still be present at the start of the study based on anecdotal reports and CCT species inventories (2012–2017) were not recorded and include grey rhebuck (Pelea capreolus), black-backed jackal (Canis mesomelas), bat-eared fox (Otocyon megalotis), yellow mongoose (Cynictis penicillata), striped weasel (Poecilogale albinucha), aardwolf (Proteles cristata), aardvark (Orycteropus afer) and African wild cat (Felis sylvestris cafra). African wild cat was not expected in this study, as it is assumed that any surviving cats within the CCT area would have interbred with domestic cats and potentially be locally extinct. Yellow mongoose has been anecdotally sighted at both Tygerberg and Uitkamp Wetland nature reserves, but has not been included in any database since 2011 and was not recorded in the study. Aardwolf have never been recorded on the CCT Biodiversity Database (CCT 2019d) and aardvark have not been recorded since 2004. It may be that large-scale development in urban areas reduce substrate for termite colonies and thus deprive them of sufficient food.
Grey rhebuck had previously been reintroduced to both the Tygerberg and Helderberg Nature Reserves to bolster population numbers (CCT 2019d). Despite this the species was not recorded in this study. The last anecdotal sighting in at Tygerberg Nature Reserve was in 2017, but a local extinction event was thought to have occurred shortly after a carcass was found and no further observations of the species have been made since. The current presumed presence of bat-eared fox and black-backed jackal within the study sites is largely uncertain as no official records have been logged in any reserve since 2012 and 2013 respectively. Striped weasel was last recorded on the Biodiversity Database at Helderberg Nature Reserve in 2013 and has been recorded elsewhere in the CCT area previously (Child et al. 2016), but it is thought that the study sites may have been at the edge of its range as the majority of sightings tend to occur toward the eastern boundary of the CCT municipal area. It thus appears that at least seven species that were still recorded in the past decade might now be extinct from the surveyed nature reserves.
Conclusions
Globally the recent trend has been for smaller patches (<100 ha) of natural habitat to be preserved as urban nature reserves (Volenec and Dobson 2020). Species in small reserves are more vulnerable to both edge effects and stochastic events which may over time reduce species richness if the surrounding matrix is unsuitable for animal movement to or from other patches of suitable habitat (Guirado et al. 2006; Zungu et al. 2019). This study’s findings support a large body of literature suggesting that maintaining species richness within these small patches is best done by ensuring good connectivity to suitable habitat. This suggests that natural and even low intensity agricultural areas (e.g. vineyards) that are tolerant of wildlife should be prioritised for retention in the land use matrix rather than creating narrow corridors of transformed land. While acquisition of new land for expansion of conservation areas may be difficult in an urban context, the development of stewardship agreements with neighbouring landowners may assist in securing existing corridors and suitable habitat to maintain species richness.
Small reserves and fragments tend not to support charismatic mammalian megafauna (typically area-sensitive and low-density species), which explains why most species in sampled CCT reserves were small and medium sized. Only reserves directly connected to extensive tracts of natural land (Steenbras and Helderberg) had retained an apex predator with the result that most urban reserves would have experienced the release of both mesopredators and prey species with the associated trophic cascades (Turner and Corlett 1996; Terborgh et al. 2001). As predicted, small sized generalists (e.g. Cape porcupine and grey mongoose) were the most common species and carnivores were better represented (n = 11) than herbivores (n = 7), although this ratio (1.6:1) is not dissimilar to that of species believed to have historically occurred in the area (1.5:1) (Rookmaaker 1989; Boshoff and Kerley 2001; Skead 2011). Habitat specialists including klipspringer and Hewitt’s red rock hare, and Cape clawless otter and water mongoose were only present in those reserves that provided rocky outcrops and permanent freshwater aquatic habitat respectively. Surprisingly a habitat specialist, the Cape grysbok, was found to be widely distributed, and seems capable of persisting in even small fragments of natural habitat.
The study shows that while a number of historically-occurring medium and large mammal species have already been lost in the CCT area, close to half of these have persisted despite urbanisation. Mammal species richness recorded in this study varied widely within the 12 CCT nature reserves surveyed but collectively they currently provide a refuge for 19 surviving medium and large mammal species. As natural habitat is lost from the landscape, small reserves, in addition to enhancing connectivity, can increase matrix quality through the retention of natural habitat in the landscape, which benefits species richness and abundance in reserves and fragments (Hurst et al. 2013; Correa Ayram et al. 2016).
References
Anderson PML, O’Farrell PJO (2012) An ecological view of the history of the establishment of the City of Cape Town. Ecol Soc 17:28–39
Augustine DJ, McNaughton SJ (1998) Ungulate effects on the functional species composition of plant communities: herbivore selectivity and plant tolerance. J Wildl Manag 62:1165–1183
Baker PJ, Harris S (2007) Urban mammals: what does the future hold? An analysis of the factors affecting patterns of use of residential gardens in Great Britain. Mammal Rev 37:297–315
Bateman PW, Fleming PA (2012) Big city life: carnivores in urban environments. J Zool 287:1–23
Bernardo PVS, De Melo FR (2013) Assemblage of medium and large size mammals in an urban Semideciduous seasonal Forest fragment in Cerrado biome. Biota Neotropica 13:76–80
Boscolo D, Metzger JP (2011) Isolation determines patterns of species presence in highly fragmented landscapes. Ecography 34:1018–1029
Boshoff AF, Kerley GIH (2001) Potential distributions of the medium- to large-sized mammals in the cape floristic region, based on historical accounts and habitat requirements. Afr Zool 36:245–273
Bragg C, Donaldson JS, Ryan PG (2005) Density of cape porcupines in a semi-arid environment and their impact on soil turnover and related ecosystem processes. J Arid Environ 61:261–275
Brassine MC, Parker DM (2012) Does the presence of large predators affect the diet of a mesopredator? Afr J Ecol 50:243–246
Brooker L, Brooker M, Cale P (1999) Animal dispersal in fragmented habitat: measuring habitat connectivity, corridor use, and dispersal mortality. Conserv Ecol 3:4
Burnham KP, Anderson DR, Anderson DR (2004) Multimodel inference: understanding AIC and BIC in model selection. Sociological Methods Research 33:261–304
Cavallini P, Nel JAJ (1990) The feeding ecology of the cape grey mongoose, Galerella pulverulenta (Wagner 1839) in a coastal area. Afr J Ecol 28:23–130
Ceballos G, Ehrlich PR (2006) Global mammal distributions, biodiversity hotspots, and conservation. Proc Natl Acad Sci 103:19374–19379
Ceballos G, Ehrlich PR, Soberon J, Salazar I, Fay JP (2005) Global mammal conservation: what must we manage? Science 309:603–607
Chao A, Chiu C-H (2016) Nonparametric estimation and comparison of species richness. eLS:1–11
Child MF, Rowe-Rowe D, Birss C, Wilson B, Palmer G, Stuart C, Stuart M, West S, Do Linh San E (2016) A conservation assessment of Poecilogale albinucha. In: Child MF, Roxburgh L, Do Linh San E, Raimondo D, Davies-Mostert HT (eds) The red list of mammals of South Africa, Swaziland and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa
City of Cape Town (2019a) Open Data Portal: Integrated Zoning Land Parcel. https://web1.capetown.gov.za/web1/OpenDataPortal/DatasetDetail?DatasetName=Integrated%20zoning%20land%20parcel. Accessed 4 April 2019
City of Cape Town (2019b) Open Data Portal: Terrestrial Biodiversity Network. http://odp.capetown.gov.za/datasets/4f2d7835518a4e6b8205ce12d77ff463_64. Accessed 4 April 2019
City of Cape Town (2019c) Open Data Portal: Aerial photography. https://web1.capetown.gov.za/web1/OpenDataPortal/DatasetDetail?DatasetName=Aerial%20photography. Accessed 25 July 2019
City of Cape Town (2019d) Biodiversity Database. https://biodiversity.co.za/. Accessed 4 April 2019
Colwell RK (2013) EstimateS version 9.1.0. Computer software, University of Connecticut, United States Of America
Colyn R, Radloff FGT, O’Riain MJ (2017) Camera trapping mammals in the scrubland’s of the cape floristic kingdom - the importance of effort, spacing and trap placement. Biodivers Conserv 27:503–520
Correa Ayram CA, Mendoza ME, Etter A, Salicrup DRP (2016) Habitat connectivity in biodiversity conservation: a review of recent studies and applications. Prog Phys Geogr 40:7–37
Cowling RM, MacDonald IAW, Simmons MT (1996) The cape peninsula, South Africa: physiographical, biological and historical to an extraordinary hot-spot of biodiversity. Biodivers Conserv 5:527–550
Crooks KR (2002) Relative sensitivities of mammalian carnivores to habitat fragmentation. Conserv Biol 16:488–502
Crooks KR, Burdett CL, Theobald DM, Rondinini C, Boitani L (2011) Global patterns of fragmentation and connectivity of mammalian carnivore habitat. Philosophical Transactions of the Royal Society B: Biological Sciences 366:2642–2651
De Stefano S, De Graaf RM (2003) Exploring the ecology of suburban wildlife. Front Ecol Environ 1:95–101
Diamond JM (1975) The island dilemma: lessons of modern biogeographic studies for the design of natural reserves. Biol Conserv 7:129–146
Dormann CF, Elith J, Bacher S, Buchmann C, Carl G, Carr G, Garc JR, Gruber B, Lafourcade B, Leit PJ, Tamara M, McClean C, Osborne PE, Der BS, Skidmore AK, Zurell D, Lautenbach S (2013) Collinearity : a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36:27–46
Druce DJ, Brown JS, Kerley GIH, Kotler BP, MacKey RL, Slotow R (2009) Spatial and temporal scaling in habitat utilization by klipspringers (Oreotragus oreotragus) determined using giving-up densities. Austral Ecology 34:577–587
Ewers RM, Didham RK (2007) The effect of fragment shape and species’ sensitivity to habitat edges on animal population size: contributed papers. Conserv Biol 21:926–936
Fahrig L, Arroyo-Rodríguez V, Bennett JR, Boucher-Lalonde V, Cazetta E, Currie DJ, Eigenbrod F, Ford AT, Harrison SP, Jaeger JA, Koper N (2019) Is habitat fragmentation bad for biodiversity? Biol Conserv 230:179–186
Fischer J, Lindenmayer DB (2007) Landscape modification and habitat fragmentation: a synthesis. Glob Ecol Biogeogr 16:265–280
Fischer JD, Cleeton SH, Lyons TP, Miller JR, Fischer JD, Cleeton SH, Timothy P (2012) Urbanization and the predation paradox: the role of trophic dynamics in structuring vertebrate communities. BioScience 62:809–818
Fox J, Weisberg S (2019) An {R} companion to applied regression. Third Edition. Thousand Oaks CA: Sage. https://socialsciences.mcmaster.ca/jfox/Books/Companion/
Garden J, McAlpine C, Peterson A, Jones D, Possingham H (2006) Review of the ecology of Australian urban fauna: a focus on spatially explicit processes. Austral Ecology 31:126–148
Gonçalves MSS, Gil-Delgado JA, Gosálvez RU, López-Iborra GM, Ponz A, Velasco Á (2018) Seasonal differences in drivers of species richness of waders in inland wetlands of La Mancha Húmeda biosphere reserve. Aquat Conserv Mar Freshwat Ecosyst 28:1414–1423
Guirado M, Pino J, Roda F (2006) Understorey plant richness and composition in metropolitan forest archipelagos: effects of forest size, adjacent land use and distance to the edge. Glob Ecol Biogeogr 15:50–62
Hansen AJ, Knight RL, Marzluff JM, Powell S, Gude PH, Jones K (2011) Effects of exurban Development on biodiversity: patterns, mechanisms, and research needs. Ecol Appl 15:1893–1905
Hardt RA, Forman RTT (1989) Boundary form effects on Woody colonization of reclaimed surface mines. Ecology 70:1252–1260
Harris C, Burgers C, Miller J, Rawfoot F (2010) O- and H-isotope record of Cape Town rainfall from 1996 to 2008. S Afr J Geol 113:33–56
Heegaard E, Økland RH, Bratli H, Dramstad WE, Engan G, Pedersen O, Solstad H (2007) Regularity of species richness relationships to patch size and shape. Ecography 30:589–597
Helzer CJ, Jelinski DE (1999) The relative importance of patch area and perimeter-area ratio to grassland breeding birds. Ecol Appl 9:1448–1458
Herse MR, With KA, Boyle WA (2018) The importance of core habitat for a threatened species in changing landscapes. J Appl Ecol 55:2241–2252
Hoffman TS, O’Riain MJ (2012a) Troop size and human-modified habitat affect the ranging patterns of a chacma baboon population in the cape peninsula, South Africa. Am J Primatol 74:853–863
Hoffman TS, O’Riain MJ (2012b) Landscape requirements of a primate population in a human-dominated environment. Front Zool 9:1
Hoffman TS, O’Riain MJ (2012c) Monkey management: using spatial ecology to understand the extent and severity of human–baboon conflict in the cape peninsula, South Africa. Ecol Soc 17(3):13
Holmes PM, Rebelo AG, Dorse C, Wood J (2012) Can Cape Town’s unique biodiversity be saved? Balancing conservation imperatives and development needs Ecology and Society 17
Hurst ZM, McCleery RA, Collier BA, Fletcher Jr RJ, Silvy NJ, Taylor PJ, Monadjem A (2013) Dynamic edge effects in small mammal communities across a conservation-agricultural interface in Swaziland PLoS One 8
iNaturalist Network (2019) City Nature Challenge 2019. https://www.inaturalist.org/projects/city-nature-challenge-2019. Accessed 20 October 2019
Jewitt D, Goodman PS, Erasmus BF, O'Connor TG, Witkowski ET (2015) Systematic land-cover change in KwaZulu-Natal, South Africa: implications for biodiversity. S Afr J Sci 111:01–09
Kerley GIH, Pressey RL, Cowling RM, Boshoff AF, Sims-Castley R (2003) Options for the conservation of large and medium-sized mammals in the cape floristic region hotspot, South Africa. Biol Conserv 112:169–190
Kigozi F, Kerley GIH, Lessing JS (2008) The diet of cape grysbok (Raphicerus melanotis) in Algoa dune Strandveld, Port Elizabeth, South Africa. S Afr J Wildl Res 38:79–81
Kindlmann P, Burel F (2008) Connectivity measures: a review. Landsc Ecol 23:879–890
Lagro J (1991) Assessing patch shape in landscape mosaics. Photogrammetric Engineering, Remote Sensing 57:285–293
Leighton GR, Bishop JM, O’Riain MJ, Broadfield J, Meröndun J, Avery G, Avery DM, Serieys LE (2020) An integrated dietary assessment increases feeding event detection in an urban carnivore. Urban Ecosystems 20:1–5
Lowry H, Lill A, Wong BBM (2013) Behavioural responses of wildlife to urban environments. Biol Rev 88:537–549
Mann GKH, Lagesse JV, O’Riain MJ, Parker DM (2015) Beefing up species richness? The effect of land-use on mammal diversity in an arid biodiversity hotspot. African Journal of Wildlife Research 45:321–331
Martins Q, Harris S (2013) Movement, activity and hunting behaviour of leopards in the Cederberg mountains, South Africa. Afr J Ecol 51:571–579
Matthee C, Wilson B, Robinson TJ, Child MF (2016) A conservation assessment of Pronolagus spp. in: the red list of mammals of South Africa, Swaziland and Lesotho. Child MF, Roxburgh L, do Linh san E, Raimondo D, Davies-Mostert HT (ed) south African National Biodiversity Institute and endangered wildlife trust, South Africa
Matthies SA, Rüter S, Schaarschmidt F, Prasse R (2017) Determinants of species richness within and across taxonomic groups in urban green spaces. Urban Ecosyst 20:897–909
Mazerolle MJ (2019) AICcmodavg: model selection and multimodel inference based on (Q)AIC(c). R package version 2.2–2. https://cran.r-project.org/package=AICcmodavg
McCleery R (2010) Urban mammals. In: urban ecosystem ecology, agronomy monographs. 55. ASA, CSSA, SSSA, Madison, WI, pp 87-102
McDonald RI, Kareiva P, Forman RTT (2008) The implications of current and future urbanization for global protected areas and biodiversity conservation. Biol Conserv 141:1695–1703
McKinney ML (2002) Urbanization, biodiversity and conservation. BioScience 52:883–890
Nams VO (2011) Emergent properties of patch shapes affect edge permeability to animals PLoS ONE 6
Nattrass N, Drouilly M, O’Riain MJ (2020) Learning from science and history about black-backed jackals Canis mesomelas and their conflict with sheep farmers in South Africa. Mammal Rev 50:101–111
Newsome SD, Ralls K, Job CVH, Fogel ML, Cypher BL (2010) Stable isotopes evaluate exploitation of anthropogenic foods by the endangered San Joaquin kit fox (Vulpes macrotis mutica). J Mammal 91:1313–1321
O’Brien TG (2008) On the use of automated cameras to estimate species richness for large- and medium-sized rainforest mammals. Anim Conserv 11:179–181
Okes NC, O’Riain MJ (2017) Otter occupancy in the cape peninsula: estimating the probability of river habitat use by cape clawless otters, Aonyx capensis, across a gradient of human influence. Aquat Conserv Mar Freshwat Ecosyst 27:706–716
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Eduard Szoecs E, Wagner H (2019) Vegan: community ecology package. R package version 2.5-5. https://cran.R-project.org/package=vegan
Olwell RK, Chang XM, Jing C (2004) Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 85:2717–2727
Ordeñana MA, Crooks KR, Boydston EE, Fisher RN, Lyren LM, Siudyla S, Haas CD, Harris S, Hathaway SA, Turschak GM, Miles AK, Van Vuren DH (2010) Effects of urbanization on carnivore species distribution and richness. J Mammal 91:1322–1331
Orrock JL, Danielson BJ, Burns MJ, Levey DJ (2003) Spatial ecology of predator-prey interactions: corridors and patch shape influence seed predation. Ecology 84:2589–2599
Palomares F, Delibes M (1990) Habitat preference of large grey mongooses Herpestes ichneumon in Spain. Acta Theriol 35:1–6
Patterson L, Kalle R, Downs CT (2018) Factors affecting presence of vervet monkey troops in a suburban matrix in KwaZulu-Natal, South Africa. Landsc Urban Plan 169:220–228
Pekin BK, Pijanowski BC (2012) Global land use intensity and the endangerment status of mammal species. Divers Distrib 18:909–918
Pickett STA, Cadenasso ML, Grove JM, Boone CG, Groffman PM, Irwin E, Kaushal SS, Marshall V, McGrath BP, Nilon CH, Pouyat RV, Szlavecz K, Troy A, Warren P (2011) Urban ecological systems: scientific foundations and a decade of progress. J Environ Manag 92:331–362
QGIS Development Team (2019) QGIS geographic information system. Open Source Geospatial Foundation Project. http://qgis.osgeo.org
R Core Team (2019). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Radloff FGT, Mucina L, Bond WJ, le Roux PJ (2010) Strontium-isotope analysis of large herbivore habitat preferences in the cape Fynbos region. Oecologia 164:567–578
Ramesh T, Kalle R, Downs CT (2016) Predictors of mammal species richness in KwaZulu-Natal, South Africa. Ecol Indic 60:385–393
Rebelo AG (1992) Red data book species in the cape floristic region: threats, priorities and target species. Transactions Royal Society of South Africa 48:55–86
Rebelo AG, Boucher C, Helme N, Mucina L, Rutherford MC (2006). Fynbos biome. In: the vegetation of South Africa, Lesotho and Swaziland. Mucina L, Rutherford MC (ed) Strelitzia 19, south African National Biodiversity Institute, Pretoria, pp 52-219
Rebelo AG, Holmes PM, Dorse C, Wood J (2011) Impacts of urbanization in a biodiversity hotspot: conservation challenges in metropolitan Cape Town. S Afr J Bot 77:20–35
Rookmaaker LC (1989) The zoological exploration of southern Africa 1650–1790. A. A. Balkema, Rotterdam
Saito M, Koike F (2013) Distribution of wild mammal assemblages along an urban-rural-Forest landscape gradient in warm-temperate East Asia. PLoS One 8:e65464
Šálek M, Drahníková L, Tkadlec E (2015) Changes in home range sizes and population densities of carnivore species along the natural to urban habitat gradient. Mammal Rev 45:1–14
Serieys LEK, Bishop J, Okes N, Broadfield J, Winterton DJ, Poppenga RH, Viljoen S, Wayne RK, O’Riain MJ (2019) Widespread anticoagulant poison exposure in predators in a rapidly growing south African city. Sci Total Environ 666:581–590
Shannon CE (1948) A mathematical theory of communication. Bell System Technical Journal 27:379–423
Si X, Kays R, Ding P (2014) How long is enough to detect terrestrial animals? Estimating the minimum trapping effort on camera traps. PeerJ 2:e374
Skead CJ (2011) Historical incidence of the large land mammals in the broader northern and Western cape. 2nd ed. Port Elizabeth, Nelson Mandela Metropolitan University
Skinner JD, Chimimba CT (2005) The mammals of the southern African subregion. Cambridge University Press, Cape Town
Söndgerath D, Schröder B (2002) Population dynamics and habitat connectivity affecting the spatial spread of populations - a simulation study. Landsc Ecol 17:57–70
South African National Biodiversity Institute (SANBI) (2016) Vegetation Map of South Africa, Lesotho and Swaziland. http://bgis.sanbi.org/SpatialDataset/Detail/18. Accessed 28 March 2017
Stevens VM, Leboulengé É, Wesselingh RA, Baguette M (2006) Quantifying functional connectivity: experimental assessment of boundary permeability for the natterjack toad (Bufo calamita). Oecologia 150:161–171
Taylor P, Fahrig L, Henein K, Merriam G (1993) Connectivity is a vital element of landscape structure. Oikos 68:571–573
Taylor P, Fahrig L, With K (2006) Landscape connectivity: a return to the basics. In: Crooks K, Sanjayan M (ed) connectivity conservation, conservation biology. Cambridge University press pp 29-43
Terborgh J, Lopez L, Nuñez P, Rao M, Shahabuddin G, Orihuela G, Riveros M, Ascanio R, Adler GH, Lambert TD, Balbas L (2001) Ecological meltdown in predator-free forest fragments. Science 294:1923–1926
Tischendorf L, Fahrig L (2000) On the usage and measurement of landscape connectivity. Oikos 90:7–19
Tobler M (2015) Camera Base 1.7. http://www.atrium-biodiversity.org/tools/camerabase. Accessed 4 April 2019
Tobler MW, Carrillo-Percastegui SE, Leite Pitman R, Mares R, Powell G (2008) An evaluation of camera traps for inventorying large- and medium-sized terrestrial rainforest mammals. Anim Conserv 11:169–178
Torres-Romero EJ, Olalla-Tárraga M (2015) Untangling human and environmental effects on geographical gradients of mammal species richness: a global and regional evaluation. J Anim Ecol 84:851–860
Turgeon K, Kramer DL (2012) Compensatory immigration depends on adjacent population size and habitat quality but not on landscape connectivity. J Anim Ecol 81:1161–1170
Turner IM, Corlett RT (1996) The conservation value of small, isolated fragments of lowland tropical rain forest. Trends Ecol Evol 11:330–333
Visconti P, Pressey RL, Giorgini D, Maiorano L, Bakkenes M, Boitani L, Alkemade R, Falcucci A, Chiozza F, Rondinini C (2011) Future hotspots of terrestrial mammal loss. Philos Trans R Soc Lond B Biol Sci 366:2693–2702
Volenec ZM, Dobson AP (2020) Conservation value of small reserves. Conserv Biol 34:66–79
Watson LH, Chadwick P (2007) Management of Cape mountain zebra in the Kammanassie nature reserve, South Africa. S Afr J Wildl Res 37:31–39
Zungu MM, Maseko MST, Kalle R, Ramesh T, Downs CT (2019) Effects of landscape context on mammal richness in the urban forest mosaic of EThekwini municipality, Durban, South Africa. Glob Ecol Conserv 21:e00878
Acknowledgements
The authors wish to thank the City of Cape Town (CCT) Biodiversity Management Branch, for landowner permission and organisational support to conduct this research within the various CCT nature reserves and for the incredibly generous logistics support, use of equipment and invaluable field knowledge of all staff and students who assisted with this research. The authors also wish to thank NCC Environmental Services (Pty) LTD for their generous loan of cameras, and the Friends of Helderberg Nature Reserve and National Research Foundation via Cape Peninsula University of Technology for contributing to funding of equipment. Research methodology was approved through CapeNature permit 0052-AAA041-00019.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
ESM 1
(PDF 642 kb)
Rights and permissions
About this article

Cite this article
Schnetler, A.K., Radloff, F.G.T. & O’Riain, M.J. Medium and large mammal conservation in the City of Cape Town: factors influencing species richness in urban nature reserves. Urban Ecosyst 24, 215–232 (2021). https://doi.org/10.1007/s11252-020-01027-w
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-020-01027-w