
Abstract
Urban natural areas are often heavily invaded by non-native plants. To restore the biodiversity and ecosystem function of these areas, land managers commonly remove invasive species. Partnering with Portland Parks and Recreation, I studied a management-scale removal of Hedera spp., Clematis vitalba, Ilex aquifolium and Prunus laurocerasus from a 59 ha forested natural area in Portland, OR. Over four years, I compared the community composition of this area with contiguous control areas where invasive species remained intact. Hedera, the most abundant invader, was dramatically reduced in removal areas, along with Clematis and Ilex. Both the abundance and the diversity of native herbs and woody seedlings were substantially higher in removal areas, with Claytonia sibirica, Galium spp., Hydrophyllum tenuipes, Trillium ovatum, and Acer macrophyllum seedlings all showing significant increases. By 2015, the minimum native species richness for removal areas was 37, as determined by the Chao 2 estimator, versus 10 native species for control areas. In contrast, native shrubs and ferns showed no increase in response to invasive removal; in fact, control plots typically contained significantly higher stem densities of native shrubs and ferns than removal plots did. Few non-native species recolonized removal areas, except for seedlings of Clematis. Collectively, these results indicate that passive recovery of the herbaceous layer is possible at large scales following invasive species removal, but that active replanting of woody species may be needed. Partnerships between land managers and academic researchers are key to documenting the effects of management-scale removals.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Natural areas provide many benefits for cities and their residents, including the preservation of biodiversity (Sadler et al. 2010). However, fragmented natural areas existing within an urbanized matrix are particularly susceptible to invasions by non-native plants (Duguay et al. 2007; Gavier-Pizarro et al. 2010; Klotz and Kühn 2010; Pennington et al. 2010; Ramalho et al. 2014). In response to the biodiversity losses that can result from these invasions (Andreu and Vilà 2011; Vilá et al. 2011), managers of urban natural areas routinely engage in efforts to control non-native plant invasions chemically and/or mechanically (D’Antonio and Meyerson 2002; Davy 2002; Reid et al. 2009; Kettenring and Adams 2011).
The intent of these efforts is to reduce or eradicate the invasive species and to return the affected area to a more native-rich plant community better approximating its condition before invasion, with improved ecosystem function (D’Antonio and Meyerson 2002). Reaching this goal is complicated by a variety of factors. Depending on the length of the invasion and its intensity, some native plant species may have been reduced in density or even driven locally extinct by interspecific competition with an aggressive non-native invader, leaving no propagule sources (Andreu and Vilà 2011). In addition, the removal efforts themselves may have unintentional negative effects on the native plant community, with the potential for mechanical pulling to uproot small individuals and seedlings, or for herbicide drift to cause mortality of non-target plant species. The invasive species may leave behind a legacy of seeds that may germinate in the newly-opened space created by the removal of the non-native vegetation (D’Antonio and Meyerson 2002; Gioria et al. 2012; Clements and Bierzychudek 2017). And even when there is no persistent seed bank, if neighboring areas are occupied primarily by horticultural species and species typical of disturbed ground, the potential is very high for the newly-created space to be colonized by propagules of the same invasive species that were removed, or by new, even more aggressive invaders (Buckley et al. 2007; Reid et al. 2009; Kettenring and Adams 2011; Cordell et al. 2016).
It is perhaps not surprising, then, that the results of invasive plant removals are often disappointing. In 2011, Kettenring and Adams published a meta-analysis of experimental invasive plant removals. They reported that while invasives were often reduced in these studies, especially when herbicide was used for control, increases in percent cover, biomass, or density of native species were rarely significant, even when active methods of revegetation were employed.
It is hard to know, however, how applicable these results are to management-scale removals. Eighty-eight percent of the studies reviewed by Kettenring and Adams (2011) applied removal treatments to areas ≤1000 m2, while most management-scale removals involve areas many hectares in size (Reid et al. 2009). The results of small-scale removals may not accurately predict the consequences of larger-scale manipulations. For example, Erskine Ogden and Rejmánek (2005) found that native species did not recover as successfully after removal of fennel (Foeniculum vulgare) from 30 m × 30 m blocks as they did in 1 m2 removal plots. In addition, most of the studies reviewed by Kettenring and Adams (2011) monitored results for no more than one growing season. Because recovery from invasion and from the disturbance associated with invasive removal is likely to take years, not months, we can expect short-term monitoring to provide a less optimistic view of recovery than longer monitoring might (Parker and Wiens 2005; Runkle et al. 2007)). For these reasons, we still know too little about the degree of community recovery that can be expected from management-scale invasive species removals, or about what factors influence that recovery.
In the Pacific Northwest (PNW), the escaped ornamental species Hedera hibernica and H. helix are pernicious invaders of urban natural areas (Boersma et al. 2006; Portland Parks and Recreation 2018). The two species are difficult to distinguish morphologically, but most of the invasive PNW populations sampled by Clarke et al. (2006) were found to be H. hibernica. Planted as a fast-growing, hardy groundcover throughout the region, Hedera’s aggressive colonization of natural areas and apparent displacement of native species (Dlugosch 2005) led the Oregon Department of Agriculture to ban its sale in 2010. Currently, efforts by municipalities, land trusts and citizens’ groups are focused on removing Hedera spp. from tree trunks, canopies and forest understories. Several other aggressive non-native plant species are often included in these removal efforts, particularly the non-native woody liana Clematis vitalba and the woody evergreen species Ilex aquifolium and Prunus laurocerasus. However, there have been no reported attempts to document the recovery of natural areas where the removal of these species has occurred.
This knowledge is particularly important for making prudent management decisions regarding urban natural areas. Is removal effective over the long term? Will native species recover passively after invasives are removed? Must land managers also invest in replanting native species? Only a small fraction of the studies reviewed by Kettenring and Adams (2011) followed removals with active replanting, and there were too few such studies to evaluate their effects.
I report here on the aftermath of the removal of Hedera spp. and other invasive woody plant species from a 59-ha urban forest in Portland, Oregon. For four growing seasons after the removal, I compared the plant community in the treated area with that of adjacent reference areas where invasive species remained intact. My goals were to assess the effectiveness of the treatment in reducing the abundance of the invasive species, and to evaluate the degree of passive recovery of the native community, by documenting changes in the species composition and abundance of native species in the treated removal areas relative to the untreated control areas. In particular, I asked these questions:
-
1.
How did treatment affect the abundance of ground and climbing Hedera spp.? How did it affect the abundance of the other treated invasives?
-
2.
Did native species increase in abundance and/or richness in areas where invasives were treated? If so, which species responded, and how much time was required?
-
3.
Did any non-native species increase in abundance or richness in treated areas? If so, which species responded, to what extent, and how quickly?
This study is unusual for several reasons (Kettenring and Adams 2011): the invasive removal occurred on a management-level scale; I observed forest recovery for four years after the removal, documenting how dozens of different plant species responded to the treatment; and I was able to attribute changes in community composition to the removal itself, rather than to other environmental changes, because the study included both a spatial and temporal control. These characteristics highlight the benefits of academic collaboration with land managers, in this case Portland Parks and Recreation. Managers rarely have the time or resources for extensive follow-up studies (Reid et al. 2009), and academic researchers are not typically able to carry out such large-scale removal experiments.
Methods
The River View Natural Area (RVNA), a 59 ha parcel of undeveloped forest, was purchased by the city of Portland, Oregon in 2011. Selectively logged sometime in the first half of the twentieth century, the upland portion of this hillside tract (the site of this study) has a closed canopy consisting primarily of Acer macrophyllum, Pseudotsuga menziesii, Abies grandis, Thuja plicata and Tsuga heterophylla individuals less than 100 years old, with abundant Acer circinnatum, Oemleria cerasiformis, and Corylus cornuta in the understory. This mix represents the most common pre-urbanization plant community in the Portland area (Christy et al. 2009). Common shrubs, ferns, and herbs include Polystichum munitum, Berberis spp., Claytonia sibirica, and Hydrophyllum tenuipes. The area has no formal trail system but contains a network of demand trails created by walkers and mountain bicyclists.
At the time of purchase, RVNA was heavily invaded by non-native Hedera (primarily H. hibernica). The invasion was sufficiently advanced that most of the forest floor was completely covered and many trees supported climbing Hedera stems of 3 cm diameter or more (P. Bierzychudek, personal observation). Hedera had likely been present in RVNA since at least the 1930s (S. Beckham 2012, Lewis & Clark College, Portland, OR, personal communication), when it was used to landscape portions of the adjacent Fir Acres Estate (now the Lewis & Clark College campus). Prunus, Ilex, and (on the periphery) Clematis were also present in RVNA, though at much lower abundances than Hedera. These species were also targeted for removal.
In October–November 2011, ground Hedera in RVNA was sprayed with 4% glyphosate/2% triclopyr 3. Glyphosate is a broad-spectrum herbicide absorbed by plant leaves; treatment occurred in late autumn to reduce potential damage to non-target deciduous herbs and shrubs. All woody stems of Hedera, Clematis, Ilex and Prunus were cut and the cut surfaces treated with 50% triclopyr 3. In September 2012, 2013 and 2014, surviving ground Hedera was spot-treated with glyphosate/triclopyr (T. Query, 2016, City of Portland, OR, personal communication). Each year, crews from Multnomah County Soil and Water Conservation District visited the study area (both control and removal areas) and spot-treated any flowering Alliaria petiolata, which is beginning to invade the region.
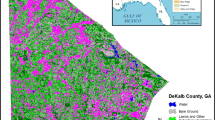
On RVNA’s northern and southern boundaries there are additional parcels of undeveloped forest belonging to River View Cemetery and Lewis & Clark College, respectively. These parcels have a similar mix of dominant species, and an examination of unpublished aerial photographs taken in 1936 indicates that the logging history of these areas was also similar to that of RVNA. These areas were as densely covered by invasive species as RVNA had been, but their invasives were still intact when this study began, allowing them to serve as control areas. To reduce pseudoreplication (Wiens and Parker 1995) and any differences between treated and control areas due to environmental variation, I divided RVNA into two sampling blocks (Fig. 1). Block 1 paired a larger northern portion of RVNA with the control area within River View Cemetery to the north, while block 2 paired a smaller southern region of RVNA with the control area within the Lewis & Clark College campus to the south.

Map of study area. Sampling plots located in areas outlined by ovals
Starting in summer 2012, seven months after the initial treatment occurred, and for three additional growing seasons, I sampled both the treated area and the two matched control areas (constituting a spatial-temporal level-by-time design). In this standard modification of a BACI (before-after control-impact) design (Wiens and Parker 1995), a change in the relative conditions of the treated and control areas over time constitutes evidence of a treatment effect. Because I used uniform sampling protocols over multiple years, this design is robust to the effects of spatial and temporal correlation that often result from pseudoreplication, and does not assume a steady-state environmental equilibrium (Wiens and Parker 1995; Parker and Wiens 2005).
In late 2013, in response to the apparent success of the invasive removal, the owners of the control areas began managing the invasive species on their lands as well. This action complicated the interpretation of subsequent events, but it also added a temporal control to the design. If differences between control and removal areas observed in 2012 and/or 2013 were due to the removal of targeted invasives, then I expected to see parallel changes in former control areas when pre-treatment years were compared to post-treatment years.
The management strategies for the two former control areas differed somewhat. In the block 1 former control area, after the 2013 census, climbing vines of Hedera and Clematis were cut from trees, and trunks of Ilex and Prunus were cut and treated with triclopyr. Ground Hedera was not treated (M. Ahr, 2014, West Multnomah Soil and Water Conservation District, Portland, OR, personal communication). In the block 2 former control area, after the 2013 census, climbing vines of Hedera were cut from trees, and, after the 2014 census, climbing vines of Clematis were cut, trunks of Ilex were girdled, and trunks of Prunus were cut and their stumps treated with triclopyr. In addition, after the 2014 census, ground Hedera in the block 2 former control area was treated with glyphosate/triclopyr, but this treatment took a more cautious approach to herbicide application than that in the original RVNA treated area (A. Dvorak 2015, Lewis & Clark College, Portland, OR, personal communication).
Data collection took place in May–June of 2012–2015, beginning in the first growing season after invasive removal. To compare the species composition of the plant community in the removal areas to that in the control areas, I selected ~200 random points, equally divided between treated and control areas. About 75% of the points fell in the larger block 1. I excluded points that were less than 10 m apart, in riparian areas, on slopes greater than 29°, or less than 5 m from trail edges or property boundaries. I selected a new set of random points each year. Each point served as the center of a circular plot 10 m in diameter (total area 78.5 m2) in which I recorded the numbers of stems and species identity of all ferns, shrubs and trees. Each point also served as the corner of a nested set of three square frequency plots (0.5 m, 1 m, and 2 m on a side, for total areas of 0.25 m2, 1 m2, and 4 m2) in which I recorded the incidence (presence/absence) and the identity of all herbaceous species, ground vines, and woody seedlings <10 cm tall. I ignored graminoids because of the impossibility of accurately identifying nonflowering individuals. Incidence is measured far more consistently among observers and years than percent cover; this consistency is an important assumption of the spatial-temporal level-by-time design (Wiens and Parker 1995). Using three sizes of frequency plots allowed me to track changes in the incidence of abundant and rare species with similar accuracy (Elzinga et al. 2001). My authority for species identifications was the Oregon Flora Project (www.oregonflora.org/index.php). A few nonflowering taxa could not be identified to the species level; all Galium species were pooled, and Tellima/Tolmeia were pooled.
In February of 2014 and 2015, before the study was completed, selected areas of RVNA were replanted with bare-root shrubs and coniferous tree saplings by the Multnomah County Soil and Water Conservation District. Because of the uniform sizes of the transplants and the fact that the shrubs were always planted in clusters of three or five stems, it was easy to distinguish these transplants from naturally-occurring individuals. Here I report data for the naturally-occurring individuals only.
For species recorded in the large circular plots, I present two measures of abundance: density (the number of stems per plot) and frequency (the proportion of plots in which a species is present). Because the densities of most species were non-normal, I report medians rather than means. For species recorded in the smaller nested frequency plots, I used a species’ frequency as a measure of its abundance, and analyzed frequency data from the plot size appropriate for each species’ abundance (Elzinga et al. 2001). For Hedera, which reached very high frequencies, I used data from the 0.25 m2 plots, and for all other species, I used data from the 4 m2 plots.
I performed all analyses in R (R Core Team 2014). Because the control areas of the two blocks were managed somewhat differently, I analyzed data from each block separately. To analyze frequency data, I used glm (family = binomial) to test specific hypotheses with planned contrasts. For data from block 1, where climbing vines of Hedera and Clematis were cut from trees in the former control area after the 2013 census, and trunks of Ilex and Prunus were cut and treated with triclopyr, I created planned contrasts to answer these questions:
-
1.
Was there a response to the original treatment? H0 = (control area in 2012–2015) = (treated area in 2012–2015).
-
2.
Was there a response to the treatment of the former control area? H0 = (control area in 2012–3) = (control area in 2014–2015).
-
3.
If there was such a response, could it have been due to a more general temporal change rather than to the treatment? H0 = (treated area in 2012–3) = (treated area in 2014–2015)
For data from block 2, where climbing vines of Hedera were cut from trees in the former control area after the 2013 census, and climbing vines of Clematis, trunks of Ilex and Prunus, and ground Hedera were treated after the 2014 census, I created planned contrasts to answer these questions:
-
1.
Was there a response to the original treatment? H0 = (control area in 2012–2014) = (treated area in 2012–2014)
-
2.
Was there a response to the treatment of climbing Hedera in the former control area? H0 = (control area in 2012) = (control area in 2013–2014)
-
3.
If there was such a response, could it have been due to a more general temporal change rather than to the treatment? H0 = (treated area in 2012) = (treated area in 2013–2014)
-
4.
Was there a response to the treatment of the other invasive species in the former control area? H0 = (control area in 2012–2014) = (control area in 2015)
-
5.
If there was such a response, could it have been due to a more general temporal change rather than to the treatment? H0 = (treated area in 2012–2014) = (treated area in 2015)
When the planned contrasts were orthogonal, I combined them into a single glm; otherwise I analyzed them individually. To analyze density data, I used the same planned contrasts, but with anova rather than glm. In the sections that follow, I refer to a response to the original treatment in either block as a “spatial treatment effect” and a response to the treatment of former control areas as a “temporal treatment effect.”
I created sample-based rarefaction curves (Gotelli and Colwell 2001, 2011) using the R package “vegan” (Oksanen et al. 2015) to compare how observed species richness of native herbs and woody seedlings in control and removal areas changed between 2012 and 2015. I used the Chao2 estimator (Gotelli and Colwell 2011), which adjusts for the number of species that were present but undetected, to estimate the minimum species richness for each treatment-year combination.
I visualized differences in post-treatment species composition between control and treated plots with nonmetric multidimensional scaling (NMDS), using the R package “vegan.” I used incidence data from the 2015 censuses for native and non-native herbs, woody seedlings, ferns and shrubs. I excluded treated invasives from this analysis to prevent their dramatic difference in frequency between control and removal plots from obscuring the pattern of recovery. I also excluded any species that occurred in fewer than two plots, and any plots that contained fewer than two species. Three dimensions were needed to obtain a convergent solution with a stress level below 0.2. I used the function ‘adonis’ to partition variance among treatments.
Results
Effectiveness of ivy removal
Treatment of Hedera in 2011 produced dramatic changes in the overall appearance of RVNA within a few weeks (P. Bierzychudek, personal observation); even casual visitors remarked on the disappearance of the Hedera carpet. At the time of the 2012 census, ground Hedera was present in 94.8% of the 0.25 m2 control plots, but only 45.2% of removal plots (Fig. 2). By 2013, after one year of re-treatment, Hedera’s frequency in the removal plots had dropped below 20%, while remaining high in control plots (Fig. 2). The difference between control and removal areas was even more dramatic than these data suggest, because even a single Hedera stem was sufficient to count as an occurrence. While control plots usually had a thick tangle of multiple stems, removal plots rarely contained more than one or two small stems/plot (P. Bierzychudek, personal observation). In both blocks, there was a significant spatial treatment effect (glm, p < 0.0001); this difference between control and removal plots was maintained over time. Treatment of former control areas did not produce a temporal treatment effect (glm, p = 0.28). This is not surprising, since in block 1’s former control area, ground Hedera was not treated at all, and in the control area of block 2, treatment of ground Hedera was highly conservative, producing only a modest decrease in its frequency (Fig. 2).

Mean proportion (± 1 s.e.) of control and removal plots containing treated invasive species, by species and census year. Solid circles = control plots; open circles = removal plots. Removal areas were treated 7 months prior to the 2012 census. Before the 2014 census, climbing Clematis and Hedera (but not ground Hedera), I. aquifolium, and P. laurocerasus were treated in the former control area of block 1. Before the 2015 census, these species plus ground Hedera were treated in the former control area of block 2
Cutting of Hedera trunks from trees also produced visible changes in the canopy and in the amount of light penetrating the forest. In 2012, climbing Hedera stems were present in 69.8% of the plots in control areas, but were completely absent from removal areas, creating a significant spatial treatment effect (glm, p < 0.0001). Similar differences persisted in 2013. Once climbing Hedera in former control areas was treated, it was absent from all sample plots there as well (Fig. 2), leading to a significant temporal treatment effect in both blocks (glm, p < 0.0001).
Effectiveness of removal of other invasive species
The other invasive species – Clematis, Ilex, and Prunus – were less abundant than Hedera prior to treatment, and experienced less dramatic reductions. Climbing Clematis, restricted to the sunny periphery of the forest, occurred in 11.1% of block 1’s control plots in 2012, compared to 1.5% of removal plots (Fig. 2). This pattern persisted in 2013, leading to a spatial treatment effect in block 1 (glm, p < 0.0001). Because climbing Clematis was less abundant in block 2 (4.2% of control plots), its removal created no significant spatial treatment effect (glm, p = 0.489). Treatment of climbing Clematis in the former control areas led to a significant temporal treatment effect in block 1 (Fig. 2, glm, p < 0.0001), but not in block 2 (glm, p = 0.244).
In 2012 and 2013, Ilex was present in 30% of the control plots, compared to 8% of removal plots, leading to a significant spatial treatment effect in both blocks (glm, p < 0.05). Treatment of the former control area in block 1 was followed by a reduction in Ilex frequency there to levels similar to that in the original treatment area (Fig. 2; Table 2), leading to a significant temporal treatment effect (glm, p < 0.0001); in block 2, where Ilex was infrequent in the control areas, there was no temporal treatment effect.
Prunus responded differently to treatment than the other invasive species did (Fig. 2). After being treated in 2012, its frequency was similar in control and removal plots of block 1 (12.5% vs. 14.7%, Fig. 2). Once cut, large trunks of laurel were typically replaced by numerous smaller stems resprouting from the roots (personal observation). In block 2, however, treated areas had a significantly lower incidence of Prunus than controls, 16.7% vs. 8%, a significant spatial treatment effect (glm, p < 0.05).
Responses of native herbs/woody seedlings: Changes in native species frequency and richness
Concomitant with observed differences in occurrence of invasive species between control and removal areas, there were substantial differences between treatments in the abundance and diversity of native herbs and native woody seedlings in the understory. This increase was apparent at both the individual plot level and at the whole forest level.
Compared to control plots, removal plots consistently contained significantly greater numbers of native species of herbs and of woody seedlings (Fig. 3). Even in 2012, only seven months after initial treatment, removal plots already contained more native species than control plots, and the size of that difference increased over time. There was a significant spatial treatment effect for species richness in both blocks (anova, p < 0.005). Native species richness in the treatment areas increased significantly over time (anova, p < 0.01). There was no temporal treatment effect for species richness, likely because ground Hedera did not significantly decrease when former control areas were treated.

Mean (± 1 s.e.) species richness of native herbs and woody seedlings per plot, by treatment, year and block. Solid circles = control plots; open circles = removal plots. N for block 1 = 68–75, depending on year and treatment; N for block 2 = 22–30, depending on year and treatment
Claytonia, a fall-germinating annual herb, showed one of the strongest responses to invasive removal. It was absent from the control plots at the 2012 census, but present in 14% of removal plots, and often reached higher frequencies thereafter (Table 1). There was a significant spatial treatment effect for Claytonia in block 2 (glm, p < 0.01) Other native species showing a significant spatial treatment effect (glm, p < 0.05) in one or both blocks were Galium spp., Hydrophyllum, Trillium ovatum, and seedlings of A. macrophyllum, Table 1). Few of these species showed a temporal treatment effect.
At the whole forest level, the rarefaction analysis demonstrated that, compared to the two control areas, the two removal areas contained many more species of native herbs and woody seedlings. Even in the first growing season after treatment, I observed twice as many species of native herbs and woody seedlings in the removal areas than in the control areas (14 vs. 7 species, Fig. 4a). By 2015, after the original control areas had been treated, their species richness had increased from seven to ten species, but the native species richness of the original treatment areas had risen even higher, to 21 (Fig. 4b). Several unusual species were found in the removal plots that were never observed in the control plots, e.g. Monotropa uniflora and Cephalanthera austiniae. These and many other native species in the removal areas were represented in only one or two plots. The existence of so many singletons implies that additional sampling would detect even more species. The Chao2 estimate of minimum species richness for the control areas was 10 ± 0, which had been reached by 2015, whereas the removal areas were estimated to contain 37 ± 16.5 species; this estimate was much greater than the number of observed species (Fig. 4).

Rarefaction curves for native herbs and native woody seedlings in control areas (dashed line, darker confidence interval cloud) and removal areas (solid line, lighter cloud), in 2012 (4a), a few months after treatment, and in 2015 (4b). Confidence interval clouds represent 2 se
Responses of native ferns and shrubs
In contrast to the responses of native herbs and woody seedlings, native ferns and shrubs showed no evidence of increasing in response to invasive removal. In fact, compared to control plots, removal plots typically contained only half as many stems of native shrubs (all species pooled, Fig. 5). This led to a significant spatial treatment effect, but in the opposite direction of the pattern that would indicate recovery (F test, p < < 0.001). Oemleria, one of the most abundant of the native shrubs, displayed this pattern, as did the less-abundant Rubus parviflorus, Vaccinium parvifolium, Berberis spp., Rosa sp., Disporum hookeri and Gaultheria shallon (Table 2; F test for effect of spatial treatment, p always <0.01). No shrub species were significantly denser in removal plots than in control plots.

Mean (± 1 s.e.) density of native shrub stems per plot, by treatment, year and block. Solid circles = control plots; open circles = removal plots. N for block 1 = 68–75, depending on year and treatment; N for block 2 = 22–30, depending on year and treatment. Before the 2015 census in block 2, ground Hedera in former control area was treated with glyphosate
Ferns showed a similar pattern, but to a lesser degree. Fern stems (all species pooled) were approximately half as dense in removal plots as they were in control plots in block 1. Densities did not differ between treatments in block 2 (Fig. 6). Treatment x block and treatment x year interactions explained a significant amount of the variation in total fern densities among plots (F tests, p < 0.02). Individual analyses of the two most abundant ferns, P. munitum and Pteridium aquilinum, supported this pattern. Athyrium felix-femina showed a more straightforward significant effect of spatial treatment (p < 0.05), with significantly fewer stems in removal plots than in control plots (Table 2).

Mean (± 1 s.e.) density of native fern stems per plot, by treatment, year and block. Solid circles = control plots; open circles = removal plots. N for block 1 = 68–75, depending on year and treatment; N for block 2 = 22–30, depending on year and treatment. Before the 2015 census in block 2, ground Hedera in former control area was treated with glyphosate
Responses of non-native herbs/woody seedlings
The median number of non-native species (including Hedera and Clematis) in removal plots was always 0, even four years after the initial treatment. In some blocks and years, removal plots contained somewhat higher frequencies of Alliaria, seedling Hedera, Lapsana communis, Mycelis muralis, Ranunculus repens, and Oxalis corniculata than control plots did, but these species’ frequencies were typically low (Table 1), and these differences were never significant (χ2 tests, p always >0.05). There is no evidence that their frequencies have increased over time (Table 1).
The most prevalent non-native colonist by far was Clematis. By 2015, seedling Clematis were significantly more frequent in 4 m2 removal plots than in control plots (Table 1, glm, p < 0.0001). They also showed a significant temporal treatment effect in block 2, where ground Hedera was treated (Table 1, glm, p < 0.0001).
Overall plant community responses
By 2015, control and removal plots were quite distinct in species composition (Fig. 7, stress level of the analysis = 0.19). Compared to control plots, where ground and climbing Hedera, climbing Clematis, Ilex and Prunus were abundant, along with Rubus armeniacus, removal plots had high incidences of native herbs (esp. Claytonia, Galium spp., Hydrophyllum, and Tolmeia/Tellima) and of seedlings of native trees (esp. A. macrophyllum). Removal plots also tended to have relatively high frequencies of Clematis seedlings. Treatment explained a highly significant amount of the variance in species composition among plots (Adonis test, p < 0.001). A portion of this difference is likely due to slightly but significantly greater variability in species composition of the control plots (Fig. 7, ANOVA, p = 0.026).

Non-metric multidimensional scaling plot for incidence data for native and non-native herbs, woody seedlings, ferns and shrubs (excluding treated invasive species) in 2015. Control plots are filled circles; removal plots are open circles. Blocks are pooled to facilitate comparison of years and treatments. Vectors indicate the species whose incidences were strongly correlated (p < .01) with the ordination. The lengths of the vectors are scaled to the r2 value for each species. Non-native species vectors are shown in red. The first two dimensions of a three-dimensional analysis are displayed; the stress value of the analysis is 0.19
Discussion
Treatment with glyphosate led to significant reductions in the frequency of Hedera spp. and of two of the other woody invasives: Clematis vitalba and Ilex aquifolium. Treatment was ineffective at reducing the incidence of Prunus laurocerasus because of this species’ tendency to resprout vigorously from cut stumps. These reductions, particularly of ground Hedera, created opportunities for native herbs and tree seedlings to re-colonize the treated parts of the forest. Several species of native herbs became more abundant within less than a year, with additional species recruiting in subsequent seasons. By 2016, the treated parts of the forest were three times as species-rich as the untreated parts. In addition, the restriction of many species to just a single sample plot implies that there are even more species in the treated area that were not detected in our sampling.
The speed with which herbaceous species appeared in removal areas is likely due to their presence as buried seeds. Seeds of Claytonia and Tolmeia/Telllima are abundant in soil sampled from these forests (Clements and Bierzychudek 2017); herbaceous species, especially annuals and biennials, often produce seeds that can remain viable in the seed bank for many years (Baskin and Baskin 1998). While most tree species do not form persistent seed banks (Pickett and McDonnell 1989), there are mature trees in the forest canopy, producing new propagules annually, that germinate if microsite conditions are appropriate. Interestingly, there have been reports of limited conifer seedling recruitment in urban forests in Portland and Seattle (Broshot 2011; O’Brien et al. 2012; Ettinger et al. 2017), most of which are invaded by Hedera. While I found that removal of invasive Hedera increased the abundance of tree seedlings, Ettinger et al.’s (2017) experimental study found that seed and dead wood availability were more important constraints on conifer seedling germination and establishment than the presence of ground Hedera.
In contrast to herbs and tree seedlings, I saw no increase in fern and shrub abundance in removal areas. Fern spores are typically aerially dispersed (Kato 1993), but fern gametophytes are inconspicuous and slow-growing, and may have been missed in the surveys. Plants of stable habitats like forests do not typically form persistent seed banks (Thompson et al. 1998); Clements and Bierzychudek (2017) found only Rubus and Sambucus in soil samples. Thus shrub recruitment into removal areas must rely principally on seed dispersal. Most of the shrub species common in this forest produce fleshy fruits that are likely bird-dispersed (e.g. Berberis, Gaultheria, Oemleria, Vaccinium). However, existing individuals rarely flower or set fruit in the dark understory of the forest; most of the reproduction takes place on the sunnier forest edges or in light gaps. Thus, shrub propagules are not abundant and must be carried by dispersers into the forest interior. More time may be required before these species recruit into the removal areas.
Vegetative growth of woody shrubs might be expected to increase in response to a possible reduction in competition from treated invasive species. However, in contrast to the vigorous response of native herbs and tree seedlings, woody shrub stems were not denser in removal areas. In fact, stem densities of native shrubs were significantly lower in removal areas than in control areas. These lowered densities could be evidence of damage to non-target perennial species by glyphosate. However, we did not observe significant numbers of dead shrub stems, as might have been the case if this had occurred. It is also possible that control and removal areas had different densities of shrub stems before the treatment was implemented. We had hoped to investigate this hypothesis by looking at whether shrub densities in block 2’s control area decreased after it was treated. While there was a (non-significant) decrease in shrub densities in the former control area of block 2 in 2015, a decrease of similar magnitude also took place in the control area of block 1, where ground Hedera was not treated, suggesting that the decrease was not a response to the herbicide. In addition, there were differences in the management approaches by the two crews (the block 2 crew in 2014 was far more conservative in their herbicide application than the original removal crew was) that complicated this comparison. The lower density of ferns in removal areas is easier to attribute to herbicide drift. The most common fern species by far was Polystichum, and it is one of the few plant species that can be seen emerging among thick carpets of ground Hedera. It is hard to imagine how Hedera could be sprayed without affecting Polystichum, and the fronds of many Polystichum individuals in removal areas were sparse and misshapen.
We also observed an increase in the frequency of non-native herbs and seedlings in removal areas. However, in contrast to studies in which the treated invasives quickly returned to pre-treatment levels (e.g. Mattingly et al. 2016), the frequency of non-native species was quite low, and consisted primarily of species typical of disturbed ground that require relatively high light levels. For this reason, it seems unlikely that these species will increase to problematic levels. Seedlings of Hedera, Ilex, and Prunus were quite rare. Even Clematis, the most abundant non-native recruit, is likely to require more light than is typically available in the forest understory, except for light gaps. However, over time, birds might be expected to carry the fleshy seeds of Hedera, Ilex, and Prunus into the forest. Maintaining these species at low densities will therefore require constant monitoring.
Reid et al. (2009) made a plea for researchers and land managers to work together more often in weed management programs. This project represents a fruitful collaboration between academic science and Portland Parks and Recreation (PPR), a land management agency. The collaboration provided access to an experimental manipulation at an unusually large scale that would have been difficult to achieve without PPR support. The results provided the agency with a long-term, quantitative assessment of how the plant community was affected by their management actions. This level of assessment is normally not possible given agency priorities and constraints, and is not typical for weed management programs generally (Reid et al. 2009).
At the same time, agency interests were sometimes at odds with research goals. In particular, this study of passive removal was complicated by PPR’s decision to replant bare-root shrubs and trees while the study was in progress. In hindsight, replanting was in fact an important strategy to implement, given the reduced density of shrub stems and ferns in treated areas, the low numbers of fruiting individuals, and the paucity of shrub seeds in the seed bank (Clements and Bierzychudek 2017).
According to Parker and Wiens (2005), “recovery occurs when the state of the system after the impact matches its state before the impact or that of a similar, unimpacted reference area.” In many urban areas, there are no suitable reference areas remaining, so judging the extent of recovery is difficult. While the plant community in the removal area of this forest is now much more diverse than that in the control area, including some uncommon species such as the mycoheterotrophs M. uniflora and C. austiniae, herb abundance and diversity are still quite low relative to those of nonurban sites. Many herbaceous species typical of nonurban sites are rare or absent in the study area. The absence of these species may be a consequence of many factors, including the fragmentary nature of urban forests and the urban heat island effect (Gaston et al. 2010). Indeed, the loss of these species may even have preceded the non-native invasion, and paved the way for it to take place (Davis et al. 2000).
However, full recovery of native species is not the only benchmark of success of a restoration project. If reduction of non-native species improves the forest’s functioning, for example either as wildlife habitat or in its capacity for water quality protection, then restoration should also be judged successful. This study, like most others (Reid et al. 2009; Kettenring and Adams 2011) did not have the resources to pursue this important question.
References
Andreu J, Vilà M (2011) Native plant community response to alien plant invasion and removal. Managing Biological Invasions 2:81–90
Baskin CC, Baskin JM (1998) Seeds: ecology, biogeography, and evolution of dormancy and germination. Academic Press, San Diego
Boersma PD, Reichard SH, Van Buren AN (eds) (2006) Invasive species in the Pacific Northwest. University of Washington Press, Seattle
Broshot NE (2011) Mortality and recruitment in an urban forest (Forest Park in Portland, Oregon) between 1993 and 2003. Urban Ecosyst 14:553–567
Buckley YM, Bolker BM, Rees M (2007) Disturbance, invasion and re-invasion: managing the weed-shaped hole in disturbed ecosystems. Ecol Lett 10:809–817
Christy JA, Kimpo A, Marttala V, Gaddis PK, Christy NL (2009) Urbanizing flora of Portland, Oregon, 1806-2008. Native Plant Society of Oregon Occasional Paper 3:1–319
Clarke MM, Reichard SH, Hamilton CW (2006) Prevalence of different horticultural taxa of ivy (Hedera spp., Araliaceae) in invading populations. Biol Invasions 8:149–157
Clements H, Bierzychudek P (2017) Can the persistent seed bank contribute to the passive restoration of urban forest fragments after invasive species removal? Ecol Restor 35:156–166
Cordell S, Ostertag R, Michaud J, Warman L (2016) Quandaries of a decade-long restoration experiment trying to reduce invasive species: beat them, join them, give up, or start over? Restor Ecol 24:139–144
D’Antonio C, Meyerson LA (2002) Exotic plant species as problems and solutions in ecological restoration: a synthesis. Restor Ecol 10:703–713
Davis MA, Grime JP, Thompson K (2000) Fluctuating resources in plant communities: a general theory of invasibility. J Ecol 88:528–534
Davy AJ (2002) Establishment and manipulation of plant populations and communities in terrestrial systems. In: Perrow MR, Davy AJ (eds) Handbook of ecological restoration, Volume 1. Cambridge University Press, Cambridge, pp 223–241
Dlugosch KM (2005) Understory community changes associated with English ivy invasions in Seattle’s urban parks. Northwest Science 79:52–59
Duguay S, Eigenbrod F, Fahrig L (2007) Effects of surrounding urbanization on non-native flora in small forest patches. Landsc Ecol 22:589–599
Elzinga CL, Salzer DW, Willoughby JW, Gibbs JP (2001) Monitoring plant and animal populations. Blackwell Science, Malden, Massachusetts
Erskine Ogden JA, Rejmánek M (2005) Recovery of native plant communities after control of a dominant invasive species, Foeniculum vulgare: implications for management. Biol Conserv 125:427–439
Ettinger AK, Lee BR, Montgometry S (2017) Seed limitation and lack of downed wood, not invasive species, threaten conifer regeneration in an urban forest. Urban Ecosyst 20:877–887
Gaston KJ, Davies ZG, Edmondson JL (2010) Urban environments and ecosystem functions. In: Gaston KJ (ed) Urban ecology. Cambridge University Press, Cambridge, pp 35–52
Gavier-Pizarro GI, Radeloff VC, Stewart SI, Huebner CD, Keuler NS (2010) Housing is positively associated with invasive exotic plant species richness in New England, USA. Ecol Appl 20:1913–1925
Gioria M, Pyšek P, Moravcová L (2012) Soil seed banks in plant invasions: promoting species invasiveness and long-term impact on plant community dynamics. Preslia 84:327–350
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett 4:379–391
Gotelli NJ, Colwell RK (2011) Estimating species richness. In: Magurran AE, McGill BJ (eds) Biological diversity. Frontiers in Measurement and Assessment. Oxford U. Press, Oxford, pp 39–54
Kato M (1993) Biogeography of ferns: dispersal and vicariance. J Biogeogr 20:265–274
Kettenring KM, Adams CR (2011) Lessons learned from invasive plant removal experiments: a systematic review and meta-analysis. J Appl Ecol 48:970–979
Klotz S, Kühn I (2010) Urbanisation and alien invasion. In: Gaston KJ (ed) Urban ecology. Cambridge University Press, Cambridge, pp 120–133
Mattingly KZ, McEwan RW, Paratley RD, Bray SR, Lempke JR, Arthur MA (2016) Recovery of forest floor diversity after removal of the nonnative, invasive plant Euonymus fortunei. Journal of the Torrey Botanical Society 143:103–116
O’Brien AM, Ettinger AK, HilleRisLambers J (2012) Conifer growth and reproduction in urban forest fragments: predictors of future responses to global change? Urban Ecosyst 15:879–891
Oksanen J, Guillaume Blanchet F, Kindt R, Legendre P, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2015) Vegan: community ecology package. R package version 2:2–1 http://CRAN.R-project.org/package=vegan
Parker KR, Wiens JA (2005) Assessing recovery following environmental accidents: environmental variation, ecological assumptions, and strategies. Ecol Appl 15:2037–2051
Pennington DN, Hansel JR, Gorchov DL (2010) Urbanization and riparian forest communities: diversity, composition, and structure within a metropolitan landscape. Biol Conserv 143:182–194
Pickett STA, McDonnell MJ (1989) Seed bank dynamics in temperate deciduous forest. In: Leck MA, Parker VT, Simpson RL (eds) Ecology of soil seed banks. Academic Press, San Diego, pp 123–147
Portland Parks and Recreation (2018). The Ivy Files. https://www.portlandoregon.gov/parks/article/201781 - No Ivy League Project Statistics Accessed 11 July 2018
R Core Team (2014) R: a language and environment for statistical computing. R foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/
Ramalho CE, Laliberté E, Poot P, Hobbs RJ (2014) Complex effects of fragmentation on remnant woodland plant communities of a rapidly urbanizing biodiversity hotspot. Ecology 95:2466–2478
Reid AM, Morin L, Downey PO, French K, Virtue JG (2009) Does invasive plant management aid the restoration of natural ecosystems? Biol Conserv 142:2342–2349
Runkle JR, DiSalvo A, Graham-Gibson Y, Dorning M (2007) Vegetation release eight years after removal of Lonicera maackii in west-Central Ohio. Ohio J Sci 107:125–127
Sadler J, Bates A, Hale J, James P (2010) Bringing cities alive: the importance of urban green spaces for people and biodiversity. In: Gaston KJ (ed) Urban ecology. Cambridge University Press, Cambridge, pp 230–260
Thompson K, Bakker JP, Bekker RM, Hodgson JG (1998) Ecological correlates of seed persistence in soil in the north-west European flora. J Ecol 86:163–169
Vilá M, Espinar JL, Hejda M, Hulme PE, Jaroŝík V, Maron JL, Pergl J, Schaffner U, Sun Y, Pyŝek P (2011) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities, and ecosystems. Ecol Lett 14:702–708
Wiens JA, Parker KR (1995) Analyzing the effects of accidental environmental impacts: approaches and assumptions. Ecol Appl 5:1069–1083
Acknowledgments
P. G. Kennedy provided the original idea for the study, helped develop the sampling design (along with L. Bogar), and provided valuable advice on the analysis and manuscript. Many Lewis & Clark students helped census study plots: S. Blizzard, L. Bogar, H. Clements, V. Engebretson, J. Hull, J. Nelson, F. Novak, S. Reed, K. Von Burg, and especially M. Garfias. J. Nelson developed the GIS routines. K. Peterson-Morgan from Portland Parks and Recreation, A. Dvorak from Lewis & Clark College, and D. Noble at River View Cemetery facilitated access to study sites. Financial support was provided by Lewis & Clark College’s J. S. Rogers Fund and the J. F. and M. L. Miller Foundation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Bierzychudek, P. Passive recovery of an urban forest in the Pacific Northwest after removal of invasive plants. Urban Ecosyst 23, 1023–1038 (2020). https://doi.org/10.1007/s11252-020-00966-8
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-020-00966-8