
Abstract
Prolactin (PRL) has been long deemed as a hormone involved only in female reproduction. However, PRL is a surprising hormone and, since its identification in the 1970s, its attributed functions have greatly increased. However, its specific role in male health is still widely unknown. Recently, low PRL has been associated with reduced ejaculate and seminal vesicle volume in infertile subjects. In addition, in men consulting for sexual dysfunction, hypoprolactinemia has been associated with erectile dysfunction and premature ejaculation, findings further confirmed in the general European population and infertile men. Several metabolic derangements, recapitulating metabolic syndrome, have also been associated with low PRL both in men with sexual dysfunction and from the general European population. In men with sexual dysfunction, followed-up for more than 4 years, low PRL was identified as an independent predictor of the incidence of major adverse cardiovascular events. Finally, an association with anxiety or depressive symptoms has been found in men with sexual dysfunction and from the general European population. While a direct role for impaired PRL function in the pathogenesis of these reproductive, sexual, metabolic and psychological disorders is conceivable, the possibility that low PRL is a mirror of an increased dopaminergic or a decreased serotonergic tone cannot be ruled-out. Hyperactivity of the dopaminergic system can explain only a few of the aforementioned findings, whereas a hypo-serotonergic tone fits well with the clinical features associated with low PRL, and there is significant evidence supporting the hypothesis that PRL could be a mirror of serotonin in the brain.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Prolactin (PRL) is a single-chain non-glycosilated polypeptide hormone made up of 199 amino acids. Its gene is located on chromosome 6. It is mainly expressed by the anterior pituitary gland by two populations of cells, lactotroph cells, which represent 15–25 % of the anterior pituitary cells and secrete only PRL, and mammosomatotroph cells, a minor cell population, which co-secrete PRL and growth hormone (GH). PRL, GH and placental lactogen share a similar structure because they derive from a common ancestral gene. The common origin of PRL and GH is also demonstrated by the dependence of lactotroph on somatotroph cells during ontogenesis. In fact, in transgenic mice with somatotroph targeted ablation, induced by the germ-line expression of GH-diphtheria toxin and GH-thymidine kinase fusion genes, lactotrophs are almost completely absent, suggesting that lactotrophs derive from post-mitotic differentiation of somaotrophs [1]. Beside the pituitary, 20 % of PRL concentration relies on extrapituitary production. Extrapituitary PRL is produced by several tissues, such as the brain, mammary epithelial cells and tumors, endometrium, myometrium, lacrimal and sweat glands, skin fibroblasts and lymphoid organs and cells [2], and this contribution seems to be very important, since treating hypophysectomized rats with anti-PRL antibodies results in immune dysfunction and death [3].
PRL acts through the interaction with its receptor (PRLR). It is part of the class 1 cytokine receptor, which includes also receptors for several interleukins, colony stimulating factors, erythropoietin and thrombopoietin. Similarly to their ligands, PRLR and GH receptors share several structural and functional characteristics. PRLR is a single pass trans-membrane chain that is encoded by its gene on chromosome 5. It is subject to alternative splicing and this mechanism provides three different isoforms that differ in the length of the cytoplasmic domain (short, intermediate and long PRLR) [see 4–6 for review]. In addition, a soluble form, identical to the extracellular domain of the trans-membrane PRLR, has been identified [see 7–9 for review]. PRL binds to its receptor through a binding site, forming an inactive complex, which allows for the interaction of a second PRL binding site to another PRLR, thus forming a trimeric complex [5, 10]. This mechanism activates a cascade of intracellular events that mediates the transduction of the PRL signal. Several signaling pathways are activated by the PRL-PRLR interaction [see 11 for review], but the most important is the phosphorylation of JAK/STAT molecules that leads to gene transcription [5]. Beside the mammary gland, PRLR is also expressed in several tissues involved in reproduction, such as the brain, ovaries, endometrium, testes, epididymis, seminal vesicles and prostate [11]. Besides PRL functions in immunity, other roles, such as water and electrolyte balance, growth, development and tumorigenesis, are also emerging [see 11 for review]. The most important and known function of PRL in humans is the development of the mammary gland during pregnancy and lactation. In addition, PRL plays a role in other functions tightly linked to maternity, as suggested by the observation that in mice with altered expression of PRLR, ovulation is impaired [11] and induction of maternal behavior is compromised [12]. Recently, this role of PRL in parental behavior has been suggested also in men [13]. Although in the pituitary PRL is present in equal amounts in males and females, its physiological role, well established for female reproduction, is still obscure for the male counterpart.
Concerning pathologic functions, the effect of abnormally high PRL levels has been extensively investigated both in males and females [14]. Conversely, the effects of low PRL have not received similar attention, and little is known about the associated clinical features. However, recent and emerging evidence shows that low PRL levels are associated with different pathological correlates and, for this reason, hypoprolactinemia may deserve a specific chapter in clinical manuals, as a new clinical syndrome.
The aim of this review is to provide an overview of the available knowledge on the role of PRL in men, with a particular consideration of the most novel data concerning the clinical correlates of low PRL levels.
2 Regulation of PRL secretion
PRL differentiates from the other pituitary hormones because of its regulation. In fact, PRL secretion has a prevalent inhibitory control carried out by dopamine (DA). Furthermore, PRL regulation is not dependent on a signaling from its peripheral target and PRL itself can stimulate DA release, leading to a self-regulated short-loop feedback. DA is not the only PRL inhibitory factor. In fact, several molecules, including endothelin-1, transforming growth factor-β1, and calcitonin, which act in an endocrine or paracrine fashion, have been identified. PRL is also regulated by stimulating agents. Among them, serotonin (5-hydroxy-triptamine, 5-HT), thyrotropin stimulating hormone (TRH), basic fibroblast growth factor, epidermal growth factor, vasoactive intestinal peptide, oxytocin, estrogen and opiates should be mentioned. The detailed description of the numerous factors involved in PRL regulation is beyond the aim of the present review and a complete overview is provided elsewhere [see 15 for review]. However, for the importance in PRL metabolism and for its possible implication in recent findings on PRL clinical correlates in men, the role of DA and 5-HT deserve a brief description.
2.1 Dopamine
Dopamine is secreted by neurons within the arcuate nucleus of the hypothalamus. These dopaminergic cells are not homogenous from an anatomical point of view, and they include three different types of neurons. In particular, the tuberoinfundibolar dopaminergic neurons (TIDA) are located in the dorso-medial arcuate nucleus and their axons end in the external zone of the median eminence. The tuberohypophyseal neurons (THDA) are located in the rostral part of the arcuate nucleus and project to the neural lobe of the pituitary. Finally, the periventricular hypophyseal (PHDA) are in the periventricular nucleus and terminate in the intermediate lobus of the pituitary. TIDA release DA into the pituitary portal vessels, while THDA and PHDA neurons secrete DA into the neurohypophysis and it is led to the anterior part of the gland by short portal vessels. Despite these differences, from a functional point of view, TIDA, THDA and PHDA neurons can be considered a single functional unit of a PRL-inhibiting system. DA in the pituitary interacts with its receptors D2 and D4 [16, 17] and it decreases the secretion of PRL [18], but it is also involved in the down-regulation of PRL gene expression [19] and in inhibiting lactotroph proliferation [20]. In turn, PRL regulates DA synthesis and release through a short-loop feedback. The PRL receptor (PRLR) is expressed in TIDA, THDA and PHDA neurons. Intracerebroventricular administration of PRL [21] induces hypothalamic DA synthesis and release of DA from TIDA neurons into pituitary stalk blood [22]. Conversely, hypophysectomy or use of D2 agonists decrease DA synthesis [23].
2.2 Serotonin
Serotonin is a neurotransmitter widely represented in the brain and it is involved in several functions and behaviors. A significant amount of evidence supports the role of 5-HT as a PRL stimulating factor. In rats, the direct intraventricular infusion of 5-HT or the intraperitoneal administration of 5-HT precursors, such as 5-hydroxy-tryptophan, leads to an increase in serum PRL levels [24, 25], and a 5-hydroxy-tryptophan-induced increase in serum PRL has been demonstrated also in humans [26]. Furthermore, p-chlorophenylalanine induced 5-HT depletion in the brain decreases basal PRL levels and their suckling-dependent increase [27, 28]. Similarly, a decrease in basal and stimulated PRL after the administration of 5-HT blockers, such as cyproheptadine, metergoline and methysergide [29, 30], has been demonstrated whereas the administration of fluoxetine, a selective 5-HT re-uptake inhibitor (SSRI), led to an increase in PRL secretion [25]. The 5-HT neurons that are mainly responsible for the serotonergic signaling involved in PRL regulation are located in the dorsal raphe nucleus [31, 32]. Lactotroph cells express 5-HT receptors [33, 34], however, when these cells are treated in vitro with 5-HT, no response in PRL secretion is observed [35–37]. Furthermore, the infusion of 5-HT into the anterior pituitary or into cannulated portal vessels does not result in increments in serum PRL levels [24]. These data suggest that 5-HT does not directly affect pituitary PRL secretion, but its activity is indirect and mediated by other PRL stimulating factors. Among the 5-HT receptors, 5HT1A, 5HT2A, and 5HT2C have been identified as those involved in the release of PRL. In particular, 5HT2A and 5HT2C receptors act indirectly through the stimulation of the paraventricular hypothalamic nucleus, as demonstrated by the blunted PRL secretion after the administration of m-chlorophenylpiperazine, a 5-HT2C receptor agonist, or 1-(2,5-dimethoxy-4-iodophenyl)2-aminopropane, a 5-HT2A/2C agonist, in rats with a selective lesion of the paraventricular hypothalamic nucleus [38, 39]. Conversely, PRL release after administration of a 5-HT1A receptor agonist, ipsapirone, is not affected [38, 39].
3 Prolactin in males: role in reproduction
The role of PRL in male reproduction and fertility has been studied in several animal models, leading, however, to conflicting results. PRLR is expressed in the testis of rodents and mammals [11], including men [40]. In particular, PRLR expression has been demonstrated in Leydig, Sertoli and in germ cells [41, 42]. PRL can affect steroidogenesis by modulating the expression of luteinizing hormone (LH) receptors [43, 44], or by regulating the activity of steroidogenetic enzymes [45, 46], such as 5α-reductase, 3β-HSD and 17β-HSD [47, 48]. In animal models, PRL regulates spermatogenesis as well. PRL induces the expression of follicle stimulating hormone (FSH) on Sertoli cells and stimulates the progression of germ cells from spermatocyte to spermatide morphology [49]. However, PRLR knock-out in male mice was associated with infertility, without disorders of mating behavior, only in 20–40 % of cases [50–53] and, in most cases, the homozygous deletion of PRLR was associated with delayed fertility, rather than complete infertility [52, 53]. Furthermore, PRL knockout mice do not show any impairment in fertility [54].
Taken together these data suggest an ancillary role of PRL in male fertility. This is in keeping with the hypothesis that PRL displays weak gonadotropic activity, which becomes evident when LH and FSH action is absent [55] and that is redundant when gonadotropin function is normal. In men, studies on the effects of PRL on fertility are scanty and, since mutations in PRL gene have not been described and mutations in PRLR were found only in women [56], no model of spontaneous defective PRL action is available. In men treated with GnRH analogs for prostate cancer, adding bromocriptine led to a worse impairment in semen parameters than adding cyproterone acetate [57]. Administration of exogenous PRL or the increase of endogenous PRL levels, by treating with metoclopramide, in 20 infertile men with hypoprolactinemia led to an improvement in sperm concentration and morphology [58]. A study conducted in human sperms suggested that PRL is involved in the survival of these cells, since, after incubation with PRL, their motility was preserved for a longer time and spontaneous DNA strand fragmentation was decreased [59]. However, a recent study conducted on 269 male partners of infertile couples failed to demonstrate any significant association between semen parameters and serum PRL levels [60].
PRL has a trophic effect on male seminal accessory glands. One of the earliest demonstrations comes from the evidence of an androgen-independent increase in seminal vesicles in rats and mice with hyperprolactinemia induced by the grafting of a female anterior pituitary [61, 62]. Treating mice with erocornine, an inhibitor of PRL secretion, was associated with a decrease in seminal vesicle weight [63], a result that is replicated in PRL knock-out mice [54]. PRL and its receptor are expressed in the prostate and studies in vitro and in vivo demonstrated that their regulation is androgen dependent [64]. The role of PRL in the prostate has been clarified by the use of transgenic mice with overexpression of the PRL gene, which developed a dramatic prostate hyperplasia [65]. Moreover, PRL seems to be involved also in the secretory activity of male accessory glands, in fact the increase or suppression of PRL levels has been associated with a change in seminal vesicle and prostate secretion composition in different animal models [66–68]. PRLR expression has been demonstrated in human seminal accessory glands [40, 69]. However, the clinical implication of PRL activity on the male genital tract is still unclear. PRL has been proposed as being involved in the pathogenesis of prostate benign hyperplasia and cancer [70], however, the available studies provide conflicting results and their scarceness did not allow us to draw final conclusions. Concerning seminal vesicles in men, only one study is available [60]. In this study, the seminal tract and accessory glands of 269 men from infertile couples were assessed with transrectal ultrasound before and after ejaculation [60]. Low Serum PRL levels were significantly associated with decreased semen volume, diameter of deferential ampullas, seminal vesicle volume either before and after ejaculation, and with an inhomogeneity of seminal vesicle texture, thus suggesting not only a trophic effect of PRL on the male genital tract, but also a remodeling activity [60].
4 Prolactin in males: role in sexuality
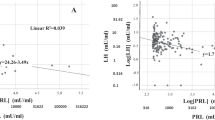
The role of PRL in male sexuality has been extensively studied with regard to its pathologically increased levels [see 14 for review]. However, recent evidence is showing a role in sexual pathology also for low PRL levels. In a study conducted on more than 2500 men consulting for sexual dysfunction, low PRL levels were associated with impaired penile blood flow, as assessed by color Doppler ultrasound [71], which is an objective index of arteriogenic erectile dysfunction (ED). Accordingly, Fig. 1 panel A shows that, in a larger population of men with sexual dysfunction, after excluding those with hyperprolactinemia or taking drugs affecting prolactinemia, men reporting moderate or severe ED have lower PRL levels. In men with sexual dysfunction, low PRL levels are also associated with the complaint of premature ejaculation (PE) (see Fig. 1 panel B and [71]). These findings on the relationship between PRL and sexual functioning were further confirmed in a later study on almost 3000 subjects, aged 40–79 years, from the general European population involved in the European Male Ageing Study (EMAS) [72]. In this study, men who reported a worsening in their sexuality during the previous year had lower PRL levels [72]. Interestingly, among the investigated domains of sexuality, low PRL was associated with a worsening in sleep- or sex-related erections and lower ejaculatory time [72]. The association of hypoprolactinemia with PE was found also in a population of infertile men, whose Premature Ejaculation Diagnostic Tool (PEDT) score was significantly higher – suggesting a higher risk of PE - in subjects with low PRL levels than in controls [60]. Ejaculation is the result of a spinal reflex arising from genital and/or brain stimulations that are integrated by the spinal ejaculatory generator, which, in turn, regulates the sympathetic, parasympathetic and motor nerve activity, thus providing the nervous outflow that leads to the ejection of seminal fluid [73]. Besides the nervous control, ejaculation is regulated also by hormones [73], as already demonstrated in a population of men consulting for sexual dysfunction, whose ejaculatory latency time was positively associated with thyroid stimulating hormone (TSH) and PRL and negatively associated with testosterone [73, 74]. Accordingly, Fig. 1 panel B shows that lower PRL, increased thyroid activity and testosterone levels are risk factors for PE, independently from each other and from other possible confounding factors.

Association between PRL and sexual symptoms. Data were derived from a consecutive series of 4159 patients (mean age 51.6 ± 13.1 years) seeking medical care at the Sexual Medicine and Andrology Unit of the Department of Experimental and Clinical Biomedical Sciences, University of Florence, Florence, Italy. All data are adjusted for age, smoking habits, alcohol consumption, waist circumference, total testosterone and TSH after excluding subjects with hyperprolactinaemia (defined as >735 mU/l or 35 ng/ml) or taking antidepressants. Abbreviations: PRL: prolactin, TSH: thyreotropin stimulating hormone
These findings are in apparent contrast with the well-known sexual inhibitory effects that are often described in hyperprolactinemia. However, it must be noted that, while the negative effect of pathologically high PRL levels on sexual desire has been consistently demonstrated [75–79], their role on erectile function and orgasm or ejaculatory disturbances is controversial [80–82]. Abnormally high PRL levels down-regulate GnRH-gonadotropin-testis axis, thus leading to hypogonadism which is per se associated with sexual dysfunctions, such as low libido and ED [83]. However, when treating hyperprolactinemic men with testosterone replacement, sexual desire is not restored [84], whereas the use of PRL-lowering drugs can improve the symptom [77, 85]. This demonstrates that PRL itself is involved in the pathogenesis of reduced libido. Conversely, the role of high PRL in ED seems not to be direct, but mediated by hypogonadism, as demonstrated by the lack of significance of the relationship between ED and hyperprolactinemia, when the association is adjusted for testosterone levels [78, 86]. Hence, these data, overall, support an effect of hypoprolactinemia, rather than hyperprolactinemia, on ED and PE.
How PRL stimulates sexual function is matter of speculation. In animal models, the intraventricular injection of PRL can stimulate sexual behavior [87]. Furthermore, studies with functional magnetic resonance in men demonstrated that brain response to erotic visual stimuli correlated with serum PRL levels [88, 89]. However, it should be taken into consideration that PRL can be a marker of different causative mechanisms, more than the direct effector of sexual stimulation. In particular, as previously mentioned, low PRL can be the result of an increased DA activity. Evidence shows that DA is involved in modulating male sexual behavior. In animal models, the systemic or intraventricular administration of the DA precursor L-DOPA or direct DA receptor agonists, such as apomorphine, is associated with the stimulation of the appetitive phase of sexual behavior, as well as with an increase in the number of ejaculations obtained and with a reduction in the number of mounts and intromissions required to achieve ejaculation [90–92]. These stimulatory effects of DA on sexual activity and ejaculation were also confirmed in castrated animals, in whom the administration of apomorphine was able to restore copulatory activity, even without testosterone replacement [93, 94]. Accordingly, the use of DA antagonists, such as haloperidol or pimozide induces a dramatic reduction in the number of ejaculations and increases ejaculatory latency time in a dose dependent manner [95–97]. Hence, an increased dopaminergic tone, as reflected by low PRL levels, can explain the association with reduced ejaculatory latency time and PE. However, it is well known that dopaminergic central neurotransmission is pivotal to inducing penile erection both in animals and in humans [see for review 98] and a hyper-activated central dopaminergic tone does not explain the association between low PRL and erectile dysfunction.
Another possible explanation for the association between hypoprolactinemia and both erectile and ejaculatory dysfunctions could be represented by a decrease in serotonergic central tone. As aforementioned, 5-HT is a PRL stimulating factor and PRL itself can reflect the central serotonergic tone. In an animal model of non-human primates (Macaca Mulata), 5-hydroxiindolacetic acid was the only monoamine whose concentration in cerebrospinal fluid correlated with the concentration of PRL in saliva [99]. Moreover, PRL response to a challenge test with fenfluramine or citalopram – a 5-HT agonist and a SSRI, respectively – has been widely used as a non-invasive index of central nervous 5-HT responsiveness in humans [100–105]. The role of 5-HT in ejaculation has been hypothesized since the 1970s when, in contrast with the widespread opinion that ejaculatory disorders were psychological diseases, several scientists started to investigate neuronal control of ejaculation. Administration in rats of lysergic acid diethylamide or quipazine, two non-selective 5-HT2C agonists, delays ejaculation [106] and a similar result is obtained by treatment with 2,5-dimethoxy-4-iodophenyl-2-aminopropane, a 5-HT2A and 5-HT2C agonist [107].
Conversely, the selective stimulation of 5-HT2A receptor with 2,5-dimethoxy-4-methyl-amphetamine does not affect ejaculatory latency time [106]. Furthermore, the stimulation of 5-HT1A receptor with a selective agonist, 8-hydroxy-2-di-n-propylaminotetralin, decreased the ejaculatory latency time and the number of intromissions required for obtaining ejaculation [106]. These observations led Waldinger [108] to formulate a neurobiological hypothesis for PE, which states that perturbation of the central 5-HT pathway, due to 5HT2C receptor hyposensitivity and/or 5-HT1A receptor hypersensitivity, might be the origin of some cases of lifelong PE [108]. This hypothesis has been proven in humans by two randomized, placebo-controlled, clinical trial which demonstrated that nefazodone, an antidepressant drug that potently antagonizes 5-HT2 receptors, and mirtazapine, an antidepressant that blocks 5-HT2C and 5-HT3 receptors, were not able to delay ejaculation in patients with PE, whereas SSRIs, such as paroxetine and sertraline were [109, 110]. Concerning erection, it has been demonstrated that different 5-HT agonists, such as fenfluramine, m-chlorophenyl-piperazine (m-CPP), quizapine and 5-hydroxytryptophan, can induce erection in rodents and non-human primates [111, 112], and DA induced erection is prevented by 5-HT antagonists [113]. The use of 5-HT receptor agonists with different affinity for the different 5-HT receptor subtypes led to the acquisition that 5-HT1C receptors mediate a stimulatory effect on erection, whereas 5-HT1A and 5-HT2 have an inhibitory action [114, 115].
From the data presented, it turns out that the emerging association between low PRL and ED or PE, confirmed in different populations, can be explained by a low central 5-HT tone that, among its manifestations, induces a reduction in serum PRL levels. Conversely, a hyperactivity of DA pathway cannot fully explain the clinical features associated with hypoprolactinemia.
5 Prolactin in males: role in metabolism
A role for PRL as a marker of metabolic disorders is also emerging. The first report was obtained by our group in a population of 2531 men who attended our Sexual Medicine and Andrology Unit at the University of Florence, seeking medical care for sexual dysfunction [71]. Among them, men with lower PRL levels had a worse metabolic profile, as demonstrated by a higher body mass index and prevalence of diabetes mellitus as well as dyslipidemia [71]. Furthermore, the study demonstrated a stepwise negative relationship between serum PRL levels and number of metabolic syndrome (MetS) components [71]. These results have now been replicated in an extended sample of 4159 men, as shown in Fig. 2. The role of low PRL as a marker of a disadvantageous metabolic profile was later confirmed in the EMAS population (almost 3000 community-dwelling men from eight European Countries), in which, beside a correlation between low PRL and obesity, hyperglycemia and increasing number of MetS components, also a relationship with physical inactivity was found [72]. More recently, in a population-based study involving more than 2000 Chinese subjects, low PRL was confirmed as a correlate of a poor glycemic profile [117]. These data have also been confirmed in a longitudinal study on a pediatric population of about 130 patients, which showed that obese children had significantly lower PRL levels as compared to their healthy counterparts and PRL levels were significantly and inversely correlated with body mass index, HOMA-IR and interleukin-6 and positively with HDL-cholesterol [118]. Furthermore, low PRL was significantly associated with incidence of MetS during one year of follow-up and, in children having MetS at baseline and who started a lifestyle intervention, hypoprolactinemia represented a negative prognostic factor for the improvement of metabolic parameters [118]. Consistently with the adverse metabolic profile, we recently demonstrated that low PRL is a predictor for the occurrence of major adverse cardiovascular events (MACE) during a mean follow-up of 4.4 years [119]. As shown in Fig. 3, the association between low PRL and forthcoming MACE is independent from other cardiovascular risk factors both conventional, such as age, smoking habits and waist circumference and unconventional and ED-specific, such as lack of partner’s sexual interest [120, 121] and reduced flaccid penile acceleration [122]. The association between low PRL and negative cardiac outcomes has also been confirmed by data from the Study of Health in Pomerania (SHIP). SHIP is a population-based study, involving almost 2000 men and 2000 women, and it showed that in men, but not in women, low PRL levels at baseline were associated with a higher incidence of left ventricular altered geometry and hypertrophy during five years of follow-up [123].

Association between PRL and metabolic parameters. Data were derived from a consecutive series of 4159 patients (mean age 51.6 ± 13.1 years) seeking medical care at the Sexual Medicine and Andrology Unit of the Department of Experimental and Clinical Biomedical Sciences, University of Florence, Florence, Italy. All data are adjusted for age, smoking habits, alcohol consumption, total testosterone and TSH after excluding subjects with hyperprolactinaemia (defined as >735 mU/l or 35 ng/ml) or taking antidepressants. Abbreviations: PRL: prolactin; IFG: impaired fasting glucose; DM: diabetes mellitus; MetS: metabolic syndrome; IDF: International Diabetes Federation [116]

Hazard ratio and 95 % confidence interval (log-scale) for incident major adverse cardiovascular events as a function of different conventional and unconventional cardiovascular risk factors. Data were derived from 1385 patients (mean age 53.1 ± 12.4 years) followed-up for a mean time of 4.4 ± 2.6 years at the Sexual Medicine and Andrology Unit of the Department of Experimental and Clinical Biomedical Sciences, University of Florence, Florence, Italy. Men with hyperprolactinaemia (defined as >735 mU/l or 35 ng/ml) or taking antidepressants were excluded from the analysis
Obesity has been previously associated with alterations in circulating PRL concentrations and with an impairment of PRL responsiveness to pharmacological stimulation [see in 124]. The PRL response to insulin-induced hypoglycemia [125], TRH stimulation [126, 127] and other stimulatory factors [128] is, in fact, diminished in obese individuals. In addition, obesity alters the 24-h spontaneous release of PRL with a generalized dampening of its release [129]. Weight reduction, with the accompanying decrease in plasma insulin levels, leads to a normalization of prolactin responses in most circumstances [130]. In animal models, the role of PRL in regulating body weight and metabolism is conflicting [131]. Buntin et al. [132] observed that prolactin strongly stimulates body weight gain in ring doves (Streptopelia risoria). Silverin [133] concluded that the body weight of prolactin treated pied flycatchers (Ficedula hypoleuca) was significantly higher than that of the control group. A study by Perez-Villamil et al. [134] indicated that PRL secretion has an effect on the increase in the body weight of mice. A very recent study demonstrated that, in male rats with high fat diet-induced obesity, PRL decreases by almost 50 % both in serum and in the anterior pituitary [135]. In healthy men, the infusion of a lipid/heparin emulsion was able to decrease the PRL response to a citalopram challenge test [136].
Animal models have suggested that PRL might be involved in stimulating insulin release and beta-cell proliferation [131, 137, 138], adipogenesis [131, 139] and lipid metabolism [131, 139]. Accordingly, PRLR deficient mice are characterized by features of incipient diabetes, with islet and β-cell hypoplasia, reduced pancreatic insulin mRNA levels, blunted insulin secretory response to glucose and glucose intolerance [140]. In addition, experimental PRL deficiency, induced by bromocriptine treatment, was found to cause an increased lipogenesis in rats [131, 141]. Treating lactating rats with bromocriptine exposes the offspring to a higher risk of obesity, dyslipidemia and insulin resistance in adulthood, increasing the risk for MetS development [142]. Only few reports have investigated the relevance of PRL in metabolic control in humans. For at least 30 years it has been known that PRL response to various stimuli is impaired in obese subjects [125, 127, 143–145]. It has been demonstrated that human adipose tissue expresses PRL and its receptor. In adipose tissue, PRL is able to reduce the bio-synthesis of adipose lipoprotein lipase and fatty acid synthase [146, 147], thus decreasing lipogenesis. Furthermore, PRL modulates the expression of several adipokines, including adiponectin, interleukin-6 and leptin [148, 149] with a consequent effect on adipose and peripheral insulin sensitivity. These observations contrast with increased the insulin-resistance, lipid levels and body weight observed in subjects with prolactinoma-induced hyperprolactinemia [150–152]. However, it should be noted that elevated PRL levels within the normal range are associated with a compensatory increase in β-cell mass and insulin secretion in response to hyperglycemia and are also associated with improved insulin sensitivity [150, 153–155]. These data, overall, are in favor of a positive metabolic role of PRL, as long as it is in the normal range, even though definitive conclusions cannot yet be drawn [156, 157]. In addition, the results of our studies themselves do not support the hypothesis of a direct effect of PRL on an adverse cardio-metabolic profile, as shown by the comparison between hyperprolactinemic men and matched controls [71, 72, 119].
Food intake as a function of energy storage in adipose tissue is regulated by several molecules, including leptin, insulin and ghrelin. DA is not primarily involved in the homeostasis of energy balance, nonetheless it takes part in the mechanisms of food intake because it is the neurotransmitter involved in reward. Sight, smell and taste can activate the reward mechanism and provide motivation for food intake with a strength which can override the homeostatic mechanisms [158]. The dopaminergic system has been extensively evaluated in the pathogenesis of obesity. In obese subjects, similarly to drug addicts, DA receptors, in particular D2 subtype, are hypo-expressed and hypo-functioning in thestriate nucleus [159–162]. Furthermore, a polymorphism of the D2 receptor, leading to significantly reduced expression [163], has been associated with a greater food reinforcement, which leads to an increased energy intake, more evident in obese subjects [164, 165]. More recently, this polymorphism has been found significantly more prevalent in subjects with type 2 diabetes mellitus than in non-diabetic ones [166]. Besides these effects on food intake, a role for DA in glucose metabolism has also been described. In particular, in hamsters, D2 receptor agonist administration demonstrated an insulin sensitizing effect [167] and, in humans, treatment with DA agonists was associated with the improvement of diabetes metabolic control [168]. In addition, the adverse effects of DA antagonists on glucose metabolism is well known [169].
Given these data, the association between low PRL and metabolic derangement cannot be explained as the consequence of an increased dopaminergic tone. Conversely, a reduced brain serotonergic tone can justify these findings. In fact, 5-HT controls several functions in the brain, including eating behavior [170]. 5-HT is mainly synthetized by the raphe nuclei but serotonergic signaling reaches all regions of the brain diffusely, including striate nucleus and hypothalamus which are the main central structures involved in feeding regulation. 5-HT2C is the most represented receptor in these areas. In rodents, infusion of 5-HT into hypothalamic nuclei results in a decreased food intake [171, 172] and in weight loss [173], whereas 5-HT neurochemical depletion leads to hyperphagia and weight gain [174–176]. Lorcaserin, a 5-HT2 agonist, is marketed in the U.S. for treatment of obesity and the short-term use of SSRI is associated with weight loss [177]. In addition to its role in food intake and weight control, an effect of 5-HT on glucose metabolism has been hypothesized, but whether its effects are mediated by a central or a peripheral 5-HT signaling is still unclear [178–184]. It has been also demonstrated that polymorphisms in the promoter of 5-HT2A and 5-HT2C receptors and 5-HT transporter are associated with a different predisposition to obesity, type 2 diabetes, coronary and carotid artery disease [185–190]. Hence, low PRL levels could be the mirror of a low serotonergic central tone and, in line with this hypothesis, an impaired PRL response to a challenge test with 5-HT agonists, such as fenfluramine or citalopram, has been associated with MetS, physical inactivity and preclinical carotid artery atherosclerosis [191–193].
6 Prolactin in males: role in psychological health
Another unexpected correlate of low PRL, emerging from our study on men with sexual dysfunction, is anxiety [71, 73], which has been further confirmed in a similar, but extended, population, as shown in Fig. 4, panel A. Accordingly, men in the lowest quartile of PRL score the highest for the SIEDY Scale 3 [Fig. 4, panel B], a validated index of psychopathology in subjects with sexual dysfunction [194]. In a large sample from the general European population (EMAS), among depressed men, serum PRL was associated with severity of depressive symptoms, as assessed by the Beck Depression Inventory [72]. In addition, men reporting a higher number of adverse life events in the previous 6 months had low PRL levels [72].

The association with PRL levels and psychological and anxiety scores. Data were derived from a consecutive series of 4159 patients (mean age 51.6 ± 13.1 years) seeking medical care at the Sexual Medicine and Andrology Unit of the Department of Experimental and Clinical Biomedical Sciences, University of Florence, Florence, Italy. Adjusted r (Adj. r) values indicate the age, smoking habits, alcohol consumption, total testosterone and thyrotropin stimulating hormone adjusted data after excluding subjects with hyperprolactinaemia (defined as >735 mU/l or 35 ng/ml) or taking antidepressants. Abbreviations: PRL: Prolactin; MHQ-A: Middlesex Hospital Questionnaire-free-floating anxiety subscale; SIEDY: Structured Interview on Erectile Dysfunction
PRL is involved in response to stress. It has been demonstrated that intraventricular administration of PRL in rats is able to reduce anxiety-related behaviors [195], prevent the stress-induced formation of gastric ulcers and shows antidepressant effects during forced swimming [196, 197]. Anxiolytic effects are produced also by intravenous administration of PRL [195] and this finding, together with the observation of an increased expression of PRLR in choroid plexus during chronic stress [198] suggests that peripheral PRL is actively and selectively up-taken by choroid plexus cells into the cerebrospinal fluid during stressful events [199]. In addition, down-regulating the PRLR expression in the brain, by antisense targeting, increases anxiety-related behaviors in rats subjected to emotional stressors [195]. According to Theorell theory [200], serum PRL levels change during different psychosocial conditions and, in particular, higher PRL levels are found when stressful events are faced with a passive coping attitude, whereas, during active coping, lower PRL levels are observed [200]. Interestingly, in the study from the EMAS population, when analyzing the single adverse life events occurring in the previous 6 months, an inverse association with serum PRL levels was found only for events which marginally affected individual wellbeing, such as having a financial crisis or problems with close friends or colleagues, and which can be overcome by positive and active coping strategies.
Although in the neurobiology of depression 5-HT and noradrenaline play a pivotal role, evidence shows that also DA have a major function [201]. In animal models of depression and chronic stress, a decreased DA concentration [202, 203] and D2/D3 receptor binding [204] is found in nucleus accumbens and these alterations are reversed by administration of tricyclic antidepressants or fluoxetine [204, 205]. Studies in humans demonstrated that the concentration of DA metabolites in cerebrospinal fluid was lower in depressed than in non-depressed subjects [206–209] and the administration of α-methylparatyrosine, an inhibitor of tyrosine hydroxylase, is associated with a relapse in depressive symptoms in patients with remission induced by norepinephrine reuptake inhibitors [210]. The responsiveness of PRL to the administration of apomorphine has been evaluated as a possible test for the assessment of central dopaminergic tone, however, while a study showed an impaired PRL suppression of apomorphine in bipolar depressed patients [211], other studies did not [212]. Clinical trials on the effectiveness of DA agonists on depression are scanty and flawed by small sample size. However, available data suggest their efficacy in improving depressive symptoms when used in monotherapy [213–217] or as adjuvants during tricyclic antidepressant therapy [218–220]. Hence, a hypo-dopaminergic tone seems to play a role in mood pathology but this would cause a trend towards increased, rather than decreased, PRL levels, thus non supporting the data on PRL and mood.
Serotonin has been extensively proven to be implicated in depression [221] and the main pharmacological strategies for depression treatment rely on drugs that enhance serotonergic tone in the brain. A large body of evidence supports the role of 5-HT in depression. Serotonin and its metabolites are low in the cerebrospinal fluid of depressive patients [222, 223] and 5-HT in the brain of depressed men who committed suicide is lower than in controls [224]. Administration of tryptophan or 5-hydroxytryptophan, which are precursors of 5-HT, in depressed patients improved their symptomatology even in the absence of a standard antidepressant therapy [225] and prevented the relapse of the disease [226]. The challenge test with fenfluramine or citalopram has been used for testing the central serotonergic tone and, in depressed men, PRL response to these 5-HT agonists was blunted [227, 228], with the severity of the clinical feature associated with the extent of the impairment in PRL response [227, 229]. Considering these data, basal low serum PRL, similarly to what is observed in challenge tests, could be a mirror of an impaired 5-HT tone in the brain and could identify a depressive phenotype.
Even in anxiety, the role of the serotonergic pathway is well known [230]. Early studies on the neurobiology of anxiety identified an over-activity of the serotonergic system as the underlying mechanism. However, further research demonstrated that different serotonergic circuitries and receptors can mediate different aspects of anxiety [102]. The simple observation that most of the drugs used in anxiety treatment are 5-HT agonists can lead to the conclusion that 5-HT is anxiolytic, however, the system is complex and only partially understood. Several studies confirmed a role of 5-HT1A receptor in the pathogenesis of anxiety [231–233] and, consistently, buspirone, a 5-HT1A agonist, as well as SSRI which, by increasing 5-HT in the synaptic cleft stimulating post-synaptic 5-HT1A receptors, can control anxiety symptoms [234]. Genetic predisposition plays an important role and polymorphisms in the sodium-dependent 5-HT transporter, which is pivotal in the regulation of the 5-HT system, have been identified as important factors conditioning variability in emotional regulation. A common repeat polymorphism in the promoter region of the gene encoding for the serotonin transporter is present. A 43-base pair deletion represents the short form of the transporter and it has been associated with less effective serotonergic function, as compared to the long allelic form [235]. Short allelic form carriers have a sharpened reactivity of the amygdala to stress, lower gray matter volume in the amygdala and medial prefrontal cortex, and an increased reactivity of hypothalamic-pituitary-adrenal axis to stress [236, 237]. In addition, as shown by a recent meta-analysis, short allelic form carriers display a greater attention to negative stimuli, a characteristic that is strongly implicated in the etiology and maintenance of anxiety disorders [238]. Furthermore, in 77 male volunteers, it has been shown that carriers of the short allele exhibit blunted PRL response to the fenfluramine challenge test [239]. Although serotonergic pathway and receptor system is complicated and only partially known, experimental and clinical data have unassailably demonstrated that a low serotonergic tone is responsible for anxiety disorders. A blunted PRL response to fenfluramine or citalopram has been used as a mirror of brain serotonergic tone. Recent studies showed an association between basal serum PRL levels and anxiety and depressive symptoms. These data can be interpreted considering that not only stimulated but also basal PRL level can mirror 5-HT cerebral tone.
7 Conclusion
Prolactin is a surprising hormone. It was initially and for a long time believed to be a hormone involved only in mammary gland development and in lactation. For this reason, studies in males are scanty and prolactin’s role in men still widely unknown. Recent and emerging data are now showing that low prolactin has a role in male reproduction, sexuality, metabolism and psychological health. Whether prolactin plays a direct role in these physiologic functions, with low prolactin causing their impairment, or it is an epiphenomenon of different mechanisms, is still matter of speculation and deserves further studies. Prolactin has been demonstrated to be a trophic factor for male reproductive tract, to have a role in improving sexual behavior and lipid and glucose metabolism as well as to have an anxiolytic effect. In this perspective, it should be reasonable to suggest maintaining prolactin within its normal range during therapy with dopaminergic agents. The possibility that low serum prolactin is a mirror of central serotonergic activity is intriguing. Serotonin is involved in several functions, including mood, fear, stress reaction, eating, locomotion, sleep, thermogenesis, cardiovascular control and insulin secretion. Serotonin central hypo-activity could explain the emerging evidence on the clinical correlates of low prolactin levels in men.
References
Burrows HL, Birkmeier TS, Seasholtz AF, Camper SA. Targeted ablation of cells in the pituitary primordia of transgenic mice. Mol Endocrinol. 1996;10:1467–77.
Ben-Jonathan N, Mershon JL, Allen DL, Steinmetz RW. Extrapituitary prolactin: distribution, regulation, functions, and clinical aspects. Endocr Rev. 1996;17:639–69.
Nagy E, Berczi I. Hypophysectomized rats depend on residual prolactin for survival. Endocrinology. 1991;128:2776–84.
Kelly PA, Djiane J, Postel-Vinay MC, Edery M. The prolactin/growth hormone receptor family. Endocr Rev. 1991;12:235–51.
Goffin V, Kelly PA. The prolactin/growth hormone receptor family: structure/function relationships. J Mammary Gland Biol Neopl. 1997;2:7–17.
Goffin V, Ferrag F, Kelly PA. Molecular aspects of prolactin and growth hormone receptors. In: LeRoith D, editor. Advances in molecular and cellular endocrinology, vol. 2. Amsterdam: Elsevier Science; 1998. p. 1–331998.
Postel-Vinay MC, Belair L, Kayser C, Kelly PA, Djiane J. Identification of prolactin and growth hormone binding proteins in milk. Proc Natl Acad Sci USA. 1991;188:6687–90.
Fuh G, Wells JA. Prolactin receptor antagonists that inhibit the growth of breast cancer cell lines. J Biol Chem. 1995;270:13133–7.
Amit T, Dibner C, Barkey RJ. Characterization of prolactin and growth hormone-binding proteins in milk and their diversity among species. Mol Cell Endocrinol. 1997;130:167–80.
Goffin V, Shiverick KT, Kelly PA, Martial JA. Sequence function relationships within the expanding family of prolactin, growth hormone, placental lactogen and related proteins in mammals. Endocr Rev. 1996;17:385–410.
Bole-Feysot C, Goffin V, Edery M, Binart N, Kelly PA. Prolactin (PRL) and its receptor: actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. Endocr Rev. 1998;19:225–68.
Feldman M, Ruan W, Cunningham BC, Wells JA, Kleinberg DL. Evidence that the growth hormone receptor mediates differentiation and development of the mammary gland. Endocrinology. 1993;133:1602–8.
Gettler LT, McDade TW, Feranil AB, Kuzawa CW. Prolactin, fatherhood, and reproductive behavior in human males. Am J Phys Anthropol. 2012;148:362–70.
Maggi M, Buvat J, Corona G, Guay A, Torres LO. Hormonal causes of male sexual dysfunctions and their management (hyperprolactinemia, thyroid disorders, GH disorders, and DHEA). J Sex Med. 2013;10:661–77.
Freeman ME, Kanyicska B, Lerant A, Nagy G. Prolactin: structure, function, and regulation of secretion. Physiol Rev. 2000;80:1523–631.
Matsumoto M, Hidaka K, Tada S, Tasaki Y, Yamaguchi T. Full-length cDNA cloning and distribution of human dopamine D4 receptor. Brain Res Mol Brain Res. 1995;29:157–62.
Valerio A, Alberici A, Tinti C, Spano P, Memo M. Antisense strategy unravels a dopamine receptor distinct from the D2 subtype, uncoupled with adenylyl cyclase, inhibiting prolactin release from rat pituitary cells. J Neurochem. 1994;62:1260–6.
Lledo PM, Legendre P, Israel JM, Vincent JD. Dopamine inhibits two characterized voltage-dependent calcium currents in identified rat lactotroph cells. Endocrinology. 1990;127:990–1001.
Elsholtz HP, Lew AM, Albert PR, Sundmark VC. Inhibitory control of prolactin and pit-1 gene promoters by dopamine. Dual signaling pathways required for D2 receptor-regulated expression of the prolactin gene. J Biol Chem. 1991;266:22919–25.
Ishida M, Mitsui T, Yamakawa K, Sugiyama N, Takahashi W, Shimura H, Endo T, Kobayashi T, Arita J. Involvement of cAMP response element-binding protein in the regulation of cell proliferation and the prolactin promoter of lactotrophs in primary culture. Am J Physiol Endocrinol Metab. 2007;293:E1529–37.
Demarest K, Riegle G, Moore K. Prolactin-induced activation of tuberoinfundibular dopaminergic neurons: evidence for both a rapid “tonic” and a delayed “induction” component. Neuroendocrinology. 1984;38:467–75.
Gudelsky GA, Porter JC. Release of dopamine from tuberoinfundibular neurons into pituitary stalk blood after prolactin or haloperidol administration. Endocrinology. 1980;106:526–9.
Demarest KT, McKay DW, Riegle GD, Moore KE. Sexual differences in tuberoinfundibular dopamine nerve activity induced by neonatal androgen exposure. Neuroendocrinology. 1981;32:108–13.
Kamberi IA, Mical RS, Porter JC. Effects of melatonin and serotonin on the release of FSH and prolactin. Endocrinology. 1971;88:1288–93.
Clemens JA, Sawyer BD, Cerimele B. Further evidence that serotonin is a neurotransmitter involved in the control of prolactin secretion. Endocrinology. 1977;100:692–8.
Kato Y, Nakai Y, Imura H, Chihara K, Ogo S. Effect of 5-hydroxytryptophan (5-HTP) on plasma prolactin levels in man. J Clin Endocrinol Metab. 1974;38:695–7.
Chen HJ, Meites J. Effects of biogenic amines and TRH on release of prolactin and TSH in the rat. Endocrinology. 1975;96:10–4.
Kordon C, Blake CA, Terkel J, Sawyer CH. Participation of serotonin-containing neurons in the suckling-induced rise in plasma prolactin levels in lactating rats. Neuroendocrinology. 1973-1974;13:213–23.
Gallo RV, Rabii J, Moberg GP. Effect of methysergide, a blocker of serotonin receptors, on plasma prolactin levels in lactating and ovariectomized rats. Endocrinology. 1975;97:1096–105.
Lawson DM, Gala RR. The influence of pharmacological manipulation of serotonergic and dopaminergic mechanisms on plasma prolactin in ovariectomized, estrogen-treated rats. Endocrinology. 1978;102:973–81.
Fessler RG, Deyo SN, Meltzer HY, Miller RJ. Evidence that the medial and dorsal raphe nuclei mediate serotonergically-induced increases in prolactin release from the pituitary. Brain Res. 1984;299:231–7.
Van de Kar LD, Bethea CL. Pharmacological evidence that serotonergic stimulation of prolactin secretion is mediated via the dorsal raphe nucleus. Neuroendocrinology. 1982;35:225–30.
Calogero AE, Bagdy G, Burrello N, Polosa P, D’Agata R. Role for serotonin3 receptors in the control of adrenocorticotropic hormone release from rat pituitary cell cultures. Eur J Endocrinol. 1995;133:251–4.
Calogero AE, Bagdy G, Moncada ML, D’Agata R. Effect of selective serotonin agonists on basal, corticotrophin-releasing hormone- and vasopressin-induced ACTH release in vitro from rat pituitary cells. J Endocrinol. 1993;136:381–7.
Lamberts SW, MacLeod RM. The interaction of the serotonergic and dopaminergic systems on prolactin secretion in the rat. The mechanism of action of the “specific” serotonin receptor antagonist, methysergide. Endocrinology. 1978;103:287–95.
Lamberts SW, MacLeod RM. Metergoline and other peripheral serotonin antagonists inhibit prolactin secretion through mechanisms unrelated to serotonin. Proc Soc Exp Biol Med. 1979;162:75–9.
Delitala G, Yeo T, Stubbs WA, Jones A. Besser GM. In: Cattibeni G, Racagni G, Spano PF, Costa E, editors. Long term effects of neuroleptics. New York: Raven; 1979. p. 443–4.
Bagdy G, Makara GB. Hypothalamic paraventricular nucleus lesions differentially affect serotonin-1A (5-HT1A) and 5-HT2 receptor agonist-induced oxytocin, prolactin, and corticosterone responses. Endocrinology. 1994;134:1127–31.
Bagdy G, Makara GB. Paraventricular nucleus controls 5-HT2C receptor-mediated corticosterone and prolactin but not oxytocin and penile erection responses. Eur J Pharmacol. 1995;275:301–5.
Hair WM, Gubbay O, Jabbour HN, Lincoln GA. Prolactin receptor expression in human testis and accessory tissues: localization and function. Mol Hum Reprod. 2002;8:606–11.
Jabbour HN, Lincoln GA. Prolactin receptor expression in the testis of the ram: localisation, functional activation and the influence of gonadotrophins. Mol Cell Endocrinol. 1999;148:151–61.
Ishida M, Yoshida M, Fukuta S, Uemura K, Iijima M, Horiguchi K, Harigaya T. Analysis of prolactin gene expression and cleaved prolactin variants in the mouse testis and spermatozoa. J Reprod Dev. 2010;56:567–74.
Takase M, Tsutsui K, Kawashima S. Effects of prolactin and bromocryptine on the regulation of testicular luteinizing hormone receptors in mice. J Exp Zool. 1990;256:200–9.
Dombrowicz D, Sente B, Closset J, Hennen G. Dose-dependent effects of human prolactin on the immature hypophysectomized rat testis. Endocrinology. 1992;130:695–700.
Chandrashekar V, Bartke A. Influence of endogenous prolactin on the luteinizing hormone stimulation of testicular steroidogenesis and the role of prolactin in adult male rats. Steroids. 1988;51:559–76.
Rubin RT, Poland RE, Tower BB. Prolactin-related testosterone secretion in normal adult men. J Clin Endocrinol Metab. 1976;42:112–6.
Takeyama M, Nagareda T, Takatsuka D, Namiki M, Koizumi K, Aono T, Matsumoto K. Stimulatory effect of prolactin on luteinizing hormone-induced testicular 5 alpha-reductase activity in hypophysectomized adult rats. Endocrinology. 1986;118:2268–75.
Gunasekar PG, Kumaran B, Govindarajulu P. Prolactin and leydig cell steroidogenic enzymes in the bonnet monkey (macaca radiata). Int J Androl. 1988;11:53–9.
Gunasekar PG, Kumaran B, Govindarajulu P. Role of prolactin on leydig, sertoli and germ cellular neutral lipids in bonnet monkeys, macaca radiata. Endocrinologia Japonica. 1991;38:1–8.
Grosdemouge I, Bachelot A, Lucas A, Baran N, Kelly PA, Binart N. Effects of deletion of the prolactin receptor on ovarian gene expression. Reprod Biol Endocrinol. 2003;1:12–27.
Binart N, Melaine N, Pineau C, Kercret H, Touzalin AM, Imbert-Bolloré P, Kelly PA, Jégou B. Male reproductive function is not affected in prolactin receptor-deficient mice. Endocrinology. 2003;144:3779–82.
Robertson FG, Harris J, Naylor MJ, Oakes SR, Kindblom J, Dillner K, Wennbo H, Törnell J, Kelly PA, Green J, Ormandy CJ. Prostate development and carcinogenesis in prolactin receptor knockout mice. Endocrinology. 2003;144:3196–205.
Ormandy CJ, Camus A, Barra J, Damotte D, Lucas B, Buteau H, Edery M, Brousse N, Babinet C, Binart N, Kelly PA. Null mutation of the prolactin receptor gene produces multiple reproductive defects in the mouse. Genes Dev. 1997;11:167–78.
Steger RW, Chandrashekar V, Zhao W, Bartke A, Horseman ND. Neuroendocrine and reproductive functions in male mice with targeted disruption of the prolactin gene. Endocrinology. 1998;139:3691–5.
Lincoln GA, Clarke IJ, Sweeney T. ‘Hamster-like’ cycles in testicular size in the absence of gonadotrophin secretion in HPD rams exposed to long-term changes in photoperiod and treatment with melatonin. J Neuroendocrinol. 1996;8:855–66.
Newey PJ, Gorvin CM, Cleland SJ, Willberg CB, Bridge M, Azharuddin M, Drummond RS, van der Merwe PA, Klenerman P, Bountra C, Thakker RV. Mutant prolactin receptor and familial hyperprolactinemia. N Engl J Med. 2013;369:2012–20.
Huhtaniemi I, Parvinen M, Venho P, Rannikko S. Combination of a GnRH agonist with an antiandrogen or bromocriptine in the treatment of prostatic cancer; slight potentiation of antigonadal effects. Int J Androl. 1991;14:374–86.
Ufearo CS, Orisakwe OE. Restoration of normal sperm characteristics in hypoprolactineamic infertile men treated with metaclopromide and exogenous prolactin. Clin. Pharmacol. Therapeut. 1995;58:354–9.
Pujianto DA, Curry BJ, Aitken RJ. Prolactin exerts a prosurvival effect on human spermatozoa via mechanisms that involve the stimulation of Akt phosphorylation and suppression of caspase activation and capacitation. Endocrinology. 2010;151:1269–79.
Lotti F, Corona G, Maseroli E, Rossi M, Silverii A, Degl’innocenti S, Rastrelli G, Forti G, Maggi M. Clinical implications of measuring prolactin levels in males of infertile couples. Andrology. 2013;1:764–71.
Negro-Vilar A, Saad WA, McCann SM. Evidence for a role of prolactin in prostate and seminal vesicle growth in immature male rats. Endocrinology. 1977;100:729–37.
Bartke A, Smith MS, Michael SD, Peron FG, Dalterio S. Effects of experimentally-induced chronic hyperprolactinemia on testosterone and gonadotropin levels in male rats and mice. Endocrinology. 1977;100:182–6.
Bartke A Effects of inhibitors of pituitary prolactin release on testicular cholesterol stores, seminal vesicles weight, fertility, and lactation in mice. Biol Reprod. 1974;11:319–25.
Nevalainen MT, Valve EM, Ahonen T, Yagi A, Paranko J, Härkönen PL. Androgen-dependent expression of prolactin in rat prostate epithelium in vivo and in organ culture. FASEB J. 1997;11:1297–307.
Wennbo H, Kindblom J, Isaksson OG, Tornell J. Transgenic mice overexpressing the prolactin gene develop dramatic enlargement of the prostate gland. Endocrinology. 1997;138:4410–5.
Ravault JP, Courot M, Garnier D, Pelletier J, Terqui M. Effect of 2-bromo-α-ergocryptine (CB154) on plasma prolactin, LH and testosterone levels, accessory reproductive glands and spermatogenesis in lambs during puberty. Biol Reprod. 1977;17:192–7.
Arunakaran J, Balasubramanian K, Srinivasan N, Aruldhas MM, Govindarajulu P. Effects of androgens, prolactin and bromocryptine on seminal vesicular enzymes of the pyruvate malate cycle involved in lipogenesis in castrated mature monkeys, macaca radiata. Int J Androl. 1988;11:133–9.
Nicoll C Ontogeny and evolution of prolactins' function. Fed Proc. 1980;39:2563–6.
Nevalainen MT, Valve EM, Ingleton PM, Nurmi M, Martikainen PM, Harkonen PL. Prolactin and prolactin receptors are expressed and functioning in human prostate. J Clin Invest. 1997b;99:618–27.
Jacobson EM, Hugo ER, Borcherding DC, Ben-Jonathan N. Prolactin in breast and prostate cancer: molecular and genetic perspectives. Discov Med. 2011;11:315–24.
Corona G, Mannucci E, Jannini EA, Lotti F, Ricca V, Monami M, Boddi V, Bandini E, Balercia G, Forti G, Maggi M. Hypoprolactinemia: a new clinical syndrome in patients with sexual dysfunction. J Sex Med. 2009;6:1457–66.
Corona G, Wu FC, Rastrelli G, Lee DM, Forti G, O’Connor DB, O’Neill TW, Pendleton N, Bartfai G, Boonen S, Casanueva FF, Finn JD, Huhtaniemi IT, Kula K, Punab M, Vanderschueren D, Rutter MK, Maggi M, EMAS Study Group. Low prolactin is associated with sexual dysfunction and psychological or metabolic disturbances in middle-aged and elderly men: the European male aging study (EMAS). J Sex Med. 2014;11:240–53.
Corona G, Jannini EA, Vignozzi L, Rastrelli G, Maggi M. The hormonal control of ejaculation. Nat Rev Urol. 2012;9:508–19.
Corona G, Jannini EA, Lotti F, Boddi V, De Vita G, Forti G, Lenzi A, Mannucci E, Maggi M. Premature and delayed ejaculation: two ends of a single continuum influenced by hormonal milieu. Int J Androl. 2011;34:41–8.
Melmed S, Casanueva FF, Hoffman AR, Kleinberg DL, Montori VM, Schlechte JA. Wass JA; endocrine society. Diagnosis and treatment of hyperprolactinemia: an endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2011;96:273–88.
Buvat J Hyperprolactinemia and sexual function in men: a short review. Int J Impot Res. 2003;15:373–7.
Ciccarelli A, Guerra E, De Rosa M, Milone F, Zarrilli S, Lombardi G, Colao A. PRL secreting adenomas in male patients. Pituitary. 2005;8:39–42.
Corona G, Mannucci E, Fisher AD, Lotti F, Ricca V, Balercia G, Petrone L, Forti G, Maggi M. Effect of hyperprolactinemia in male patients consulting for sexual dysfunction. J Sex Med. 2007;4:1485–93.
Corona G, Rastrelli G, Ricca V, Jannini EA, Vignozzi L, Monami M, Sforza A, Forti G, Mannucci E, Maggi M. Risk factors associated with primary and secondary reduced libido in male patients with sexual dysfunction. J Sex Med. 2013;10:1074–89.
Nair S, Milsom S. Anejaculation as the presenting feature of pituitary microadenoma. Fertil Steril. 2012;2008(90):e21–3.
Ishikawa H, Kaneko S, Ohashi M, Nakagawa K, Hata M. Retrograde ejaculation accompanying hyperprolactinemia. Arch Androl. 1993;30:153–5.
Corona G, Mannucci E, Petrone L, Fisher AD, Balercia G, de Scisciolo G, Pizzocaro A, Giommi R, Chiarini V, Forti G, Maggi M. Psychobiological correlates of delayed ejaculation in male patients with sexual dysfunctions. J Androl. 2006;27:453–8.
Rastrelli G, Carter EL, Ahern T, Finn JD, Antonio L, O’Neill TW, Bartfai G, Casanueva FF, Forti G, Keevil B, Maggi M, Giwercman A, Han TS, Huhtaniemi IT, Kula K, Lean ME, Pendleton N, Punab M, Vanderschueren D, Wu FC, EMAS Study Group. Development of and recovery from secondary hypogonadism in aging men: prospective results from the EMAS. J Clin Endocrinol Metab. 2015;100:3172–82.
Carter JN, Tyson JE, Tolis G, Van Vliet S, Faiman C, Friesen HG. Prolactin-screening tumours and hypogonadism in 22 men. N Engl J Med. 1978;299:847–52.
Prescott RW, Johnston DG, Kendall-Taylor P, Crombie A, Hall K, McGregor A, Hall R. Hyperprolactinemia in men-response to bromocriptine therapy. Lancet. 1982;1:245–8.
Buvat J, Maggi M, Gooren L, Guay AT, Kaufman J, Morgentaler A, Schulman C, Tan HM, Torres LO, Yassin A, Zitzmann M. Endocrine aspects of male sexual dysfunctions. J Sex Med. 2010;7:1627–56.
Cruz-Casallas PE, Nasello AG, Hucke EE, Felicio LF. Dual modulation of male sexual behavior in rats by central prolactin: relationship with in vivo striatal dopaminergic activity. Psychoneuroendocrinology. 1999;24:681–93.
Seo Y, Jeong B, Kim JW, Choi J. Plasma concentration of prolactin, testosterone might be associated with brain response to visual erotic stimuli in healthy heterosexual males. Psychiatry Investig. 2009;6:194–203.
Redouté J, Stoléru S, Grégoire MC, Costes N, Cinotti L, Lavenne F, Le Bars D, Forest MG, Pujol JF. Brain processing of visual sexual stimuli in human males. Hum Brain Mapp. 2000;11:162–77.
Paglietti E, Peliegrini-Quarantotti B, Mereu G, Gessa GL. Apomorphine and L-DOPA lower ejaculation threshold in the male rat. Physiol Behav. 1978;20:559–62.
Tagliamonte A, Fratta W, Del Fiacco M, Gessa GL. Possible stimulatory role of brain dopamine in the copulatory behavior of male rats. Pharmacol Biochem Behav. 1974;2:257–60.
Tagliamonte A, Fratta W, Gessa GL. Aphrodisiac effect of L-DOPA and apomorphine in male sexually sluggish rats. Experientia. 1974;30:381–2.
Malmnas CO. The significance of dopamine vs. other catecholamines for L-DOPA-induced facilitation of sexual behavior in the castrated male rat. Pharmacol Biochem Behav. 1976;4:521–6.
Malmnas CO. Dopaminergic reversal of the decline after castration of rat copulatory behaviour. J Endocr. 1977;73:187–8.
Napoli-Farris L, Fratta W, Gessa GL. Stimulation of dopamine autoreceptors elicits "premature ejaculation" in rats. Pharmacol Biochem Behav. 1984;20:69–72.
Pfaus JG, Phillips AA. Differential effects of dopamine receptor antagonists on the sexual behavior of male rats. Psychopharmacology. 1989;98:363–8.
Pfaus JG, Phillips AG. Role of dopamine in anticipatory and consummatory aspects of sexual behavior in the male rat: I. Effects of systemic administration of dopamine antagonists. Behav Neurosci. 1991;105:727–43.
Argiolas A, Melis MR. Neuromodulation of penile erection: an overview of the role of neurotransmitters and neuropeptides. Prog Neurobiol. 1995;47:235–55.
Lindell SG, Suomi SJ, Shoaf S, Linnoila M, Higley JD. Salivary prolactin as a marker for central serotonin turnover. Biol Psychiatry. 1999;46:568–72.
Flory JD, Manuck SB, Perel JM, Muldoon MF. A comparison of d, l-fenfluramine and citalopram challenges in healthy adults. Psychopharmacology. 2004;174:376–80.
Murphy DL, Mellow AM, Sunderland T, Aulakh CS, Lawlor BL, Zohar J. Strategies for the study of serotonin in humans. In: Coccaro EF, Murphy DL editors. Serotonin in major psychiatric disorders. Washington D.C.: American Psychiatric Press; 1990. pp 2–25
Yatham LN, Steiner M. Neuroendocrine probes of serotonergic function: a critical review. Life Sci. 1993;53:447–63.
Seifritz E, Baumann P, Muller MJ, Annen O, Amey M, Hemmeter U, Hatzinger M, Chardon F, Holsboer-Trachsler E. Neuroendocrine effects of a 20-mg citalopram infusion in healthy males. Neuropsychopharmacology. 1996;14:253–63.
Attenburrow MJ, Mitter PR, Whale R, Terao T, Cowen PJ. Low-dose citalopram as a 5-HT neuroendocrine probe. Psychopharmacology. 2001;155:323–6.
Lotrich FE, Bies R, Muldoon MF, Manuck SB, Smith GS, Pollock BG. Neuroendocrine response to intravenous citalopram in healthy control subjects: pharmacokinetic influences. Psychopharmacology. 2005;178:268–75.
Ahlenius S, Larsson K, Svensson L, Hjorth S, Carlsson A, Lindberg P, Wikström H, Sanchez D, Arvidsson LE, Hacksell U, Nilsson JL. Effects of a new type of 5-HT receptor agonist on male rat sexual behaviour. Pharmacol Biochem Behav. 1981;15:785–92.
Foreman MM, Hall JL, Love RL. The role of the 5-HT2 receptor in the regulation of sexual performance of male rats. Life Sci. 1989;45:1263–70.
Waldinger MD. The neurobiological approach to premature ejaculation. J Urol. 2002;168:2359–67.
Waldinger MD, Zwinderman AH, Olivier B. Antidepressants and ejaculation: a double-blind, randomized, placebo-controlled, fixed-dose study with paroxetine, sertraline, and nefazodone. J Clin Psychopharmacol. 2001;21:293–7.
Waldinger MD, Zwinderman AH, Olivier B. Antidepressants and ejaculation: a double-blind, randomized, fixed-dose study with mirtazapine and paroxetine. J Clin Psychopharmacol. 2003;23:467–70.
Baraldi M, Benassi-Benelli A, Lolli M. Penile erections in rats after fenfluramine admimstration. Rio Farmacoi Ter. 1977;8:375–9.
Szele FG, Murphy DL, Garrick NA. Effects of fenfluramine, m-chlorophenylpiperazine. And other serotonin related agonists and antagonists on penile erection in nonhuman primates. Life Sci. 1988;43:1297–304.
Berendsen HHG, Broekkamp CLE. Drug-induced penile erection in rats: indications of serotoninla receptor mediation. Eur J Pharmac. 1987;135:279–87.
Berendsen HHG, Jenck F, Broekkamp CLE. Involvement of 5-HT,c-receptors in drug-induced penile erections in rats. Psychopharmacology. 1990;101:57–61.
Pomerantz SM, Hepner BC, Wertz M. Serotoninergic mfluences on male sexual behavior of rhesus monkeys: Effect of serotonin agonists. Psychophnrmaco1ogy 1993;111:47–54
Alberti KG, Zimmet P, Shaw J. Metabolic syndrome–a new world-wide definition. A consensus statement from the international diabetes federation. Diabet Med. 2006;23:469–80.
Wang T, Lu J, Xu Y, Li M, Sun J, Zhang J, Xu B, Xu M, Chen Y, Bi Y, Wang W, Ning G. Circulating prolactin associates with diabetes and impaired glucose regulation: a population-based study. Diabetes Care. 2013;36:1974–80.
Chirico V, Cannavò S, Lacquaniti A, Salpietro V, Mandolfino M, Romeo PD, Cotta O, Munafò C, Giorgianni G, Salpietro C, Arrigo T. Prolactin in obese children: a bridge between inflammation and metabolic-endocrine dysfunction. Clin Endocrinol. 2013;79:537–44.
Corona G, Rastrelli G, Boddi V, Monami M, Melani C, Balzi D, Sforza A, Forti G, Mannucci E, Maggi M. Prolactin levels independently predict major cardiovascular events in patients with erectile dysfunction. Int J Androl. 2011;34:217–24.
Corona G, Monami M, Boddi V, Cameron-Smith M, Lotti F, de Vita G, Melani C, Balzi D, Sforza A, Forti G, Mannucci E, Maggi M. Male sexuality and cardiovascular risk. A cohort study in patients with erectile dysfunction. J Sex Med. 2010;7:1918–27.
Rastrelli G, Corona G, Fisher AD, Silverii A, Mannucci E, Maggi M. Two unconventional risk factors for major adverse cardiovascular events in subjects with sexual dysfunction: low education and reported partner’s hypoactive sexual desire in comparison with conventional risk factors. J Sex Med. 2012;9:3227–38.
Rastrelli G, Corona G, Lotti F, Aversa A, Bartolini M, Mancini M, Mannucci E, Maggi M. Flaccid penile acceleration as a marker of cardiovascular risk in men without classical risk factors. J Sex Med. 2014;11:173–86.
Haring R, Völzke H, Vasan RS, Felix SB, Nauck M, Dörr M, Wallaschofski H. Sex-specific associations of serum prolactin concentrations with cardiac remodeling: longitudinal results from the study of health Pomerania (SHIP). Atherosclerosis. 2012;221:570–6.
Smith SR. The endocrinology of obesity. Endocrinol Metab Clin N Am. 1996;25:921–42.
Weaver JU, Noonan K, Kopelman PG. An association between hypothalamic-pituitary dysfunction and peripheral endocrine function in extreme obesity. Clin Endocrinol (Oxford). 1991;35:97–102.
Donders SH, Pieters GF, Heevel JG, Ross HA, Smals AG, Kloppenborg PW. Disparity of thyrotropin (TSH) and prolactin responses to TSH-releasing hormone in obesity. J Clin Endocrinol Metab. 1985;61:56–9.
Lala VR, Ray A, Jamias P, Te D, Orteza N, Fiscina B, Noto R. Prolactin and thyroid status in prepubertal children with mild to moderate obesity. J Am Coll Nutr. 1988;7:361–6.
Röjdmark S, Rössner S. Decreased dopaminergic control of prolactin secretion in male obesity: normalization by fasting. Metabolism. 1991;40:191–5.
Copinschi G, De Laet MH, Brion JP, Leclercq R, L’Hermite M, Robyn C, Virasoro E, Van Cauter E. Simultaneous study of cortisol, growth hormone and prolactin nyctohemeral variations in normal and obese subjects. Influence of prolonged fasting in obesity. Clin Endocrinol. 1978;9:15–26.
Kopelman PG. Physiopathology of prolactin secretion in obesity. Int J Obes Relat Metab Disord. 2000;24(Suppl 2):S104–8.
Ben-Jonathan N, LaPensee CR, LaPensee EW. What can we learn from rodents about prolactin in Humans? Endocr Rev. 2008;29:1–41.
Strader AD, Buntin JD. Changes in agouti-related peptide during the ring dove breeding cycle in relation to prolactin and parental hyperphagia. J Neuroendocrinol. 2003;15:1046–53.
Silverin B, Goldsmith AR. Plasma prolactin concentrations in breeding pied flycatchers (ficedula hypoleuca) with an experimentally prolonged brooding period. Horm Behav. 1990;24:104–13.
Pérez-Villamil B, Bordiú E, Puente-Cueva M. Involvement of physiological prolactin levels in growth and prolactin receptor content of prostate glands and testes in developing male rats. J Endocrinol. 1992;132:449–59.
Lemini M, Ruiz-Herrera X, Ledesma-Colunga MG, Díaz-Lezama N, De Los Ríos EA, López-Barrera F, Méndez I. Martínez de la escalera G, macotela Y, Clapp C. Prolactin anterior pituitary expression and circulating levels are reduced in obese and diabetic rats: role of TGF-β and TNF-α. Am J Physiol Regul Integr Comp Physiol. 2015;308:R792–9.
Sondermeijer BM, Klein Twennaar CF, Kastelein JJ, Franssen EJ, Hutten BA, Dallinga-Thie GM, Stroes ES, Fliers E, Twickler MT, Serlie MJ. Infusion of a lipid emulsion in healthy men decreases the serotonergic response. Neuroendocrinology. 2012;95:325–31.
Arumugam R, Horowitz E, Noland RC, Lu D, Fleenor D, Freemark M. Regulation of islet beta-cell pyruvate metabolism: interactions of prolactin, glucose, and dexamethasone. Endocrinology. 2010;15:3074–83.
Yamamoto T, Mita A, Ricordi C, Messinger S, Miki A, Sakuma Y, Timoneri F, Barker S, Fornoni A, Molano RD, Inverardi L, Pileggi A, Ichii H. Prolactin supplementation to culture medium improves beta-cell survival. Transplantation. 2010;89:1328–35.
Brandebourg TD, Bown JL, Ben JN. Prolactin upregulates its receptors and inhibits lipolysis and leptin release in male rat adipose tissue. Biochem Biophys Res Commun. 2007;357:408–13.
Freemark M, Avril I, Fleenor D, Driscoll P, Petro A, Opara E, Kendall W, Oden J, Bridges S, Binart N, Breant B, Kelly PA. Targeted deletion of the PRL receptor: effects on islet development, insulin production, and glucose tolerance. Endocrinology. 2002;143:1378–85.
Flint DJ, Clegg RA, Vernon RG. Prolactin and the regulation of adipose-tissue metabolism during lactation in rats. Mol Cel Endocrinol. 1981;22:265–75.
de Moura EG, Bonomo IT, Nogueira-Neto JF, de Oliveira E, Trevenzoli IH, Reis AM, Passos MC. Lisboa PC maternal prolactin inhibition during lactation programs for metabolic syndrome in adult progeny. J Physiol. 2009;587:4919–29.
Kopelman PG, Pilkington TR, Jeffcoate SL, White N. Persistence of defective hypothalamic control of prolactin secretion in some obese women after weight reduction. Br Med J. 1980;281:358–9.
Kopelman PG, White N, Pilkington TR, Jeffcoate SL. Impaired hypothalamic control of prolactin secretion in massive obesity. Lancet. 1979;1:747–50.
Plewe G, Schneider U, Krause U, Beyer J. Naloxone increases the response of growth hormone and prolactin to stimuli in obese humans. J Endocrinol Investig. 1987;10:137–41.
Ling C, Svensson L, Odén B, Weijdegård B, Edén B, Edén S, Billig H. Identification of functional prolactin (PRL) receptor gene expression: PRL inhibits lipoprotein lipase activity in human white adipose tissue. J Clin Endocrinol Metab. 2003;88:1804–8.
Hogan JC, Stephens JM. The regulation of fatty acid synthase by STAT5A. Diabetes. 2005;54:1968–75.
Nilsson L, Binart N, Bohlooly YM, Bramnert M, Egecioglu E, Kindblom J, Kelly PA, Kopchick JJ, Ormandy CJ, Ling C, Billig H. Prolactin and growth hormone regulate adiponectin secretion and receptor expression in adipose tissue. Biochem Biophys Res Commun. 2005;331:1120–6.
Hugo ER, Brandebourg TD, Comstock CE, Gersin KS, Sussman JJ, Ben-Jonathan N. LS14: a novel human adipocyte cell line that produces prolactin. Endocrinology. 2006;147:306–13.
Doknic M, Pekic S, Zarkovic M, Medic-Stojanoska M, Dieguez C, Casanueva F, Popovic V. Dopaminergic tone and obesity: an insight from prolactinomas treated with bromocriptine. Eur J Endocrinol. 2002;147:77–84.
Berinder K, Nyström T, Höybye C, Hall K, Hulting AL. Insulin sensitivity and lipid profile in prolactinoma patients before and after normalization of prolactin by dopamine agonist therapy. Pituitary. 2011;14:199–207.
Galluzzi F, Salti R, Stagi S, La Cauza F, Chiarelli F. Reversible weight gain and prolactin levelsdlong-term follow-up in childhood. J Pediatr Endocrinol Metab. 2005;18:921–4.
Park S, Kim S, Daily JW, Kim SH. Serum prolactin concentrations determine whether they improve or impair b-cell function and insulin sensitivity in diabetic rats. Diabetes Metab Res Rev. 2011;27:564–74.
Park S, Kang S, Lee HW, Ko BS. Central prolactin modulates insulin sensitivity and insulin secretion in diabetic rats. Neuroendocrinology. 2012;95:332–43.
Lyons DJ, Hellysaz A, Broberger C. Prolactin regulates tuberoinfundibular dopamine neuron discharge pattern: novel feedback control mechanisms in the lactotrophic axis. J Neurosci. 2012;32:8074–83.
Vigas M, Klimes I, Jurcovicová J, Jezová D. Acute elevation of endogenous prolactin does not influence glucose homeostasis in healthy men. Physiol Res. 1993;42:341–5.
Katz EJ, Donald RA, Beaven DW, Espiner EA. Lack of effect of hyperprolactinemia on glucose disposal and insulin secretion in patients with prolactinomas. Horm Metab Res. 1991;13:667–9.
Baik JH. Dopamine signaling in food addiction: role of dopamine D2 receptors. BMB Rep. 2013;46:519–26.
Stice E, Yokum S, Zald D, Dagher A. Dopamine-based reward circuitry responsivity, genetics, and overeating. Curr Top Behav Neurosci. 2011;6:81–93.
Salamone JD, Correa M. Dopamine and food addiction: lexicon badly needed. Biol Psychiatry. 2013;73:e15–24.
Wang GJ, Volkow ND, Thanos PK, Fowler JS. Imaging of brain dopamine pathways: implications for understanding obesity. J Addict Med. 2009;3:8–18.
Volkow ND, Fowler JS, Wang GJ, Baler R, Telang F. Imaging dopamine’s role in drug abuse and addiction. Neuropharmacology. 2009;56:3–8.
Ritchie T, Noble EP. Association of seven polymorphisms of the D2 dopamine receptor gene with brain receptor-binding characteristics. Neurochem Res. 2003;28:73–82.
Epstein LH, Wright SM, Paluch RA, Leddy JJ, Hawk LW, Jaroni JL, Saad FG, Crystal-Mansour S, Shields PG, Lerman C Relation between food reinforcement and dopamine genotypes and its effect on food intake in smokers. Am J Clin Nutr. 2004;80:82–8.
Epstein LH, Temple JL, Neaderhiser BJ, Salis RJ, Erbe RW, Leddy JJ. Food reinforcement, the dopamine D2 receptor genotype, and energy intake in obese and nonobese humans. Behav Neurosci. 2007;121:877–86.
Barnard ND, Noble EP, Ritchie T, Cohen J, Jenkins DJ, Turner-McGrievy G, Gloede L, Green AA, Ferdowsian H. D2 dopamine receptor Taq1A polymorphism, body weight, and dietary intake in type 2 diabetes. Nutrition. 2009;25:58–65.
Luo S, Liang Y, Cincotta AH. Intracerebroventricular administration of bromocriptine ameliorates the insulin-resistant/glucose-intolerant state in hamsters. Neuroendocrinology. 1999;69:160–6.
Defronzo RA. Bromocriptine: a sympatholytic, d2-dopamine agonist for the treatment of type 2 diabetes. Diabetes Care. 2011;34:789–94.
Starrenburg FC, Bogers JP. How can antipsychotics cause diabetes mellitus? Insights based on receptor-binding profiles, humoral factors and transporter proteins. Eur Psychiatry. 2009;24:164–70.
Donovan MH, Tecott LH. Serotonin and the regulation of mammalian energy balance. Front Neurosci. 2013;7:1–15.
Fletcher PJ, Paterson IA. A comparison of the effects of tryptamine and 5-hydroxytryptamine on feeding following injection into the paraventricular nucleus of the hypothalamus. Pharmacol Biochem Behav. 1989;32:907–11.
Leibowitz SF, Weiss GF, Suh JS. Medial hypothalamic nuclei mediate serotonin’s inhibitory effect on feeding behavior. Pharmacol Biochem Behav. 1990;37:735–42.
Fetissov SO, Meguid MM. Serotonin delivery into the ventromedial nucleus of the hypothalamus affects differently feeding pattern and body weight in obese and lean zucker rats. Appetite. 2010;54:346–53.
Breisch ST, Zemlan FP, Hoebel BG. Hyperphagia and obesity following serotonin depletion by intraventricular pchlorophenylalanine. Science. 1976;192:382–5.
Saller CF, Stricker EM. Hyperphagia and increased growth in rats after intraventricular injection of 5,7-dihydroxytryptamine. Science. 1976;192:385–7.
Waldbillig RJ, Bartness TJ, Stanley BG. Increased food intake, body weight, and adiposity in rats after regional neurochemical depletion of serotonin. J Comp Physiol Psychol. 1981;95:391–405.
Colman E, Golden J, Roberts M, Egan A, Weaver J, Rosebraugh C. The FDA’s assessment of two drugs for chronic weight management. N Engl J Med. 2012;367:1577–9.
Berglund ED, Liu C, Sohn JW, Liu T, Kim MH, Lee CE, Vianna CR, Williams KW, Xu Y, Elmquist JK. Serotonin 2C receptors in pro-opiomelanocortin neurons regulate energy and glucose homeostasis. J Clin Invest. 2013;123:5061–70.
Chen X, Margolis KJ, Gershon MD, Schwartz GJ, Sze JY. Reduced serotonin reuptake transporter (SERT) function causes insulin resistance and hepatic steatosis independent of food intake. PLoS One. 2012;7:e3251.
Ghaeli P, Shahsavand E, Mesbahi M, Kamkar MZ, Sadeghi M, Dashti-Khavidaki S. Comparing the effects of 8-week treatment with fluoxetine and imipramine on fasting blood glucose of patients with major depressive disorder. J Clin Psychopharmacol. 2004;24:386–8.
Iordanidou M, Tavridou A, Petridis I, Arvanitidis KI, Christakidis D, Vargemezis V, Manolopoulos VG. The serotonin transporter promoter polymorphism (5-HTTLPR) is associated with type 2 diabetes. Clin Chim Acta. 2010;411:167–71.
Furman BL, Wilson GA. Further studies on the effects of 5-hydroxytryptophan on plasma glucose and insulin in the mouse. Diabetologia. 1980;19:386–90.
Yamada J, Sugimoto Y, Kimura I, Takeuchi N, Horisaka K. Serotonin-induced hypoglycemia and increased serum insulin levels in mice. Life Sci. 1989;45:1931–6.
Hajduch E, Rencurel F, Balendran A, Batty IH, Downes CP, Hundal HS. Serotonin (5-hydroxytryptamine), a novel regulator of glucose transport in rat skeletal muscle. J Biol Chem. 1999;274:13563–8.
Fumeron F, Betoulle D, Nicaud V, Evans A, Kee F, Ruidavets JB, Arveiler D, Luc G, Cambien F. Serotonin transporter gene polymorphism and myocardial infarction: etude caste’moins de’ l’infarctus du myocarde (ECTIM). Circulation. 2002;105:2943–5.
Coto E, Reguero JR, Alvarez V, Morales B, Batalla A, Gonzalez P, Martin M, Garcia-Castro M, Iglesias-Cubero G, Cortina A. 5-hydroxytryptamine 5-HT2A receptor and 5-hydroxytryptamine transporter polymorphisms in acute myocardial infarction. Clinical Science. 2003;104:241–5.
Yuan X, Yamada K, Ishiyama-Shigemoto S, Koyama W, Nonaka K. Identification of polymorphic loci in the promoter region of the serotonin 5-HT2C receptor gene and their association with obesity and type II diabetes. Diabetologia. 2000;43:373–6.
Rosmond R, Bouchard C, Bjorntorp P. Increased abdominal obesity in subjects with a mutation in the 5-HT2A receptor gene promoter. Ann N Y Acad Sci. 2002;967:571–5.
Wolff B, Grabe HJ, Volzke H, Ludemann J, Schwahn C, Freyberger H, John U, Langes M, Cascorbi I, Felix SB. A functional serotonin transporter (SLC6A4) polymorphism modifies the association of smoking and diabetes with asymptomatic carotid atherosclerosis. Thromb Haemost. 2005;93:180–2.
Arinami T, Ohtsuki T, Yamakawa-Kobayashi K, Amemiya H, Fujiwara H, Kawata K, Ishiguro H, Hamaguchi H. A synergistic effect of serotonin transporter gene polymorphism and smoking in association with CHD. Thromb Haemost. 1999;81:853–6.
Muldoon MF, Mackey RH, Korytkowski MT, Flory JD, Pollock BG, Manuck SB. The metabolic syndrome is associated with reduced central serotonergic responsivity in healthy community volunteers. J Clin Endocrinol Metab. 2006;91:718–21.
Muldoon MF, Mackey RH, Williams KV, Korytkowski MT, Flory JD, Manuck SB. Low central nervous system serotonergic responsivity is associated with the metabolic syndrome and physical inactivity. J Clin Endocrinol Metab. 2004;89:266–71.
Muldoon MF, Mackey RH, Sutton-Tyrrell K, Flory JD, Pollock BG, Manuck SB. Lower central serotonergic responsivity is associated with preclinical carotid artery atherosclerosis. Stroke. 2007;38:2228–33.
Corona G, Ricca V, Bandini E, Rastrelli G, Casale H, Jannini EA, Sforza A, Forti G, Mannucci E, Maggi M. SIEDY scale 3, a new instrument to detect psychological component in subjects with erectile dysfunction. J Sex Med. 2012;9:2017–26.
Torner L, Toschi N, Pohlinger A, Landgraf R, Neumann ID. Anxiolytic and anti-stress effects of brain prolactin: improved efficacy of antisense targeting of the prolactin receptor by molecular modeling. J Neurosci. 2001;21:3207–14.
Drago F, Continella G, Conforto G, Scapagnini U. Prolactin inhibits the development of stress-induced ulcers in the rat. Life Sci. 1985;36:191–7.
Drago F, Pulvirenti L, Spadaro F, Pennisi G. Effects of TRH and prolactin in the behavioral despair (swim) model of depression in rats. Psychoneuroendocrinology. 1990;15:349–56.
Fujikawa T, Soya H, Yoshizato H, Sakaguchi K, Doh-Ura K, Tanaka M, Nakasima K. Restraint stress enhances the gene expression of prolactin receptor long form at the choroid plexus. Endocrinology. 1995;136:5608–13.
Walsh RJ, Slaby FJ, Posner BI. A receptor-mediated mechanism for the transport of prolactin from blood to cerebrospinal fluid. Endocrinology. 1987;120:1846–50.
Theorell T Prolactin—a hormone that mirrors passiveness in crisis situations. Integr Physiol Behav Sci. 1992;27:32–8.
Dunlop BW, Nemeroff CB. The role of dopamine in the pathophysiology of depression. Arch Gen Psychiatry. 2007;64:327–37.
Salamone JD, Aberman JE, Sokolowski JD, Cousins MS. Nucleus accumbens dopamine and rate of responding: neurochemical and behavioral studies. Psychobiology. 1999;27:236–47.
Neill DB, Fenton H, Justice JB. Increase in accumbal dopaminergic transmission correlates with response cost not reward of hypothalamic stimulation. Behav Brain Res. 2002;137:129–38.
Papp M, Klimek V, Willner P. Parallel changes in dopamine D2 receptor binding in limbic forebrain associated with chronic mild stress-induced anhedonia and its reversal by imipramine. Psychopharmacology. 1994;115:441–6.
Ichikawa J, Meltzer HY. Effect of antidepressants on striatal and accumbens extracellular dopamine levels. Eur J Pharmacol. 1995;281:255–61.
Randrup A, Munkvad I, Fog R, Gerlach J, Molander L, Kielberg B, Scheel-Kruger J. Mania, depression and brain dopamine. In: Essman WB, Valzelli S, editors. Current developments in psychopharmacology. New York: NY Spectrum Publications; 1975. p. 206–48.
Banki CM. Correlation between cerebrospinal fluid amine metabolites and psychomotor activity in affective disorders. J Neurochem. 1977;28:255–7.
Bowers Jr MB, Heninger GR, Gerbode F. Cerebrospinal fluid 5-hydroxyindoleacetic acid and homovanillic acid in psychiatric patients. Int J Neuropharmacol. 1969;8:255–62.
Mendels J, Frazer A, Fitzgerald RG, Ramsey TA, Stokes JW. Biogenic amine metabolites in cerebrospinal fluid of depressed and manic patients. Science. 1972;175:1380–2.
Miller HL, Delgado PL, Salomon RM, Berman R, Krystal JH, Heninger GR, Charney DS. Clinical and biochemical effects of catecholamine depletion on antidepressant-induced remission of depression. Arch Gen Psychiatry. 1996;53:117–28.
Monreal J, Duval F, Mokrani MC, Pinault G, Macher JP. Exploration de la fonction dopaminergique dans les depressions bipolares et unipolares. Ann Med Psychol (Paris). 2005;163:399–404.
McPherson H, Walsh A, Silverstone T. Growth hormone and prolactin response to apomorphine in bipolar and unipolar depression. J Affect Disord. 2003;76:121–5.
Post RM, Gerner RH, Carman JS, Gillin JC, Jimerson DC, Goodwin FK, Bunney Jr WE. Effects of a dopamine agonist piribedil in depressed patients: relationship of pretreatment homovanillic acid to antidepressant response. Arch Gen Psychiatry. 1978;35:609–15.
Nordin C, Siwers B, Bertilsson L. Bromocriptine treatment of depressive disorders. Clinical and biochemical effects. Acta psychiatr. Scand. 1981;64:25–33.
Waehrens J, Gerlach J. Bromocriptine and imipramine in endogenous depression. A double-blind controlled trial in out-patients. J Affect Disord. 1981;3:193–202.
Bouras N, Bridges P. Bromocriptine in depression. Curr Med Res Opin. 1982;8:150–3.
Theohar C, Fischer-Cornelssen K, Brosch H, Fischer E, Petrovic D. A comparative, multicenter trial between bromocriptine and amitriptyline in the treatment of endogenous depression. Arzneimittelforschung. 1982;32:783–7.
Izumi T, Inoue T, Kitagawa N, Nishi N, Shimanaka S, Takahashi Y, Kusumi I, Odagaki Y, Denda K, Ohmori T, Koyama T. Open pergolide treatment of tricyclic and heterocyclic antidepressant-resistant depression. J Affect Disord. 2000;61:127–32.
Bouckoms A, Mangini L. Pergolide: an antidepressant adjuvant for mood Disorders? Psychopharmacol Bull. 1993;29:207–11.
Inoue T, Tsuchiya K, Miura J, Sakakibara S, Denda K, Kasahara T, Koyama T. Bromocriptine treatment of tricyclic and heterocyclic antidepressant-resistant depression. Biol Psychiatry. 1996;40:151–3.
Kalia M Neurobiological basis of depression: an update. Metabolism. 2005;4:24–7.
Åsberg M, Nordström P, Träskman-Bendz L. Cerebrospinal fluid studies in suicide. An Overview Ann NY Acad Sci. 1986;487:243–55.
Asberg M, Traskman L, Thoren P. 5-HIAA in the cerebrospinal fluid. A biochemical suicide predictor? Arch Gen Psychiatry. 1976;33:1193–7.
Murphy DL, Campbell L, Costa JL. Current status of the indoleamine hypothesis of the affective disorders. In: Lipton MA, DiMascio A, Killam KF, editors. Psychopharmacology: a generation of progress. New York: Raven Press; 1978. p. 1235–47.
Meltzer HY, Lowy MT. The serotonin hypothesis of depression. In: Meltzer HY, editor. Psychopharmacology: the third generation of progress. New York: Raven Press; 1987. p. 513–26.
van Praag HM, de Hann S. Chemoprophylaxis of depression: an attempt to compare lithium with 5-hydroxytryptophan. Acta Psychiatr Scand. 1981;63:191–205.
Newman ME, Shapira B, Lerer B. Evaluation of central serotonergic function in affective and related disorders by the fenfluramine challenge test: a critical review. Int J Neuropsychopharmacol. 1998;1:49–69.
Kapitany T, Schindl M, Schindler SD, Hesselmann B, Füreder T, Barnas C, Sieghart W, Kasper S. The citalopram challenge test in patients with major depression and in healthy controls. Psychiatry Res. 1999;88:75–88.
Malone KM, Corbitt EM, Li S, Mann JJ. Prolactin response to fenfluramine and suicide attempt lethality in major depression. Br J Psychiatry. 1996;168:324–9.
Gordon JA, Hen R. The serotonergic system and anxiety. Neruomol Med. 2004;5:27–40.
Heisler LK, Chu HM, Brennan TJ, Danao JA, Bajwa P, Parsons LH, Tecott LH. Elevated anxiety and antidepressant-like responses in serotonin 5-HT1A receptor mutant mice. Proc Natl Acad Sci U S A. 1998;95:15049–54.
Parks CL, Robinson PS, Sibille E, Shenk T, Toth M. Increased anxiety of mice lacking the serotonin 1A receptor. Proc Natl Acad Sci U S A. 1998;95:10734–9.
Ramboz S, Oosting R, Amara DA, Kung HF, Blier P, Mendelsohn M, Mann JJ, Brunner D, Hen R. Serotonin receptor 1A knockout: an animal model of anxiety-related disorder. Proc Natl Acad Sci U S A. 1998;95:14476–81.
Griebel G 5-hydroxytryptamine-interacting drugs in animal models of anxiety disorders: more than 30 years of research. Pharmacol Therap. 1995;65:319–95.
Heils A, Teufel A, Petri S, Stober G, Riederer P, Bengel D. Lesch KP. Allelic Variation of Human Serotonin Transporter Gene Expression J Neurochem. 1996;66:2621–4.
Caspi A, Hariri AR, Holmes A, Uher R, Moffitt TE. Genetic sensitivity to the environment: the case of the serotonin transporter gene and its implications for studying complex diseases and traits. Am J Psychiatry. 2010;167:509–27.
Hariri AR, Drabant EM, Munoz KE, Kolachana BS, Mattay VS, Egan MF, Weinberger DR. A susceptibility gene for affective disorders and the response of the human amygdala. Arch Gen Psychiatry. 2005;62:146–52.
Pergamin-Hight L, Bakermans-Kranenburg MJ, van Ijzendoorn MH, Bar-Haim Y. Variations in the promoter region of the serotonin transporter gene and biased attention for emotional information: a meta-analysis. Biol. Psychiatr. 2012;71:373–9.
Reist C, Mazzanti C, Vu R, Tran D, Goldman D. Serotonin transporter promoter polymorphism is associated with attenuated prolactin response to fenfluramine. Am J Med Genet. 2001;105:363–8.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The Authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
Informed consent
Original data derive from a cohort of patients attending the Outpatient Clinic of Sexual Medicine and Andrology Unit of the University of Florence. All the data provided were collected as part of the routine clinical procedure. An informed consent was obtained from all patients.
Rights and permissions
About this article

Cite this article
Rastrelli, G., Corona, G. & Maggi, M. The role of prolactin in andrology: what is new?. Rev Endocr Metab Disord 16, 233–248 (2015). https://doi.org/10.1007/s11154-015-9322-3
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11154-015-9322-3