
Abstract
Aims
In this study, we aimed to understand effects and mechanisms of legume species and their relative abundance on soil N, species recruitment, and productivity in mixed grassland. We also assessed the utilization prospect of several legumes in natural grassland.
Methods
In 2006, grass-legume combinations (GLCs) with different ratios (GLR) were established using Leymus chinensis and four legume species (Medicago ruthenica, Lespedeza daurica, Medicago falcata, and Medicago sativa). In 2009, plant and soil samples were taken to examine the differences in soil nitrogen (N) properties, biological N fixation (BNF), recruitment characteristics, and aboveground biomass of the community under different mixture patterns.
Results
Soil total N and available N concentration increased when legume component increased from GLR 1:0 (grass monoculture) to GLR 1:1 (grass:legume 1:1) but decreased as legume component increased further from GLR 1:3 (grass:legume 1:3) to GLR 0:1 (legume monoculture). GLR 1:1 had the highest BNF in most GLCs except for L. chinensis-M. falcata. For any GLR with legume, L. chinensis-M. sativa combination (L-MS) had higher soil total N, available N, water content, and BNF. The legume combinations with a GLR lower than 1:1 resulted in more species recruitments compared with the grass monoculture, and L-MS induced relatively more species recruitments than other GLCs, which were correlated with improved soil water and NO3 −-N status. The grassland productivity increased as the GLR decreased; in any GLRs with legume, higher productivity was found under L-MS combination, and current results showed that grassland productivity was positively linked to soil NO3 −-N availability influenced by grass-legume mixture.
Conclusions
We concluded that the species and relative abundance of legumes had great impact on soil N status, species recruitment, and productivity in this temperate grassland ecosystem. In this study site, legume introduction can be considered as an alternative to N fertilization for increasing grassland productivity. M. sativa has the greatest economic and ecological potential species to be mixed into natural temperate steppe grassland, and grass-legume ratio 1:1 is the best combination in mixed communities.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
As a key element to support plant growth and build proteins, nitrogen (N) is a primary limiting factor for forage production and quality in grazing ecosystems (LeBauer and Treseder 2008; Karki et al. 2009). Traditionally, N fertilization has frequently been used to increase productivity and improve forage quality in grasslands (Schellberg et al. 1999). However, chemical fertilizers require a large monetary investment and do not benefit all grassland species due to interspecific differences in nutrient capture and utilization efficiency. Thus, chronic or intensive N fertilization can cause loss of plant species diversity, which maintains grassland ecosystem structure and functional stability (Suding et al. 2005; Clark and Tilman 2008; Bai et al. 2010). Historical and experimental evidence shows a trend of declining species diversity with N deposition and artificial N addition in native grassland ecosystems (Wedin and Tilman 1996; Gough et al. 2000).
Legumes are generally considered high quality forage due to their low fiber and high protein content (Mortenson et al. 2004). More importantly, it is well-known that legumes can increase N availability in soils by fixing atmospheric N in an available form (Spehn et al. 2002). The fixed N can be transferred to co-existing plants for utilization through root exudation (Lory et al. 1992; Paynel et al. 2008) and decomposition of dead roots (Trannin et al. 2000). Compared with N fertilization, N fixation from legumes can provide a more slow-released and persistent N source. This may reduce the competitive advantage of more efficient N consuming species and thus be beneficial for species coexistence. In addition, many legumes have well-developed shoot and root systems, which may maintain soil water content by increasing canopy shade and water retention (Armstrong et al. 1999). Moreover, legumes decompose quickly due to their high litter quality, which accelerates nutrient cycling and can improve soil productivity (Thomas and Asakawa 1993; Spehn et al. 2002; Milcu et al. 2008). Improved soil fertility can provide more niches for other plants to establish, which can be advantageous for maintaining higher species diversity and increasing resource complementarity (van Ruijven and Berendse 2005). Therefore, legumes can have huge economic and ecological potential as N donors to native grasslands.
Currently, inter-seeding legumes in native grasslands and establishing grass-legume mixed grasslands have been widely adopted to overcome the forage N deficit in grazing systems. Research across many sites has revealed that the introduction of legumes into grasslands leads to more soil N accumulation and higher productivity (Sleugh et al. 2000; Mortenson et al. 2004, 2005; Rao et al. 2007). These benefits from grass-legume mixtures depend on the legume species (Spehn et al. 2002; Fornara and Tilman 2008) and the trade-off with competition and facilitation effects for limiting resources between legumes and other plants (Lithourgidis et al. 2006). The introduction of different legume species and their relative abundance in mixed grasslands may influence soil N properties and community productivity due to the differences in N fixation and competitive effects of different species and mixtures. Kirwan et al. (2007) showed that grass-legume mixtures improved productivity greatly compared to grass only monocultures and that the effect of mixtures depended on the relative abundance of legumes based on results from 28 sites in European grassland. Nyfeler et al. (2009) found that mixtures fertilized with 50 kg N ha−1 year−1 produced equivalent forage yields to grass monocultures fertilized with 450 kg N ha−1 year−1 if the legume proportion was about 50 to 70 %. However, previous studies on mixed grasslands either had artificially controlled community structure (Spehn et al. 2002; Fornara and Tilman 2008) or only considered the effect of legumes on soil N status and productivity (Mortenson et al. 2004, 2005; Lithourgidis et al. 2006). Few studies revealed effects of legume presence, in particular legume species and their relative abundance, on species recruitment, coexistence, and subsequent species diversity in mixed grasslands (Carino and Daehler 2002), although this information would be very useful for introducing legumes as a resource in native grasslands.
Therefore, in this study, we aimed to a) understand effects and mechanisms of legume species and their relative density in grass-legume mixed grassland on soil N, species recruitment, and productivity and b) assess the utilization prospect of several legumes in natural grasslands. Our hypotheses were as follows: a) soil N and water status would be improved with increased legume density in mixed grasslands, b) increased relative density of legumes in mixed grasslands could promote species recruitment and enhance productivity through improving soil N and water status, and c) the benefit of grass-legume mixtures would depend on the legume species included in the mixture.
Materials and methods
Study site
This study was conducted at the Grassland Farming Research Station (E123°31′, N44°33′, elevation 145 m) of the Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, which is located at the eastern edge of the Eurasian steppe and is characterized as Eurasian continental temperate grassland. The site has a semi-arid continental climate. Mean annual temperature is 4.9 °C; annual precipitation is approximately 410 mm, with 70 % falling from June to September. The soil type is meadow chernozem soil. Soil pH is 8.1 with 16 g kg−1 organic matter and 1.1 g kg−1 total N in the 0–30 cm depth. The climax vegetation type is meadow steppe dominated by Leymus chinensis, a perennial C3 rhizome grass, which accounts for 80 % of the aboveground biomass. However, from 1980s, part of the L. chinensis meadow had been reclaimed as cropland due to the increasing economic benefit from grain production. Our experiment was conducted in a section of the meadow that was converted to cropland in 1995, but subsequently abandoned in 2002.
Field experiment design
The experiment was a split-plot design with four grass-legume combinations (GLCs) as main plots and five grass-legume mixture ratios as sub-plots randomized allocated in the main plots, with four replicates. The four GLCs were L. chinensis-Medicago ruthenica (L-MR), L. chinensis-Lespedeza daurica (L-LD), L. chinensis-Medicago falcata (L-MF), and L. chinensis-Medicago sativa (L-MS). Under each L. chinensis-legume combination, total plant density was 600 plant individuals per square meter, which reflected the mean plant density of the natural L. chinensis grassland in this region. The grass-legume ratios (GLRs) in the mixtures were 1:0 (grass monoculture), 3:1 (3 grass:1 legume), 1:1 (1 grass:1 legume), 1:3 (1 grass:3 legume), and 0:1 (legume monoculture), respectively, by altering the sowing rate and thinning the seedlings later.
On 5 July 2006, after vegetation was removed and the soil was turned over at a depth of 0–30 cm, all 80 plots with 3 × 3 m in size were sown with the proper number of L. chinensis and legume seeds adjusted with actual seed germination rate. In each plot, all sown seeds were mixed then uniformly broadcasted into plots in strips with row space of 15 cm. Before sowing, all legume seeds were soaked in 98 % H2SO4 for 0.5 h to soften seed coats and ensure viable germination. No inoculant was applied when sown assuming the soil contains enough rhizobia as the site had history of legume growing which was confirmed by the pot experiment described below. Plots were irrigated when necessary to ensure emergence and survival of the sown species. On 5 August 2006, we thinned the plots to preserve the targeted plant species densities and GLRs. From seeding date to June 2007, plots were kept weed-free by hand-weeding to ensure successful establishment of L. chinensis and the sown legumes and to eliminate the impact of the original soil seed bank on species recruitment.
Pot experiment for legume nodulation
Nodulation of the four legume species were confirmed by growing them in pots with soil from the same field as the experimental grassland. Soil at 0–40 cm depth was collected in field experiment site and then to be passed through a 2-mm sieve. A 15-kg sieved soil was filled into a 30 × 35 cm (diameter × height) pot, with a total of 24 pots. Each legume species was planted into randomly selected six plots with 40 acid treatment seeds in each pot. Pots were randomly placed in greenhouse, and their positions were exchanged every 3 days. For keeping soil moisture, equal water was sprayed in each pot every 3 days if needed. Emerged seedlings were thinned to retain 15 plants in each pot. At full flowering, the soil in each pot was washed to harvest shoot and root of each plant. Total and effective (pink or light pink) nodule number were counted, and nodule fresh weight was determined in each pot.
Plant sampling and analysis
In early August 2009, a 1 × 1 m quadrat was arranged in the center of each plot, and plants were counted and identified to species in each quadrat. The aboveground part of each species was clipped at the soil surface and taken back to the laboratory in paper bags. All newly present species in the plots that were not from the initially sown species pool were defined as recruited species. Plant materials were oven-dried at 65 °C for 48 h and weighed to determine the dry biomass. Shoot samples of sown legumes and non-legume species L. chinensis, Hierochloe glahra, Setaria viridis, and Kalimeris integrifoliain in grass monocultures were finely ground to determine the biological N fixation (BNF) rate using 15 N natural abundance technique (Unkovich et al. 2008). BNF from sown legumes was estimated using the 15 N natural abundance method. The percent of N derived from the atmosphere (%Ndfa) in legume biomass was estimated using the following formula (Unkovich et al. 2008):
To avoid possible N transfer between legumes and non-legumes species, we used L. chinensis, H. glahra, S. viridis, and K. integrifolia in grass monocultures as the reference plants. These four species were all common species in mixture and grass monoculture communities. They represent three function groups including C3 grass, C4 grass, and forb and likely reflect different N uptake characteristics. The B value for legume species were determined by growing them under N-free medium (sand + perlite + nutrient solution) inoculated with a soil suspension from experiment site in temperature-controlled glasshouse and harvesting shoot material for 15 N analysis at a stage of growth that is same as our field analyses (Unkovich et al. 2008). Total BNF was calculated based on % N from fixation, legume shoot biomass, and shoot N concentration. Total N was analyzed using the continuous flow Isotope Ratio Mass Spectrometer (Stable Isotope Facility, UC Davis).
Soil sampling and analysis
After all plant material was collected, we used an auger (4 cm in diameter) to collect three soil samples at depths of 0–40 cm in each quadrat. These soil samples were sealed into three previously weighed aluminum containers and then taken back to laboratory to determine their fresh weight and then the dry weight after being oven-dried at 105 °C for 48 h. Soil water content was calculated based on the soil’s fresh and dry weight. Three more soil cores were collected and bulked at depths of 0–40 cm in each quadrat. With visible plant materials and other debris removed, part of mixed soil samples were frozen at 0 °C, and the remaining samples were air-dried in the dark and then ground to pass through a 0.2-mm sieve for subsequent analysis. NH4 + and NO3 − were extracted with 2 M KCl from frozen soil samples (Miller and Keeney 1982), and their concentrations were determined by a Bran-Luebbe AA3 autoanalyzer (Bran and Luebbe, Hamburg, Germany). Soil total nitrogen (TN) was measured by the Kjeldahl method using air-dried soil samples (Sparks et al. 1996). All soil nutrient traits were calculated based on units of dry soil weight.
Data analysis
All data were assessed to verify model assumptions of normality and equality of variance. Three-way ANOVA based on a split-plot design was conducted to determine the main and interactive effects of GLC and GLR with block as random factor. Means comparisons were conducted by one-way ANOVA, followed by Duncan’s HSD test. Stepwise multiple regression analysis was conducted to examine the dependence of species recruitment on soil N availability and water content, with species recruitment characteristics as dependent variables and soil NH4 +-N, NO3 −-N, and water content as independent variables. A similar stepwise multiple regression analysis was performed to examine the dependency of aboveground biomass (AGB) on species number, soil N availability, and water content. Significance for all statistical tests were evaluated at P = 0.05. All data analyses were conducted with the SPSS 16.0 software (Chicago, IL, USA).
Results
Soil N and water properties
Both the GLC and GLR had significant impact on the soil available N, NO3 −-N, and water content. In addition, GLR had a significant effect on soil NH4 +-N and total N (Table 1). Across all GLCs, the soil total N, available N, and its components at the depth of 0–40 cm decreased in the order: GLR 1:1 > GLR 1:3 > GLR 0:1 > GLR 3:1 > GLR 1:0. Soil available N and its components had significantly higher values under GLR 1:1 compared with the GLR 1:0, GLR 3:1, and GLR 0:1 treatments (Table 2). In all GLCs, the mixture treatments had higher soil water content than monoculture, particularly GLR 3:1, under which soil water content was significantly higher than both grass and legume monocultures (Table 2). For any GLR treatment with legumes, the highest values of soil total N, available N, and water content were found in L-MS and L-MR combinations, and the lowest values in L-MF combination (Table 2).
Legume nodulation
In pot experiment, the MS had significant higher total and effective nodule number compared with other three legume species (Fig. 1a). The total nodule number was 29 %, 45 %, and 90 % higher in MS than MR, LD, and MF, respectively, with the effective nodule number being 27 %, 48 %, and 103 % higher in MS than MR, LD, and MF, respectively. The fresh weight of total and effective nodule showed similar interspecific difference with nodule number (Fig. 1b). However, there was no significant difference in total plant biomass between four legume species (Fig. 1c), indicating that the enhanced nodulation in MS compared to the other legumes was specific nodulation response but did not correlate with enhanced growth.

Nodule number (a) and fresh weight (b) of four legume species and plant biomass (c) of four legume species in plots experiment. Values are shown as means + SE. Different lowercase letters above bars indicate significant differences of effective nodule (P < 0.05) between legume species. Different uppercase letters above bars indicate significant differences of total nodule (P < 0.05) between legume species in Fig. 1a, b. NS no significant difference
Shoot δ15N of legumes and BNF
The δ15N values of legume species increased with enhanced proportion of legumes in the mixtures for all GLCs. The δ15N values of L. chinensis in mixture communities were significantly lower than grass monoculture with the lowest values in GLR 1:1, indicating that L. chinensis taken up N transferred from legumes when grown together with legumes, and more N was transferred from legume to L. chinensis under GLR 1:1 compared to other GLRs. The %Ndfa and BNF of legumes showed similar trend for GLC and GLR with different reference species. Compared with reference species, L. chinensis and K. integrifolia had relatively higher estimate of %Ndfa and BNF compared with H. glahra and S. viridis (Table 3). Increasing relative legume density significantly decreased %Ndfa of legumes. Averaged across GLC, the GLR 1:1 fixed significantly more nitrogen than GLR 3:1 treatments, and GLR 1:1 had the highest BNF compared to other GLRs in most GLCs except L-MF. For any given GLR, L-MS combination fixed more atmosphere N, especially compared to L-MF; L-MS combination had significant higher BNF (Tables 1 and 3).
L. chinensis and sown legumes
The plant density of L. chinensis significantly declined with decreased GLR, while the plant density of sown legumes significantly increased in all communities (Tables 1 and 4). Moreover, the plant density of L. chinensis significantly increased under GLR 1:0, 3:1, and 1:1 treatments compared with initial sowing density in all L. chinensis-legume combinations (all P < 0.05; Table 4). The aboveground biomass of L. chinensis and sown legumes also depended on GLR and GLC (Table 1). The AGB of L. chinensis significantly declined in communities as GLR decreased, in contrast to a statistically significant increase of AGB of sown legume species (Table 4). Under mixed treatments, the aboveground biomass of L. chinensis was higher in L-MS and L-MR than L-LD and L-MF combinations.
Species recruitment
GLC, GLR, and their interaction had statistically significant impacts on species recruitment as measured by the number of recruited species and their aboveground biomass (Table 1). The recruitment of legume species only occurred in communities with initial GLR ≤ 1, and the total plant density of recruited legumes was less than 4 plants m−2 (Table 5). All recruited species are commonly found in the natural steppe grassland, but none of the species recruited were dominated species. For all GLCs, the number of recruited species significantly increased from treatment with GLR 1:0 to GLR 1:1 and then significantly decreased from GLR 1:1 to GLR 0:1 (Table 6). The plant density and AGB of recruited species were significantly higher under GLR 3:1 and GLR 1:1 compared with other GLR treatments in most GLCs excluding L-MF (Table 6). For all GLR treatments with legumes, plant density and AGB of recruited species were lower in the L-MF combination compared with other combinations (Table 6). Stepwise multiple regression analysis showed that the number of recruited species positively correlated to soil water and NO3 −-N content. The combination of soil water and NO3 −-N content explained the 44.5 % variation in the number of recruited species (Table 7). The plant density and AGB of recruited species only had a significant correlation with soil water content, which explained 35.8 % and 39.5 % of the variations in plant density and AGB of recruited species, respectively (Table 7).
Aboveground biomass of the community
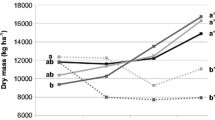
Aboveground biomass (AGB) varied greatly under different GLC and GLR (Table 1). Across all L. chinensis-legume combinations, the AGB significantly increased with decreased GLR. However, no significant difference in AGB was found between GLR 1:1 and 0:1 (Fig. 2). In all GLR treatments with legumes, AGB of the community decreased in the following order: L-MS > L-LD > L-MR > L-MF (Fig. 2). AGB was positively correlated to soil NO3 −-N content, which explained 44.3 % variations in AGB of the community (Table 7).

Aboveground biomass (AGB) of the community under different L. chinensis-legume combinations and L. chinensis-legume ratios. Values are shown as means + SE
Discussion
Effects of grass-legume ratios on soil N
Until now, the impact of legumes proportion on soil N in a mixed temperate steppe grassland was not well-documented, even though several studies examined effects of legume proportion on N transfer, productivity, soil moisture, and soil carbon flux (Nyfeler et al. 2009, 2011; Zhang et al. 2013). Our results showed that soil total N and available N increased with GLR ranging from 1:0 to 1:1 and then decreased as the proportion of legumes further increased. This indicates that effects of grass-legumes mixtures on soil N depend on the relative abundance of legumes in this grassland, and that GLR 1:1 was the most ideal density relationship between L. chinensis and legumes for improving soil N status.
The difference in BNF is the major driving factor for the changed soil N status under the same site condition and fertilization management. Our results showed that BNF and soil N had similar responses to different GLRs. One of the possible explanations for the change of soil N status with GLR was that, in grass-legume mixed grasslands, decreased GLR may initially enhance BNF due to increased legume density and stimulating effects of grass species by N transfer (see δ15N values of L. chinensis in Table 3) (Hartwig 1998; Ehrenfeld 2003; Schipanski and Drinkwater 2012). For example, Ledgard et al. (1985) showed that the N fixation of Trifolium repens increased by 20 % when mixed into ryegrass grassland. However, when legume density is excessive, the intraspecific competition increased, resulting in less interaction between grass and legume in N utilization, which may inhibit BNF from legume (Ledgard and Steele 1992), and induce the physiological transformation from fixation of atmospheric N to uptake of soil N, consequently decreasing N fixation and soil N status (Tables 2 and 3). Furthermore, soil water content showed similar patterns as BNF in response to GLR in our experiment. Thus, GLR may impact BNF indirectly by regulating soil water content. Kowalenko and Cameron (1976) and Austin et al. (2004) reported that improved soil water status can enhance BNF of legumes. Therefore, initially increased legume density may promote BNF by increasing soil water content (Armstrong et al. 1999). However, relatively excessive legume density may result in more water transpiration due to larger shoot and leaf structures of the four legumes associated with L. chinensis. This could cause soil water to become a limiting factor for BNF, consequently inhibiting the potential increase in soil N (Sprent and Bradford 1977).
Effects of grass-legume ratios on species recruitment
In arid and semi-arid ecosystems, soil water may also be an important limiting factor for species recruitment (Brown and Archer 1990; Carino and Daehler 2002). With regards to the modifying effects of legumes on soil N and water status (Armstrong et al. 1999), we expected that increased relative abundance of legumes in this grassland would promote species recruitment. Our results showed that the introduction of legumes into this grassland increased all recruitment characteristics values when the GLR was lower than one in all L. chinensis-legume combinations; however, the legume monocultures showed inhibition effects on plant density and aboveground biomass of recruited species. Further regression analysis indicated that soil water content was the only control factor for all recruitment characteristics. This means that in this semi-arid grassland ecosystem, soil water content rather than soil N may be a more important environment factor to determine which species can successfully establish in a community, and legume abundance may promote or restrict species recruitment through regulating soil water status when introduced into grasslands (Brown and Archer 1990; Armstrong et al. 1999). However, when soil water content was excluded, we still found that soil NO3 −-N significantly correlated with the number of recruited species. Experimental studies have shown that a higher NO3 −-N availability promotes recruitment, while communities with lower levels of NO3 −-N are often more resistant to the establishment of recruited species (Fargione and Tilman 2005; Roscher et al. 2009). In addition, previous studies showed that N enrichment may enhance plant tolerance to limited water through physiological regulation (Saneoka et al. 2004; Wu et al. 2008). Therefore, soil NO3 −-N may indirectly impact community recruitment by regulating the drought adaptability of recruited communities in this grassland ecosystem.
In our study, the density of sown legumes and L. chinensis in all mixed treatments remained steady following recruitment of new species, as compared with initial values, and significantly increased under GLR 3:1 and 1:1 in all GLCs. This indicates that the replacement of L. chinensis with legumes does not induce the passively competitive outcome for L. chinensis and legumes, and appropriate GLR may promote the development of a sustainable L. chinensis and legume population, while increasing species richness.
Effects of grass-legume ratios on grassland productivity
Nitrogen is a main limiting factor for grassland productivity (Karki et al. 2009). It has been widely reported that introducing legumes into grasslands leads to higher soil N availability and productivity (Sleugh et al. 2000; Mortenson et al. 2004; Rao et al. 2007). Our study demonstrated that the increased relative abundance of legumes persistently increased aboveground productivity of a semi-arid grassland, which was partly attributed to higher individual biomass of legumes compared with L. chinensis (Table 4), and also resulted from BNF by legumes (Table 3). The increased N input from legumes improved soil N availability and ultimately increased grassland productivity (Table 2). Nyfeler et al. (2009) similarly showed that the introduction of legumes induced consistently higher production compared with grass monocultures in European grassland. The facilitation effect of a grass-legume mixture on grassland productivity was more intense under low soil N availability (Nyfeler et al. 2009).
In the current study, the GLR1:1 treatment had 50–60 % greater aboveground production than a grass monoculture. Alternatively, grassland production can also be improved by N fertilization. In the same steppe region of the current study, Bai et al. (2010) showed that 3 years of N fertilization at 10.5 g m−2 year−1 in this mature grassland dominated by L. chinensis increased aboveground production approximately 50 % compared with no fertilization (the AGB was around 300 g m−2). However, in grassland ecosystems, species diversity loss induced by N enrichment has become a concern (Wedin and Tilman 1996; Gough et al. 2000; Suding et al. 2005; Clark and Tilman 2008). Increased productivity by applying N fertilizer would be at a cost of loss of species richness. However, if the same forage production is obtained by introducing legumes into the grassland, the species richness may increase because of the facilitation effect of legumes on species recruitment. Therefore, as an alternative to N fertilization, introducing legumes in a temperate steppe grassland with GLR 1:1 can be a better choice to maximize the benefits of forage production and diversity conservation.
Species difference in grass-legume mixtures
Different species have different impacts on community recruitment and productivity as certain species drives specialized interspecific interaction (Vilà and Weiner 2004; Lankau et al. 2009), influences soil nutrient cycling (Liao et al. 2008), and possibly results in a sampling effect (Vilà et al. 2011). Our results showed that GLC had a significant impact on species recruitment and grassland productivity, which can partly be attributed to different effects of the four legume species on BNF, soil water, and N status (Table 2; Table 3; Table 7). The current experiment showed that L-MS had a higher N fixation compared with other GLCs which may result from relatively higher nodulation response and population biomass of M. sativa (Fig. 1; Table 4). Meanwhile, M. sativa showed stronger water retention capacity than the other three legumes when mixed with L. chinensis (Table 2). These factors helped to promote species recruitment and grassland productivity by improving soil N and water status. Considering the total effects of GLC on soil N, species recruitment, and grassland productivity, we suggest that M. sativa has more advantages for introduction into grasslands to enhance forage production and maintain species diversity, with L. daurica and M. ruthenica following, with M. falcata performing the worst.
Critical reflection about the assessment method of legume nodulation and %Ndfa
We used soil from the field site to assess legume nodulation in a pot experiment as it would be very hard to collect intact legume root for nodulation measurement in field without destructive harvest of the sample plots. However, the growth conditions in greenhouse indeed can never perfectly mimic the conditions in field regarding factors such as temperature, moisture, and competition for soil nutrient and water depending on species composition and plant development. Therefore, our nodulation data may not absolutely represent actual field values. In our pot experiment, we kept lower plant density and higher soil moisture compared with field condition. The attained legume nodulation data may reflect the inherent differences of nodulation response for four legume species in non-competition condition. Therefore, the species specific pattern of these nodulation data can be valuable for understanding the similar difference of BNF between legume species.
15 N natural abundance method is commonly used to estimate %Ndfa (Carlsson et al. 2009; Frankow-Lindberg and Dahlin 2013). The accuracy of the natural abundance technique ultimately depends upon the magnitude of δ15N in the plant-available soil N pool which can be reflected by δ15N value of reference plant (Unkovich et al. 2008). Generally, higher δ15N concentration of plant-available soil N causes lower BNF estimate error (Ledgard and Steele 1992). Unkovich et al. (1994) recommended 2‰ as the lowest δ15N value of reference plant for detecting a reliable %Ndfa. In our results, the δ15N values of reference plants ranged 3.74–4.09‰, which met the lowest δ15N value requirement suggested by Unkovich et al. (1994). The use of correct B value also is crucial for the accuracy of the 15 N natural abundance method (Unkovich et al. 2008; Carlsson et al. 2009). Some published B values can be chosen for Medicago spp. but these B values frequently had difference in different literatures (Unkovich et al. 2008). To attain accurate values of %Ndfa, here we generated our own B values for all four legume species in a glasshouse experiment. An assumption of the 15 N natural abundance method is that the δ15N of reference plants identically reflects the δ15N of soil N utilized by the legume. Chosen reference species influences the calculation of %Ndfa, especially when the δ15N of soil mineral N changes markedly with depth or fluctuates during the growing season because different reference species may have different N uptake and isotopic fractionation characteristics during utilization of soil N (Ledgard and Steele 1992; Unkovich et al. 2008). Since it is nearly impossible to know the “true” δ15N of the soil N available to the legume and equally difficult to identify one optimal reference species, it has been suggested that the best choice is to use several reference species (Unkovich et al. 2008; Carlsson et al. 2009). In current study, we selected four species as reference plants which likely reflected different N uptake and isotopic fractionation characteristics. We calculated the %Ndf using each reference species and also calculated a mean %Ndf (Table 3). The data showed some differences in %Ndfa when different reference δ15N values were used, especially when %Ndfa was at low level. If direct N transfer from legumes to non N-fixing plants occurs, a choice of reference plants growing in close association with legumes may greatly underestimate the %Ndfa estimates (Ledgard and Steele 1992; Unkovich et al. 2008). In a previous study at the same site, it appeared that the direct N transfer from legume to neighboring plants had occurred (unpublished data). In the current study, the significant difference in δ15N of L. chinensis between monoculture and mixture plots also indicated that direct N transfer from legume to grass had likely occurred; therefore, we used plants in grass monocultures as reference plants. However, this may impose other errors since it fails to accurately mimic the δ15N of the soil N utilized by legume (Unkovich et al. 2008; Carlsson et al. 2009). In fact, even though there is evidence for direct N transfer from legumes to reference plants in study sites, it does not exclude the possibility for N transfer in other directions, which is an argument for using reference plants grown together with the legume (Carlsson et al. 2009). For example, Carlsson and Huss-Danell (2014) indicated that N transfer not only occur from legumes to neighboring non-legume plants but also in the opposite direction, from non-legumes to legumes and partial N transferred from a legume plant should be considered equally available for re-uptake by the legume plant itself as for neighboring legume and non-legume plants. This makes the error weak for using non-legume plants in mixtures to reflect 15 N availability in soil for neighboring legume. In contrast, N2 fixation may be overestimated by using a reference plant in pure stand (Carlsson and Huss-Danell 2014). Based on these studies, during N2 fixation estimate, a precise choice for reference plants should be done after considering the directions and amount of N transfer.
Conclusions
Introducing legumes in a temperate steppe grassland dominated by L. chinensis can improve soil N and water status. However, effects of grass-legume mixtures depend on the relative abundance of legumes, with the GLR 1:1 inducing the most positive effects on soil N and water contents. Appropriate GLR can promote species recruitment by enhancing soil water and NO3 −-N content, consequently increasing the species diversity of a grassland. Aboveground production of plant communities persistently increased with relative abundance of legumes, which was correlated to increased soil NO3 −-N availability. The different legume species tested had different effects on soil N, species recruitment, and grassland productivity in the grass-legume mixtures. In general, M. sativa had the most positive effects on soil N, species recruitment, and grassland productivity when mixed with L. chinensis.
In conclusion, in this semi-arid steppe grassland, legume introduction can be considered an alternative to nitrogen fertilization for increasing grassland productivity without compromising grassland species diversity. M. sativa has the greatest economic and ecological potential to be mixed into grasslands, and 1:1 ratio for grass and legume is the best ratio in mixed communities.
References
Armstrong RD, Kuskopf BJ, Millar G, Whitbread AM, Standley J (1999) Changes in soil chemical and physical properties following legumes and opportunity cropping on a cracking clay soil. Anim Prod Sci 39(4):445–456
Austin AT, Yahdjian L, Stark JM, Belnap J, Porporato A, Norton U, Ravetta DA, Schaeffer SM (2004) Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 141(2):221–235
Bai YF, Wu JG, Clark CM, Naeem S, Pan QM, Huang JH, Zhang LX, Han XG (2010) Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from inner Mongolia Grasslands. Glob Chang Biol 16(1):358–372
Brown JR, Archer S (1990) Water relations of a perennial grass and seedling vs adult woody plants in a subtropical savanna, Texas. Oikos 57(3):366–374
Carino DA, Daehler CC (2002) Can inconspicuous legumes facilitate alien grass invasions? Partridge peas and fountain grass in Hawaii. Ecography 25(1):33–41
Carlsson G, Huss-Danell K (2014) Does nitrogen transfer between plants confound 15N-based quantifications of N2 fixation? Plant Soil 374(1–2):345–358
Carlsson G, Palmborg C, Jumpponen A, Scherer-Lorenzen M, Högberg P, Huss-Danell K (2009) N2 fixation in three perennial Trifolium species in experimental grasslands of varied plant species richness and composition. Plant Ecol 205(1):87–104
Clark CM, Tilman D (2008) Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 451:712–715
Ehrenfeld JG (2003) Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems 6(6):503–523
Fargione J, Tilman D (2005) Diversity decreases invasion via both sampling and complementarity effects. Ecol Lett 8(6):604–611
Fornara DA, Tilman D (2008) Plant functional composition influences rates of soil carbon and nitrogen accumulation. J Ecol 96(2):314–322
Frankow-Lindberg BE, Dahlin AS (2013) N2 fixation, N transfer, and yield in grassland communities including a deep-rooted legume or non-legume species. Plant Soil 370(1–2):567–581
Gough L, Osenberg CW, Gross KL, Collins SL (2000) Fertilization effects on species density and primary productivity in herbaceous plant communities. Oikos 89(3):428–439
Hartwig UA (1998) The regulation of symbiotic N2 fixation: a conceptual model of N feedback from the ecosystem to the gene expression level. Perspect Plant Ecol 1(1):92–120
Karki U, Goodman MS, Sladden SE (2009) Nitrogen source influences on forage and soil in young southern-pine silvopasture. Agric Ecosyst Environ 131(1):70–76
Kirwan L, Lüscher A, Sebastia MT, Finn JA, Collins RP, Porqueddu C, Helgadottir A et al (2007) Evenness drives consistent diversity effects in intensive grassland systems across 28 European sites. J Ecol 95(3):530–539
Kowalenko CG, Cameron DR (1976) Nitrogen transformations in an incubated soil as affected by combinations of moisture content and temperature and adsorption-fixation of ammonium. Can J Soil Sci 56(2):63–70
Lankau R, Nuzzo V, Spyreas G, Davis AS (2009) Evolutionary limits ameliorate the negative impact of an invasive plant. PNAS 106(36):15362–15367
LeBauer DS, Treseder KK (2008) Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 89(2):371–379
Ledgard SF, Steele KW (1992) Biological nitrogen fixation in mixed legume/grass pastures. Plantsoil 141(1–2):137–153
Ledgard SF, Simpson JR, Freney JR, Bergersen FJ (1985) Field evaluation of 15N techniques for estimating nitrogen fixation in legume-grass associations. Crop Pasture Sci 36(2):247–258
Liao CZ, Peng RH, Luo YQ, Zhou XH, Wu XW, Fang CM, Chen JK, Li B (2008) Altered ecosystem carbon and nitrogen cycles by plant invasion: a meta-analysis. New Phytol 177(3):706–714
Lithourgidis AS, Vasilakoglou IB, Dhima KV, Dordas CA, Yiakoulaki MD (2006) Forage yield and quality of common vetch mixtures with oat and triticale in two seeding ratios. Field Crop Res 99(2):106–113
Lory JA, Russelle MP, Heichel GH (1992) Quantification of symbiotically fixed nitrogen in soil surrounding alfalfa roots and nodules. Agron J 84(6):1033–1040
Milcu A, Partsch S, Scherber C, Weisser WW, Scheu S (2008) Earthworms and legumes control litter decomposition in a plant diversity gradient. Ecology 89(7):1872–1882
Miller RH, Keeney DR (1982) Methods of soil analysis. Part 2: chemical and microbiological properties, 2nd edn. American Society of Agronomy, Soil Science Society of America, Madison
Mortenson MC, Schuman GE, Ingram LJ (2004) Carbon sequestration in rangelands interseeded with yellow-flowering alfalfa (Medicago sativa ssp. falcata). Environ Manag 33(1):475–481
Mortenson MC, Schuman GE, Ingram LJ, Nayigihugu V, Hess BW (2005) Forage production and quality of a mixed-grass rangeland interseeded with Medicago sativa ssp. falcata. Rangel Ecol Manag 58(5):505–513
Nyfeler D, Huguenin-Elie O, Suter M, Frossard E, Connolly J, Lüscher A (2009) Strong mixture effects among four species in fertilized agricultural grassland led to persistent and consistent transgressive overyielding. J Appl Ecol 46(3):683–691
Nyfeler D, Huguenin-Elie O, Suter M, Frossard E, Lüscher A (2011) Grass-legume mixtures can yield more nitrogen than legume pure stands due to mutual stimulation of nitrogen uptake from symbiotic and non-symbiotic sources. Agric Ecosyst Environ 140(1):155–163
Paynel F, Lesuffleur F, Bigot J, Diquélou S, Cliquet JB (2008) A study of 15N transfer between legumes and grasses. Agron Sustain Dev 28(2):281–290
Rao SC, Northup BK, Phillips WA, Mayeux HS (2007) Interseeding novel cool-season annual legumes to improve bermudagrass paddocks. Crop Sci 47(1):168–17
Roscher C, Beßler H, Oelmann Y, Engels C, Wilcke W, Schulze ED (2009) Resources, recruitment limitation and invader species identity determine pattern of spontaneous invasion in experimental grasslands. J Ecol 97(1):32–47
Saneoka H, Moghaieb REA, Premachandra GS, Fujita K (2004) Nitrogen nutrition and water stress effects on cell membrane stability and leaf water relations in Agrostis palustris Huds. Environ Exp Bot 52(2):131–138
Schellberg J, Möseler BM, Kühbauch W, Rademacher IF (1999) Long-term effects of fertilizer on soil nutrient concentration, yield, forage quality and floristic composition of a hay meadow in the Eifel mountains, Germany. Grass Forage Sci 54(3):195–207
Schipanski ME, Drinkwater LE (2012) Nitrogen fixation in annual and perennial legume-grass mixtures across a fertility gradient. Plant Soil 357(1–2):147–159
Sleugh B, Moore KJ, George JR, Brummer EC (2000) Binary legume–grass mixtures improve forage yield, quality, and seasonal distribution. Agron J 92(1):24–29
Sparks DL, Page AL, Helmke PA, Loeppert RH, Soltanpour PN, Tabatabai MA, Johnston CT, Sumner M (1996) Methods of soil analysis. Part 3: chemical methods. American Society of Agronomy, INC, Madison, pp 1085–1121
Spehn EM, Scherer-Lorenzen M, Schmid B, Hector A (2002) The role of legumes as a component of biodiversity in a cross-European study of grassland biomass nitrogen. Oikos 98(2):205–218
Sprent JI, Bradford AM (1977) Nitrogen fixation in field beans (Vicia faba) as affected by population density, shading and its relationship with soil moisture. J Agric Sci 88:303–310
Suding KN, Collins SL, Gough L, Clark CM, Cleland EE, Gross KL, Milchunas DG, Pennings S (2005) Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. PNAS 102(12):4387–4392
Thomas RJ, Asakawa NM (1993) Decomposition of leaf litter from tropical forage grasses and legumes. Soil Biol Biochem 25(10):1351–1361
Trannin WS, Urquiaga S, Guerra G, Ibijbijen J, Cadisch G (2000) Interspecies competition and N transfer in a tropical grass-legume mixture. Biol Fertil Soils 32(6):441–448
Unkovich MJ, Pate JS, Sanford P, Armstrong EL (1994) Potential precision of the δ15N natural abundance method in field estimates of nitrogen fixation by crop and pasture legumes in Southwest Australia. Crop Pasture Sci 45(1):119–132
Unkovich M, Herridge D, Peoples M, Cadish G, Boddey B, Giller K, Alves B, Chalk P (2008) Measuring plant-associated nitrogen fixation in agricultural systems. Australian Centre for International Agricultural Research, Canberra
van Ruijven J, Berendse F (2005) Diversity–productivity relationships: initial effects, long-term patterns, and underlying mechanisms. PNAS 102(3):695–700
Vilà M, Weiner J (2004) Are invasive plant species better competitors than native plant species? Evidence from pair-wise experiments. Oikos 105(2):229–238
Vilà M, Espinar JL, Hejda M, Hulme PE, Jarošík V, Maron JL, Pergl J, Schaffner U, Sun Y, Pyšek P (2011) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecol Lett 14(7):702–708
Wedin DA, Tilman D (1996) Influence of nitrogen loading and species composition on the carbon balance of grasslands. Science 274(5293):1720–1723
Wu F, Bao W, Li F, Wu N (2008) Effects of drought stress and N supply on the growth, biomass partitioning and water-use efficiency of Sophora davidii seedlings. Environ Exp Bot 63(1):248–255
Zhang ZH, Duan JC, Wang SP, Luo CY, Zhu XX, Xu BRBY, Chang XF, Cui SJ (2013) Effects of seeding ratios and nitrogen fertilizer on ecosystem respiration of common vetch and oat on the Tibetan plateau. Plant Soil 362(1–2):287–299
Acknowledgments
This study was supported by China’s National Key Basic Research Program (2011CB403203) and Strategic Science and Technology Guide Project of CAS (XDA05050401).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Euan K. James.
Rights and permissions
About this article

Cite this article
Li, Q., Song, Y., Li, G. et al. Grass-legume mixtures impact soil N, species recruitment, and productivity in temperate steppe grassland. Plant Soil 394, 271–285 (2015). https://doi.org/10.1007/s11104-015-2525-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-015-2525-2