
Abstract
Anthropogenic alteration of the landscape has facilitated plant community change and non-native species invasion in urban areas. We used species occurrence data from over 2000 wetlands within the Chicago metropolitan region to classify urban wetlands into community types and examined non-native species composition across community types. Non-native species were widespread across the region, occurring in over 99% of wetlands. On average, 35% of the plant species in individual wetlands were non-native. A single non-native species, Phalaris arundinacea, was present in 74% of wetlands. Six wetland community types were identified (wet meadows, marshes, forested wetlands, farmed wetlands/mudflats, roadside marshes, and an undetermined wetland type), with each having aggressively spreading non-natives amongst the most common plant species. We conducted canonical correspondence analysis to evaluate the contribution of surrounding land cover, roads, and location of wetlands to plant community composition in these wetlands, and found that similar changes to the landscape have resulted in similar combinations of native and non-native species. Differences in species composition reflected spatial gradients in land use from urban to rural areas across the region, as well as proximity to major roads. Anthropogenic drivers have resulted in profound and pervasive changes to wetland communities across the region, creating novel habitats and ultimately novel community types.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Factors associated with urbanization are major drivers of plant community assembly in urbanized regions. Urbanization is associated with changes in the local environment via reduction of habitat area, elimination of unique community types, introduction of non-native species, eutrophication of wetlands, and alteration of hydrology (Reinelt et al. 1998; McKinney 2002, 2006; Schwartz et al. 2006; Hogan and Walbridge 2007). These changes in the local environment have resulted in increased susceptibility to invasion by non-native plant species (Alpert et al. 2000; McKinney 2002). As a result, the presence and extent of urban land cover around a site are reliable indicators of non-native plant species presence and abundance (Aronson et al. 2014; González-Moreno et al. 2014; Thomas and Moloney 2015). Once a population of non-native species establishes in a site, propagules can be dispersed to nearby sites (Hutchinson and Vankat 1997; Foxcroft et al. 2004), increasing the likelihood that sites in close proximity will share the same group of non-native species and increasing overall community similarity.
In urban areas, the proximity and density of roads are also reliable indicators of community composition (Gavier-Pizarro et al. 2010). Road construction results in the removal of native vegetation, soil disturbance, and re-vegetation of roadsides with both native and non-native species (Forman and Alexander 1998; Forman et al. 2003; Gelbard and Belnap 2003). Roadside maintenance, including mowing and herbicide application, as well as de-icing salt run-off, can negatively affect native vegetation (Forman and Alexander 1998; Forman et al. 2003; Hansen and Clevenger 2005). Together, the construction and on-going maintenance of roads facilitate the establishment and spread of non-native plant species by providing disturbed habitats and colonization sites (Gelbard and Belnap 2003; Kalwij et al. 2008; Joly et al. 2011; Meunier and Lavoie 2012). Furthermore, linear landscape features like roadside habitats can provide corridors that link patches through seed transport (Maheu-Giroux and de Blois 2006; Thiele et al. 2008; Brisson et al. 2010). Passing vehicles, particularly vehicles moving at highway speeds, create air currents that blow seeds or plant material along roadways (von der Lippe and Kowarik 2007, 2008; von der Lippe et al. 2013). Vehicles can also directly carry attached seeds or plant material (Hodkinson and Thompson 1997; Hansen and Clevenger 2005). Connections between patches increase the likelihood that seeds will be dispersed and that non-native populations will establish (Thiele et al. 2008; Minor et al. 2009; Minor and Gardner 2011; Vilà and Ibáñez 2011).
Wetlands, due to their position as landscape sinks, are subject to inputs of both non-native propagules and pollutant- and nutrient-laden runoff (Zedler and Kercher 2004). As a consequence, anthropogenically disturbed wetlands are often dominated by invasive plants such as Phalaris arundinacea, Phragmites australis, Lythrum salicaria, and Typha x glauca (Galatowitsch et al. 1999). However, different forms of anthropogenic disturbance, combined with different underlying environmental conditions among wetland types, can result in divergent communities of non-native species (Choi and Bury 2003). Numerous classification systems have been developed for wetlands based on characteristics of vegetation, hydrology, water chemistry, substrate, and landform (Tiner 2017), but with the notable exception of Cowardin et al. (1979), most wetland classifications do not explicitly consider human alterations to wetlands. Native wetland types in our study region (Chicago, Illinois, USA) included forested wetlands in floodplains or upland depressions, wet prairies, sedge meadows, marshes, and fens (Mohlenbrock 2002; Wilhelm and Rericha 2017). However, wetlands in the region have undergone extensive excavation, impoundment, draining and filling, and altered water and sediment chemistry, leading to plant communities unlike those present at the time of European settlement (Choi and Bury 2003; Wilhelm and Rericha 2017).
The creation of new habitat conditions through urbanization, and the consequent increased abundance of non-native plant species, suggests that plant communities of urban ecosystems may be profoundly different from those in native ecosystems. The term “novel community” has been used to describe a variety of conditions including new assemblages of species originating from different locations, communities with no natural analog, and changes in community composition in response to climate or atmospheric changes (Hobbs et al. 2006; Seastedt et al. 2008; Truitt et al. 2015). Similar anthropogenic changes to the local environment may result in similar non-native-dominated plant communities throughout an urbanizing region, suggesting the formation of coherent novel communities. The objectives of this study were to: (1) classify wetland communities in the Chicago metropolitan region (2) characterize the non-native components of these communities, and (3) relate anthropogenic alterations of the local environment and landscape configuration to patterns of species composition.
Methods
Study area
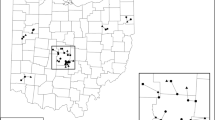
This study was conducted within the Chicago metropolitan region in northeastern Illinois, USA, an area that includes Cook, DuPage, Kane, Lake, McHenry and Will Counties (approximately 9708 km2) (Fig. 1). We selected this area because it includes a gradient of urban-to-rural land uses and the region is heavily impacted by invasive species (Early Detection and Distribution Mapping System 2015). The climate of the Chicago metropolitan region is continental with an average annual temperature of 9.94 C, an average annual precipitation of 93.7 cm (including both rain and snow water equivalent), and an average annual snowfall depth of 93.2 cm (Angel 2011).

Study location map showing the location of sampled wetlands
INHS-IDOT dataset
Wetlands used in this study were investigated and mapped by the Illinois Natural History Survey (INHS) in advance of Illinois Department of Transportation (IDOT) construction projects, following U.S. Army Corps of Engineers methodology. To be considered a wetland, an area must possess hydrophytic vegetation, indicators of wetland hydrology, and hydric soils (Environmental Laboratory 1987; U.S. Army Corps of Engineers 2010). The size of the wetlands ranged from 0.001 to 69.120 ha within the INHS survey limits. Survey limits were determined by the project requirements of IDOT, and as a result not all wetland boundaries were mapped in their entirety. INHS botanists compiled a list of all plant species observed (presence/absence only) in each wetland.
We considered all wetlands mapped by INHS within the study area between 2002 and 2013, for inclusion in this study. Although some changes may have occurred in the regional flora during this time span, we opted to maximize the number of sampling locations and the total area sampled by including data from the full 12-years period. We included only wetlands investigated between April and October because wetlands investigated outside the growing season were likely to have incomplete species lists. Due to the original purposes of the data collection, spatial overlaps existed among some sampled areas. Where spatial overlaps occurred, we selected the most recent investigation. The resulting sample contained 2005 wetlands (Fig. 1).
Predictor variables
Predictor variables included measures of local environmental factors and landscape configuration. All variables were measured using ArcGIS 10.2.2 (ESRI 2014) or R with packages raster and rgdal (Bivand et al. 2014; Hijmans 2014; R Core Team 2014). All GIS datasets were projected to planar coordinates, and all distances are planar measurements. We used the below listed predictor variables.
-
1.
Land Cover: We described the land cover/use of the area surrounding each wetland’s centroid. Wetland centroid points were buffered at 100 m and 1 km, representing the local and neighborhood contexts, respectively. Proportions of eight land cover types (agriculture, developed, forest, shrubland, grassland, wetland, water, and barren ground) present within the buffers were calculated for every wetland. We used the National Land Cover 2011 Dataset for Illinois (30-m grid cells) for this analysis (U.S. Geological Survey 2014). Detailed land cover classes were aggregated into general classes, e.g. high, medium, and low intensity developed land classes were classified as developed land. We retained agriculture, developed, forest, wetland and water land cover categories in the analysis because other land cover/use types were uncommon (< 5% cover within 100-m and 1-km buffers).
-
2.
Distance to Road: We measured distance between the edge of each wetland and the nearest road centerline. This distance serves as a measure of the likelihood that a wetland will be affected by a road. Road locations were obtained from ESRI’s Street Map North America dataset (ESRI and Tom Tom North America 2013).
-
3.
Road Type: We classified the road type adjacent to each wetland using five categories (Table 1). Road categories were intended to incorporate multiple factors that contribute to a road’s function in dispersing or providing habitat for non-native species; these include traffic volume, vehicle speed, and disturbance from road construction and ongoing roadway maintenance. Each of these factors increases with increasing road class ranking, e.g. interstate highways (Road Class 5) have the greatest speeds, traffic volume, and disturbance from construction and maintenance. Use of road type as a proxy for these factors is supported by findings that non-native species abundance increases with road improvement (Gelbard and Belnap 2003; Joly et al. 2011). Road type was modified from ESRI’s Street Map North America dataset (ESRI and Tom Tom North America 2013).
-
4.
Distance to Chicago: We measured the distance between each wetland centroid and a point within the city of Chicago. Urbanization across the region decreases with distance from the city center. The point location within Chicago was placed at the junction of Interstate Highways 90, 94, and 290. This point was selected because major roads within the region radiate from this junction point. Roads provide functional connectivity between the city and wetlands in the surrounding area (von der Lippe and Kowarik 2008).
-
5.
Geographic Coordinates: We determined the location of each wetland (easting and northing) using UTM coordinates.
Classification of plant communities and species
We used hierarchical clustering to assign wetlands into community type clusters based on species occurrences. Clustering was based on Sørensen’s dissimilarity index, computed for each pair of wetlands using species presence-absence data for each wetland. Ward’s minimum variance clustering, a method that splits the data based on minimizing the within group sum of squares, was used to partition groups (Borcard et al. 2011). Clusters were computed using R and the hclust function (R Core Team 2014). Selection of an appropriate number of clusters was made by reviewing the resulting dendrogram and the number of sites within each cluster.
Indicator species analysis was performed to identify characteristic species for each previously defined community cluster using R and the indicspecies package (De Cáceres and Legendre 2009; R Core Team 2014). Indicator species are those species that show fidelity to a particular cluster, as well as exclusivity to that cluster. In addition to indicator species, we characterized community clusters based on their most common species (i.e., those with the greatest number of occurrences within the community type). We assigned community type names to each cluster based on their indicator species and most common species.
Designation of species as native or non-native followed Mohlenbrock (2002) with some exceptions. Several species which are native to far southern Illinois are considered introduced in the Chicago region (Swink and Wilhelm 1994; Mohlenbrock 2002). Therefore, we considered Catalpa speciosa, Crataegus phaenopyrum, Pentaphylloides floribunda, Physocarpus opulifolius, Robinia pseudoacaia, Taxodium distichum, Thuja occidentalis and Viburnum recognitum to be non-native for this study. Phragmites australis is considered native to Illinois (Mohlenbrock 2002) and the Chicago region (Swink and Wilhelm 1994). Although native genotypes are present in the region, an introduced Eurasian genotype is more common, particularly in disturbed roadside habitats (Saltonstall 2002, 2003; Price et al. 2014). Because the majority of wetlands in this study were located along roadways, all occurrences of P. australis were assumed to be the non-native variety.
Relationship between plant communities and predictor variables
Non-native species richness and the proportion of non-native species in each of the wetland were modeled using multiple regressions. We fitted a global model containing land cover (100-m buffer), distance to road, road type, distance to Chicago, and geographic coordinates. We then used a best subsets analysis to compare all possible models based on combinations of the predictor variables. We selected the models for richness and proportion of non-natives that minimized the Mallows’ Cp and Akaike Information Criterion (AIC) scores. Tests were performed using R and the olsrr and vegan package (Hebbali 2017; Oksanen et al. 2013; R Core Team 2014).
We evaluated the relationships among species presence/absence, wetland sites and the predictor variables using canonical correspondence analysis (CCA). CCA is an approach that combines regression and ordination to relate the structure of one data set to the structure of a second data set (Borcard et al. 2011). Our first data set was a matrix of wetland sites and presence/absence of each of the species observed across the study area. The second data set was a matrix of wetland sites and the local environment and landscape configuration predictor variables. To minimize noise from infrequently occurring species, we excluded all species that occurred in fewer than 5% of wetlands. Significance testing of the model and ordination axes were performed using Monte Carlo simulations with 999 permutations. Tests were performed using R and the vegan package (Oksanen et al. 2013; R Core Team 2014).
We used analysis of variance (ANOVA) to determine whether surrounding land cover differed among the wetland community types identified from the cluster analysis. We first used principal components analysis (PCA) to reduce the five land cover types to two uncorrelated axes representing land cover gradients. ANOVA was conducted separately for the two PCA axes, and Tukey’s honest significant difference (HSD) test was used for post hoc comparisons among wetland community types.
Results
Vegetation
Of the 2005 wetlands selected for analysis, 99.25% contained at least one non-native species. Among the 1990 wetlands with non-natives, the number of non-native species ranged from 1 to 35 (mean ± S.E. = 7.11 ± 0.09), and the number of native species ranged from 0 to 145 (mean ± S.E. = 16.20 ± 0.27). On average, non-natives composed over a third of the species present in each wetland (mean ± S.E. = 34.7 ± 0.4%, median = 31.6%). In 18 sites, all species were non-native. A total of 233 non-native species were identified across the study sites. The most abundant non-native species within the sampled wetlands were Phalaris arundinacea, Typha angustifolia, and Rumex crispus (Fig. 2). Phalaris arundinacea, the most common non-native species, occurred in 73.4% of wetlands, whereas 86% of the 223 non-native species observed occurred in less than 5% of wetlands. By comparison, the most common native species, Vitis riparia, occurred in only 46.4% of wetlands.

Percentage of wetlands in which the 10 most common non-native species occurred
Non-native richness
The best approximating model for non-native species richness included the amount of water and wetland land cover, distance to the nearest road, road type, distance to Chicago, and geographic coordinates (Table 2). As expected, non-native richness was greatest in proximity to Chicago and along major roads. However, the model explained little of the variation in non-native richness (adjusted R2 = 0.04). The best approximating model for the proportion of non-natives included the amount of water, forest, agriculture, and wetland land cover; distance to the nearest road; road type; distance to Chicago; and geographic coordinates (adjusted R2 = 0.18, Table 2). Thus, the proportion of non-natives increased with a higher proportion of anthropogenic land use, the proportion increasing both with increasing agricultural land cover and decreasing proximity to Chicago. The proportion was also higher near roads and in close proximity to major roads.
Community type clusters
We used cluster analysis to separate wetlands into community types. After a visual review of the resulting dendrogram, we cut the dendrogram at a level resulting in six clusters (Online Resource 1). Common and indicator species characteristic of cluster 1 included a mix of species from forested wetlands and open, herbaceous-dominated wet meadows and marshes (Table 3). We could not easily classify this cluster as an identifiable wetland type in the region. The remaining five clusters could be identified as distinct community types based upon the common and indicator species (Table 3).
Relationships between wetland communities and human land use
Canonical correspondence analysis revealed patterns in species assemblages relative to the selected environmental predictor variables (Fig. 3). After excluding uncommon species, the resulting sample included 1999 wetlands and 115 species. Similar relationships between species occurrence data and predictor variables were observed using land cover within both 100 m and 1 km of the wetlands, with the model containing land cover at 100 m accounting for a greater amount of variance (CCA with land cover at 100 m, constrained inertia = 0.042, CCA with land cover at 1 km, constrained inertia = 0.038). The total amount of inertia explained was low, but the overall models were statistically significant (p ≤ 0.001), and all model terms were significant when tested independently (p ≤ 0.001).

Canonical correspondence analysis (CCA) ordination biplot for wetlands in community clusters 2 through 6 and the environmental variables with land cover at 100-m. The ordination with land cover variables measured at 1 km was similar, and is not shown. Grey circles denote wetland sites (n = 1999. Grey ellipses denote wetland clusters (one standard deviation around the cluster centroid): 1 – undetermined wetland type, 2 – wet meadow, 3 – marsh, 4 – forested wetland, 5 – farmed wetland/mudflat, and 6 – roadside marsh
Standard deviational ellipses, overlain on the wetlands within each cluster in the ordination plot revealed separation among some of the clusters, indicating that the community types identified in the cluster analysis could be distinguished to some degree based on the predictor variables (Fig. 3). The first CCA axis corresponded with a gradient in surrounding land cover from forests and wetlands to more intensively developed lands (Fig. 3). The second axis reflected a regional gradient from the agricultural areas west and north of Chicago to the heavily urbanized east. Species assemblages of wet meadows (cluster 2) and marshes (cluster 3) were not closely associated with the predictor variables, and these wetlands were positioned near the origin of the 2-dimensional CCA biplot (Fig. 3). Species assemblages characteristic of forested wetlands (cluster 4), and to some extent the undefined wetland cluster (cluster 1), were arrayed along the axis containing wetland and forest land cover types and were negatively associated with developed land cover and road variables (Fig. 3). Species assemblages typical of farmed wetlands and open mudflats (cluster 5) were associated with greater anthropogenic land cover, including agricultural land cover (Fig. 3). Species assemblages that are typical of roadside marshes (cluster 6) were associated with developed land cover and higher order road types (Fig. 3), often occurring near multi-lane, high volume roads. Notably, 52.7% of the roadside marshes occurred along interstate highways, whereas only 9.7% of all other wetland community types occurred along interstate highways.
Major land cover categories differed among wetland types (Fig. 4, Table 4). We used PCA to reduce land cover variables to two axes, which together explained 62% of the variation in land cover. The first axis (PC1) represented a gradient from developed to agriculture land cover, and the second axis (PC2) represented a gradient from natural (forest, water, and wetland) to anthropogenic (developed and agriculture) land cover. Cover of developed land was greater surrounding roadside marshes than all other wetland types (Fig. 4). Cover of agricultural land was greater surrounding the farmed wetland/mudflat type than undetermined wetlands, marshes, forested wetlands, and roadside marshes (Fig. 4). Natural land cover (forest and wetland) was greatest surrounding the undetermined wetlands and forested wetlands, and least surrounding roadside marshes (Fig. 4, Table 4).

Percentage of land cover types within the 100-m buffer around wetland centroids for all wetlands and each of the community type clusters
Discussion
Non-native species composition
Two objectives of our study were to classify wetland communities and characterize their non-native components. The presence of at least one non-native plant species in nearly every sampled wetland indicates that non-native plants are widespread across the Chicago region. Many non-native plant species occurred infrequently, but a few were present in a large number of sampled wetlands. Considering only rates of occurrence of individual species, there appear to be a few widespread aggressive invaders and many infrequently occurring non-natives.
Furthermore, many of the most abundant non-natives found in our sites are considered to be highly invasive, including Phalaris arundinacea (74.1% of wetlands), Typha angustifolia (51.2%), Rhamnus cathartica (37.6%), and Phragmites australis (31.4%) (Maurer and Zedler 2002; Knight et al. 2007; Lelong et al. 2007; Shih and Finkelstein 2008). Although we do not have species abundance data for the sampled wetlands, these species often become dominant where established. For example, P. arundinacea is increasing in frequency and abundance in Illinois wetlands, resulting in significant taxonomic homogenization across the state (Price et al. in press). Widespread presence of non-native species, including these widely distributed, aggressively spreading non-natives, has resulted in plant assemblages that no longer resemble native wetland communities. The most common species in both wet meadow and marsh communities was P. arundinacea, and it was the second most common species in the farmed wetland/mudflat communities. The top five indicator species of roadside marsh were non-native, including T. angustifolia and P. australis.
By classifying wetland community types and then characterizing their non-native species composition, we show that wetland plant communities in the Chicago metropolitan region reflect discernable community types. Although there were overlaps in the occurrence of the most common species, such as P. arundinacea, which was frequent in most community types, there were unique non-native indicator species for each of the community types. Thus, novel elements have been incorporated into every type of wetland in the region. For example, Rhamnus cathartica has become one of the most frequent species in forested wetlands in the Chicago region.
Drivers of plant community composition
Another objective of our study was to relate patterns of species composition to anthropogenic alterations of the landscape and local environment. We focused on two components of the local environment that may be contributing to the assembly of these non-native-dominated communities: (1) human land use, and (2) roads. Intensive human land use includes developed and agricultural land uses, and together these are the predominant land uses in the Chicago metropolitan region (U.S. Geological Survey 2014). Relationships with land cover at the local site context (100-m buffer) were stronger than those at the larger neighborhood context (1-km buffer), which is expected since disturbances in the immediate vicinity of a wetland will have the largest effect upon species composition (Bowman Cutway and Ehrenfeld 2009, 2010).
Prior to urbanization, the Chicago region was first converted from native vegetation to agricultural land uses (Iverson 1988). Agriculture remains the predominant land use along the landward periphery of the study area (U.S. Geological Survey 2014). Annual soil disturbance from plowing of agricultural fields creates open areas that can be colonized by weedy native species (e.g. Echinochloa muricata), as well as non-natives (e.g. Rumex crispus). This is particularly true for areas that are too wet for crops to be established in most years.
Developed land is now the predominant land cover within the study region (U.S. Geological Survey 2014), and was the primary land cover type within 100 m of wetlands, ranging from a low of 57.0% for the wet meadow community to 91.1% for the roadside marsh community (Fig. 4). This gradation from rural to more intense urban land cover was reflected in the primary CCA axis, with the roadside marsh communities located at the urban end of this axis. Urbanization is a driver of change in wetland communities through disturbances such as hydrologic alteration, increased nutrient inputs, and dumping and filling (Reinelt et al. 1998; Kentula et al. 2004). In addition, urbanization can generate new wetlands, for example via excavation to route or retain stormwater run-off (Moore and Hunt 2012). New wetlands are also created unintentionally via land grading or due to the increase in impervious surfaces, which results in increased run-off into adjacent low-lying areas. Flooding and increases in nutrient laden run-off from urban areas have been shown to favor the establishment of non-natives including P. arundinacea (Galatowitsch et al. 2000; Kercher and Zedler 2004). The presence of P. arundinacea in over 74% of our sites indicates pervasive changes to wetland ecosystems throughout this urbanized region.
In addition to urbanization in general, roads in particular are important determinants of plant community composition in urban wetlands. Some non-natives, especially halophytes, occur most frequently near roads in this region (Skultety and Matthews 2017). Proximity to roads and road type were important predictors for the roadside marsh community, presumably because these communities are dominated by species tolerant of high sodium and chloride concentrations found in soils adjoining roads where de-icing salts are used (Forman and Alexander 1998; Cunningham et al. 2008). Salt contamination of roadside soils acts as a strong filter on plant community composition, resulting in the replacement of intolerant glycophytic species with maritime and other salt-tolerant species (Moore 1982; Scott and Davison 1982, 1985). Indicator species in these wetlands included Phragmites australis, Solidago sempervirens, Typha angustifolia, and Hordeum jubatum, all of which are salt tolerant and common along roads (Scott and Davison 1982; Wilcox 1986; Brauer and Geber 2002; Brisson et al. 2010). Just over 52% of the wetlands in this community cluster were located alongside interstate highways and limited access state and federal highways, roads that are likely to receive the greatest applications of de-icing salts.
The CCA indicated that the pattern of species composition in urban wetlands was related not only to anthropogenic alterations to the local environment but also to the distribution of those changes across the region. The second axis of the CCA indicated that species composition changed along a gradient related to the distance to Chicago and easting. Proximity to Chicago may reflect a dispersal gradient, for example if non-native species are dispersing outward from an urban core. As reported in previous studies (Matthews et al. 2009; Aronson et al. 2015), we found that non-native species richness and proportion of non-native species decreases as distance from the urban core increases, suggesting that non-natives often first establish in major cities then spread outward. Alternatively, proximity to Chicago may serve as a proxy for land cover, particularly the degree of urbanization. Within the Chicago metropolitan area, urbanization generally decreases outward from the heavily urbanized city center. Geographic coordinates are likely capturing these same urbanization and/or dispersal gradients. Easting reflects the position of Chicago on the east side of the study area and less developed, more agricultural areas along the west side. Northing, however, does not capture a similar gradient as Chicago is in the center of the north-south axis.
Novel ecosystems
In urban areas where land cover has been greatly altered and non-native plants are abundant and widely distributed, labelling communities as simply degraded or invaded does not fully convey the extent to which urbanization reorganizes ecosystems (Pincetl 2015). The pervasiveness of non-native species in wetlands of the Chicago region suggests that some of the region’s wetlands should be considered “novel ecosystems.” We acknowledge that there is considerable disagreement regarding the usefulness of the novel ecosystem concept (Aronson et al. 2014; Hobbs et al. 2014; Murcia et al. 2014), and the presence of numerous and widely distributed non-native species in urban wetlands does not alone indicate that individual wetlands in this study are novel ecosystems. However, the fact that similar changes to the landscape have resulted in similar combinations of species suggests that anthropogenic drivers, including roads and human land uses, are creating novel environmental conditions that favor the assembly of distinct species communities dominated by particular assemblages of non-natives. In some cases, native communities have been wholly replaced. Nevertheless, there is no single agreed upon definition or way of determining novel ecosystem status (Kowarik 2011; Hallett et al. 2013; Hobbs et al. 2013; Morse et al. 2014; Truitt et al. 2015). Although there is general consensus that novel ecosystems represent a departure from historic ecosystems, there is disagreement on how much change is required before the label “novel” should be applied.
Some of the changes to wetland communities observed in this study, particularly the widespread invasion by P. arundinacea and the recurrence of unique roadside marsh assemblages along major roads, represent profound departures from historic conditions that can be interpreted as “novel.” Ecosystems fall along a continuum of change that ranges from merely degraded to truly novel, with intermediaries described as “hybrid ecosystems” (Hobbs et al. 2006, 2013). One measure of where an ecosystem falls along this continuum is whether it can be effectively restored, or whether the ecosystem has passed thresholds that make restoration difficult or prevent restoration entirely (Hobbs et al. 2009, 2013; Hallett et al. 2013). At the regional level, wetlands are difficult to restore to a natural condition, due largely to invasion by P. arundinacea (Matthews and Spyreas 2010). It is an open question whether this apparent irreversibility represents an ecological threshold (e.g., via ecological feedbacks that drive and maintain P. arundinacea dominance; Zedler 2009) or a socioeconomic threshold (e.g., a lack of effort or funding necessary for full restoration; Murcia et al. 2014). Regardless of the restorability of individual sites, however, the widespread infiltration of wetlands by non-native species and the massive restructuring of wetland communities are undoubtedly irreversible at the regional level.
References
Alpert P, Bone E, Holzapfel C (2000) Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants. Perspect Plant Ecol 3(1):52–66. https://doi.org/10.1078/1433-8319-00004
Angel J (2011) Illinois climate normals. Illinois State Water Survey, Prairie Research Institute: University of Illinois at Urbana-Champaign. http://www.isws.illinois.edu/atmos/statecli/newnormals/newnormals.htm. Accessed 5 Jan 2017
Aronson J, Murcia C, Kattan GH, Moreno-Mateos D, Dixon K, Simberloff D (2014) The road to confusion is paved with novel ecosystems: a reply to Hobbs et al. Trends Ecol Evol 29(12):646–647. https://doi.org/10.1016/j.tree.2014.09.011
Aronson MFJ, Handel SN, La Puma IP, Clemants SE (2015) Urbanization promotes non-native woody species and diverse plant assemblages in the New York metropolitan region. Urban Ecosyst 18(1):31–45. https://doi.org/10.1007/s11252-014-0382-z
Bivand R, Keitt T, Rowlingson B (2014) Rgdal: bindings for the geospatial data abstraction library. R package version 0.9–1. http://cran.r-project.org/package=rgdal
Borcard D, Gillet F, Legendre P (2011) Numerical ecology with R. Springer, New York. https://doi.org/10.1007/978-1-4419-7976-6
Bowman Cutway H, Ehrenfeld JG (2009) Exotic plant invasions in forested wetlands: effects of adjacent urban land use type. Urban Ecosyst 12(3):371–390. https://doi.org/10.1007/s11252-009-0088-9
Bowman Cutway H, Ehrenfeld JG (2010) The influence of urban land use on seed dispersal and wetland invasibility. Plant Ecol 210(1):153–167. https://doi.org/10.1007/s11258-010-9746-5
Brauer J, Geber MA (2002) Population differentiation in the range expansion of a native maritime plant, Solidago sempervirens L. Int J Plant Sci 163(1):141–150. https://doi.org/10.1086/324047
Brisson J, de Blois S, Lavoie C (2010) Roadside as invasion pathway for common reed (Phragmites australis). Invasive Plant Sci Manag 3(04):506–514. https://doi.org/10.1614/IPSM-09-050.1
Choi YD, Bury C (2003) Process of floristic degradation in urban and suburban wetlands in northwestern Indiana, USA. Nat Areas J 23:320–331
Cowardin LM, Carter V, Golet FC, LaRoe ET (1979) Classification of wetlands and deepwater habitats of the United States. U.S. Department of the Interior, fish and wildlife service: Washington, DC. FWS/OBS-29–31
Cunningham MA, Snyder E, Yonkin D, Ross M, Elsen T (2008) Accumulation of deicing salts in soils in an urban environment. Urban Ecosyst 11(1):17–31. https://doi.org/10.1007/s11252-007-0031-x
De Cáceres M, Legendre P (2009) Associations between species and groups of sites: indices and statistical inference. Ecology 90(12):3566–3574. https://doi.org/10.1890/08-1823.1
Early Detection & Distribution Mapping System (2015) Status of invasive plants in Illinois. Center for Invasive Species and Ecosystem Health, University of Georgia. http://www.eddmaps.org/tools/statereport.cfm?id=us_il. Accessed 5 Jan 2017
Environmental Laboratory (1987) Corps of Engineers wetland delineation manual. Technical Report Y-87-1. US Army Corps of Engineers, Vicksburg, MS
ESRI (2014) ArcGIS 10.2.2 for desktop. ESRI, Redlands
ESRI, Tom Tom North America (2013) U.S. and Canada detailed streets, data & maps for ArcGIS, 2013 - Street Map North America. ESRI, Redlands
Forman R, Alexander L (1998) Roads and their major ecological effects. Annu Rev Ecol Syst 29(1):207–231. https://doi.org/10.1146/annurev.ecolsys.29.1.207
Forman RTT, Sperling D, Bissonette JA, Clevenger AP, Cutshall CD, Dale VH, Fahrig L, France R, Goldman CR, Heanue K, Jones JA, Swanson FJ, Turrentine T, Winter TC (2003) Road ecology: science and solutions. Island Press, Washington DC
Foxcroft LC, Rouget M, Richardson DM, Mac Fadyen S (2004) Reconstructing 50 years of Opuntia stricta invasion in the Kruger National Park, South Africa: environmental determinants and propagule pressure. Divers Distrib 10(5-6):427–437. https://doi.org/10.1111/j.1366-9516.2004.00117.x
Galatowitsch SM, Anderson NO, Ascher PD (1999) Invasiveness in wetland plants in temperate North America. Wetlands 19(4):733–755. https://doi.org/10.1007/BF03161781
Galatowitsch SM, Whited DC, Lehtinen R, Husveth J, Schik K (2000) The vegetation of wet meadows in relation to their land-use. Environ Monit Assess 60(2):121–144. https://doi.org/10.1023/A:1006159028274
Gavier-Pizarro GI, Radeloff VC, Stewart SI, Huebner CD, Keuler NS (2010) Housing is positively associated with invasive exotic plant species richness in New England, USA. Ecol Appl 20(7):1913–1925. https://doi.org/10.1890/09-2168.1
Gelbard JL, Belnap J (2003) Roads as conduits for exotic plant invasions in a semiarid landscape. Conserv Biol 17(2):420–432. https://doi.org/10.1046/j.1523-1739.2003.01408.x
González-Moreno P, Diez JM, Ibáñez I, Font X, Vilà M (2014) Plant invasions are context-dependent: multiscale effects of climate, human activity and habitat. Divers Distrib 20(6):720–731. https://doi.org/10.1111/ddi.12206
Hallett L, Standish R, Hulvey K, Gardner M, Suding KN, Starzomski B, Murphey S, Harris JA (2013) Towards a conceptual framework for novel ecosystems. In: Hobbs RJ, Higgs ES, Hall CM (eds) Novel ecosystems: intervening in the new ecological world order. John Wiley & Sons, West Sussex, pp 16–28. https://doi.org/10.1002/9781118354186.ch3
Hansen MJ, Clevenger AP (2005) The influence of disturbance and habitat on the presence of non-native plant species along transport corridors. Biol Conserv 125(2):249–259. https://doi.org/10.1016/j.biocon.2005.03.024
Hebbali A (2017) Olsrr: tools for building OLS regression models. R package version 0.4.0. http://cran.r-project.org/package=olsrr
Hijmans RJ (2014) Raster: geographic data analysis and modeling. R Packag Version 2:3–12 http://cran.r-project.org/package=raster
Hobbs RJ, Arico S, Aronson J, Baron JS, Bridgewater P, Cramer VA, Epstein PR, Ewel JJ, Klink CA, Lugo AE, Norton D, Ojima D, Richardson DM, Sanderson EW, Valladares F, Vilà M, Zamora R, Zobel M (2006) Novel ecosystems: theoretical and management aspects of the new ecological world order. Glob Ecol Biogeogr 15(1):1–7. https://doi.org/10.1111/j.1466-822X.2006.00212.x
Hobbs RJ, Higgs E, Harris JA (2009) Novel ecosystems: implications for conservation and restoration. Trends Ecol Evol 24(11):599–605. https://doi.org/10.1016/j.tree.2009.05.012
Hobbs RJ, Higgs ES, Hall CM (2013) Defining novel ecosystems. In: Hobbs RJ, Higgs ES, Hall CM (eds) Novel ecosystems: intervening in the new ecological world order. John Wiley & Sons, West Sussex, pp 58–60. https://doi.org/10.1002/9781118354186.ch6
Hobbs RJ, Higgs ES, Harris JA (2014) Novel ecosystems: concept or inconvenient reality? A response to Murcia et al. Trends Ecol Evol 29(12):645–646. https://doi.org/10.1016/j.tree.2014.09.006
Hodkinson DJ, Thompson K (1997) Plant dispersal: the role of man. J Appl Ecol 34(6):1484–1496. https://doi.org/10.2307/2405264
Hogan DM, Walbridge MR (2007) Urbanization and nutrient retention in freshwater riparian wetlands. Ecol Appl 17(4):1142–1155. https://doi.org/10.1890/06-0185
Hutchinson T, Vankat J (1997) Invasibility and effects of Amur honeysuckle in southwestern Ohio forests. Conserv Biol 11(5):1117–1124. https://doi.org/10.1046/j.1523-1739.1997.96001.x
Iverson LR (1988) Land-use changes in Illinois, USA: the influence of landscape attributes on current and historic land use. Landsc Ecol 2(1):45–61. https://doi.org/10.1007/BF00138907
Joly M, Bertrand P, Gbangou RY, White MC, Dubé J, Lavoie C (2011) Paving the way for invasive species: road type and the spread of common ragweed (Ambrosia artemisiifolia). Environ Manag 48(3):514–522. https://doi.org/10.1007/s00267-011-9711-7
Kalwij JM, Milton SJ, McGeoch MA (2008) Road verges as invasion corridors? A spatial hierarchical test in an arid ecosystem. Landsc Ecol 23(4):439–451. https://doi.org/10.1007/s10980-008-9201-3
Kentula ME, Gwin SE, Pierson SM (2004) Tracking changes in wetlands with urbanization: sixteen years of experience in Portland, Oregon, USA. Wetlands 24(4):734–743. https://doi.org/10.1672/0277-5212(2004)0240734:TCIWWU2.0.CO;2
Kercher SM, Zedler JB (2004) Flood tolerance in wetland angiosperms: a comparison of invasive and noninvasive species. Aquat Bot 80(2):89–102. https://doi.org/10.1016/j.aquabot.2004.08.003
Knight KS, Kurylo JS, Endress AG, Stewart JR, Reich PB (2007) Ecology and ecosystem impacts of common buckthorn (Rhamnus cathartica): a review. Biol Invasions 9(8):925–937. https://doi.org/10.1007/s10530-007-9091-3
Kowarik I (2011) Novel urban ecosystems, biodiversity, and conservation. Environ Pollut 159(8-9):1974–1983. https://doi.org/10.1016/j.envpol.2011.02.022
Lelong B, Lavoie C, Jodoin Y, Belzile F (2007) Expansion pathways of the exotic common reed (Phragmites australis): a historical and genetic analysis. Divers Distrib 13(4):430–437. https://doi.org/10.1111/j.1472-4642.2007.00351.x
Maheu-Giroux M, de Blois S (2006) Landscape ecology of Phragmites australis invasion in networks of linear wetlands. Landsc Ecol 22:285–301
Matthews JW, Spyreas G (2010) Convergence and divergence in plant community trajectories as a framework for monitoring wetland restoration progress. J Appl Ecol 47(5):1128–1136. https://doi.org/10.1111/j.1365-2664.2010.01862.x
Matthews JW, Peralta AL, Soni A, Baldwin P, Kent AD, Endress AG (2009) Local and landscape correlates of non-native species invasion in restored wetlands. Ecography 32(6):1031–1039. https://doi.org/10.1111/j.1600-0587.2009.05863.x
Maurer DA, Zedler JB (2002) Differential invasion of a wetland grass explained by tests of nutrients and light availability on establishment and clonal growth. Oecologia 131(2):279–288. https://doi.org/10.1007/s00442-002-0886-8
McKinney ML (2002) Urbanization, biodiversity, and conservation. Bioscience 52(10):883–890.
McKinney ML (2006) Urbanization as a major cause of biotic homogenization. Biol Conserv 127(3):247–260. https://doi.org/10.1016/j.biocon.2005.09.005
Meunier G, Lavoie C (2012) Roads as corridors for invasive plant species: new evidence from smooth bedstraw (Galium mollugo). Invasive Plant Sci Manag 5(01):92–100. https://doi.org/10.1614/IPSM-D-11-00049.1
Minor ES, Gardner RH (2011) Landscape connectivity and seed dispersal characteristics inform the best management strategy for exotic plants. Ecol Appl 21(3):739–749. https://doi.org/10.1890/10-0321.1
Minor ES, Tessel SM, Engelhardt KAM, Lookingbill TR (2009) The role of landscape connectivity in assembling exotic plant communities: a network analysis. Ecology 90(7):1802–1809. https://doi.org/10.1890/08-1015.1
Mohlenbrock RH (2002) Vascular flora of Illinois: a field guide, 3rd edn. Southern Illinois University Press, Carbondale and Edwardsville
Moore P (1982) Coastal plants take to the road. Nature 297(5867):537–538. https://doi.org/10.1038/297537b0
Moore TLC, Hunt WF (2012) Ecosystem service provision by stormwater wetlands and ponds - a means for evaluation? Water Res 46(20):6811–6823. https://doi.org/10.1016/j.watres.2011.11.026
Morse NB, Pellissier PA, Cianciola EN, Brereton RL, Sullivan MM, Shonka NK, Wheeler TB, McDowell WH (2014) Novel ecosystems in the Anthropocene: a revision of the novel ecosystem concept for pragmatic applications. Ecol Soc 19:12
Murcia C, Aronson J, Kattan GH, Moreno-Mateos D, Dixon K, Simberloff D (2014) A critique of the ‘novel ecosystem’ concept. Trends Ecol Evol 29(10):548–553. https://doi.org/10.1016/j.tree.2014.07.006
Oksanen J, Blanchet F, Kindt R, Legendre P, Minchin PR, O’Hara R, Simpson G, Solymos P, Stevens H, Wagner H (2013) Vegan: community ecology package. R package version 2.0–10. http://cran.r-project.org/package=vegan
Pincetl S (2015) Cities as novel biomes: recognizing urban ecosystem services as anthropogenic. Front Ecol Evol 3:1–5
Price AL, Fant JB, Larkin DJ (2014) Ecology of native vs. introduced Phragmites australis (common reed) in Chicago-area wetlands. Wetlands 34(2):369–377. https://doi.org/10.1007/s13157-013-0504-z
Price EPF, Spyreas G, Matthews JW (in press) Biotic homogenization of regional wetland plant communities within short time-scales in the presence of an aggressive invader. J Ecol. https://doi.org/10.1111/1365-2745.12883
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Reinelt L, Horner R, Azous A (1998) Impacts of urbanization on palustrine (depressional freshwater) wetlands—research and management in the Puget sound region. Urban Ecosyst 2(4):219–236. https://doi.org/10.1023/A:1009532605918
Saltonstall K (2002) Cryptic invasion by a non-native genotype of the common reed, Phragmites australis, into North America. Proc Natl Acad Sci U S A 99(4):2445–2449. https://doi.org/10.1073/pnas.032477999
Saltonstall K (2003) Genetic variation among north American populations of Phragmites australis: implications for management. Estuaries 26(2):444–451. https://doi.org/10.1007/BF02823721
Schwartz MW, Thorne JH, Viers JH (2006) Biotic homogenization of the California flora in urban and urbanizing regions. Biol Conserv 127(3):282–291. https://doi.org/10.1016/j.biocon.2005.05.017
Scott NE, Davison AW (1982) De-icing salt and the invasion of road verges by maritime plants. Watsonia 14:41–52
Scott NE, Davison AW (1985) The distribution and ecology of coastal species on roadsides. Vegetatio 62(1-3):433–440. https://doi.org/10.1007/BF00044771
Seastedt TR, Hobbs RJ, Suding KN (2008) Management of novel ecosystems: are novel approaches required? Front Ecol Environ 6(10):547–553. https://doi.org/10.1890/070046
Shih JG, Finkelstein SA (2008) Range dynamics and invasive tendencies in Typha latifolia and Typha angustifolia in eastern North America derived from herbarium and pollen records. Wetlands 28(1):1–16. https://doi.org/10.1672/07-40.1
Skultety D, Matthews JW (2017) Urbanization and roads drive non-native plant invasion in the Chicago metropolitan region. Biol Invasions 19(9):2553–2566. https://doi.org/10.1007/s10530-017-1464-7
Swink F, Wilhelm GS (1994) Plants of the Chicago region, 4th edn. Indiana Academy of Science, Indianapolis
Thiele J, Schuckert U, Otte A (2008) Cultural landscapes of Germany are patch-corridor-matrix mosaics for an invasive megaforb. Landsc Ecol 23(4):453–465. https://doi.org/10.1007/s10980-008-9202-2
Thomas SM, Moloney KA (2015) Combining the effects of surrounding land-use and propagule pressure to predict the distribution of an invasive plant. Biol Invasions 17(1):477–495. https://doi.org/10.1007/s10530-014-0745-7
Tiner RW (2017) Wetland indicators: a guide to wetland formation, identification, delineation, classification, and mapping. CRC Press, Boca Raton
Truitt AM, Granek EF, Duveneck MJ, Goldsmith KA, Jordan MP, Yazzie KC (2015) What is novel about novel ecosystems: managing change in an ever-changing world. Environ Manag 55(6):1217–1226. https://doi.org/10.1007/s00267-015-0465-5
U.S. Army Corps of Engineers (2010) Regional supplement to the Corps of Engineers wetland delineation manual: Midwest Region (Version 2.0). In: Wakely JS, Lichvar RW, Noble CV, Berkowitz JF (eds) ERDC/EL TR-10-16. US Army Corps of Engineers, Vicksburg
U.S. Geological Survey (2014) 2011 National Land Cover Dataset. USGS Earth Resources Observation and Science Center, Sioux Falls
Vilà M, Ibáñez I (2011) Plant invasions in the landscape. Landsc Ecol 26(4):461–472. https://doi.org/10.1007/s10980-011-9585-3
von der Lippe M, Kowarik I (2007) Long-distance dispersal of plants by vehicles as a driver of plant invasions. Conserv Biol 21(4):986–996. https://doi.org/10.1111/j.1523-1739.2007.00722.x
von der Lippe M, Kowarik I (2008) Do cities export biodiversity? Traffic as dispersal vector across urban-rural gradients. Divers Distrib 14(1):18–25. https://doi.org/10.1111/j.1472-4642.2007.00401.x
von der Lippe M, Bullock JM, Kowarik I, Knopp T, Wichmann MC (2013) Human-mediated dispersal of seeds by the airflow of vehicles. PLoS One 8(1):e52733. https://doi.org/10.1371/journal.pone.0052733
Wilcox DA (1986) The effects of deicing salts on vegetation in Pinhook bog, Indiana. Can J Bot 64(4):865–874. https://doi.org/10.1139/b86-113
Wilhelm G, Rericha L (2017) Flora of the Chicago region: a floristic and ecological synthesis. Indiana Academy of Science, Indianapolis
Zedler JB (2009) Feedbacks that might sustain natural, invaded, and restored states in herbaceous wetlands. In: Hobbs RJ, Suding KN (eds) New models for ecosystem dynamics and restoration. Island Press, Washington, DC, pp 236–258
Zedler JB, Kercher S (2004) Causes and consequences of invasive plants in wetlands: opportunities, opportunists, and outcomes. Crit Rev Plant Sci 23(5):431–452. https://doi.org/10.1080/07352680490514673
Acknowledgements
We would like to thank the Illinois Department of Transportation for funding the original wetland delineations and the staff of the Illinois Natural History Survey who conducted the wetland delineations. Thanks to Jennifer Fraterrigo, Jonathan Greenberg, George Geatz, Adrianna Krzywicka, Edward Price and three anonymous reviewers for their helpful feedback on this manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
ESM 1
(DOCX 10 kb)
Rights and permissions
About this article

Cite this article
Skultety, D., Matthews, J.W. Human land use as a driver of plant community composition in wetlands of the Chicago metropolitan region. Urban Ecosyst 21, 447–458 (2018). https://doi.org/10.1007/s11252-018-0730-5
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-018-0730-5