
Abstract
Aims
Soil heterogeneity is a primary mechanism explaining plant species diversity. Yet, controlled experiments yield inconsistent soil heterogeneity-diversity (SHD) relationships, ranging from positive, neutral to negative.
Methods
Here we investigated the SHD relationship by experimentally alternating nutrient-rich and nutrient-poor substrate in three dimensions, creating four levels of soil configurational heterogeneity (cell sizes 0, 12, 24 and 48 cm). Across each mesocosm, a mixture of species with high and low nitrogen requirements was evenly sown.
Results
Contrary to earlier experimental findings, this approach yielded a unimodal SHD relationship, peaking at cell size 12 cm. This pattern originated mainly from increased plant diversity of species with high nitrogen requirement. Diversity increases with configurational heterogeneity were not due to greater variation in light niches, and diversity decreases were not due to success of fast growing species. Strikingly, plant density increased monotonically with increasing configurational heterogeneity, indicating that not only more species but also more individuals could coexist.
Conclusions
This study provides experimental evidence for unimodal SHD curves in plant communities, which has hitherto only been predicted by models. Our results carry a striking similarity with other unimodal response patterns of plant species diversity, notably in diversity–disturbance and diversity–productivity relationships.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Spatial heterogeneity is likely to be an ultimate driver of plant species diversity (Tilman 1982, 1988; Tilman and Pacala 1993). However, the nature of the soil heterogeneity-diversity (SHD) relationship is not consistent across studies, and several theories have been put forward to explain the underlying mechanisms. The classical one is niche theory, which assumes that heterogeneous environments offer more niches than homogeneous environments, thus allowing more species to coexist (Tilman and Pacala 1993; Rosenzweig 1995; Williams and Houseman 2014). Positive SHD’s in line with this theory were indeed found in several experimental studies (e.g. Richardson et al. 2012; Williams and Houseman 2014). In contrast, other experiments have found negative SHD relationships (e.g. Gazol et al. 2013). The decreasing pattern was attributed to rapid depletion of resource-rich patches in heterogeneous soils by species with good foraging abilities, thus suppressing other species through asymmetric root competition (Hutchings et al. 2003; Wijesinghe et al. 2005). The easy access to patchily distributed soil resources would in turn also enhance shoot biomass, further suppressing competitors through asymmetric competition for light (Hautier et al. 2009; Lamb et al. 2009; DeMalach et al. 2017). Sometimes, neutral SHD relationships are found. In the experiment of Reynolds et al. (2007), this was attributed to clonal species obscuring the effect of soil heterogeneity on plant diversity (De Kroon and Bobbink 1997; Eilts et al. 2011; Baer et al. 2015). However, to our knowledge, no experimental study simultaneously tested many possible mechanisms, which hampers assessing their relative importance.
Investigating the SHD relationship in nature is complex because soil heterogeneity has a qualitative component (texture, nutrients, moisture, pH, etc.) and a configurational component (the size and distribution of patches) (Kelly and Canham 1992; Maestre and Cortina 2002; Dufour et al. 2006), and both these components vary in space and time (Tilman and Pacala 1993; Maestre et al. 2006; Maestre and Reynolds 2006). Experimental manipulation of soil heterogeneity, on the other hand, may bring more control and repeatability, but suffers from the lack of a standard method to vary soil heterogeneity. Some experimental studies have injected nutrients or spread fertilizer in a clumped pattern (Richardson et al. 2012), but doing so may not lead to stable patch sizes. Others have spatially redistributed soil from different layers at the same location, or soil from different locations (García-Palacios et al. 2011; Wubs and Bezemer 2016, 2017). While this may bring more realism, legacies from previous plant-soil feedback can confound current plant responses to soil heterogeneity (Brandt et al. 2013). Moreover, the studies that experimentally explored effects on plant species diversity have varied soil heterogeneity in two dimensions, yet soils are heterogeneous in three dimensions (Stewart et al. 2000). Finally, differences in species composition may explain some of the contrasting SHD findings, as in the aforementioned case of clonal species, or when N-fixing species change the original soil heterogeneity through local N-fixation.
Here we explore the SHD relationship with a mesocosm experiment where soil configurational heterogeneity is systematically varied in three dimensions using a recently developed technique (Fig.1a, Liu et al. 2017a), whilst excluding species that may significantly alter soil heterogeneity or blur response patterns such as N-fixing and clonal species (García-Palacios et al. 2012; Tamme et al. 2016). To allow different species to thrive and coexist on different substrates, as would be the case in nature, we apply the same seed rain to all mesocosms (Gazol et al. 2013). Compared with the existing literature, novel potential mechanisms are put forward as well as mechanisms proposed earlier. Our hypotheses are: (1) At patch scale, high availability of soil resource promotes biomass production, which in turn reduces light availability (and vice versa). The fine-scale alternation of small resource-rich and resource-poor patches at high levels of soil heterogeneity therefore creates greater spatial variation in light intensity at mesocosm scale, and thus more light niches, than the course-scale alternation of large resource-rich and resource-poor patches at low levels of soil heterogeneity. As a result, soil heterogeneity would indirectly increase diversity by weakening competition for light (Fig. 1b). (2) Alternatively, at mesocosm scale, soil heterogeneity decreases diversity, because, when cell size is small, slow growing species are eliminated by species with better foraging abilities (often fast growers, Fransen et al. 1999; Kembel and Cahill 2005) that can better exploit the more dispersed soil resources (Fig. 1c). Soil heterogeneity would thus lower diversity by accelerating species exclusion. (3) High soil heterogeneity (small patches) facilitates root access to resources from adjacent patches because of the shorter distance. At patch scale, communities growing on nutrient-poor patches will thus more easily gain resources from neighbouring nutrient-rich patches when the patch size is small, and this will enhance their productivity and light competition and reduce their species diversity (Fig. 1d-f). In contrast, communities growing on nutrient-rich patches will more easily lose resources to ingrowing neighbours from adjacent nutrient-poor patches when the patch size is small, thus reducing their productivity and light competition, and allowing more species to coexist. The balance of these changes on the two substrates will determine the species diversity response to heterogeneity at mesocosm scale (note that this is the case for all hypotheses).

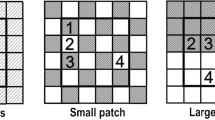
a Three-dimensional view of the mesocosms with the two substrates used in the experiment, where black and white colour indicates nutrient-rich and nutrient-poor substrate, respectively. Configurational heterogeneity decreases from left to right, from fine (small cells) to coarse (large cells) distribution of resources. The cell size of the mixture of the two substrates on the left can be considered as approximately zero. b At mesocosm scale, predicted pattern of the coefficient of variation of PAR transmission [CV(TPAR)] and plant species diversity along a gradient of increasing soil heterogeneity (decreasing cell size), under Hypothesis 1. c At mesocosms scale, predicted pattern of the species diversity of high N and low N species along a gradient of increasing soil heterogeneity (decreasing cell size), under Hypothesis 2. (d,e,f) At substrate scale, predicted pattern of biomass, PAR transmission (TPAR) and species diversity within nutrient-rich and nutrient-poor patches along a gradient of increasing soil heterogeneity (decreasing cell size), under Hypothesis 3
Materials and methods
Experimental design
As details of the experimental site and design are described in Liu et al. (2017b), who studied root responses of plant communities to soil heterogeneity in the same model ecosystems, we give a succinct description here. The experiment was conducted at University of Antwerp in Wilrijk, Belgium (51°09′41″N, 04°24′29″E), which is characterized by mild winters and cool summers, with average annual air temperature 10.6 °C and rainfall 832 mm, equally distributed throughout the year (Royal Meteorological Institute of Belgium). In spring 2015 we established four levels of three-dimensional soil heterogeneity in cubic mesocosms of the same size (48 cm × 48 cm × 48 cm), by varying the cell size within these mesocosms from 0 to 12, 24 and 48 cm (Fig. 1a). The cells were filled with nutrient-rich and nutrient-poor substrate, created by thoroughly mixing potting soil and sand in a 4:1 and a 1:4 ratio, respectively, in a cement mixer. Nutrients were the main difference between these two substrates, since soil water in the experiment was kept optimal. Each level of soil heterogeneity was constructed with the same amounts of the two substrates, so that only configurational heterogeneity was varied (via cell size) and qualitative heterogeneity was kept constant (see method in Liu et al. 2017a). Mesocosms with cell size 48 cm were filled with either nutrient-rich or nutrient-poor substrate; mesocosms with cell size 24 and 12 cm were filled with nutrient-rich and nutrient-poor substrate alternating in all directions; mesocosms with the smallest cell size were filled with a mixture of the two substrates, i.e. with both of them alternating at very short distance. The exact size could no longer be accurately measured as small aggregates of both substrates remained, but for convenience we named this cell size “0” cm.
We replicated the mesocosms with cell sizes 0, 12 and 24 cm five times, and the mesocosms with cell size 48 cm ten times, five with nutrient-rich and five with nutrient-poor substrate because they jointly constitute the mesocosm-scale response at 48 cm (they were lumped in mesocosm-scale analyses), but also to know the separate effects of both substrates. The mesocosms were contained in wooden boxes with drainage holes in the bottom. Liu et al. (2017a) provide further details on the technique to create soil heterogeneity in three dimensions.
The same seed rain was applied to all mesocosms, consisting of 24 species that naturally occur in grasslands in Belgium. This seed mixture covered a broad range of Ellenberg’s Indicator Nitrogen Values (Ellenberg et al. 1991), in order to allow potentially different communities to develop on nutrient-poor and nutrient-rich patches, as could be expected in nature. The species were classified in two groups, i.e. low N (Ellenberg 1–4) and high N (Ellenberg 6–8), with each group being represented by 12 species in order to avoid bias from uneven composition in the seed rain (Table 1). Low N and high N species tend to be slow growing and fast growing, respectively (Grime 1977; Chapin 1980; Franzaring et al. 2007). Seeds were obtained from commercial suppliers (Herbiseed, Reading, UK and Cruydt-Hoeck, Nijeberkoop, The Netherlands). We tested the germination rates and emergence times of these seeds 3 weeks before the start of experiment, and took them into account when composing the seed rain to have equal representation (aiming at six individuals per species) and germination timing (within a 2-week window) of all species. Only germination rate needed to be corrected. On 19 May 2015, each mesocosm received a uniform seed rain of 423 mixed seeds, sown randomly on the surface and covered with a few mm of the relevant substrate (i.e. nutrient-poor substrate on nutrient-poor cells and vice versa). This seed rain aimed for a distance of 4 cm between germinating individuals in each mesocosm. Mesocosms were kept moist to ensure optimal germination and establishment; later on, water was added at the prevailing frequency of rainfall events in the region (every 2 days) where natural rainfall fell short. Fungicide was added twice, one at the end of June and once 1 week later. Weeds were regularly removed.
Measurements and calculations
To assess the light environment of the plants, the horizontal distribution of photosynthetically active radiation (PAR) was measured with a custom-made miniature sensor in each mesocosm 5 cm above the soil surface, at every 2 cm along two parallel lines placed at respectively 18 and 30 cm from the edge of the wooden box (S1). These PARbelow canopy measurements were made on a cloudy day (1 September 2015) to avoid disturbance by sunflecks and to obtain an average across a range of solar angles. Incident PAR (PARabove canopy) was measured at the same time, yielding PAR transmission (TPAR):
The horizontal variation in PAR in each mesocosm, required to test Hypothesis 1, was assessed with the coefficient of variation of PAR transmission:
Abundance (density) was recorded by species, in four samples in mesocosms with cell size 0, and eight samples in mesocosms with cell sizes 12, 24 and 48 cm (four on nutrient-rich and four on nutrient-poor patches), during the last week of August 2015. Sample size was 12 cm × 12 cm and the squares were randomly placed within the substrate type. Values converted to m2 at mesocosm and substrate scale are shown in S2. At mesocosm scale, we also calculated whole-community abundance and abundance by group of species (high N or low N), likewise converted to m2.
The same data were used to assess species diversity, its components species richness and species evenness, and similarity in species composition between the two substrate types in a mesocosm (Table 2). Species richness at mesocosm scale refers to the total number of different species in the four 12 cm × 12 cm samples in a mesocosm, while species richness at substrate scale reflects the same for a given substrate in a mesocosm. Species richness at mesocosm scale was also separated into high N and low N species, required to test Hypothesis 2. Simpson’s diversity, Simpson’s evenness, Shannon-Wiener’s diversity and Shannon-Wiener’s evenness were calculated from the relative abundances of the species, likewise at mesocosm scale or by substrate type. Similarity indices (Sorensen and Bray-Curtis) assess the similarity of the species composition between the two substrate types in a mesocosm. Reflecting β-diversity, these indices connect α-diversity (substrate scale) with γ-diversity (mesocosm scale).
At the end of the experiment, on 2 September 2015, plant shoots in each mesocosm were cut 2–3 cm above the soil surface, separated by substrate type, oven dried at 70 °C for 4 days and weighed. Shoot biomass was calculated at mesocosm and at substrate scale by converting to m2. Average shoot biomass of individual plants in a mesocosm (not separated by species) was calculated as shoot biomass / plant density.
Statistical analysis
We first examined the nature of the SHD relationship. At mesocosm scale, one-way MANOVA was used to explore the effect of cell size (0, 12, 24 and 48 cm) on community shoot biomass, abundance, diversity indices and CV(TPAR). At substrate scale, we investigated the effects of cell size, substrate type and their interaction with two-way MANOVA on shoot biomass, abundance, diversity indices and TPAR of the local community on that substrate. Moreover, GLMM was conducted to test the performance of high N and low N species on different substrates. Cell size, species type, substrate type and their interactions were the fixed factors. Box identity was a random factor, and cell size 0 was excluded as substrates could not be distinguished in this treatment. In all these analyses, non-significant explanatory variables were excluded stepwise, and significant differences among treatments were explored further with post-hoc analysis (pairwise comparisons with Fisher’s LSD). All statistics were conducted with SPSS 23.0 (IBM Corp., 2015).
Next, to test the assumptions involved in hypothesis 1 and 3, structural equation modeling (SEM) was conducted (Gazol et al. 2013). Corresponding with hypothesis 1 and 3, we assumed that soil heterogeneity (cell size) influences diversity indirectly via changes in plant shoot biomass which themselves alter light availability (hypothesis 3), or its spatial variation (hypothesis 1). Yet, we also allowed for possible other relationships between SEM variables, for example a direct effect of soil heterogeneity on diversity, in order to test for possible alternative response pathways not included in the hypotheses. Because the relationship between cell size and the diversity indices was unimodal (see Results), with a positive response from cell size 0–12 cm and a negative response from cell size 12–48 cm, the underlying mechanisms were tested at mesocosm scale in separate SEMs for these ranges. However, SEMs at mesocosm scale only lead to an acceptable model when both the cell size 0 and the biomass variation between the two substrates within the mesocosm were removed. We therefore present results for SEMs on cell size 12–24-48 cm with effects of soil heterogeneity on PAR variation and subsequently on plant diversity, whilst keeping also the direct path from soil heterogeneity to plant diversity referred to above. Apart from these SEMs at mesocosm scale, we also conducted SEMs at substrate scale, but here only the response from 12 to 48 cm could be tested as responses to nutrient-rich and nutrient-poor substrate cannot be distinguished at cell size 0 cm. The overall fit of each SEM model was assessed by the χ2 statistic and the root square mean error of approximation (RMSEA), with non-significant χ2 and significant RMSEA indicating an acceptable fit of the model. In these final SEM diagrams, values along the path arrow refer to the standardized path coefficients and values above the variable refer to the proportion of variance that can be explained by relationships with other variables. SEM analyses were conducted with IBM SPSS Amos 23.0.
To test hypothesis 2, generalized linear mixed models (GLMMs) were applied to test effects of cell size, species type (high N or low N) and their interaction on community abundance and species richness, with box identity as a random factor. Finally, one-way ANOVA was performed to test the effect of cell size on community abundance, on the calculated average biomass of individual plants and on the similarity indices between the two substrate types in a mesocosm. In all these analyses, non-significant explanatory variables were excluded stepwise, and significant differences among treatments were explored further with post-hoc analysis (pairwise comparisons with Fisher’s LSD). Statistics in this section were conducted with SPSS 23.0 (IBM Corp., 2015).
Results
The MANOVA analyses revealed that cell size significantly affected community performance (F3,24 = 3.02, P < 0.005), whilst marginally significantly interacting with substrate type (F16,34 = 1.87, P = 0.062). The relationship between species richness and cell size at mesocosm scale was unimodal, with a peak at cell size 12 cm (Fig. 2b; Table 3). A similar pattern was observed for species diversity (Fig. 2c,d; Table 3), consistent with species evenness not being affected by cell size (Table 3; mean Simpson’s and Shannon-Wiener’s evenness were 0.79 ± SE 0.02 and 0.94 ± SE 0.01, respectively). The unimodal species richness response originated from nutrient-rich patches, as cell size did not affect richness on nutrient-poor patches (Fig. 3b; Table 4). Beta diversity between nutrient-rich and nutrient-poor patches did not contribute to the richness peak at 12 cm either, as Sorensen similarity was insensitive to cell size (Fig. 3g, Table 3; as mentioned above, community richness at mesocosm scale can be seen as gamma diversity, produced by the alpha diversities on both nutrient-rich and nutrient-poor patches, and the beta diversity between them). The unimodal response trend of species diversity to cell size likewise originated from the nutrient-rich patches (Fig. 3c,d, Table 4; again, cell size had no effect on nutrient-poor patches). However, in this case, beta diversity between nutrient-rich and nutrient-poor patches dampened the peak at mesocosm scale by reaching a minimum, i.e. Bray-Curtis similarity reaching a maximum (Fig. 3h), at 12 cm (we use Bray-Curtis similarity here instead of Sorensen because species diversity takes into account relative abundances). The GLMMs suggest that the higher richness and diversity going from cell size 48 cm to 12 cm mainly originated from the increase of high N species (Fig. 2g, Table 3), an increase that was observed on both nutrient-rich and nutrient-poor patches (not shown). These patterns being established, we can now move to the underlying hypotheses. In itself, a unimodal relationship excludes none of them, as it encompasses both an increasing and a decreasing response.

At mesocosm scale, mean ± SE of shoot biomass (a), species richness (b), Simpson’s diversity (c), Shannon-Wiener’s diversity (d), coefficient of variation of PAR transmission [CV(TPAR)] (e) and plant abundance (f) of the community, and mean ± SE of species richness (g) and plant abundance (h) of high N and low N species separately, all as a function of cell size. In (a-h), the grey symbol at 48 cm represents the average of the measurements on nutrient-rich (black symbol) and nutrient-poor (white symbol) mesocosms. Significant differences between treatments are indicated by different letters (post hoc analysis with Fisher’s LSD)

At substrate scale, mean ± SE of shoot biomass (a), species richness (b), Simpson’s diversity (c), Shannon-Wiener’s diversity (d), local community abundance (e), PAR transmission (TPAR) (f), Sorensen similarity (g) and Bray-Curtis similarity (h) as a function of cell size, on nutrient-rich (black) and nutrient-poor (white) patches. Response variables at cell size 0 are grey. Significant differences between treatments in (a-f) are indicated by different letters (post hoc analysis with Fisher’s LSD), with small and capital letters for nutrient-poor and nutrient-rich patches, respectively
Under Hypothesis 1 we expected greater diversity at higher soil heterogeneity (smaller cell size) because more edges between productive vegetation on nutrient-rich patches and unproductive vegetation on nutrient-poor patches would create more light niches. Cell size did not affect CV(TPAR) from 12 to 24-48 cm (Fig. 2e and SEM in Fig. 4). This result was similar when we excluded the edges of the mesocosms and only used the inner 24 × 24 cm area (not shown). Probably the shoot biomass on the two substrates was not different enough to generate much spatial variation of light, (Fig. 3a, see also corresponding effects on PAR transmission in Fig. 3b). Cell size 48 cm was the exception, with much less productive plants on nutrient-poor than on nutrient-rich patches, but these productivity differences cannot cause light variation within mesocosms either because there are no edges with adjacent patches (in fact 48 cm represents an ‘infinite’ cell size). Surprisingly, CV(TPAR) did not influence richness at cell size 12–24-48 cm (SEM in Fig. 4). Probably, CV(TPAR) – diversity relationships are hard to pinpoint across the very small ranges of CV(TPAR) observed in this experiment (cf. Fig. 2e). The SEMs also detected a direct negative effect of cell size on species richness from 12 to 24-48 cm (Fig. 4), corroborating the declining phase of the SHD relationship in Fig. 2b. Altogether, the support for Hypothesis 1 was thus limited. Paths observed in SEMs for species diversity (Simpson and Shannon-Wiener index, Fig. S3–4) were highly similar compared with those for species richness.

At mesocosm scale, structural equation model (SEM) relating cell size (12–24-48 cm) to coefficient of variation of PAR transmission [CV(TPAR)] and species richness. The statistics of SEM fitting are: χ2 = 0.355, P = 0.551, GFI = 0.988, RMSEA <0.001. Values above the variables refer to the proportion of variance that can be explained by relationships with other variables; values along the path arrows reflect the standardized path coefficients. Negative effects are indicated with ‘-’. Significant (P < 0.05) and marginally significant (P < 0.10) pathways are indicated with solid thick and dashed line, respectively
Under Hypothesis 2 we postulated lower diversity at greater soil heterogeneity, owing to fast growing species with good foraging abilities depleting the resource-rich cells more easily, at the expense of slower growers. Although species richness did decline from cell size 12 to zero (Fig. 2b), this hypothesis was not supported because the richness of high N species decreased at mesocosm scale over this range of cell sizes (Fig. 2g, Table 3), opposite to expectation.
Under Hypothesis 3 we assumed that, with increasing soil heterogeneity, a low-productive and thus species-richer community on nutrient-poor substrate will become more productive because root access to neighbouring nutrient-rich substrate is improved by the shorter distance. This would enhance light competition on the poor patches and diminish plant diversity there. This was not confirmed: although shoot biomass did increase (Fig. 3a and SEM in Fig. 5a) and PAR transmission did decrease (Fig. 3f and SEM in Fig. 5a) towards smaller cell size on nutrient-poor patches, as expected, a connection in the SEM between these two changes was not observed, nor did TPAR influence species richness. On nutrient-rich substrate, a high-productive and thus species-poorer community was expected to become less productive towards smaller cell size, because resources are then more easily lost to (more nearby) neighbouring species on nutrient-poor substrate. This would decrease light competition on the nutrient-rich patches and thus promote plant diversity there. This was not confirmed either: none of these paths were retained in the SEM for nutrient-rich substrate (Fig. 5b). Still, cell size negatively influenced species richness directly, corroborating the unimodal pattern in Fig. 3b. Combined for nutrient-rich and nutrient-poor patches, the trend in richness and diversity predicted by Hypothesis 3 is confirmed, at least from cell size 48 towards 12 cm (Fig. 2b), but not the underlying mechanism. Paths observed in SEMs for species diversity (Simpson and Shannon-Wiener index, Fig. S5–6) were highly similar compared with those for species richness.

At substrate scale, structural equation model (SEM) relating cell size (12, 24 and 48 cm) to root biomass, shoot biomass, PAR transmission and species richness, separately for nutrient-poor (a) and nutrient-rich substrate (b). The statistics of SEM fitting are: (a) χ2 = 1.020, P = 0.601, GFI = 0.964, RMSEA <0.001; (b) χ2 = 0.487, P = 0.784, GFI = 0.983, RMSEA <0.001. Values above the variables refer to the proportion of variance that can be explained by relationships with other variables; values along the path arrows reflect the standardized path coefficients. Negative effects are indicated with ‘-’. Significant (P < 0.05), marginally significant (P < 0.10), and nonsignificant pathways are indicated with solid thick, solid thin and dashed line, respectively
Interestingly, community abundance at mesocosm scale increased with smaller cell size (greater soil heterogeneity) (Fig. 2f, Table 3). This originated mainly from nutrient-rich patches (Fig. 3e, Table 4), and high N species also contributed more to this increase than low N species (Fig. 2h). At the same time, cell size did not affect the average biomass of individual plants in a mesocosm (P = 0.600). Likewisely, cell size did not affect shoot biomass per unit area at mesocosm scale (Fig. 2a), in agreement with the contrasting responses on the two substrates (Fig. 3a).
Discussion
The idea that soil heterogeneity drives plant species diversity has attracted much attention in recent decades (Tilman 1982, 1988; Hutchings et al. 2003; Williams and Houseman 2014). However, lack of a standard method to create soil heterogeneity experimentally and the inclusion of species that can blur fundamental trends in empirical tests (e.g. clonal or N-fixing species) may have prevented consistent SHD relationships from emerging. In the current experiment we systematically varied the patch size of each of the two used substrates (one nutrient rich, one nutrient poor), from small to large in three dimensions, whilst avoiding confounding by specific species. Contrary to previous findings in controlled experiments, we identified a novel pattern in the form of a unimodal SHD relationship, with diversity first increasing and then decreasing across the 3-D cell size range. Surprisingly, our hypothesized mechanisms to explain these responses, several of which were based on earlier assumptions in the heterogeneity-diversity literature, were not confirmed. However, our results do point to other potential mechanisms.
To test Hypothesis 1 that soil heterogeneity promotes species diversity by generating light niches, induced by the productivity differences between nutrient-rich and nutrient-poor patches, the spatial variation of PAR transmission in mesocosms was measured. This was not explicitly considered in previous studies on soil heterogeneity (Borer et al. 2014). Soil heterogeneity generating light niches is analogous to species diversity (i.e. heterogeneity in plant traits) generating light niches (Spehn et al. 2000), which in turn allows species to coexist. Yet, in our experiment, cell size did not affect the variation of light transmission [CV(TPAR)] through modifying biomass (including biomass variation did not lead to an acceptable SEM) at mesocosm scale: shoot biomass was too similar on nutrient-rich and nutrient-poor patches (Fig. 2a), as well as high enough to produce low TPAR values, across the 0–12-24 cm cell size range (Fig. 3f), thus offering little potential for light niche differentiation. Liu et al. (2017b) attributed this shoot biomass similarity to easier root access of plants growing on nutrient-poor patches to soil resources in neighbouring nutrient-rich patches when cell size is smaller. However, we cannot exclude that the observed positive SHD response across part of the cell size range was caused by the presence of more light niches before full light interception was reached (Sapijanskas et al. 2014; Vojtech et al. 2008), as biomass would be expected to increase faster on nutrient-rich than on nutrient-poor patches. Interestingly, in the SEMs, cell size directly reduced plant diversity from 12 to 48 cm. This points to other mechanisms than those hypothesized here, and thus requires further research.
Contrary to Hypothesis 1, Hypothesis 2 postulated impoverished communities at high soil heterogeneity, owing to fast growing species outcompeting slow growers through rapid depletion of resource-rich cells. While species richness did drop across one part of the SHD range (from 12 to 0 cm), the underlying cause was opposite: high N species were lost instead of low N species. Tamme et al. (2010) and Laanisto et al. (2013) proposed that negative SHD relationships might also ensue from increased isolation and lack of connectivity among patches at high levels of heterogeneity, but the question remains whether these principles from landscape fragmentation apply across the 12 to 0 cm cell size range. Possibly, very small pockets of nutrient-rich substrate (i.e., smaller than the plant size) offer insufficient resources to maintain a large diversity of fast growing species because they co-occur with nutrient-poor cells, thus locally reducing the mean resource availability relative to larger nutrient-rich cells where plant individuals only ‘sense’ the most favourable substrate. This would also explain why these fast growers could not outcompete the slow growing species from cell size 12 to 0 cm. The explanation of insufficient resources at very small cell size would not be incompatible with the observed increase of the diversity of the high N species on another part of the cell size range, i.e. from 48 to 12 cm. The latter could arise from relaxation of intense competition among these fast growing species, and thus low diversity, from cell size 48 towards 12 cm, especially in nutrient-rich patches. Note that this mechanism is opposite to Hypothesis 2. Moreover, the 3D structure of soil heterogeneity in this experiment may also explain this, as the roots in a nutrient-rich patch quickly encounter poor soil when they grow deeper, resulting in reduced nutrient availability which would likewise relax competition. Different underlying reasons for species impoverishment at both very small and large cell sizes may thus explain the unimodal SHD relationship, similar to other unimodal plant diversity patterns such as diversity-productivity (Fraser et al. 2015) and diversity-disturbance (Kondoh 2001).
Hypothesis 3 was based on the aforementioned greater resource loss from nutrient-rich patches through extraction by species on neighbouring nutrient-poor patches as cell size gets smaller (Liu et al. 2017b, Fig. 3g), thus reducing the productivity and light competition and increasing the species diversity on nutrient-rich patches. However, such easier root access should increase the productivity and light competition on nutrient-poor patches, reducing species diversity there. Depending on the balance of these processes, increasing as well as decreasing SHD relationships at mesocosm scale might thus arise, in principle also giving rise to unimodal curves. Yet, though we observed higher biomass and reduced light availability on nutrient-poor patches as cell size decreased, diversity on these patches was not reduced. On nutrient-rich patches, on the other hand, we found increases in diversity even though the expected lower biomass and decreased light availability was not observed. Nevertheless, we think that the mechanisms in Hypothesis 3, which basically consider only shading within a patch, might still hold, but could be blurred by the associated, simultaneous effects of shading by the neighbouring patches (Anten and Hirose 1999; DeMalach et al. 2016, 2017).
Previous studies on heterogeneity have to our knowledge not measured plant density along a range of controlled soil heterogeneity. In our mesocosms, plant density increased monotonically towards small cell size, so not only more species were able to coexist on the same area (up to cell size 12 cm), but also more individuals (up to cell size 0 cm). As cell size did not affect community shoot biomass, the more numerous plants growing on small cells would be expected to be less productive, which was not the case. We also considered whether small-cell mesocosms contained more species of small stature. This was not confirmed either: the average species height (derived from www.try-db.org on 19 March 2018), weighted by their relative abundance as observed in our mesocosms, was highly similar (49 and 51 cm at cell size 48 and 12 cm, respectively), indicating that the community composition probably did not shift to intrinsically smaller species and that the enhanced coexistence of more individuals may be caused by other factors. In any case, starting from the seed rain, less competitive exclusion occurred at cell size 12 than at 48 cm since species richness was higher there, pointing at the same conclusion of improved coexistence (assuming equal germination across cell sizes because mesocosms contained equal amounts of nutrient-rich and nutrient-poor surface soil). Note that our findings of greater density and similar community biomass in mesocosms with smaller cell size also point to the law of constant final yield (Kira et al. 1953; Weiner 2004).
We analysed effects of soil heterogeneity in line with the method in our earlier publications (Liu et al. 2017a, b), along a gradient from very large to very small substrate patches. This variation in cell size thus considers heterogeneity in the spatial, physical sense. It could be argued that the way these patches of varying size are perceived, depends very much on the organism. As such, heterogeneity would be different for trees, forbs, mosses, and soil bacteria on a soil with the same absolute, physical spatial heterogeneity (‘cell size’). What is very heterogeneous to one organism, could be sensed as homogeneous by another, for example a large plant on our 12 cm cells versus a bacteria in the middle of that cell. Moreover, this perception can also change during the organism’s life cycle, for instance, from seedling to large plant. This makes it very difficult to quantify ‘perceived’ heterogeneity in a multi-species community consisting of very differently-sized organisms, and makes the physical (cell size) approach more practical in analyses. Nevertheless, much like in studies on other environmental drivers, where for example the same air temperature may be perceived very differently by various coexisting plant species depending on their thermal traits (e.g. Michaletz et al. 2015), the interpretation of the analyses should take into account potential varying perception of the driver (here: spatial heterogeneity). It is unclear whether the perception of heterogeneity changes towards the smallest cell sizes in general, i.e. whether plants perceive their environment as increasingly homogeneous when substrate patches get very small, which would mean spatial (physical) and perceived heterogeneity start diverging at some point. To shed more light on this, new studies would need to include additional levels of spatial heterogeneity on side of the gradient (between cell sizes 0 and 12 cm).
We conclude that species diversity responses to small-scale spatial soil heterogeneity can be unimodal, which to our knowledge was not experimentally observed before for plant communities, although it has been studied in simulation modelling at large spatial scale where it was attributed to greater extinction risk of small plant populations from inbreeding depression and stochastic events (Kadmon and Allouche 2007; Allouche et al. 2012). These mechanisms clearly do not operate at the scale of our experiment. The location of the SHD peak at 12 cm suggests different underlying mechanisms, which seem to switch around this point, as proposed earlier (Fitter 1994). Studies at the very small scale may therefore hold the key to progress in this domain. However, future studies should also include longer-term community dynamics mediated by further competitive exclusion (beyond the level observed here) or by possible influences of soil heterogeneity on seed production and dispersal. For example, the probability of dispersal of species adapted to nutrient-rich patches into surrounding less suitable nutrient-poor habitats may increase with decreasing cell size, because these less suitable habitats are then more nearby. This might alter the competitive balance between species, and thus species diversity. Longer-term experiments would also create their own seed rain, possibly altering species recruitment.
References
Allouche O, Kalyuzhny M, Moreno-Rueda G, Pizarro M, Kadmon R (2012) Area-heterogeneity tradeoff and the diversity of ecological communities. Proc Natl Acad Sci 109:17495–17500
Anten NPR, Hirose T (1999) Interspecific differences in above-ground growth patterns result in spatial and temporal partitioning of light among species in a tall-grass meadow. J Ecol 87:583–597
Baer SG, Blair JM, Collins SL (2015) Environmental heterogeneity has a weak effect on diversity during community assembly in tallgrass prairie. Ecol Monogr 86:94–106
Borer ET, Seabloom EW, Gruner DS, Harpole WS, Hillebrand H, Lind EM, Adler PB, Alberti J, Anderson TM, Bakker JD, Biederman L, Blumenthal D, Brown CS, Brudvig LA, Buckley YM, Cadotte M, Chu C, Cleland EE, Crawley MJ, Daleo P, Damschen EI, Davies KF, DeCrappeo NM, du G, Firn J, Hautier Y, Heckman RW, Hector A, HilleRisLambers J, Iribarne O, Klein JA, Knops JMH, la Pierre KJ, Leakey ADB, Li W, MacDougall AS, McCulley RL, Melbourne BA, Mitchell CE, Moore JL, Mortensen B, O'Halloran LR, Orrock JL, Pascual J, Prober SM, Pyke DA, Risch AC, Schuetz M, Smith MD, Stevens CJ, Sullivan LL, Williams RJ, Wragg PD, Wright JP, Yang LH (2014) Herbivores and nutrients control grassland plant diversity via light limitation. Nature 508:517–520
Brandt AJ, De Kroon H, Reynolds HL, Burns JH (2013) Soil heterogeneity generated by plant–soil feedbacks has implications for species recruitment and coexistence. J Ecol 101:277–286
Chapin FS (1980) The mineral nutrition of wild plants. Annu Rev Ecol Syst 11:233–260
De Kroon H, Bobbink R (1997) Clonal plant dominance under elevated nitrogen deposition, with species reference to Brachypodium pinnatum in chalk grassland. In: De Kroon H, Van Groenendael J (eds) The ecology and evolution of clonal plants. Backhuys, Leiden, pp 359–379
DeMalach N, Zaady E, Weiner J, Kadmon R (2016) Size asymmetry of resource competition and the structure of plant communities. J Ecol 104:899–910
DeMalach N, Zaady E, Kadmon R (2017) Light asymmetry explains the effect of nutrient enrichment on grassland diversity. Ecol Lett 20:60–69
Dufour A, Gadallah F, Wagner HH, Guisan A, Buttler A (2006) Plant species richness and environmental heterogeneity in a mountain landscape: effects of variability and spatial configuration. Ecography 29:573–584
Eilts JA, Mittelbach GG, Reynolds HL, Gross KL (2011) Resource heterogeneity, soil fertility, and species diversity: effects of clonal species on plant communities. Am Nat 177:574–588
Ellenberg H, Webber HE, Düll R, Wirth V, Werner W, Paulissen D (1991) Zeigerwerte von Pflanzen in Mitteleuropa. Scripta Geobotanica 18:1–248
Fitter AH (1994) Architecture and biomass allocation as components of the plastic response of root systems to soil heterogeneity. In: Caldwell MM, Pearcy RM (eds) Exploitation of environmental heterogeneity by plants. Academic Press, San Diego, pp 305–323
Fransen BH, De Kroon H, De Kovel CGF, Van den Bosch F (1999) Disentangling the effects of root foraging and inherent growth rate on plant biomass accumulation in heterogeneous environments: a modelling study. Ann Bot 84:305–311
Franzaring J, Fangmeier A, Hunt R (2007) On the consistencies between CSR plant strategies and Ellenberg ecological indicator values. J Appl Bot Food Qual 81:86–94
Fraser LH, Pither J, Hentsch A, Sternberg M, Zobel M, Askarizadeh D et al (2015) Worldwide evidence of a unimodal relationship between productivity and plant species richness. Science 349:302–305
García-Palacios P, Maestre FT, Gallardo A (2011) Soil nutrient heterogeneity modulates ecosystem responses to changes in the identity and richness of plant functional groups. J Ecol 99:551–562
García-Palacios P, Maestre FT, Bardgett RD, De Kroon H (2012) Plant responses to soil heterogeneity and global environmental change. J Ecol 100:1303–1314
Gazol A, Tamme R, Price JN, Hiiesalu I, Laanisto L, Pärtel M (2013) A negative heterogeneity-diversity relationship found in experimental grassland communities. Oecologia 173:545–555
Grime JP (1977) Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am Nat 111:1169–1194
Hautier Y, Niklaus PA, Hector A (2009) Competition for light causes plant biodiversity loss after eutrophication. Science 324:636–638
Hutchings MJ, John EA, Wijesinghe DK (2003) Toward understanding the consequences of soil heterogeneity for plant populations and communities. Ecology 84:2322–2334
Kadmon R, Allouche O (2007) Interacting the effects of area, isolation, and habitat heterogeneity on species diversity: a unification of island biogeography and niche theory. Am Nat 170:443–454
Kelly VR, Canham CD (1992) Resource heterogeneity in oldfields. J Veg Sci 3:545–552
Kembel SW, Cahill JF (2005) Plant phenotypic plasticity belowground: a phylogenetic perspective on root foraging trade-offs. Am Nat 166:216–230
Kira T, Ogawa H, Shinozaki K (1953) Intraspecific competition among higher plants I. competition-yield-density interrelationships in regularly dispersed populations. Journal of the Institute of Polytechnics. Series D 4:1–16
Kondoh M (2001) Unifying the relationship of species richness to productivity and disturbance. P Roy Soc B-Bio Sci 268:269–271
Laanisto L, Tamme R, Hiiesalu I, Szava-Kovats R, Gazol A, Pärtel M (2013) Microfragmentation concept explains non-positive environmental heterogeneity-diversity relationships. Oecologia 171:217–226
Lamb EG, Kembel SW, Cahill JF (2009) Shoot, but not root, competition reduces community diversity in experimental mesocosms. J Ecol 97:155–163
Liu Y, De Boeck HJ, Wellens MJ, Nijs I (2017a) A simple method to vary soil heterogeneity in three dimensions in experimental mesocosms. Ecol Res 32:287–295
Liu Y, Bortier MF, De Boeck HJ, Nijs I (2017b) Root distribution responses to three-dimensional soil heterogeneity in experimental mesocosms. Plant Soil 421:353–366
Maestre FT, Cortina J (2002) Spatial patterns of surface soil properties and vegetation in a Mediterranean semi-arid steppe. Plant Soil 241:279–291
Maestre FT, Reynolds JF (2006) Small-scale spatial heterogeneity in the vertical distribution of soil nutrients has limited effects on the growth and development of Prosopis glandulosa seedlings. Plant Ecol 183:65–75
Maestre FT, Bradford MA, Reynolds JF (2006) Soil heterogeneity and community composition jointly influence grassland biomass. J Veg Sci 17:261–270
Michaletz ST, Weiser MD, Zhou J, Kaspari M, Helliker BR, Enquist BJ (2015) Plant thermoregulation: energetics, trait–environment interactions, and carbon economics. Trends Ecol Evol 30:714–724
Reynolds HL, Mittelbach GG, Darcy-Hall TL, Houseman GR, Gross AL (2007) No effect of varying soil resource heterogeneity on plant species richness in a low fertility grassland. J Ecol 95:723–733
Richardson PJ, MacDougall AS, Larson DW (2012) Fine-scale spatial heterogeneity and incoming seed diversity additively determine plant establishment. J Ecol 100:939–949
Rosenzweig ML (1995) Species diversity in space and time. Cambridge University Press, Cambridge
Sapijanskas J, Paquette A, Potvin C, Kunert N, Loreau M (2014) Tropical tree diversity enhances light capture through crown plasticity and spatial and temporal niche differences. Ecology 95:2479–2492
Spehn EM, Joshi J, Schmid B, Alphei J, Körner C (2000) Plant diversity effects on soil heterotrophic activity in experimental grassland systems. Plant Soil 224:217–230
Stewart AJA, John EA, Hutchings MJ (2000) The world is heterogeneous: ecological consequences of living in a patchy environment. In: Hutchings MJ, John EA, Stewart AJA (eds) The ecological consequences of environmental heterogeneity. Blackwell Science, Cambridge, pp 1–8
Tamme R, Hiiesalu I, Laanisto L, Szava-Kovats R, Pärtel M (2010) Environmental heterogeneity, species diversity and co-existence at different spatial scales. J Veg Sci 21:796–801
Tamme R, Gazol A, Price JN, Hiiesalu I, Pärtel M (2016) Co-occurring grassland species vary in their responses to fine-scale soil heterogeneity. J Veg Sci 27:1012–1022
Tilman D (1982) Resource competition and community structure. In: Princeton University press, Princeton
Tilman D (1988) Plant strategies and the dynamics and structure of plant communities. In: Princeton University press, Princeton
Tilman D, Pacala S (1993) The maintenance of species richness in plant communities. In: Ricklefs R, Schluter D (eds) Species diversity in ecological communities. University of Chicago Press, Chicago, pp 13–25
Vojtech E, Loreau M, Yachi S, Spehn EM, Hector A (2008) Light partitioning in experimental grass communities. Oikos 117:1351–1361
Weiner J (2004) Allocation, plasticity and allometry in plants. Perspect Plant Ecol 6:207–215
Wijesinghe DK, John EA, Hutchings MJ (2005) Does pattern of soil resource heterogeneity determine plant community structure? An experimental investigation. J Ecol 93:99–112
Williams BM, Houseman GR (2014) Experimental evidence that soil heterogeneity enhances plant diversity during community assembly. J Plant Ecol 7:461–469
Wubs ERJ, Bezemer TM (2016) Effects of spatial plant-soil feedback heterogeneity on plant performance in monocultures. J Ecol 104:364–376
Wubs ERJ, Bezemer TM (2017) Plant community evenness responds to spatial plant-soil feedback heterogeneity primarily through the diversity of soil conditioning. Funct Ecol 32:509–521
Acknowledgments
We thank Eddy De Smet, Eleni Meers, Evelyne Elst, Joanna Horemans, Marc Wellens, Niels Van Putte, Sigi Berwaers and Toon Ramsdonck for field assistance, and Joanna Horemans for statistical advice. This research was supported by National Natural Science Foundation of China (41571505) and Research Foundation – Flanders (FWO) (G.0490.16 N). Yongjie Liu holds a research grant from the China Scholarship Council (CSC).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Jeffrey Walck.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOCX 285 kb)
Rights and permissions
About this article

Cite this article
Liu, Y., De Boeck, H.J., Li, Z. et al. Unimodal relationship between three-dimensional soil heterogeneity and plant species diversity in experimental mesocosms. Plant Soil 436, 397–411 (2019). https://doi.org/10.1007/s11104-019-03938-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-019-03938-w