
Abstract
Several studies have shown that urbanization usually leads to severe biotic homogenizing, i.e. the local extirpation of many native species and the expansion to regional scales of a small group of “urban-adaptors”, some of them exotics. Margarita Island (Venezuela) is a tropical Caribbean island that has undergone an accelerated urban development in the last 40 years. Because the island also has a high bird diversity with several endemic and threatened species, we evaluated the effect of urban development on bird species richness, assemblage structure, seasonal changes and feeding guilds. We defined an urban gradient from areas with high vegetation cover (remnant woodlands) through areas with intermediate vegetation (traditional towns) to areas with next to no vegetation (recent suburbs). Each experimental unit was replicated 3 times and birds were surveyed during the dry and rainy seasons. Richness decreased as urbanization increased, being severely depleted in the recent suburbs. The bird assemblage consisted of native species, including six endemic sub-species, but only one exotic, Columba livia. There were no seasonal changes in assemblage structure. We identified the species most tolerant and most sensitive to urbanization. Omnivorous birds were common along the gradient and granivores were also tolerant to urban development. Specialized insectivores and frugivores were the most negatively affected groups. The considerable amount of native woodland and other vegetation present in the traditional towns we evaluated, and their proximity to natural protected areas, favors the persistence of native bird species in these urban areas on Margarita Island.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Worldwide land-use conversion to urban development and the expansion of agriculture are the current main factors in the destruction of natural environments with the subsequent degradation and fragmentation of habitats, and loss of global biodiversity (Sala et al. 2000; Sanderson et al. 2002). In recent years, research has focused on the study of birds in the efforts to determine the capacity of animals to adapt to urban environments (Marzluff 2001; Hunter 2007; Müller et al. 2010). However, most studies have focused on how bird community structure and composition changes with urbanization, whilst offering limited insights into the causal factors for those changes. More detailed investigation into the effects of urban development on birds is thus needed (Müller et al. 2010).
Several studies have shown that urbanization usually leads to severe biotic homogenizing, i.e. the local extirpation of many native species together with the expansion to regional scales of a small group of “urban-adaptable” species, some of them exotics (Blair 2001; Chace and Walsh 2006; McKinney 2006; Clergeau et al. 2006). Thus, at both local and regional levels, exotic species frequently compensate the loss of native species in urban areas (Olden and Rooney 2006). Several studies have shown that in urban bird communities, the decrease in species richness is usually accompanied by a general increase in the density of these “urban-adaptable” species, which makes them more common there than in their native habitats (Clergeau et al. 1998, Melles et al. 2003; Caula et al. 2010).
Urban bird communities are affected by different selective pressures to those in the wild. They are usually in close proximity to humans and unusual predators, as well as exotic species that can act as potential competitors. Contemporary cities, however, offer habitat heterogeneity, and can also reduce pressure and generate kindly conditions including easy access to food as well as protection from adverse climatic conditions and seasonal variations (Hunter 2007; Caula et al. 2013).
A significantly high volume of urban bird studies has been carried out in northern or temperate regions. Nevertheless studies in the tropics, particularly in the Caribbean, are lacking (for a review see: Marzluff 2001; Müller et al. 2010). The high biodiversity that characterizes this zone, their insular character and the relatively small size of the islands, makes them vulnerable to change. Urban development could, thus, be detrimental to the many native and endemic animals and plants living in this region. Nueva Esparta is the only insular state in Venezuela, and is comprised by three islands in the Caribbean Sea: Margarita, Coche and Cubagua. Margarita Island is the largest and most diverse of the three, both in natural landscapes and in the flora and fauna they harbor. There are 20 endemic subspecies and two endemic species of terrestrial vertebrates, nine of which are endangered. Among these subspecies, 13 are endemic birds. Furthermore, Margarita is one of the Caribbean islands with the highest biodiversity of ecosystems and species compared to those of similar size (Sanz 2007).
The Margarita Island landscape results from a long-lasting interaction between humans and ecological factors. There is evidence of the existence of indigenous Guaiqueries since 2300 BC (Cruxent and Rouse 1961; Ayala 1996). The Guaiqueries were mostly fishermen, although they also practiced some very rudimentary agriculture, and settled on the sea shores. In 1532, Spaniard settlers began to expand agro-pastoral activities on the island, which became the main livelihood of their descendants until recently (Vila 1991). However, after the Free Tax Zone (1971) and Free Tax Port (1974) decrees, the traditional activities of fishing and agriculture declined and the economy became based on tourism and commerce. As a result, towns and cities experienced rapid expansion, with a growing public demand for infrastructure and services. Over the last 40 years, the resident population has increased from 118 000 inhab. in 1970 to 490 500 inh. in 2011, 96 % living in urban environments (INE 2012). In addition, the island regularly receives a large floating population: an average of 2 700 000 visitors came to the island yearly between 2007 and 2012 (CORPOTUR 2013). The principal urban centers have developed on the plains, and currently cover 17 % of the island territory (16 000 ha) (Sanz et al. 2011). From 1990 to 2001, Nueva Esparta state had the fourth highest growth rate in Venezuela (41.7 %; INE 2011).
Although there is clear evidence of the loss and degradation of natural ecosystems due to the accelerated urban development, as well as threats to, and even the extinction of, local species, no studies on the direct effects of urbanization on wildlife populations have as yet been undertaken (Sanz et al. 2010, 2011).
Given this situation, we specifically addressed six questions: (1) How rich is the urban bird assemblage compared with the natural one? (2) Which native bird species are found in urban areas and how are they distributed along an urban gradient? (3) What is the proportion of the urban avifauna that is of conservation interest? (4) Does bird richness and assemblage structure vary seasonally? (5) What is the foraging guild structure in the assemblage of urban birds (6) Is the urban bird community simplified, i.e. composed of only a few generalist species?
Methods
Study area
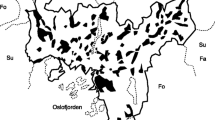
Margarita Island (952 km2, 490 500 inhab, 515 inhab/km2) is located in the Caribbean Sea, 25 km from the Venezuelan coast (10°40–11°11 N, 63°45–63°25 W). It has two sectors: the Macanao Peninsula to the West and the Oriental sector to the East where most of the urban development has occurred. The Oriental sector is formed by several mountain ranges running in a more or less north–south direction (Fig. 1). The highest peak reaches 920 masl. The climate is warm year around, with an average temperature of 27 °C. Average annual rainfall is 550 mm in the lowlands with a bimodal pattern: a main rainy season between November and January and a secondary one between June and August. However, due to the topography and wind patterns, the rainfall regime inland is complex, reaching 900 mm annually on the mountain tops, with large spatial and inter-annual variations (MARN 1997).

Location of the three (3) sectors and sampling units (black squares) we selected in urban areas of Margarita Island
Due to the climate, topography and soil effects on the vegetation, Margarita Island shows a mosaic of landscapes that range from salt marshes, mangroves, desert scrub and thorn-woodland on the coast and lowlands, to dry or cloud forests at higher altitudes. Scrub and woodlands of varying densities and floristic compositions are the main vegetation types, covering 42 % of the island (Sanz et al. 2011).
At the center of the Oriental sector is the Cerro El Copey National Park (7 130 ha). Woodlands and scrublands cover the lower slopes, and tropical dry forests are found along the river beds. Cloud forests develop above 500 masl. The park is surrounded by highways and towns, the most important being: La Asunción, San Juan, El Valle and Tacarigua.
Most of the island residents live in the more developed eastern sector, especially in the city of Porlamar and adjoining towns of Pampatar and Los Robles. Margarita’s main metropolitan area covers approximately 26 km2 (Sanz et al. 2011) and has a population of about 145 000 inhabitants (INE 2011). This area is growing very fast: between 2010 and 2011 the mean annual population growth rate reached 2 % (INE 2011).
Stratification of the urban landscape
The study was conducted in the eastern sector of Margarita Island. We chose three long-established traditional towns adjacent to Cerro El Copey National Park: La Asunción (23 616 inhab.), El Valle del Espíritu Santo (49 967 inhab.) and San Juan Bautista (39 490 inhab.). In each town two 200 × 200 m quadrats (approx.) were delineated: one in an established urbanized area within the town (Tt) and one in a remnant of tall woodland with natural vegetation (Rw). In addition, we marked out one quadrat in each of three recently built residential suburbs (Rr) near the largest city on the island, Porlamar, (98 000 inh.) for a total of nine census plots (Table 1). We used a geographic information systems (QGIS v. 2.4) to digitalize and calculate building and vegetation coverage in each sampling unit with the aim to quantify the percentage of different land covers in the three types of urban habitats chosen. The recent residential districts (Rr) near Porlamar showed the highest building densities (81–85 %), and mostly consisted of small two-storied houses with a low proportion of green space (7–14 %). The urbanized areas in the traditional towns (Tt) had a wider range of building density (47–74 %), consisting of small buildings and houses with ornamental gardens. There was also a mix of vegetation with dry forests along the rivers, woodlands (26–46 %) and grassland (3–4 %) in non-urbanized patches, and native and exotic trees and bushes on the streets. The remnant woodlands (Rw) within the urban matrix consisted of plots of 6–10 ha of native dry forest. Trees were 5–20 m tall and deciduous species such as Bursera simaruba, Lonchocarpus spp, Inga spp and Platymiscium diadelphum, dominated. These sample units were used to form an urbanized gradient as proposed by McDonnell and Hahs (2008): from sites with the most vegetation (remnant woodlands), followed by urbanized areas in the traditional towns that maintain vegetation along the streets and gardens and lastly, zones with the least vegetation cover: the recent residential developments in the suburbs.
Avian survey
We recorded birds within each sampling unit over four periods: two in the dry season (October 2010 and May 2011) and two in the rainy season (February and July 2011). These sampling dates were selected to include the northern migratory season in Venezuela (September to March). Each sampling unit was visited twice during each field trip. Bird counts were carried out in the morning on days with no wind or rain, beginning 15 min after sunrise and ending approximately 3 h later. During each visit we walked the whole area defined by our sampling square for 20 min in the two urban zones (Tt and Rr), but in the remnant woodlands (Rw) we surveyed at two points 150 m apart for 10 min each. We recorded all the birds heard or seen with the help of binoculars 10 × 50. Species identification was based on Hilty (2003) and Restall et al. (2007) and taxonomic classification followed SACC South American Classification Committee (2013).
Data analysis
For the bird list we included every species registered, but for the quantitative analyses we selected only the species recorded at least twice during a survey period.
We compared the species richness among the three types of urban habitats considering every location as replicates (n = 3 replicates) (Rw: remnant woodland, Tt: traditional town, Rr: recent residential area). We calculated the corrected species richness with COMDYN software (Hines et al. 1999) that accounts for heterogeneity in species detectability using the jackknife estimator.
In order to describe the variation in the distribution of each species along the urban gradient, we defined “frequency” as the number of times a species was recorded, expressed as a percentage of the total number of visits (N = 8) to each sampling unit within each habitat type (N = 9). We analyzed the structure of the bird assemblages per sample and per season using correspondence analysis considering presence/absence (MVSP 3.1 Software, Kovach Computing Services). Species were classified in trophic guilds following Poulin et al. (1994) and personal data, and their occurrences were compared among urban areas and between urban and natural habitats. In spite of the criticism of the use of guilds, several supra-specific groups (e.g. guilds, functional groups) have been widely adopted for the evaluation, management and monitoring of ecosystems (Milesi et al. 2002).
Results
We recorded 34 species of birds along the urban gradient over both seasons; this represents 58 % of all the land bird species currently found in similar nearby natural vegetation in Cerro El Copey National Park (58 species) (Sanz et al. 2010) (Appendix). Many of the species not registered in the study, but reported for the national park, were migratory species or species found at low abundances on Margarita Island (V. Sanz unpublished data).
Out of the seven endemic subspecies registered for dry forests and woodlands in Cerro El Copey N.P., six were seen, and four (Hypnelus bicinctus stoicus, Icterus nigrogularis helioeides, Quiscalus lugubris insularis, Xiphorhynchus picus longirostris) were recorded more than once along the urban gradient. Two of these were recorded in the whole gradient surveyed (Table 2). None of the species recorded was classified as threatened by any international body. No migratory birds were registered during the census, neither in October nor in February; months when northern migratory species fly over or stay on the island.
Thirty (30) species were recorded at least twice, and these are the ones considered in the following analysis. Species richness decreased with an increase in the percentage of built up area and a decrease in vegetation cover (Fig. 2). Thus, the estimated species richness was highest in the remnant woodlands (32, SE 2.4), intermediate in the traditional towns (27, SE 3.5) and lowest in the recent suburbs, (14, SE 1.38): less than half that of the remnant woodlands.

Estimated bird-species richness in the three types of urban habitats. Vertical bars represent standard errors. Total species observed in parenthesis. Rw remnant woodland; Tr traditional town, and Rr recent residential. Numbers of census plots in parenthesis at axis “X”. Data from Cerro El Copey NP come from bibliography, museum records and mist-net sampling between 2006 and 2011
We analyzed the use of species for the different habitat types surveyed (Table 2). Q. lugubris insularis, C. livia, and C. talpacoti were recorded with high frequencies in sectors of high build density, taking effective advantage of resources found within the urban area, as food, water, and shelter. Five species that showed a high frequency (>45 %) along urban gradient, can be considered generalists, mainly M. gilvus, C. squammata, and C. flaveola. However, no species was only registered from the recent residential areas: the species using the more urbanized areas were also present at least in the traditional towns.
Another group of twelve species were present in highly vegetated urban areas: remnant woodland and sectors within the traditional towns, some of them with a preference for the remnant woodland, e.g. M. rubricapillus, T. doliatus, L. verreauxi, and T. nudigenis, among others (Table 2). Five species were recorded only from remnant woodlands: O. ruficauda, F. grisea, S. arenarum, A. tobaci, and T. naevia and could thus be considered the species most sensitive to changes due to urbanization. Finally, 25 species recorded in Cerro El Copey National Park were not recorded in the city (Appendix).
For the correspondence analysis of the presence of bird species in the urban landscape we ran 18 samples (nine sampling units over two seasons) (Fig. 3). The first two axes accounted for 43.91 % of the total variance, and segregated three groups of samples: remnant woodlands, traditional towns and recent suburbs, independent of the season. This means that the assemblage of species present in each habitat type along the gradient does not change with the seasons: thus any differences are based on habitat structure. The species in the center of the diagram (C. flaveola, C. squammata, I. nigrogularis helioeides, M. gilvus, Q. lugubris insularis, T. glaucocolpa inter alia) were more associated with the traditional towns, but were also present in the other two habitats, as was mentioned above.

Scatter diagram of correspondence analysis in dry and rainy seasons: sampling unit N(9+9)=18, bird species N = 30. Cumulative percentage variance of species data by Axis I and Axis II =43.4 %. Keys: Rw remnant woodland, Tt traditional town, Rr recent residential, r rain season, d dry season. Species acronyms in Appendix
Changing the focus from individual species to functional groups, such as feeding guilds, can give us an idea of the resources the birds are using in urban areas. Although the proportion of species belonging to each guild was very similar in all the habitats surveyed (Fig. 4), seven guilds were recorded from the urban habitats, whereas in the natural woodlands species belonging to ten different guilds were registered. A more detailed analysis of guilds and species along the urban gradient shows that, as expected, omnivores become more important as urbanization increases (Fig. 5). Species such as M. gilvus, Q. lugubris insularis, I. nigrogularis helioeides, C. ani, and C. livia were found in all three habitat types, and three of these were found more frequently in urbanized areas (Table 3), where they eat several kinds of food, including human scraps. Several of these species are ground foragers and benefit from the open spaces found in cities. Another group that seems to have benefited by urban growth are the granivores, mainly pigeons and doves (Fig. 5, Table 3). These species feed off grass seeds in towns and suburbs. As for the omnivores, the granivores are able to use these open spaces as they often forage whilst walking on the ground.

Guilds frequency in urban and natural (El Copey N.P.) habitats. N = 60 species. OMN omnivore, NEC nectarivore, FRU frugivore, GR granivore, IN insectivore. CARV carnivore. Keys: Rw remnant woodland, Tt traditional town, Rr recent residential

Guilds frequency in urban sectors and El Copey N.P. N = 60 species. OMN omnivore, NEC nectarivore, FRU frugivore, GR granivore, IN insectivore. Keys: Rw remnant woodland, Tt traditional town, Rr Recent residential
In contrast, insectivorous birds were one of the guilds most negatively affected by urbanization: their abundance decreased along the gradient remnant woodlands – recent residential suburbs (Fig. 5). This is a broad group that includes species that employ several kinds of insect manipulation. Specialized insectivorous birds, such as X. picus longirostris, glean food from tree trunks, and were rare even in the remnant woodlands. Foliage gleaners, such as F. grisea or P. plumbea, need shrubs or trees to find food, and were absent from the recent residential developments with little vegetation. The only species able to use the more urban areas was the aerial feeder T. melancholicus, which was present along the whole gradient, as it benefits from open spaces (Table 3). Frugivorous or frugivorous-insectivorous species, such as O. ruficauda, T. glaucocolpa, T. palmarum, M. rubricapilus, H. flavipes were found only or mainly in the remnant woodlands and traditional towns, where fruits and insects were available (Fig. 5, Table 3). The nectarivores-insectivores, all of them hummingbirds, were uncommon species along the whole gradient; possibly their presence was underestimated as they use seasonally abundant resources, such as certain flowers.
Discussion
The overall pattern we found in Margarita Island of decreasing species richness with increasing building density and decreasing vegetation, was consistent with results from earlier studies on urban bird populations in both temperate and tropical regions (e.g. Beissinger and Osborne 1982; Cam et al. 2000; Melles et al. 2003; Sandstorm et al. 2006; Palomino and Carrascal 2006, Ortega-Álvarez and MacGregor-Fors 2009; Carbó and Zuria 2011). The intermediate disturbance hypothesis, where diversity peaks at a midpoint along a disturbance gradient (Catford et al. 2012; Fox 2013), was not supported by our results. An explanation for this could be that the only exotic species registered during our study was C. livia (all the others are natives of Margarita Island), and no species was found only in urbanized areas. Although we do not have data on the initial bird community of the urbanized areas before they were built to directly assess changes in community structure over time, we can assume that these initial bird assemblages would have been similar to those currently found in nearby natural landscapes, which can thus be used as a surrogate “before urbanization” condition . This means that urban development on Margarita Island has led to a loss of bird diversity: almost half the species reported from Cerro El Copey National Park were not detected in any of the habitat types, along the studied gradient.
It is nevertheless true that the traditional town and remnant woodland habitats are able to maintain an important number of land bird species (34) representing 31 % of the total number of land birds recorded from the island (Sanz et al. 2010). These include species of particular interest for conservation, such as endemic sub-species. In contrast, the number of species in the recent residential areas was severely depleted. This result can be explained by two factors: (1) the existence of a considerable amount of green space and vegetation in the first two habitat types, unlike the modern suburbs where vegetation is scarce or practically absent and (2) the proximity of the traditional towns to natural protected areas, that enable the movement of birds from one habitat to the other (Blair 1996; Sandstorm et al. 2006).
Our urban gradient follows a reduction in vegetation and an increase in urbanization; usually these conditions also implicate the modification of other variables as: elevated air and soil temperatures due to urban heat island effects, atmospheric and water pollution, a greater concentration of heavy metals and high noise levels (Grimm et al. 2008; McDonnell et al. 1997). To live in cities birds must adapt to these novel conditions (Hunter 2007). We observed four types of adaptive responses as urbanization increased and vegetation decreased (McDonnell and Hahs 2008): (1) the positive response of “urban exploiters”, such as Q. lugubris insularis and C. livia which are the most common birds found in built up areas. Both species are omnivores, often opportunistic, naturally inhabit open landscapes, and are well known as human commensals, (2) the intermediate response of “urban adapters” such as M. gilvus, T. glaucocolpa, I. nigrogularis, C. flaveola which are most common in highly vegetated urban areas and belong to different foraging guilds, (3) the transitional response of “urban tolerant species”, such as H. flavipes, F. grisea, O. ruficauda, M. rubricapillus, T. doliatus, and F. passerinus, which were common in remnant woodlands, and feed on fruits and/or insects, (4) the negative response of “urban intolerant species” or “urban avoiders”, such as Cardinalis phoeniceus or C. gujanensis, that are mainly insectivores and granivores. These species were not detected in any of the studied urban habitats type, but are present in natural habitats on the island. Two bird species, Turdus nudigenis and Amazilia tobaci, common from other urban areas on mainland Venezuela, were surprisingly rare or absent from the traditional towns surveyed. The first of these could have been displaced from the urban areas on Margarita Island by the extremely common M. gilvus. Similarly, the Amazilia hummingbird could have been replaced by the larger L. fallax, also common on the island and present, although in low frequencies, along the entire gradient studied.
The homogenization hypothesis (McKinney and Lockwood 1999, Rahel 2002), assumes that cities are occupied by a relatively uniform group of “urban-exploiter and “urban adaptable” species, together with some exotics, which tolerate the particular conditions of a built-up environment. Simultaneously, many native “urban avoider” species, unable to tolerate urban conditions, disappear from the city landscape. This homogenization is a regional process whereby the similarity of bird species among the more urbanized sectors is determined more by the degree of urbanization than by other local effects. In this study, we were not able to establish whether bird species homogenization occurs among cities in Venezuela. However, we are able to anticipate some of the effects of urbanization. The only exotic species on Margarita Island is Columbia livia. Thus, the displacement of native species by introduced exotics has not been observed. Nevertheless, with increasing urbanization, the assemblage of birds is becoming more simplified both at the guild and species levels, with a predominance of generalist omnivores and granivores. If this pattern is repeated at the regional level we could expect to observe the functional homogenization of bird populations among other Venezuelan cities.
The urban bird assemblages we surveyed did not change significantly with the season. These results differ from those reported by other studies where the movement of birds into the most urbanized sectors of the city has been observed during winter in North America and Europe (Clergeau et al. 1998; Fernández-Juricic and Jokimäki 2001) and during the dry season in the tropics (Torga et al. 2007). On Margarita Island there are four seasons with the dry and wet periods intercalated, thus the limiting season is shorter than on the mainland. Furthermore, the traditional towns on the island do not have big recreational parks, exotic vegetation, lagoons or fountains; development is based rather on tree-lined streets, small houses with gardens, and small fields of native vegetation. Thus, these urban areas do not offer such a contrast in resources compared to larger cities which have more resources that can be exploited by birds during the lean season.
The assemblage of trophic guilds in the urban areas was similar to that found for the remnant woodland, but with a few of the guilds absent. Although the percentages of species within each guild did not differ between urban and natural areas, the carnivore guilds were notably absent from the urban gradient. A total of eight species of raptor have been reported from Margarita Island, and two of these, Gampsonyx swainsonii and Buteo albicaudatus, were observed in nearby natural forests. None of these species were recorded, however, from the sampling units. In the literature, there are reports of Falco peregrinus and other raptors (e.g. the Osprey, Pandion haliaetus) using artificial structures for nest sites. Chace and Walsh (2006) even suggested that urban landscapes could provide superior habitats for some raptors because they are usually free from human persecution and have a high availability of abundant food. As regards omnivorous and granivorous species, these were favored by urbanization whereas specialized insectivores were negatively affected as has been reported for temperate zones (Beissinger and Osborne 1982; Clergeau et al. 1998, Carbó and Zuria 2011).
The results of our study suggest that forest-woodland species, as well as specialized insectivores or frugivores, are more affected than omnivores of open or semi open habitats as urbanization continues.
Conclusions
We found that the composition of bird communities is stable all year around with a spatial structure influenced by the abundance of vegetation. Although increased urbanization can lead to a decrease in avifauna species richness, towns do include resources that are important for a number of urban-adapted birds. This suggests that there are ways in which the conservation of bird diversity can be accommodated within urban planning frameworks.
The urban gradient selected is also a historical gradient, in which towns from colonial times up to the present day have been developed whilst maintaining vegetation in the urban areas, planting tall trees along the main streets and in the town squares, thus assuring habitat for native birds. On the contrary, in recent urban developments, particularly private housing estates built during the last 30 years, the priority has been to achieve the highest human population density possible, without setting aside spaces for parks or even leafy tree-lined streets. If this trend continues, modern urban areas will not be adequate for the conservation of an important part of bird biodiversity on the island. As we seen, however, it should be possible to increase bird species richness in modern urban developments by improving the vegetation structure so that it resembles that of the more traditional towns. Urban landscapes that contain sufficient heterogeneity, complexity and connectivity of vegetation can accommodate a diverse bird community including endemic subspecies of conservation value. The development of parks and gardens in residential areas, the planting of native trees along the streets and the maintenance of local woodlands and forests should all be taken into account for the adequate urban planning of Margarita Island.
References
Ayala C (1996) La etnohistoria prehispánica guaiquerí. Antropológica 82:5–127
Beissinger SR, Osborne D (1982) Effects of urbanization on avian community organization. Condor 84:75–83
Blair RB (1996) Land-usage and avian species diversity along an urban gradient. Ecol Appl l6:506–519
Blair RB (2001) Creating a homogenous avifauna. In: Marzluff JM, Bowman R, Donnelly R (eds) Avian ecology and conservation in an urbanizing world. Kluwer Academic Publishers, Massachusetts, pp 459–486
Cam E, Nichols JD, Sauer J, Hines J, Curtis F (2000) Relative species richness and community completeness: birds and urbanization in the Mid-Atlantic States. Ecol Appl 10:1196–1210
Carbó P, Zuria I (2011) The value of small urban greenspaces for birds in a Mexican City. Landsc Urban Plan 100:213–222
Carrascal, L.M. & Palomino, D. 2002. Determinantes de la riqueza de especies de aves en las islas Selvagem y Canarias. Ardeola 49(2), 211–221
Catford JA, Daehler CC, Murphy HT, Sheppard AW, Hardesty BD, Westcott DA (2012) The intermediate disturbance hypothesis and plant invasions: implications for species richness and management. Perspect Plant Ecol 14:231–241
Caula S, Sirami C, Marty P, Martin JL (2010) Value of an urban habitat for the native Mediterranean avifauna. Urban Ecosyst. doi:10.1007/s11252-009-0104-0
Caula S, Villalobos A, Marty P (2013) Seasonal dynamics of bird communities in urban forest of a Mediterranean city (Montpellier, Southern France). Urban Ecosyst. doi:10.1007/s11252-013-0295-2
Chace JF, Walsh JJ (2006) Urban effects on native avifauna: a review. Landsc Urban Plan 74:46–69
Clergeau P, Savard JPL, Mennechez G, Falardeau G (1998) Bird abundance and diversity along an urban–rural gradient: a comparative study between two cities on different continents. Condor 100:413–425
Clergeau P, Croci S, Jokimäki J, Kaisanlahti-Jokimäki ML, Dinetti M (2006) Avifauna homogenisation by urbanisation: analysis at different European latitudes. Biol Conserv 127:336–344
Cruxent JM, Rouse I (1961) Arqueología cronológica de Venezuela. Vol.1. Unión Panamericana-Facultad de Economía de la Universidad Central de Venezuela. Washington DC
CORPOTUR (Corporación de Turismo del Estado Nueva Esparta) (2013) Estadísticas turísticas del Estado Nueva Esparta. Resumen histórico de entrada anual de viajeros al estado Nueva Esparta por medios de transporte 1994–2009. www.corpoturmrgarita.gob.ve (cons. Agosto 2011)
Fernández-Juricic E, Jokimäki J (2001) An habitat island approach to conserving birds in urban landscapes: case studies from southern and northern Europe. Biodivers Conserv 10:2023–2043
Fox JW (2013) The intermediate disturbance hypothesis is broadly defined, substantive issues are key: a reply to Sheil and Burslem. Trends Ecol Evol 28:572–573
Grimm NB, Faeth SH, Golubiewski NE, Redman CL, Wu J, Bai X, Briggs JM (2008) Global change and the ecology of cities. Science 319:756–760
Hilty S (2003) Birds of Venezuela. Princeton University Press, Princeton
Hines JE, Boulinier T, Nichols JD, Sauer JR, Pollock KH (1999) COMDYN: software to study the dynamics of animal communities using a capture-recapture approach. Bird Study 46(suppl):S209–S217
INE (2011) Censo Población y Vivienda. Instituto Nacional de Estadística. República Bolivariana de Venezuela. http://www.ine.gov.ve/index.php?option=com_content&view=category&id=95&Itemid=26. (cons. agosto 2013)
INE (2012) Censo Población y Vivienda. Instituto Nacional de Estadística. República Bolivariana de Venezuela. http://www.ine.gov.ve/demografica/censopoblacionvivienda.asp (cons. agosto 2013)
MARN (1997) Atlas Básico del Estado Nueva Esparta. Ministerio del Ambiente y de los Recursos Naturales- Gobernación Estado Nueva Esparta- Fundación para el Desarrollo del Estado Nueva Esparta, Porlamar
Marzluff JM (2001) Worldwide urbanization and its effects on birds. In: Marzluff JM, Bowman R, Donnelly R (eds) Avian ecology and conservation in an urbanizing world. Kluwer Academic Publisher, Massachusetts
McDonnell MJ, Hahs AK (2008) The use of gradient analysis studies in advancing our understanding of the ecology of urbanizing landscapes: current status and future directions. Landsc Ecol 23:1143–1155
McDonnell MJ, Pickett STA, Groffman P, Bohlen P, Pouyat RV (1997) Ecosystems processes along an urban-to-rural gradient. Urban Ecosyst 1:21–36
McKinney ML, and Lockwood JL (1999) Biotic homogenization: a few winners replacing meny loser in the nest mass extinction. TREE, 14 (11) 450.
McKinney ML (2006) Urbanization as a major cause of biotic homogenization. Biol Conserv 127:247–260
Melles S, Glenn S, Martin K (2003) Urban bird diversity and landscape complexity: species-environment associations along a multiscale habitat gradient. Conserv Ecol [online] URL: http://www.consecol.org/vol7/iss1/art5/.
Milesi FA, Marone L, Lopez de Casenave J, Cueto VR, Mezquida ET (2002) Gremios de manejo como indicadores de las condiciones del ambiente: un estudio de caso con aves y perturbaciones del hábitat en el Monte Central, Argentina. Ecologíca Austral 12:149
Müller N, Ignatieva M, Nilon CH, Werner P, Zipperer, WC (2010) Patterns and trends in urban biodiversity and landscape design. In: Müller et al. Urban Biodiversity and Design. Wiley-Blackwell, pp 123–174
Olden JD, Rooney TP (2006) On defining and quantifying biotic homogenization. Global Ecol Biogeogr. doi:10.1111/j.1466-822X.2006.00214.x
Ortega-Álvarez R, MacGregor-Fors I (2009) Living in the big city: effects of urban land-use on bird community structure, diversity and composition. Landsc Urban Plan 90:189–195
Poulin B, Lefebvre G, McNeil R (1994) Diets of land birds from northeastern Venezuela. Condor 96:354–367
Rahel, FJ, (2002) Homogenization of freshwater faunas. Annu.Rev.Ecol.Syst. 33:291–315
Restall R, Rodner C, Lentino M (2007) Birds of Northern South America: An identification guide. Yale University Press
Sala OE, Stuart Chapin S, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwal E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M, Poff NL, Sykes MT, Walker BH, Walker M, Diana H, Wall DH (2000) Global biodiversity scenarios for the year 2100. Science 287:1770–1774
Sanderson EW, Malanding J, Levy M, Redford K, Wannebo AV, Woolmer G (2002) The human footprint and the last of the Wild. Bioscience. doi:10.1641/0006-3568(2002)052[0891:THFATL]2.0.CO;2
Sandstorm UG, Angelstam P, Mikusinski G (2006) Ecological diversity of birds in relation to the structure of urban green space. Landsc Urban Plan 77:39–53
Sanz V (2007) ¿Son las áreas protegidas de la Isla de Margarita suficientes para mantener su biodiversidad? Análisis espacial del estado de conservación de sus vertebrados amenazados. Mem Fund La Salle de Cienc Nat 167:111–130
Sanz V, Oviol L, Medina A, Moncada R (2010) Avifauna del estado Nueva Esparta, Venezuela: recuento histórico y lista actual con nuevos registros de especies y reproducción. Interciencia 35:329–339
Sanz V, Riveros M, Gutiérrez M, Moncada R (2011) Vegetación y uso de la tierra en el estado Nueva Esparta, Venezuela: un análisis desde la ecología del paisaje. Interciencia 36:881–887
South American Classification Committee (SAAC). A classification of the bird species of South America. American Ornithologists’ Union. http://www.museum.lsu.edu/~Remsen/SACCBaseline.html
Torga K, Franchin AG, Júnior OM (2007) A avifauna em uma seção da área urbana de Uberlândia, MG. Biotemas 20:7–17
Vila, P. (1991) Visiones Geohistóricas de Venezuela. Fundación de Promoción Cultural de Venezuela. Caracas, Venezuela. 465pp
Acknowledgments
This work was funded by Council for Scientific and Technological Development of University of Carabobo (CDCH-UC) No 0432–10 and Venezuelan Institute for Scientific Research (IVIC). We are grateful to two anonymous referees whose comments helped to improve the manuscript. To Rodrigo Díaz for his assistance in the field and Elysa Silva for the GIS support in the digitalization of the urban gradient.
Author information
Authors and Affiliations
Corresponding author
Additional information
Virginia Sanz and Sabina Caula have contributed equally to this publication.
Appendix
Appendix
Rights and permissions
About this article

Cite this article
Sanz, V., Caula, S. Assessing bird assemblages along an urban gradient in a Caribbean island (Margarita, Venezuela). Urban Ecosyst 18, 729–746 (2015). https://doi.org/10.1007/s11252-014-0426-4
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-014-0426-4