
Abstract
Purpose
Nitrogen (N) enrichment through either artificial N application or atmospheric N deposition often increases ecosystem aboveground net primary productivity (ANPP). Therefore, results from N addition experiments have been used to assess the effects of atmospheric N deposition on ecosystems. However, the frequency of atmospheric N deposition is higher than that of artificial N addition. Whether the frequency of N addition alters the long-term response of ecosystem ANPP remains unclear.
Methods
We conducted a N addition frequency experiment from 2010 in a temperate grassland, northern China. Plant community ANPP was collected in 2019 and 2020, and soil physicochemical properties were measured in 2020.
Results
Plant community ANPP was significantly enhanced by N addition, whereas these increments declined with the frequency of N addition. The responses of the grasses ANPP were similar to those of the plant community ANPP. Forbs ANPP was not significantly altered. Meanwhile, soil ammonium and nitrate (NO3−–N) concentrations decreased with increasing N addition frequency, while the soil water content (SWC) and pH were similar among the frequencies of N addition. Regardless of the frequency of N addition, SWC and soil NO3−–N jointly promoted grasses ANPP, ultimately increasing the plant community ANPP.
Conclusion
Our findings demonstrate that the frequency of N addition affects plant community biomass production through altering soil nitrate concentration in the semi-arid grassland. Therefore, this study illustrates that a higher frequency of N addition is more suitable for assessing the long-term impacts of atmospheric N deposition on ecosystems.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Plant community aboveground net primary productivity (ANPP) is a fundamental ecosystem functioning, providing food, fuel, and fiber (Haberl et al. 2007; Knapp et al. 2017; Migliavacca et al. 2021). The ecosystem biomass demand had been doubled in last century and was expected to increase in the coming decades (Krausmann et al. 2013). However, nitrogen (N) is limited in many terrestrial ecosystems, especially in temperate arid/semi-arid grasslands (Du et al. 2020; LeBauer and Treseder 2008; Vitousek and Howarth 1991), affecting plant community biomass production and ecosystem services.
It has been suggested that the rapidly increasing atmospheric N deposition caused by human activities (Galloway et al. 2021) could promote plant community ANPP in grasslands (Stevens et al. 2015). Similarly, N addition could increase plant community ANPP in various grasslands by partially or completely relieving N limitation through enrichment of soil N concentrations (Bai et al. 2010; LeBauer and Treseder 2008; Seabloom et al. 2021; Yue et al. 2020). Although both N fertilization and N deposition cause increments in plant community ANPP, empirical evidence regarding whether results from N addition experiments can accurately evaluate the influence of N deposition is still lacking, as there are apparent differences in N input frequency between artificial N application and atmospheric N deposition (Peng et al. 2020; Smith et al. 2009).
Clarifying whether the frequency of N input causes different effects on ecosystems is a prerequisite for accurately assessing the ecological effects of N deposition using results from N addition experiments (Smith et al. 2009). In most studies simulating N deposition (N addition) in grasslands, reactive N is input at a low frequency (once or several times) during the growing season (e.g., Bai et al. 2010; Clark and Tilman 2008; Eisenlord and Zak 2010; Peng et al. 2020). In contrast, for atmospheric N deposition, reactive N frequently enters ecosystems throughout the year (Smith et al. 2009). In the non-growing season, N input would not be immediately absorbed by plants because of the asynchrony between N availability and plant N demand in temperate herbs (Larsen et al. 2012; Ma et al. 2018, 2021). Moreover, these non-growing season N enrichments may cause N loss through leaching and/or emission. Therefore, atmospheric N deposition may result in reduced soil N availability during the growing season in comparison with that under N addition only during the growing season. This suggests that a higher plant community ANPP may be produced with a low frequency of N addition in the growing season compared to that under frequent N addition throughout the year. However, our previous studies found that higher soil ammonia emissions (Zhang et al. 2014a) and lower soil ammonium concentrations (Zhang et al. 2014b) are observed under the higher frequency of N addition, but soil nitrate and inorganic N concentrations and plant community ANPP are similar between low and high frequencies of N addition after consecutive 5-year treatments in a temperate grassland (Zhang et al. 2015).
There are three potential explanations for the above-mentioned responses of plant community ANPP between low and high frequencies of N addition. First, N applied during both the growing season and winter, even at a low frequency, cannot appropriately reflect soil N dynamics for the previous N addition during only the growing season. Second, plants in arid/semi-arid ecosystems may prefer NO3−–N (Wang and Macko 2011; Zhang et al. 2018a, 2016); however, soil NO3−–N concentrations do not show significant differences between low and high frequencies of N addition (Zhang et al. 2014b), resulting in similar responses in the plant community ANPP. Third, a significant difference in species diversity has been observed in a 5-year treatment; however, the lost species had relatively small biomass proportions (Zhang et al. 2015). The previous 5-year field experiment might be a relatively short-term study, as a recent report suggested that populations may need longer time (e.g., 10 years) to respond consistently to environmental fluctuations (Cusser et al. 2021). Therefore, it is necessary to conduct a long-term experiment with distinct frequencies of N addition, especially considering addition of N only in the growing season, to determine whether and how the results from N addition experiments can accurately verify the effects of atmospheric N deposition on ecosystem ANPP.
Grasslands in Inner Mongolia, which have high biodiversity and ecosystem productivity (Bai et al. 2004), have experienced relatively low atmospheric N deposition (Yu et al. 2019). Thus, it is an ideal model ecosystem to evaluate the impacts of N deposition on ecosystem functioning by comparing the effects of different frequencies of N addition. To evaluate the above-mentioned three potential explanations, based on our previous results with low and high frequencies of N addition (Zhang et al. 2015, 2014b, 2018c), we conducted a new field experiment with a control and three experimental frequencies (once per year, twice per year, and monthly) of N addition at 10 g N m−2 year−1. We addressed two questions: (i) whether the N-induced increments in plant community ANPP are associated with a decrease in the frequency of N addition? and (ii) if so, whether soil physicochemical factors mediate the responses of plant community ANPP to the frequency of N addition?
Materials and methods
Study site
The experiment was conducted in the Leymus chinensis grassland near the Inner Mongolia Grassland Ecosystem Research Station, Chinese Academy of Sciences (116°42′E, 43°38′N, ~ 1100 m a.s.l.), in the Inner Mongolia Autonomous Region, China. The long-term (1982–2020) mean annual precipitation was 341.3 mm, with approximately 71.4% falling during the growing season (from May to August). Annual precipitation was similar between 2019 and 2020 (Fig. S1). The long-term mean annual temperature is 1.1 °C, with a mean monthly temperature ranging from − 21.1 °C (January) to 20.0 °C (July). According to the Food and Agriculture Organization of the United Nations soil classification system, the soil is classified as Haplic Calcisol. The plant community is dominated by perennial grasses, which together account for more than 90% of the total peak aboveground biomass (Zhang et al. 2015). The field has been fenced since 1999 to exclude feeding and trampling by large herbivores. No fertilization was applied prior to the N addition experiment. The total ambient atmospheric N deposition was less than 1.0 g N m−2 year−1 in the grassland over the past three decades (Yu et al. 2019).
Experiment design
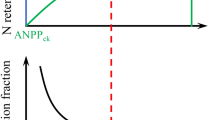
The frequency of N addition experiment (Fig. 1) was established in September 2010, with a randomized block design with six blocks (replicates). The field experiment contained a control (ambient condition; without N addition) and three frequencies of N addition (once per year [adding N in the growing season], twice per year [adding N in both the growing season and winter], and monthly) at 10 g N m−2 year−1 (the most commonly used load for assessing atmospheric N deposition in grasslands globally [Borer et al. 2014]) as analytic solid NH4NO3. Thus, there were 24 experimental plots in total; each plot was 4 × 4 m, with 1 m intervals between plots. Specifically, to match our previous N addition experiment with a low (twice per year) and high (monthly) frequency of N addition (Zhang et al. 2014b), monthly N addition (12 N additions year−1) was started on September 1, 2010, and reapplied on the first day of each following month; the twice per year N addition (2 N additions year−1) was started on November 1, 2010, and reapplied on the first day of each following June and November. In addition, for the once per year addition, N addition (1 N addition year−1) was started on June 1, 2010, and replied on the first day of each following June. Therefore, after monthly N addition in August (the peak plant community biomass period), the N loads were equal among the three frequencies of N addition (Fig. 1).

Nitrogen addition frequency experiment design and location. Randomized complete block design was employed, with six blocks. Control treatment (ambient condition without N addition) and three frequencies of N addition (once per year [1 N addition year−1], twice per year [2 N additions year−1], and monthly [12 N additions year−1]) were set in a temperate grassland in Inner Mongolia, China. Nitrogen (as NH4NO3) addition started on September 1, 2010, and reapplied on the first day of each month thereafter for the 12 N additions year−1 treatment, started on November 1, 2010, and reapplied on the first day of each June and November thereafter for the 2 N additions year−1, and started on June 1, 2011, and reapplied on the first day of each June thereafter for the 1 N addition year−1. That is, in August, the total annual N loadings equal 10 g N m−2 year−1 in each N-addition experimental plot
Field sampling and laboratory measurements
Plant community ANPP was estimated by investigating the peak aboveground biomass of the community, because plant aboveground tissues are dead during winter in temperate grasslands (Bai et al. 2004; Zhang et al. 2018b). Plant aboveground tissues, separated into grasses and forbs, were clipped using a 0.5 m × 1 m sampling quadrat in each experimental plot on August 16 in both 2019 and 2020 (the 10th year of N addition). The quadrats, without spatial overlapping, were randomly placed at least 0.5 m inside the border to avoid edge effects in each plot. Plant tissues were oven-dried at 65 °C for 48 h to a constant weight, and then weighed.
The top 10 cm soils, five cores with 3 cm diameter, were collected at the same location where plants were harvested in each plot on August 16, 2020. Each soil sample was thoroughly mixed, sieved through a 2 mm mesh, and divided into two subsamples. One was for soil water content (SWC, %) as well as soil ammonium (NH4+–N; mg kg−1 dry soil) and nitrate (NO3−–N; mg kg−1 dry soil) concentrations. For SWC, approximately 30 g of fresh soil was oven-dried at 105 °C for 48 h to a constant weight. For soil NH4+–N and NO3−–N concentrations, 10 g of fresh soil was extracted with 50 mL KCl (2.0 M). The extracts were measured using a flow injection auto analyzer (FLAstar 5000 Analyzer; Foss Tecator, Hillerød, Denmark). The other subsample was air-dried for soil pH (soil:water = 1:2.5) using a PHSJ-4A pH meter (Leici, Shanghai, China).
Statistical analyses
The relative ANPP of grasses/forbs was calculated as:
where ANPPi is the ANPP of grasses/forbs and ANPPcom is the plant community ANPP.
Repeated-measures analysis of variances (ANOVA) were used to test the effects of the frequency of N addition, year, and their interaction on the ANPP of the plant community, grasses, and forbs, and the relative ANPP of grasses/forbs, using block as a random factor. In addition, two-way ANOVAs were employed to detect the frequency of N addition, year, and their interaction on the ANPP of the plant community, grasses, and forbs, and the relative ANPP of grasses/forbs. The results were similar between repeated-measures (Table S1) and two-way (Table S2) ANOVA. One-way ANOVAs were used to test the effects of the frequency of N addition on the ANPP of the plant community, grasses, and forbs, and the relative ANPP of grasses/forbs each year.
The soil inorganic N concentration was the sum of the soil NH4+–N and NO3−–N. To satisfy the normality assumption, soil NH4+–N, NO3−–N, and inorganic N concentrations were natural logarithm-transformed. One-way ANOVAs were also used to detect the effects of the frequency of N addition on soil NH4+–N, NO3−–N, and inorganic N concentrations, soil NH4+–N/NO3−–N ratio, SWC, and soil pH, using a block as a random factor. Duncan's new multiple range test in R package ‘agricolae’ (de Mendiburu 2021) was employed at α = 0.05.
Moreover, the relationships between soil properties and the ANPP of plant community, grasses and forbs were explored through using correlation, partial correlation, stepwise regression, and path analysis. First, correlation analysis (function corr.test in package “psych”) was used. And the strong multicollinearity between variables was observed (Fig. S2). Second, partial correlation analysis was employed with controlling for the effects of the other soil variables, using the function pcor in package ‘ppcor’ (Kim, 2015). Third, stepwise regression through bidirectional elimination were employed using function step (package ‘stats’). The optimal models were selected based on the lowest Akaike's information criterion (AIC). Last, based on theory and previous empirical evidence (Zhang et al. 2015), we constructed an initial path analysis model (Fig. S3). By gradually eliminating insignificant paths using package ‘lavaan’ (Rosseel 2012), the final model fitted well with the lowest AIC. All statistical analyses and plots were performed on R 4.0.3 (R Core Team 2020). The code for all analyses can be found in the supplementary information.
Results
Effects of the frequency of N addition on plant community ANPP
Plant community ANPP was significantly increased by N addition (Fig. 2a), but the increments decreased with an increase in the frequency of N addition (Fig. 2a; Table S1; F3,15 = 6.1, P = 0.0066). In comparison with that of the control (ambient N), the mean plant community ANPP was enhanced by 57.4%, 31.9%, and 19.2% under 1 N (once per year), 2 N (twice per year), and 12 N (monthly) additions year−1, respectively (Fig. 2a). There was no significant difference between 2019 and 2020 (F1,20 = 2.5, P = 0.1282). No significant interaction effect was observed (F3,20 = 1.9, P = 0.1635).

Effects of the frequency of N addition on aboveground net primary productivity (ANPP). a Plant community, b grasses, and c forbs. Grey bars represent control treatment and blue bars present three N addition frequencies (dark blue = 1 N addition yr−1 [once per year], medium blue = 2 N additions yr−1 [twice per year], light blue = 12 N additions yr−1 [monthly]). Different letters indicate significant differences among the frequency of N addition (α = 0.05). Error bars indicate 1 SE
Effects of the frequency of N addition on the ANPP of grasses and forbs
Similar to the responses of the plant community ANPP, the ANPP of grasses was increased by N addition, and the increments significantly declined with an increase in the frequency of N addition in 2019 (Fig. 2b; F3,15 = 4.57, P = 0.0183) and 2020 (Fig. 2b; F3,15 = 3.10, P = 0.0584). The increments were 80.4%, 54.8%, and 27.0% under 1 N, 2 N, and 12 N additions year−1, respectively (Fig. 2b). The ANPP of forbs was not altered by N addition or frequency (Fig. 2c; Table S1; F3,15 = 0.2, P = 0.9041). There were no significant differences among years and the interaction of the frequency of N addition and year on the ANPP of grasses and forbs (Table S1; Ps > 0.2364). Moreover, no any significant effects were observed on the relative ANPP of grasses/forbs (Table S1; F3,15 = 0.4, P = 0.7475). On average, the relative ANPP of the grasses was 82.4%.
Effects of the frequency of N addition on soil properties
Soil NH4+–N (Fig. 3a), NO3−–N (Fig. 3b), and inorganic N concentrations (Fig. 3c) increased with N addition, whereas the increments decreased with an increase in the frequency of N addition (Fig. 3a–c). Soil NH4+–N (r = 0.92, P < 0.0001) and NO3−–N (r = 0.78, P < 0.0001) were positively correlated with the soil inorganic N concentration (Fig. S2). The soil NH4+–N/NO3−–N ratio was the highest at 1 N addition year−1 (Fig. 3d). SWC was similar among all four experimental treatments (Table S3; F3,15 = 1.1, P = 0.3619; Fig. 3e). Soil pH significantly decreased with N addition compared to that in the control, without significant differences among the three frequencies of N addition (Fig. 3f).

Effects of the frequency of N addition on soil properties: a soil ammonium concentration (NH4+–N), b soil nitrate concentration (NO3−–N), c soil inorganic N (NH4+–N + NO3−–N) concentration, d soil NH4+–N/NO3−–N ratio, e soil water content (SWC), and (e), soil pH. The soil NH4+–N, NO3−–N, and inorganic N concentrations were natural logarithm-transformed. Different letters indicate significant differences among the frequencies of N addition (α = 0.05). Error bars indicate 1 SE (n = 6)
Relationships between soil properties and plant community ANPP
Plant community ANPP was positively correlated with the ANPP of grasses (Fig. 4a; all years: R2 = 0.61, P < 0.0001) and forbs (Fig. 4b; all years: R2 = 0.18, P = 0.0025); however, the correlation between the ANPP of the plant community and that of forbs was not significant in 2019 (Fig. 4b; R2 = 0.07, P = 0.2174).

Proposed mechanisms of the effect of the frequency of N addition on plant community ANPP. Relationships between ANPP of community and (a), grasses and (b), forbs in 2019 (open symbols and red correlation) and in 2020 (closed symbols and green correlation). (c) Path analysis depicting the influence of soil properties on the ANPP of grasses and forbs in 2020. Numbers denote standardized coefficients. The proportion of variance explained (R2) is given. The goodness-of-fit statistics: χ2 = 1.28, df = 4, P = 0.865, GFI = 0.98, RMSEA < 0.001. Solid and dashed lines indicate significant (P ≤ 0.05) and nonsignificant (P > 0.05) relationships
After controlling for the effects of the other soil variables, results of partial correlation analysis showed that SWC was significantly positively associated with the ANPP of plant community (Table S4; r = 0.49, P = 0.0293) and grasses (Table S4; r = 0.61, P = 0.0040), and soil NO3−–N concentration was positively correlated with the ANPP of grasses (Table S4; r = 0.41, P = 0.0760). None of the other soil variables showed significantly partial correlation with the ANPP of plant community, grasses and forbs (Table S4; all Ps > 0.1275).
Moreover, the results of the stepwise regression analysis showed that SWC and soil NO3−–N concentration together explained 29% of the variation in the plant community ANPP (Table 1; P = 0.0268) and 51% of the variation in the ANPP of grasses (P = 0.0005), while no soil variables were obtained for the variation in the ANPP of forbs. The results of the path analysis also showed that SWC (Fig. 4c; standardized coefficient = 0.62) and soil NO3−–N concentration (standardized coefficient = 0.54) jointly promoted the ANPP of grasses. No significant relationships between soil variables and the ANPP of forbs were detected (Fig. 4c).
Discussion
We conducted a in situ study to detect the long-term effects of the frequency of N addition on plant community biomass production for evaluating whether and how the results of N addition experiments can verify the impacts of atmospheric N deposition on biomass production. We found that plant community ANPP was enhanced by N addition, but the increments decreased with an increase in the frequency of N addition. This suggests that a low frequency of N addition may overestimate the positive effect of atmospheric N deposition on plant community ANPP. The increments in the plant community ANPP were attributable to those of grasses ANPP in relation to the frequency of N addition. Moreover, irrespective of the frequency of N addition, SWC and soil NO3−–N concentration jointly promoted grasses ANPP, and ultimately enhanced the plant community ANPP. We also found that soil nitrate, not SWC, was influenced by the frequency of N addition. Taken together, our study indicates that soil nitrate mediates the responses of plant community aboveground biomass production to the frequency of N addition.
Effects of N addition on plant community ANPP
Consistent with previous reports from both field N addition experiments (Bai et al. 2010; LeBauer and Treseder 2008; Seabloom et al. 2021; Yue et al. 2020) and investigations of atmospheric N deposition gradients (Stevens et al. 2015), we found that plant community ANPP was enhanced under N addition. By providing long-term empirical evidence (continuous addition of N for 10 years), our findings confirm that N is a limiting factor of ecosystem productivity in temperate grasslands (Bai et al. 2010; Lü et al. 2018; Zhang et al. 2015).
Moreover, we found that grasses, which contributed 82.4% to the community, mediated the response of the plant community ANPP to N addition, in line with previous studies (La Pierre et al. 2016; Lü et al. 2018; Tian et al. 2020; Van Sundert et al. 2021) and in support of the mass ratio hypothesis (Grime 1998). In particular, the ANPP of grasses, not forbs, was promoted under N addition. It has been shown that N enrichment always promotes the ANPP of grasses (Bai et al. 2015; Hao et al. 2018; La Pierre et al. 2016; Tang et al. 2017; Van Sundert et al. 2021). For the ANPP of forbs, positive (La Pierre et al. 2016), neutral (Ren et al. 2021; Tang et al. 2017; Van Sundert et al. 2021), and negative (Bai et al. 2015; Lu et al. 2021) effects under N enrichment have all been reported in previous studies. On a global scale, a meta-analysis has shown that N enrichment has few impacts on alterations in the ANPP of forbs (You et al. 2017). The distinct responses between grasses and forbs may be attributed to their root morphology and proliferation. Higher specific root length and specific root area (Ravenek et al. 2016; Zheng et al. 2019; Zhou et al. 2018) can help grasses occupy N-rich patches (Šmilauerová and Šmilauer 2010) to promote the growth of aboveground parts. Meanwhile, faster growth in the ANPP of grasses may restrict forbs through light competition (Hautier et al. 2009; Van Sundert et al. 2021), resulting in non-significant increases in the ANPP of forbs under N-enriched conditions.
We further found that regardless of the frequency of N addition, soil NO3−–N rather than soil NH4+–N was positively associated with the ANPP of grasses. In arid/semi-arid ecosystems, native herbaceous plants prefer NO3−–N, which has been reported in deserts (Zhuang et al. 2020), grasslands (Ashton et al. 2010; Kahmen et al. 2006; Wang et al. 2021; Xu et al. 2011), and forests (Ma et al. 2021). Our finding may support the theory that with long-term evolutionary adaptation, preferential N form absorption by local plant species matches the characteristics of N cycles (Wang and Macko 2011; Zhang et al. 2018a, 2016). We also found that both the plant community and grasses were driven by the soil NO3−–N concentration, even though the soil NH4+–N concentration was higher (Fig. 3d). Recent empirical evidence showed that grasses mainly absorb NO3−–N in adjacent grasslands where soil NO3−–N is the most abundant form after N addition (Cao et al. 2021; Xi et al. 2017). From a physiological perspective, NO3−–N is the main N source for photosynthesis in chloroplasts (Heldt and Piechulla 2021; Tischner 2000), resulting in plant-specific NO3−–N preference. Furthermore, annual and biennial forbs, which are opportunistic species, do not show an apparent preference for N forms (Ren et al. 2021; Zhang et al. 2015). Together, these findings indicate that coevolution (Zhang et al. 2018a), rather than the N-induced NH4+–N increment, determines the impacts of N form on of plant community biomass production.
Effects of the frequency of N addition on plant community ANPP
Interestingly, we found that the plant community ANPP increased with a decrease in the frequency of N addition. The alterations in the plant community ANPP based on the frequency of N addition were attributed to changes in the ANPP of grasses, which were associated with the N addition frequency-induced soil NO3−–N concentration (Fig. 4). The frequency of N addition with seasonal events affects the soil NH4+–N, NO3−–N, and inorganic N concentrations. First, little N is absorbed by herbaceous plants (Joseph and Henry 2009; Ma et al. 2021) and immobilized by microbes in the non-growing season, that is, winter (Joseph and Henry 2009; Ma et al. 2018). Second, N addition in the winter tends to be lost from the rhizosphere through denitrification and leaching when soil moisture becomes saturated during freeze–thaw dynamics (Joseph and Henry 2009; Li et al. 2021; Müller et al. 2002). Third, Zhang et al. (2014a) reported that increased soil ammonia volatilization under a high frequency of N addition results in lower soil NH4+–N concentrations in the surface soils during the growing season. These ecological processes may have caused the reduced soil N accumulation in the surface soils during the growing seasons under the higher frequencies of N addition in our study. Therefore, stronger eutrophication at a lower frequency of N addition promotes increased plant community biomass production. This implies that experiments with a lower frequency of N addition may overestimate the positive effect of atmospheric N deposition on ecosystem productivity in the long term. Future research should clarify how the frequency of N addition affects ecosystem functioning to provide reliable parameters to accurately evaluate the ecological impacts of N deposition based on N addition field experiments.
Conclusion and outlook
By employing a 10-year field experiment with three frequencies of N addition (once per year [growing season], twice per year [both in growing season and winter], and monthly) in a semi-arid grassland, we found that the plant community ANPP increased with a decrease in the frequency of N addition. This suggests that previous N addition experiments may have overestimated the increase in plant community ANPP from atmospheric N deposition. Moreover, we found that lower soil NO3−–N concentration was associated with less ANPP of grasses and plant community at the higher frequency of N addition. Therefore, our study provides a complete evidence chain for long-term responses in productivity to verify that “using a higher frequency of N addition simulates atmospheric N deposition” (Zhang et al. 2014b). More importantly, given that (i) the alteration in plant biomass production is attributed to changes in soil NO3−–N under the frequency of N addition and (ii) NH4+–N/NO3−–N ratios have been changed in atmospheric N deposition (Du 2016; Liu et al. 2013; Yu et al. 2019), a new set of N deposition simulation experiments that account for both NH4+–N/NO3−–N ratios and the frequency of N addition are urgently needed in temperate grasslands.
Data availability
The datasets are available from the corresponding author on reasonable request.
References
Ashton IW, Miller AE, Bowman WD, Suding KN (2010) Niche complementarity due to plasticity in resource use: plant partitioning of chemical N forms. Ecology 91:3252–3260. https://doi.org/10.1890/09-1849.1
Bai W, Guo D, Tian Q, Liu N, Cheng W, Li L, Zhang W-H (2015) Differential responses of grasses and forbs led to marked reduction in below-ground productivity in temperate steppe following chronic N deposition. J Ecol 103:1570–1579. https://doi.org/10.1111/1365-2745.12468
Bai Y, Han X, Wu J, Chen Z, Li L (2004) Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature 431:181–184. https://doi.org/10.1038/nature02850
Bai Y, Wu J, Clark CM, Naeem S, Pan Q, Huang J, Zhang L, Han X (2010) Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from inner Mongolia grasslands. Glob Change Biol 16:358–372. https://doi.org/10.1111/j.1365-2486.2009.01950.x
Borer ET, Harpole WS, Adler PB, Lind EM, Orrock JL, Seabloom EW, Smith MD (2014) Finding generality in ecology: a model for globally distributed experiments. Methods Ecol Evol 5:65–73. https://doi.org/10.1111/2041-210X.12125
Cao J, Yang L, Pang S, Yang J, Hu Y, Li Y, Li L, Wang Q (2021) Convergent nitrogen uptake patterns and divergent nitrogen acquisition strategies of coexisting plant species in response to long-term nitrogen enrichment in a temperate grassland. Environ Exp Bot 185:104412. https://doi.org/10.1016/j.envexpbot.2021.104412
Clark CM, Tilman D (2008) Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 451:712–715. https://doi.org/10.1038/nature06503
Cusser S, Helms J IV, Bahlai CA, Haddad NM (2021) How long do population level field experiments need to be? Utilising data from the 40-year-old LTER network. Ecol Lett 24:1103–1111. https://doi.org/10.1111/ele.13710
de Mendiburu F (2021) agricolae: Statistical procedures for agricultural research. R package version 1.3–5. https://CRAN.R-project.org/package=agricolae (accessed by Aug. 2021)
Du E (2016) Rise and fall of nitrogen deposition in the United States. Proc Natl Acad Sci USA 113:E3594–E3595. https://doi.org/10.1073/pnas.1607543113
Du E, Terrer C, Pellegrini AFA, Ahlström A, van Lissa CJ, Zhao X, Xia N, Wu X, Jackson RB (2020) Global patterns of terrestrial nitrogen and phosphorus limitation. Nat Geosci 13:221–226. https://doi.org/10.1038/s41561-019-0530-4
Eisenlord SD, Zak DR (2010) Simulated atmospheric nitrogen deposition alters actinobacterial community composition in forest soils. Soil Sci Soc Am J 74:1157–1166. https://doi.org/10.2136/sssaj2009.0240
Galloway JN, Bleeker A, Erisman JW (2021) The human creation and use of reactive nitrogen: a global and regional perspective. Annu Rev Environ Resour 46:255–288. https://doi.org/10.1146/annurev-environ-012420-045120
Grime JP (1998) Benefits of plant diversity to ecosystems: immediate, filter and founder effects. J Ecol 86:902–910. https://doi.org/10.1046/j.1365-2745.1998.00306.x
Haberl H, Erb KH, Krausmann F, Gaube V, Bondeau A, Plutzar C, Gingrich S, Lucht W, Fischer-Kowalski M (2007) Quantifying and mapping the human appropriation of net primary production in earth’s terrestrial ecosystems. Proc Natl Acad Sci USA 104:12942–12947. https://doi.org/10.1073/pnas.0704243104
Hao T, Song L, Goulding K, Zhang F, Liu X (2018) Cumulative and partially recoverable impacts of nitrogen addition on a temperate steppe. Ecol Appl 28:237–248. https://doi.org/10.1002/eap.1647
Hautier Y, Niklaus PA, Hector A (2009) Competition for light causes plant biodiversity loss after eutrophication. Science 324:636–638. https://doi.org/10.1126/science.1169640
Heldt H-W, Piechulla B (2021) Nitrate assimilation is essential for the biosynthesis of organic matter. In: H-W Heldt, B Piechulla (eds) Plant Biochemistry (Fifth Edition). Academic Press.
Joseph G, Henry HAL (2009) Retention of surface nitrate additions in a temperate old field: implications for atmospheric nitrogen deposition over winter and plant nitrogen availability. Plant Soil 319:209–218. https://doi.org/10.1007/s11104-008-9862-3
Kahmen A, Renker C, Unsicker SB, Buchmann N (2006) Niche complementarity for nitrogen: An explanation for the biodiversity and ecosystem functioning relationship? Ecology 87:1244–1255. https://doi.org/10.1890/0012-9658(2006)87[1244:NCFNAE]2.0.CO;2
Kim S (2015) ppcor: An R Package for a Fast Calculation to Semi-partial Correlation Coefficients. CSAM 22:665–674. https://doi.org/10.5351/CSAM.2015.22.6.665
Knapp AK, Ciais P, Smith MD (2017) Reconciling inconsistencies in precipitation–productivity relationships: implications for climate change. New Phytol 214:41–47. https://doi.org/10.1111/nph.14381
Krausmann F, Erb K-H, Gingrich S, Haberl H, Bondeau A, Gaube V, Lauk C, Plutzar C, Searchinger TD (2013) Global human appropriation of net primary production doubled in the 20th century. Proc Natl Acad Sci USA 110:10324–10329. https://doi.org/10.1073/pnas.1211349110
La Pierre KJ, Blumenthal DM, Brown CS, Klein JA, Smith MD (2016) Drivers of variation in aboveground net primary productivity and plant community composition differ across a broad precipitation gradient. Ecosystems 19:521–533. https://doi.org/10.1007/s10021-015-9949-7
Larsen KS, Michelsen A, Jonasson S, Beier C, Grogan P (2012) Nitrogen uptake during fall, winter and spring differs among plant functional groups in a subarctic heath ecosystem. Ecosystems 15:927–939. https://doi.org/10.1007/s10021-012-9555-x
LeBauer DS, Treseder KK (2008) Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 89:371–379. https://doi.org/10.1890/06-2057.1
Li J, Zhao Y, Zhang A, Song B, Hill RL (2021) Effect of grazing exclusion on nitrous oxide emissions during freeze-thaw cycles in a typical steppe of Inner Mongolia. Agr Ecosyst Environ 307:107217. https://doi.org/10.1016/j.agee.2020.107217
Liu X, Zhang Y, Han W, Tang A, Shen J, Cui Z, Vitousek P, Erisman JW, Goulding K, Christie P, Fangmeier A, Zhang F (2013) Enhanced nitrogen deposition over China. Nature 494:459–462. https://doi.org/10.1038/nature11917
Lu P, Hao T, Li X, Wang H, Zhai X, Tian Q, Bai W, Stevens C, Zhang W-H (2021) Ambient nitrogen deposition drives plant-diversity decline by nitrogen accumulation in a closed grassland ecosystem. J Appl Ecol 58:1888–1898. https://doi.org/10.1111/1365-2664.13858
Lü X-T, Liu Z-Y, Hu Y-Y, Zhang H-Y (2018) Testing nitrogen and water co-limitation of primary productivity in a temperate steppe. Plant Soil 432:119–127. https://doi.org/10.1007/s11104-018-3791-6
Ma L, Liu G, Xu X, Xin X, Bai W, Zhang L, Chen S, Wang R (2018) Nitrogen acquisition strategies during the winter-spring transitional period are divergent at the species level yet convergent at the ecosystem level in temperate grasslands. Soil Biol Biochem 122:150–159. https://doi.org/10.1016/j.soilbio.2018.04.020
Ma L, Xu X, Zhang C, Lv Y, Liu G, Zhang Q, Feng J, Wang R (2021) Strong non-growing season N uptake by deciduous trees in a temperate forest: a 15N isotopic experiment. J Ecol. https://doi.org/10.1111/1365-2745.13754
Migliavacca M, Musavi T, Mahecha MD, Nelson JA, Knauer J, Baldocchi DD, Perez-Priego O, Christiansen R, Peters J, Anderson K, Bahn M, Black TA, Blanken PD, Bonal D, Buchmann N, Caldararu S, Carrara A, Carvalhais N, Cescatti A, Chen J, Cleverly J, Cremonese E, Desai AR, El-Madany TS, Farella MM, Fernández-Martínez M, Filippa G, Forkel M, Galvagno M, Gomarasca U, Gough CM, Göckede M, Ibrom A, Ikawa H, Janssens IA, Jung M, Kattge J, Keenan TF, Knohl A, Kobayashi H, Kraemer G, Law BE, Liddell MJ, Ma X, Mammarella I, Martini D, Macfarlane C, Matteucci G, Montagnani L, Pabon-Moreno DE, Panigada C, Papale D, Pendall E, Penuelas J, Phillips RP, Reich PB, Rossini M, Rotenberg E, Scott RL, Stahl C, Weber U, Wohlfahrt G, Wolf S, Wright IJ, Yakir D, Zaehle S, Reichstein M (2021) The three major axes of terrestrial ecosystem function. Nature 598:468–472. https://doi.org/10.1038/s41586-021-03939-9
Müller C, Martin M, Stevens RJ, Laughlin RJ, Kammann C, Ottow JCG, Jäger HJ (2002) Processes leading to N2O emissions in grassland soil during freezing and thawing. Soil Biol Biochem 34:1325–1331. https://doi.org/10.1016/S0038-0717(02)00076-7
Peng Y, Chen HYH, Yang Y (2020) Global pattern and drivers of nitrogen saturation threshold of grassland productivity. Funct Ecol 34:1979–1990. https://doi.org/10.1111/1365-2435.13622
R Core Team (2020). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/. (accessed by Nov. 2020)
Ravenek JM, Mommer L, Visser EJW, van Ruijven J, van der Paauw JW, Smit-Tiekstra A, de Caluwe H, de Kroon H (2016) Linking root traits and competitive success in grassland species. Plant Soil 407:39–53. https://doi.org/10.1007/s11104-016-2843-z
Ren Z, Zhang Y, Zhang Y (2021) Nitrogen deposition magnifies the positive response of plant community production to precipitation: ammonium to nitrate ratio matters. Environ Pollut 276:116659. https://doi.org/10.1016/j.envpol.2021.116659
Rosseel Y (2012) lavaan: an R package for structural equation modeling. J Stat Softw 48:1–36. https://doi.org/10.18637/jss.v048.i02
Seabloom EW, Adler PB, Alberti J, Biederman L, Buckley YM, Cadotte MW, Collins SL, Dee L, Fay PA, Firn J, Hagenah N, Harpole WS, Hautier Y, Hector A, Hobbie SE, Isbell F, Knops JMH, Komatsu KJ, Laungani R, MacDougall A, McCulley RL, Moore JL, Morgan JW, Ohlert T, Prober SM, Risch AC, Schuetz M, Stevens CJ, Borer ET (2021) Increasing effects of chronic nutrient enrichment on plant diversity loss and ecosystem productivity over time. Ecology 102:e03218. https://doi.org/10.1002/ecy.3218
Šmilauerová M, Šmilauer P (2010) First come, first served: grasses have a head start on forbs with prompt nutrient patch occupation. Plant Soil 328:327–336. https://doi.org/10.1007/s11104-009-0112-0
Smith MD, Knapp AK, Collins SL (2009) A framework for assessing ecosystem dynamics in response to chronic resource alterations induced by global change. Ecology 90:3279–3289. https://doi.org/10.1890/08-1815.1
Stevens CJ, Lind EM, Hautier Y, Harpole WS, Borer ET, Hobbie S, Seabloom EW, Ladwig L, Bakker JD, Chu C, Collins S, Davies KF, Firn J, Hillebrand H, Pierre KJL, MacDougall A, Melbourne B, McCulley RL, Morgan J, Orrock JL, Prober SM, Risch AC, Schuetz M, Wragg PD (2015) Anthropogenic nitrogen deposition predicts local grassland primary production worldwide. Ecology 96:1459–1465. https://doi.org/10.1890/14-1902.1
Tang Z, Deng L, An H, Yan W, Shangguan Z (2017) The effect of nitrogen addition on community structure and productivity in grasslands: a meta-analysis. Ecol Eng 99:31–38. https://doi.org/10.1016/j.ecoleng.2016.11.039
Tian Q, Yang L, Ma P, Zhou H, Liu N, Bai W, Wang H, Ren L, Lu P, Han W, Schultz PA, Bever JD, Zhang F-S, Lambers H, Zhang W-H (2020) Belowground-mediated and phase-dependent processes drive nitrogen-evoked community changes in grasslands. J Ecol 108:1874–1887. https://doi.org/10.1111/1365-2745.13415
Tischner R (2000) Nitrate uptake and reduction in higher and lower plants. Plant, Cell Environ 23:1005–1024. https://doi.org/10.1046/j.1365-3040.2000.00595.x
Van Sundert K, Arfin Khan MAS, Bharath S, Buckley YM, Caldeira MC, Donohue I, Dubbert M, Ebeling A, Eisenhauer N, Eskelinen A, Finn A, Gebauer T, Haider S, Hansart A, Jentsch A, Kübert A, Nijs I, Nock CA, Nogueira C, Porath-Krause AJ, Radujković D, Raynaud X, Risch AC, Roscher C, Scherer-Lorenzen M, Schuchardt MA, Schütz M, Siebert J, Sitters J, Spohn M, Virtanen R, Werner C, Wilfahrt P, Vicca S (2021) Fertilized graminoids intensify negative drought effects on grassland productivity. Glob Change Biol 27:2441–2457. https://doi.org/10.1111/gcb.15583
Vitousek PM, Howarth RW (1991) Nitrogen limitation on land and in the sea: how can it occur? Biogeochemistry 13:87–115. https://doi.org/10.1007/BF00002772
Wang L, Macko SA (2011) Constrained preferences in nitrogen uptake across plant species and environments. Plant Cell Environ 34:525–534. https://doi.org/10.1111/j.1365-3040.2010.02260.x
Wang R, Peñuelas J, Li T, Liu H, Wu H, Zhang Y, Sardans J, Jiang Y (2021) Natural abundance of 13C and 15N provides evidence for plant–soil carbon and nitrogen dynamics in a N-fertilized meadow. Ecology 102:e03348. https://doi.org/10.1002/ecy.3348
Xi N, Zhu B-R, Zhang D-Y (2017) Contrasting grass nitrogen strategies reflect interspecific trade-offs between nitrogen acquisition and use in a semi-arid temperate grassland. Plant Soil 418:267–276. https://doi.org/10.1007/s11104-017-3296-8
Xu X, Ouyang H, Richter A, Wanek W, Cao G, Kuzyakov Y (2011) Spatio-temporal variations determine plant–microbe competition for inorganic nitrogen in an alpine meadow. J Ecol 99:563–571. https://doi.org/10.1111/j.1365-2745.2010.01789.x
You C, Wu F, Gan Y, Yang W, Hu Z, Xu Z, Tan B, Liu L, Ni X (2017) Grass and forbs respond differently to nitrogen addition: a meta-analysis of global grassland ecosystems. Sci Rep 7:1563. https://doi.org/10.1038/s41598-017-01728-x
Yu G, Jia Y, He N, Zhu J, Chen Z, Wang Q, Piao S, Liu X, He H, Guo X, Wen Z, Li P, Ding G, Goulding K (2019) Stabilization of atmospheric nitrogen deposition in China over the past decade. Nat Geosci 12:424–429. https://doi.org/10.1038/s41561-019-0352-4
Yue K, Fornara DA, Li W, Ni X, Peng Y, Liao S, Tan S, Wang D, Wu F, Yang Y (2020) Nitrogen addition affects plant biomass allocation but not allometric relationships among different organs across the globe. J Plant Ecol 14:361–371. https://doi.org/10.1093/jpe/rtaa100
Zhang J, Cai Z, Müller C (2018a) Terrestrial N cycling associated with climate and plant-specific N preferences: a review. Eur J Soil Sci 69:488–501. https://doi.org/10.1111/ejss.12533
Zhang J, Wang J, Müller C, Cai Z (2016) Ecological and practical significances of crop species preferential N uptake matching with soil N dynamics. Soil Biol Biochem 103:63–70. https://doi.org/10.1016/j.soilbio.2016.08.009
Zhang Y, Feng J, Isbell F, Lü X, Han X (2015) Productivity depends more on the rate than the frequency of N addition in a temperate grassland. Sci Rep 5:12558. https://doi.org/10.1038/srep12558
Zhang Y, Han X, He N, Long M, Huang J, Zhang G, Wang Q, Han X (2014a) Increase in ammonia volatilization from soil in response to N deposition in Inner Mongolia grasslands. Atmos Environ 84:156–162. https://doi.org/10.1016/j.atmosenv.2013.11.052
Zhang Y, Loreau M, He N, Wang J, Pan Q, Bai Y, Han X (2018b) Climate variability decreases species richness and community stability in a temperate grassland. Oecologia 188:183–192. https://doi.org/10.1007/s00442-018-4208-1
Zhang Y, Lü X, Isbell F, Stevens C, Han X, He N, Zhang G, Yu Q, Huang J, Han X (2014b) Rapid plant species loss at high rates and at low frequency of N addition in temperate steppe. Glob Change Biol 20:3520–3529. https://doi.org/10.1111/gcb.12611
Zhang Y, Wang J, Stevens CJ, Lü X, He N, Wang C, Han X (2018c) Effects of the frequency and the rate of N enrichment on community structure in a temperate grassland. Journal of Plant Ecology 11:685–695. https://doi.org/10.1093/jpe/rtx041
Zheng Z, Bai W, Zhang W-H (2019) Root trait-mediated belowground competition and community composition of a temperate steppe under nitrogen enrichment. Plant Soil 437:341–354. https://doi.org/10.1007/s11104-019-03989-z
Zhou M, Bai W, Zhang Y, Zhang W-H (2018) Multi-dimensional patterns of variation in root traits among coexisting herbaceous species in temperate steppes. J Ecol 106:2320–2331. https://doi.org/10.1111/1365-2745.12977
Zhuang W, Li J, Yu F, Dong Z, Guo H (2020) Seasonal nitrogen uptake strategies in a temperate desert ecosystem depends on N form and plant species. Plant Biol 22:386–393. https://doi.org/10.1111/plb.13083
Acknowledgements
We thank Xiaoliang Wang, Yanhui Zhang, Hongying, Li Yuqin Zong, and Huiling Tian for assistance and the anonymous reviewers for their insightful comments. We appreciate the Inner Mongolia Grassland Ecosystem Research Station for the experimental facility support. This work was funded by National Natural Science Foundation of China (32071603 and 32122055) and the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA26020101).
Funding
This work was funded by National Natural Science Foundation of China (Grant numbers [32071603] and [32122055]) and the Strategic Priority Research Program of the Chinese Academy of Sciences (Grant numbers [XDA26020101]).
Author information
Authors and Affiliations
Contributions
Yunhai Zhang contributed to the study conception and design. Data collection and analysis were performed by Changchun Song, Yuqiu Zhang, Zhengru Ren, Haining Lu, Xu Chen, Ruoxuan Liu, Jungang Chen, and Yunhai Zhang. The first draft of the manuscript was written by Changchun Song and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
We declare no conflict of interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Responsible Editor: Devrim Coskun.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Song, C., Zhang, Y., Ren, Z. et al. Soil nitrate mediates the responses of plant community production to the frequency of N addition in a temperate grassland: a decadal field experiment. Plant Soil 491, 9–20 (2023). https://doi.org/10.1007/s11104-022-05435-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-022-05435-z