
Abstract
Purpose
To explain the effects of the osmolyte proline on the protein-protein interactions (PPI), viscosity and stability of highly concentrated antibody solutions in contrast to other neutral osmolytes.
Methods
The viscosity of ~225 mg/mL mAb solutions was measured with proline, glycine and trehalose as a function of pH and co-solute concentration up to 1.3 M. The stability was assessed via turbidity as well as size exclusion chromatography after 4 weeks storage at 40°C. The PPI strength was assessed qualitatively via the high concentration diffusion rate by dynamic light scattering.
Results
Increasing proline significantly reduced the mAb viscosity and increased the colloidal stability at pH 6, but not at pH 5 further from the mAb pI. In contrast, glycine and trehalose did not improve the viscosity nor stability. The normalized diffusion coefficient at high concentration, which is inversely proportional to the attractive PPI strength, increased with proline concentration but decreased with increasing glycine.
Conclusions
Proline demonstrated greater efficacy for improving mAb viscosity and stability in contrast to glycine and trehalose due to its amphipathic structure and partial charge on the pyrrolidine side chain. These properties likely allow proline to screen the attractive electrostatic and hydrophobic interactions that promote self-association and high viscosities. Binary proline-histidine formulations also demonstrated greater viscosity reduction effects than histidine alone at the same total co-solute concentration, while maintaining a lower total solution osmolarity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Formulation strategies to reduce the viscosity and enhance the stability of highly concentrated monoclonal antibody solutions are of great interest in the field of drug delivery. For the preferred delivery route of subcutaneous injection, antibodies often must be formulated at concentrations of 200 mg/mL or higher in order to deliver the required dosage in a limited injection volume of 1–1.5 ml (1). However, attractive short-ranged protein-protein interactions (PPI) become dominant at these concentrations (2,3) due to small spacings between protein molecules on the order of the molecular diameter (2). These interactions can lead to self-association (3) and high viscosities above the 15–20 cP limit for subcutaneous injection (1). The concentration-dependent viscosity increase is thought to arise from reversible protein self-association via attractive multibody interactions mediated by short-ranged PPI (4,5). Strategies to modify or disrupt these oligomers and reduce the solution viscosity therefore require an understanding of the complex landscape of PPI at high concentrations (6).
The viscosity and colloidal stability of concentrated protein solutions is determined by an intricate balance of long-ranged and local anisotropic electrostatic and hydrophobic interactions. In addition to long-ranged electrostatic repulsion between molecules due to the protein net charge (7), protein molecules in close proximity also experience strong, short-ranged anisotropic electrostatic attraction from the alignment of local charges and dipoles (8,9) or via charge-dipole and dipole-dipole interactions (7,10). As the magnitude of the difference between the pH and isoelectric point increases, the long ranged electrostatic repulsion becomes stronger. The local electrostatic attraction also becomes weaker, as more of the charges on the protein surface have the same sign, resulting in a smaller probability of attractive electrostatic contact between oppositely charged residues (7). Unfavorable hydrophobic interactions also promote self-association that may result in higher solution viscosities (11).
A strategy for lowering the solution viscosity is to weaken the attractive PPI and disrupt protein self-association. In some cases this has been accomplished by charge screening via salt addition (10). For example, chaotropic salts have been found to afford greater viscosity reductions than neutral salts such as NaCl (12), as the greater disorder in the water structure weakens hydrophobic PPI (13). Similarly, hydrophobic salts in some cases have produced marked viscosity reductions in concentrated antibody solutions (11) by charge screening and also weakening of the hydrophobic interactions. It is also possible to reduce the viscosity by modifying the hydrophobic interactions with uncharged hydrophobic amino acids such as glycine, alanine and phenylalanine (14). Charged amino acids such as protonated arginine and histidine have been found to markedly improve the viscosity of antibodies at high concentration (2,15) in addition to suppressing self-association (16,17). It appears that arginine and histidine interact with hydrophobic residues on the protein surface via cation-π interactions (18) as well as with charged residues via their guanidyl (9) and imidazole functionalities (19). In this manner, arginine and histidine are able to modify both electrostatic and hydrophobic interactions to reduce protein viscosity and self-association. Furthermore, hydrophobic interactions can be minimized by stabilizing proteins in the native folded state at high concentration by depletion attraction (20) via the addition of osmolyte depletants such as polymers and polysaccharides. The addition of preferentially-excluded osmolytes such as proline, glycine and trehalose would therefore favor the folded state of the protein to minimize the excluded volume, surface area and thus free energy at high concentration (20,21). Similarly, the attachment of protein or peptide tags, such as Cherry-Red™ can improve the thermal stability and solubility of proteins, which was demonstrated from molecular dynamics simulations to be due to modification of the protein surface net charge and hydrophobicity along with steric hindrance effects (22).
Proline has been shown to solubilize poorly-soluble proteins and suppress aggregation during protein refolding (23,24). Interestingly, proline is the most water-soluble of the common amino acids (7 M) at room temperature (25) and is often found on the solvent-exposed protein surface, despite possessing a cyclic side chain structure with three hydrophobic CH2 groups (26). Additionally, proline has been observed to cause a sharp increase in emission intensity and strong blue-shift in the emission spectra of ANS (8-anilinonaphthalene-1-sulfonic acid) dye at high concentrations, suggesting the creation of hydrophobic surfaces (24). Based on these observations, proline has been hypothesized to form amphipathic supramolecular assemblies (23,24). As a result of its amphipathic nature, proline can act as a hydrotrope in solubilizing hydrophobic proteins (23). Proline is excluded from the protein backbone (21,27) but also interacts favorably with the protein side chains (21,27), allowing it to improve protein stability by promoting refolding of chemically-denatured protein (27) and suppressing aggregation during refolding events (27). Although proline has been seen in some instances to reduce the viscosity of commercial antibodies (14,28), this effect has received little attention, and to our knowledge has not been studied systematically versus pH and co-solute concentration.
Herein, we compare the efficacy of the nonelectrolyte osmolytes proline (Pro), glycine (Gly) and trehalose (Tre) for reducing the viscosity and increasing the colloidal stability of concentrated mAb 1 solutions. Trehalose was chosen over the more widely used sucrose in order to facilitate comparison with our previous studies of depletant effects (using trehalose) on the viscosity of the mAb in this study (29,30). We demonstrate that proline improves the viscosity and colloidal stability of mAb 1 solutions (measured by size exclusion chromatography) as a function of pH and Pro concentration between 250 and 1300 mM. In contrast, these improvements are not observed for the common osmolytes glycine (up to 1300 mM) and trehalose (up to 580 mM). Proline may thus bind more strongly to the mAb than glycine and trehalose due to its more favorable interactions with the protein side chains (21) and weaken local anisotropic electrostatic attraction and hydrophobic interactions more effectively. Unlike the protonated bases arginine, histidine and imidazole, a potential advantage of the neutral co-solute proline is that it does not influence the ionic strength and does not require added counterions. Notably, we show that a binary system with Pro and His at a total co-solute concentration of 500 mM generates a greater viscosity reduction than 500 mM His alone, while greatly reducing the solution osmolarity and ionic strength due to reduced counterion levels.
Materials and Methods
Materials
The monoclonal antibody used in this study (mAb 1) is an IgG1 antibody with an isoelectric point (pI) of 9.3. mAb 1 was supplied by AbbVie at ~120 mg/mL in a proprietary buffer composition, and is the same mAb as in Hung et al. (31), Borwankar et al. (32) and Dear et al. (33). L-glycine, L-histidine, L-histidine hydrochloride monohydrate, hydrochloric acid, and imidazole were purchased from Fisher Scientific, Fairlawn, NJ. L-proline was purchased from Alpha Aesar, Ward Hill, MA. α-trehalose dihydrate (Tre) was purchased from Ferro Pfanstiehl Laboratories Inc., Waukegan, IL. Amicon Ultra-15 Ultracel – 30 K centrifugal filters were purchased from Merck Millipore Ltd. Ireland. Disposable 0.22 μm polyethersulfone (PES) bottle top filters and 13 mm syringe sterile filters were obtained from Celltreat Scientific Products, Shirley, MA (product codes 229,717 and 229,746).
Methods
Centrifugal Diafiltration and Ultrafiltration (CF)
The concentrated mAb solutions were prepared by centrifugation filtration as described previously (31,32,33). The co-solute buffers were prepared as described previously (33), where the reported co-solute concentration in the retentate, C C,R , depends on the concentration in the buffer feed, C C,F as a result of volume exclusion effects.
Here, C P is the targeted mAb concentration (230 mg/mL) and \( {\overline{\nu}}_p \) is the protein partial volume (0.7407 mL/g (34)). Consequently, in this study the final co-solute concentrations reported as 250, 400, 750 and 1300 mM correspond to feed concentrations of 300, 480, 905 and 1565 mM respectively. The buffers were sterile filtered with the Celltreat bottle top PES filters and then degassed under vacuum for 30 min.
Lyophilization Dilution (LD)
Lyophilized mAb 1 powder was prepared as described previously (33) with Pro or Tre added as a cryoprotectant and osmolyte in a 0.25:1 osmolyte:mAb mass ratio. The mAb powder was then reconstituted in concentrated aqueous buffers of His, Im or additional Pro to obtain ~100 μL of a 250 mg/mL mAb solutions at the desired co-solute concentration.
mAb Concentration Determination and Turbidity by UV-Vis Spectroscopy
The mAb concentration was measured in duplicate at 500× dilution using a Cary 60 UV-Vis spectrophotometer (Agilent Technologies, Santa Clara, CA), as described previously (31,32,33) with an extinction coefficient of 1.42 ml/mg/cm. The turbidity of the concentrated, unfiltered mAb solutions was measured at 350 nm in a micro volume size cell (A54094; Beckman Coulter, Indianapolis, IN) with a 0.2 cm path length. The turbidity was determined by subtracting the A350 of the protein-free solvent from the A350 of the sample and normalized to a 1 cm path length.
Viscosity Measurements
The solvent viscosity was measured in triplicate at 25°C using a size 50 Cannon-Fenske routine viscometer (Cannon Instrument Company, State College, PA) and averaged. The mAb viscosity was measured in triplicate using a customized capillary syringe viscometer as described previously (31,32,33).
The viscosity of protein solutions increases exponentially with protein concentration as described by colloidal viscosity models such the Ross-Minton equation (4,35). Due to small differences in sample mAb concentration between the different co-solute systems in this work, the viscosities were instead normalized by the protein concentration through the inherent viscosity
in order to more directly compare the effects of different co-solute formulations on the viscosity. Here η is the solution viscosity, η 0 is the solvent viscosity and c is the protein concentration (36). Unlike η, η inh increases linearly with concentration. Reported errors in η inh were calculated from propagation of error in concentration and viscosity measurements. Qualitatively, the inherent viscosity increases with the strength of the PPI. Given the modest variation in mAb concentrations in this study, the results were also normalized and presented as a calculated viscosity at a reference mAb concentration of 225 mg/mL, as determined using Eq. 2. Although η inh is expected to change by up to about 1.5 mL/g over the concentration range 205 to 230 mg/mL (31), for simplicity we assume a constant η inh for predicting the viscosity.
Dynamic Light Scattering
The collective diffusion coefficient D c of mAb 1 at high concentration in the different formulation conditions was measured by DLS using a Malvern Zetasizer Nano ZS equipped with a 633 nm laser (Malvern Instruments, Malvern, Worcestershire, United Kingdom). The D c was determined by fitting the autocorrelation function (ACF) with the quadratic cumulant algorithm. However, the 750 and 1300 mM proline measurements were fit with a stretched double exponential function (37) (Eq. 3) due to the presence of a visible second decay mode, with the first (fast) decay mode attributed to the diffusion of the protein.
Here, σ2 is an instrument-specific prefactor, A1 is the weighting factor by intensity for the fast decay mode, τ1 and τ2 are the characteristic relaxation times of the fast and slow decay modes respectively, and β is a stretch exponent to capture the shape of the slower exponential decay. The diffusion coefficients D i were determined from τ i via Eq. 4, where q is the scattering vector (Eq. 5) of the DLS measurement (36):
where λ is the incident laser wavelength, n is the solvent refractive index (RI), and θ is the scattering angle. The samples were sterile filtered through a 0.22 μm PES syringe filter (Celltreat Scientific Products, Shirley, MA) prior to DLS. The DLS measurements were made at 25°C in the back-scattering mode (scattering angle of 173°). Each sample was measured in triplicate for one minute per replicate, consisting of four 15-s scans which were averaged.
Accelerated Storage Stability Study
50 μL aliquots of the final mAb solution were stored in capped 300 μL HPLC vial inserts inside 1 ml HPLC vials (Thermo Fisher Scientific, Waltham, MA). The vials were sealed with three alternating layers of Parafilm and aluminum in order to minimize evaporative losses. The sealed vials were stored in a Boekel convection oven (model number 107905; Boekel Industries, Feasterville, PA) at 40°C for 4 weeks. The samples were diluted to 2 mg/mL for SEC analysis.
Size Exclusion Chromatography (SEC)
The relative level of irreversible soluble aggregates, or corresponding level of retained monomer in the final solutions before and after accelerated storage stability studies was quantified by SEC after dilution of the mAb solution to 2 mg/mL in the mobile phase (200 mM NaHPO4, 50 mM NaCl (pH 7)) and sterile filtration through a 0.22 μm PES syringe filter (Celltreat Scientific Products, Shirley, MA). As sterile filtration removes large insoluble aggregates, this method only quantifies the level of soluble aggregates, rather than total aggregates. The standard solution was prepared by diluting freshly-thawed mAb monomer stock (as provided at 120 mg/mL) in the mobile phase. A 10 μL injection of each sample was analyzed with a Waters Breeze HPLC (Waters Corporation, Milford, MA) equipped with a Tosoh Biosciences TSKgel3000SWXL column (Tosoh Corporation, Tokyo, Japan), operating at a flow rate of 0.5 ml/min. The eluate was monitored by the UV absorbance at 214 nm and 280 nm using a Waters 2489 UV/Visible detector (Waters Corporation, Milford, MA), which has a baseline noise level of <5 μAU. Typical peak heights in the spectrograph were 0.20 AU. The soluble monomer level (referred to as “% monomer” or “monomer retention” throughout the rest of this study) was quantified by the peak area ratio of the monomer peak to the monomer + aggregate peaks for the diluted samples. The monomer retention is equivalent to the mass fraction of soluble protein that remains monomeric, and not the total monomer recovery after incubation, as this method does not account for the monomer loss to formation of insoluble aggregates. Due to limited sample amounts, measurements were made without duplicates. However, the systematic instrument error was confirmed to be very small, as the standard deviation in the measured percent monomer (relative to total soluble protein content) for the mAb standard solution across 9 replicate measurements from multiple sample sets was less than 0.1% (for an average value of 99.7%).
Results
Increasing Viscosity Reduction with Increasing Proline Concentration
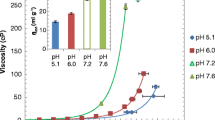
In order to investigate the effect of co-solutes on mAb 1’s viscosity, control experiments were first performed at 50 mM His-HCl (pH 5, 6). As seen in Table I, 205 and 208 mg/mL mAb 1 solutions had viscosities of 21 and 57 cP at pH 5 and 6, respectively, corresponding to inherent viscosities of 15.0 and 20.0 mL/g (Fig. 1a, Table I). In order to more directly compare different formulations, the mAb viscosity was calculated for a reference concentration of 225 mg/mL from Eq. 2 using the empirical inherent viscosity (measured at 200–230 mg/mL mAb 1). Although η inh increases modestly with mAb concentration (Eq. S2), by up to 1.7 mL/g between 190 and 230 mg/mL (31), we assumed that η inh for a given formulation remains constant between 200 and 225 mg/mL for comparison purposes. As seen in Fig. 1b, the viscosity was calculated to increase to >80 cP at 225 mg/mL and pH 6. For a phosphate (Phos) buffered system at the same pH, a higher viscosity of 80 cP was observed at 209 mg/mL mAb 1 (Table S1) with a calculated viscosity of 110 cP at 225 mg/mL (Fig. 1b).

Dependence of (a) measured inherent viscosity of concentrated 200–250 mg/mL mAb solutions and (b) calculated viscosity at 225 mg/mL mAb on proline concentration. Samples are buffered with 50 mM of histidine HCl or phosphate buffer. The corresponding viscosity data is shown in Table I and Table S1, and the colored lines are a guide to the eye.
In an attempt to reduce the mAb viscosity at pH 6 to acceptably low levels for subcutaneous injection, Pro was added at concentrations from 250 to 1300 mM. The highest concentration of 1300 mM was chosen to approach the threshold concentration of 1.5 M, above which proline has been shown to solubilize poorly-soluble proteins and suppress aggregation during protein refolding (23). While the highest co-solute concentration of 1300 mM is too high to be practical, the intermediate Pro concentration of 750 mM used here is still within the osmolarity threshold for acceptable levels of pain during injection (2× – 3× iso-osmolarity, according to various biopharmaceutical experts, and corresponding to 600–900 mM Pro). Given the small variation in mAb concentrations between samples, and considering the exponential dependence of viscosity on protein concentration, it is more meaningful to compare η inh rather than absolute viscosities, as explained in the Methods section on viscosity. As the Pro concentration was increased, similar large decreases in η inh of up to 6 mL/g for the mAb solution were observed in both the 50 mM His and Phos buffers (Fig. 1a; Table I; Table S1). The corresponding mAb viscosity in His and Phos buffer decreased by 3-fold to 20 cP at 195 mg/mL and 48 cP at 224 mg/mL, respectively (Fig. 1b; Table I; Table S1). In contrast, at pH 5 the addition of Pro in 50 mM His buffer caused a smaller reduction in η inh (Fig. 1a, Table I). As a consequence of the simultaneous increase in solvent viscosity and decrease in inherent viscosity with increasing Pro concentration, the calculated mAb viscosity at 225 mg/mL remained constant at ~25 cP over the entire concentration range (Fig. 1b). It is interesting to note that the addition of high concentrations of Pro at pH 6 reduces the mAb viscosity and inherent viscosity to approach the same value as at pH 5.
Lack of Viscosity Reduction from Preferential Exclusion with Glycine and Trehalose
To further probe the effects of neutral, preferentially excluded (20,21) osmolytes on protein viscosity, glycine and trehalose were studied. Glycine in some cases can greatly reduce the viscosity of mAb solutions (14). However, we found that at pH 5 Gly mildly increased η inh for mAb 1, by up to ~3 mL/g at the highest concentration of 1300 mM relative to the 50 mM His control (Fig. 2a, Table II). At pH 6, 250 mM Gly appears to cause a modest reduction in η inh relative to the control, even at a higher mAb concentration (Table II, S2). However, further increasing the Gly concentration to 1300 mM increased η inh relative to the control, even at a lower mAb concentration (Table II, S2). Consequently, whereas the addition of Pro caused a large reduction in the calculated mAb viscosity at pH 6, Gly caused a significant increase in the calculated viscosity at both pH 5 and 6, as seen in Fig. 2b.

Dependence of (a) measured inherent viscosity of concentrated 200–250 mg/mL mAb solutions and (b) corresponding calculated viscosity at 225 mg/mL mAb on co-solute choice (proline, glycine) and co-solute concentration. The proline data from Fig. 1 is reproduced here for visual comparison against the glycine viscosity data. Samples are buffered with 50 mM histidine HCl. The corresponding viscosity data is shown in Tables I and II, and the colored lines are a guide to the eye.
Similar to the case for Gly, the addition of Tre up to 580 mM did not reduce the mAb viscosity at pH 6 (Table S2, S3). For mAb solutions buffered in 30 mM His, the addition of up to 220 mM Tre produced a mild increase in η inh relative to the Tre-free control (Table S2; control in row 1). However, the modest increase in η inh of up to ~1.0 mL/g may be accounted for in part by the higher corresponding mAb concentrations. Similarly, the addition of up to 580 mM Tre did not significantly change η inh for mAb solutions buffered with 50 mM Phos (Table S3) relative to the Tre-free phosphate control (row 1 in Table S3). Unlike for Gly, which modestly reduces the viscosity at pH 6 and low Gly concentration, Tre does not appear to reduce the viscosity at any concentration between 150 and 580 mM.
The results for Gly and Tre indicate that depletion attraction produced by a neutral co-solute does not produce a general viscosity reduction, as was proposed in an earlier study based upon a theoretical model (30). The greater viscosity reduction for Pro versus Gly or Tre may be influenced by the stronger interactions between the Pro and protein side chains. Furthermore, the larger size of Pro compared to Gly may also contribute to greater depletion attraction at the same co-solute concentration, which in turn promotes a more compact, folded conformation for the mAb (20) and minimizes the number of solvent-exposed self-association sites.
Viscosity Reduction with Binary Co-Solutes: Proline with Histidine or Imidazole
To attempt to modulate the electrostatic and hydrophobic PPI to further reduce the viscosity, we also examined binary systems of Pro with protonated His or Im. Histidine is a pharmaceutically-acceptable co-solute that has been shown to reduce the viscosity of several mAbs (31,33,38,39), likely through preferential interactions of His with aromatic residues via its imidazole side chain as has been seen in simulations (18,19), which may screen hydrophobic interactions. Imidazole was also investigated for its same favorable preferential interactions with aromatic residues, but was also chosen due to its smaller size compared to His (it lacks the amino acid backbone of His). Due to the similar mechanisms by which the two co-solutes modify protein interactions, at 250 mM both reduced the mAb inherent viscosity at ~230 mg/mL and pH 6 by 3–4 mL/g relative to the 50 mM His control (Table S4), indicating a weakening of attractive PPI. The reduction in inherent viscosity is slightly greater for His compared to Imid, but the higher solvent viscosity for His caused the net change in viscosity to be identical between His and Imid (Table S4). The addition of 250 mM Pro further reduced η inh at ~230 mg/mL from 16 to 17 mL/g without Pro (Table S4) to 14.8 and 14.3 mL/g at ~225 mg/mL for the His and Im systems, respectively (Fig. 3a, Table S5), with the corresponding calculated viscosity at 225 mg/mL decreasing by nearly 1.5-fold from 40 to 45 cP without Pro to 25–30 cP (Fig. 3b). However, the addition of Pro in the binary systems at pH 5 did not further reduce η inh (Fig. 3a, Table S5) or viscosity (Fig. 3b) significantly. A summary of the lowest-viscosity proline formulations are presented in Fig. 4.

(a) Measured inherent viscosity and (b) calculated viscosity of 225 mg/mL mAb solutions containing 250 mM histidine or imidazole with no proline (blue bars) or an additional 250 mM proline (red bars).

Viscosity reductions for select 200–230 mg/mL solutions at pH 6 after the addition of high concentrations of proline to the mAb solution in histidine or imidazole buffer. The exact mAb and buffer concentrations are listed in mg/mL and mM, respectively.
Impressively, the use of binary Pro-His systems will now be shown to lower viscosities more than His-only formulations at the same ‘active’ co-solute concentration (i.e. not counting the counterions) but at a lower solution osmolarity. For example, the calculated viscosity of this mAb at 225 mg/mL and pH 6 was 40 cP in 250 mM His (Fig. 3b; also Fig. S11 from Dear et al. (33)). Further increases in His concentration to 500 mM led to a small reduction in the calculated viscosity to 35 cP (Table S7), but at a total co-solute osmolarity of 810 mM. In contrast, the 250:250 mM His:Pro binary system reduced the expected (calculated) viscosity at 225 mg/mL to 31 cP, at a lower total co-solute osmolarity of 655 mM. An intermediate His concentration of 360 mM (osmolarity = 583 mOsm/L), closer to the osmolarity of the binary Pro-His system, actually caused a small increase in the calculated viscosity (Table S7) to 44 cP, but the change is small and may be caused in part by the sample drift to a slightly higher final solution pH.
While binary systems of Pro with His offer larger viscosity reductions than His-only at the same total co-solute concentration, both co-solutes are much less effective than Arg for the reducing the viscosity of mAb 1 at pH 6. As seen from Fig. S11 of an earlier study with this mAb (33), the addition of 250 mM Arg.HCl at pH 6 reduces the viscosity at 225 mg/mL to 26 cP. Increasing the Pro conc at pH 6 does reduce the viscosity to this same low value, but requires over 1300 mM to do so (Fig. 1b). Similarly, the same study showed that over 800 mM His would be needed at pH 6 to reduce the viscosity to ~26 cP. It is clear that at least for this mAb at pH 6, Arg is far more effective on a molar basis for reducing the mAb viscosity.
For practical formulation development applications, Pro as the main viscosity modifier may thus not be as desirable or advantageous as Arg due to its lower molar efficacy for reducing viscosity. However, there may be certain situations where Pro is advantageous over Arg due to its neutrality, as will be explained in the Discussion section below. Furthermore, the fundamental insight gained from contrasting the effects of Pro on the mAb viscosity and stability relative to Arg and His may provide guidelines for the future selection and design of highly potent viscosity-reducing co-solutes for a wide variety of mAbs.
Effects of Osmolytes on mAb Stability Against Aggregation
To characterize the mAb colloidal stability under thermal stress, the level of irreversible soluble aggregates after 4 weeks of storage at 40°C was quantified by SEC. The pre-storage stability of the mAb was excellent across all tested formulations, with a monomer content of over 99% (relative to the total soluble protein content), as seen from Fig. 5a. However, mAb 1 showed a significant loss in soluble monomer content after 4 weeks of storage at 40°C at both pH 5 and 6 in 30 mM His(HCl) control buffers, as seen in Dear et al. (33) and Fig. 5b, c. The monomer contents of the mAb 1 pH 5 and 6 controls were 92.0 and 88.2% respectively. The addition of 220 mM Tre suppressed aggregation of mAb 1, yielding 99.5% monomer (33), consistent with protein stabilization by a general osmotic depletion effect to favor folding (20,21). Similarly, both Pro and Gly suppressed soluble mAb aggregation at all tested co-solute concentrations. As seen in Fig. 5b, c, the relative monomer content of the mAb 1 solutions in 50 mM His with 250–1300 mM Pro or Gly were greater than 96.5% after storage, compared to the 30 mM His controls with less than 92.0% monomer content. Increasing Pro concentration between 250 and 1300 mM progressively inhibited post-storage aggregation at both pH 5 (Fig. 5b) and 6 (Fig. 5c; Table S8). However, the overall effect of Pro on both viscosity and stability at pH 5 is weak. In contrast, the pH 6 Pro systems are all at comparable mAb concentrations of 195–205 mg/mL, and thus show a clear correlation between higher Pro levels and increased storage stability.

Accelerated storage stability of 193–232 mg/mL mAb solutions with addition of proline, glycine or histidine. (a) Monomer fraction (of total soluble protein content) before storage as a function of co-solute and pH. The intermediate co-solute concentration is 750 mM for both Pro systems (pH 5 and 6), and 400 mM for Gly (pH 5). No initial stability measurements were made for 250 or 400 mM Gly at pH 6. (b) Monomer fraction after 4-weeks storage at 40°C as a function of co-solute formulation at pH 5. (c) Monomer fraction after 4-weeks storage at 40°C as a function of co-solute formulation at pH 6. The 30 mM His(HCl) 4-week storage controls shown by black bars were reproduced from an earlier study on the same mAb, as reported in Dear et al.,(33) where the pH 5 control was at 242 mg/mL, and the pH 6 control was at 224 mg/mL. (d) Monomer fraction before (blue bars) and after (red bars) 4 weeks of storage with histidine at pH 5 and 6 with an additional 250 mM proline. All sample concentrations are shown in Tables S8 – S10. The approximate sample concentrations for each pH/co-solute concentration pair in (b) and (c) are also shown in text boxes on each figure. The monomer fraction was measured by SEC after dilution of the storage samples to 2 mg/mL in the SEC mobile phase.
In contrast, the stabilizing effects of Gly at pH 5 were diminished at 1300 mM relative to 250 mM (Fig. 5b, Table S9). Similarly, 1300 mM Gly at pH 6 did not increase the post-storage stability as much as 1300 mM Pro (Fig. 5c, Table S8, S9). Thus, while both Pro and Gly imparted overall stabilizing effects on mAb 1 relative to 30 mM His, Gly addition beyond 250 mM proved to be less beneficial. These differing trends can be explained in the context of the net effect of Gly and Pro on the protein stability resulting from a competition between stabilization of the peptide backbone through crowding effects and destabilization of the hydrophobic residue side chains through preferential co-solute interactions, which lead to favorable solvation energies for the side chains (20,21,40). CH-π interactions between Gly/Pro and aromatic residues create favorable solvation energies for hydrophobic side chains (40,41) and destabilize them (greater conformational instability). However, Pro may contribute more steric repulsion or crowding for greater colloidal stability due to its larger size and rigid side chain structure, which helps to compensate for the increased conformational instability at higher co-solute concentrations. In contrast, Gly’s small size results in less steric repulsion to counteract the greater conformational instability, such that the net effect at 1300 mM Gly is reduced overall protein stability relative to 250 mM Gly (but still more stable than the 30 mM His control).
Interestingly, the enhancement of mAb 1’s stability with increasing Pro concentration was more pronounced at pH 6, with a 12-fold reduction in monomer loss from 2.4% down to 0.2% at the highest Pro concentration of 1300 mM (Fig. 5c, Table S8). Proline at the same concentration also nearly eliminated aggregation at pH 5, with a monomer loss of 0.01% (Table S8). However, aggregation was less of an issue overall at pH 5, given the much smaller monomer loss at pH 5 relative to pH 6 at each Pro concentration studied (Table S8).
Similar to Pro, His improved mAb 1’s stability with increasing co-solute concentration, as seen in Fig. 5d. An increase in His concentration from 50 to 250 mM in the presence of 250 mM Pro caused a nearly five-fold reduction in monomer loss at both pH 5 and 6, down to as low as 0.2% and 0.4% respectively (Table S9). As was also the case for the Pro systems (Fig. 5b, c), the increased His concentration had a much larger effect on mAb stability at pH 6 than at pH 5, maintaining 99% monomer retention (relative to the total soluble protein content) after 4 weeks of accelerated storage, as opposed to 96.5% at 50 mM His (Fig. 5d). Furthermore at pH 6, 250 mM His stabilized the protein more effectively than Pro at the same concentration.
Contrasting Effects of Proline and Glycine on mAb Dynamics at High Concentration
The collective diffusion coefficient D c of the mAb at high concentration, as measured by DLS, is strongly influenced by protein-protein interactions via the osmotic compressibility dπ/dc (Eq. 6), where ϕ is the mAb volume fraction and f sp c s is a protein-solvent friction term (42).
As such, osmolyte effects on D c can be related to change in the strength of the protein-protein interactions, where dπ/dc is inversely proportional to the PPI strength (43). The effects of proline and glycine on D c (Table III) and viscosities (Tables I and II) were correlated. The parameters used to fit D c are given in Table S11. Interestingly, a prominent second decay mode was observed for 750 and 1300 mM proline (Fig. S4a) even though the samples had already been filtered through 0.22 um filters prior to DLS to remove large aggregates. The origin of the second decay mode is unclear, although measurements of the mAb-free Pro solvents suggest that the decay mode may correspond with entities possibly formed at ultrahigh Pro concentrations, as discussed in the Supporting Info. The ACFs were therefore fit to a stretched double exponential function (Table S11). To account for differences in mAb 1’s diffusivity due to the formulation solvent viscosities, D c was normalized as D c /D 0 , where D 0 is measured at infinite dilution in the given solvent. The value of D 0 was determined from a previous measurement for the same mAb in a 30 mM His buffer (33) and corrected for η 0 of each formulation using the Stokes-Einstein relation (Eq. 7).
The D c /D 0 of mAb 1 increased with proline concentration to above 1.0, whereas it decreased very slightly from 0.75 to 0.71 with increasing glycine concentration (Table III). The PDI of D c also increased with glycine concentration, although the values for proline and glycine at 250 mM were comparable. The second decay mode of 750 and 1300 mM proline corresponded to a very slow-moving entity/relaxation mode, with D eff /D 0 of ~0.006 (Table S11). However, the relative contribution of this slower decay mode to the overall ACF was small (~30% by intensity; Table S11). This secondary decay was not observed for glycine (Fig. S4b) even at the highest glycine concentration of 1300 mM. At the same mAb concentration, the increasingly faster D c /D 0 with proline can be attributed to more repulsive (or less attractive) protein-protein interactions via the dπ/dc contribution to D c (42). The increase in D c with proline concentration therefore suggests a progressive weakening of the attractive protein-protein net interactions, which is in line with the observed reduction in mAb viscosity. In contrast, the negligible change in D c /D 0 with increasing glycine concentration suggests that glycine has no discernible effect on the protein interactions. The possible causes for the greatly differing effects of proline and glycine on the mAb interactions, viscosity, and stability will be explored further in the Discussion section below.
Discussion
Proline as an Alternative Viscosity Modifier to Ionic Co-Solutes Such as Arg and His
The high concentrations of Pro needed to achieve an acceptable level of viscosity reduction for this mAb at pH 6 are impractical, but it will be shown below to be practical in mixtures with histidine and imidazole. Since cryoprotectants are already included in most lyophilized formulations, proline can be used in place of other cryoprotectants (such as sucrose, mannitose, etc.), with the added benefit of improving the viscosity and stability of the reconstituted protein at high concentration. Using a cryoprotectant that also functions as a viscosity modifier will help to reduce the total osmolality of the mAb drug product, as less Arg, His etc. is needed to maintain low viscosities. Pro has also been approved for use in parenteral formulations (28), and thus it would be helpful to compare its performance as a viscosity modifier against other co-solutes such as Arg and His. Pro shares some physical characteristics with Arg – it is an amphipathic molecule whose side chain (pyrrolidine) has been shown in simulations to preferentially interact with hydrophobic residues via CH-π interactions (41).
An important advancement of this work is the demonstration of greater reductions of viscosity for a Pro-His(HCl) mixture relative to pure His(HCl). As discussed earlier, the 250:250 mM His(HCl):Pro binary system was able to reduce the calculated mAb viscosity at 225 mg/mL and pH 5 even relative to 500 mM His(HCl) (same total co-solute concentration, not including the Cl− counterions), while also providing for a lower co-solute osmolarity of 655 mOsm/L, compared to 810 mOsm/L for 500 mM His(HCl) (Table S7). An osmolarity of 655 mOsm/L may be low enough for injection based on various biopharmaceutical experts. Since Pro is a neutral molecule, it is compatible with other ionic co-solutes, creating a large formulation space for binary Pro systems. However, binary Pro-Arg systems were not explored in this work since Pro-Arg systems require a third solvent component (a buffering agent) due to Arg’s poor buffering capability at clinically relevant pH values (pH 5–6), leading to a more complicated formulation.
Furthermore, there are some situations where Pro may be advantageous over ionic co-solutes such as Arg and His. For example, in formulation conditions where the mAb has a strong net charge (ex. low pH far from the pI), increasing the ionic strength may screen out the electrostatic repulsion and actually increase the net attraction (44,45,46), which has in one case led to phase separation (44). It may therefore be preferable to use a neutral viscosity modifier such as Pro instead of ionic co-solutes such as Arg and His in order to weaken the non-electrostatic PPI without screening out the beneficial electrostatic repulsion.
Inferring Protein-Protein Interaction from Viscosity and Stability
Although protein-protein interactions are commonly measured at low concentration, they have rarely been measured at concentrations above 200 mg/mL, even without added co-solutes. More recently, neutron scattering (47) and light scattering (3,4,48) at high concentration have given some insight into how charged co-solutes modify mAb self-association to lower the mAb viscosity, as well as demonstrated differences in co-solute binding to the protein surface (49). Despite the limited information, it may also be possible to begin to infer how co-solutes influence PPI from the degree by which they modify the viscosity, presumably by weakening the PPI. For example, Arg+ is known to bind to hydrophobic sites (50). Arg+ is also known to reduce the viscosity of highly concentrated mAbs significantly more than inorganic salts such as NaCl (15), which do not modify hydrophobic interactions.
The pH-dependent effect of His and Im on viscosity and stability (Table S4, Fig. 5d) can be explained in terms of the protein net charge and charge distribution, which change with pH. Since the mAb pI is 9.3, the mAb has a smaller net positive charge and a greater degree of charge anisotropy at pH 6 than at pH 5, where the charge distribution balance is shifted towards positive charges. As a result, the increase in pH from 5 to 6 both weakens the long-ranged electrostatic repulsion between mAb molecules and strengthens the local anisotropic electrostatic attraction (due to a higher number and probability of attractive electrostatic contacts between oppositely-charged sites on neighboring mAb molecules), leading to stronger protein self-association and higher viscosities. Protonated His and Im may be able to neutralize negative residues via ion pairing, and may also make neutral and positive residues more strongly positive via interactions between the imidazole functionality and the imidazole/guanidyl side chains of the His and Arg residues, respectively (19). As a result, His may be able to increase the protein net charge and reduce surface charge anisotropy, thus strengthening global electrostatic repulsion while weakening local anisotropic electrostatic attraction. Changes in the mAb net charge at high concentration caused by co-solute binding may be quantified in future studies from zeta potential measurements, although extremely low protein electrophoretic mobilities at high ionic strength and/or concentration may prove challenging for obtaining meaningful measurements. Since the mAb has a smaller net charge and greater charge anisotropy at pH 6, modification of the mAb charge and charge distribution by His is expected to have a larger effect on the mAb viscosity and stability than at pH 5, where there are fewer anisotropic attractive interactions to neutralize. The significant differences in the reduction of aggregation and viscosity (η inh ) by His at pH 5 and 6 (Table S4, Fig. 5d) are in agreement with this hypothesis, and are indicative of histidine’s effect on the electrostatic PPI. The modestly larger reduction in inherent viscosity by His compared to Imid may potentially be attributed to its larger molecular size, which allows it to contribute more steric repulsion and molecular crowding than Imid at the same molar concentration, favoring a more compact folded mAb conformation (20) that leads to fewer surface-exposed attraction sites. Correspondingly, the larger reduction in η inh for Imid with added Pro, compared to for His, may likely be attributed to the smaller size of Imid which results in less steric hindrance for Pro molecules to approach and interact with the attractive sites on the mAb surface. In addition, the relative increase in added steric repulsion from Pro is greater for Imid compared to His, given the smaller occupied volume by Imid. However, the differences in viscosity between His and Imid with Pro are not significant enough to support any conclusive statements regarding differences in the two co-solutes’ effects on the PPI.
We now address the pH-dependent effects of Pro on the mAb viscosity and stability. Polyol and sugar osmolytes are known to have a pH-dependent effect on protein conformational stability at low pH (~2–5) (51) due to changes in the protonation state of the carboxylic acid side chains (pKa ~ 4) in the aspartic acid and glutamic acid residues. The resulting change in protein hydrophobicity (52) modifies the degree of osmolyte exclusion due to repulsion between the polar osmolyte OH− groups and hydrophobic protein surface. However, given the lack of OH− groups in Pro and the higher pH (5,6) in this study, the preferential exclusion mechanism cannot explain proline’s pH-dependent effects on the mAb viscosity and stability. Instead, it is likely that Pro reduces mAb viscosity and aggregation by modifying the local attractive hydrophobic and/or electrostatic interactions directly. Although the pH dependence may reflect changes in protein hydrophobicity with pH, the similarity between proline and histidine’s beneficial effects on these properties with increasing pH may also indicate modification of the electrostatic PPI by Pro, despite proline’s net neutrality between pH 5 and 6.
It is possible that the partial positive charge on the pyrrolidine ring side chain (41) as well as the polar groups in Pro may interact with charged mAb residues. The greater molar efficacy of His for reducing viscosity and aggregation relative to Pro may be due to the larger positive charge on its side chain. For example, nearly three times as much Pro as His (750 vs 250 mM respectively; Fig. 1b; Table S4) is needed to reduce the viscosity at 225 mg/mL and pH 6 to 40 cP. Similarly, the addition of 1050 mM Pro to a reference condition (50:250 mM His:Pro; pH 6) was needed to achieve the same level of aggregation suppression as adding 200 mM His (Fig. 5c, d). However, although Pro reduces viscosity and aggregation at a lower molar efficiency than His, the observation that it does so in a similar pH-dependent basis (suggesting modification of electrostatic PPI) is surprising given its zero net charge.
Proline Amphipathic Behavior and Modification of Hydrophobic PPI
Proline has been observed to bind to aromatic residues by X-ray crystallography (53). Simulations show that Pro interacts with (and binds to) those residues through its pyrrolidine ring side chain via CH-π interactions (41). As a result, Pro can potentially shield aromatic residues and disrupt hydrophobic PPI via preferential hydration of the protein surface through its zwitterionic functionality (54). The stabilization and twofold viscosity reduction of a 200 mg/mL polyclonal IgG solution by 250 mM Pro observed in literature (28) has been attributed to this effect. However, Pro lowered the viscosity of two other mAbs at low concentration but increased the viscosity for one mAb at >200 mM Pro (55), in contrast to the monotonic behavior (up to 1300 mM Pro) for the mAb in this study. This variability likely reflects the different distributions of hydrophobic and charged interactions sites between the different mAbs. Once the interacting sites are saturated or fully screened by the Pro molecules, the addition of further Pro only serves to increase the solvent viscosity and therefore the solution viscosity.
At high proline concentration, proline’s ability to disrupt hydrophobic interactions may also be enhanced by the hypothesized formation of dimers with hydrophobic “pockets” (26) or of supramolecular aggregates (23,24) via alignment of the pyrrolidine rings. In both cases, the resulting Pro aggregate is strongly amphipathic, with a hydrophilic face (the carboxyl and amino groups) and a hydrophobic face (the stacked pyrrolidine rings). This amphipathic structure may explain why Pro at 2 M or higher has been shown to be a potent hydrotrope (23), as Pro may act as a surfactant to bridge the exposed hydrophobic mAb residues and the solvent molecules, subsequently weakening the attractive hydrophobic PPI. The unexpected observed increase in the mAb solution turbidity with decreasing viscosity at high Pro concentration (Supporting Info) may be attributed in part to the potential formation of Pro aggregates. Similarly, the development of a much slower secondary relaxation mode in the DLS ACFs for 750 and 1300 mM proline, which was not present for glycine at the same co-solute concentration nor at 250 mM proline, may also indicate structuring of the proline at high concentration and interactions of the mAb with the proline superstructure. It is unlikely that this second decay mode is caused by large mAb aggregates, as the samples were filtered in order to remove aggregates prior to DLS. There was also less irreversible aggregation observed by SEC (Fig. 5c) than at 250 mM proline, where the second decay was not present. Similarly, although a secondary decay mode is often indicative of glassy fluids (56) or percolating networks (57), it is unlikely that the high proline mAb solution is approaching a glassy state, given the low viscosities. Instead, the appearance of the secondary slow decay mode corresponds to a large reduction in mAb viscosity and increase in 4-week storage stability, suggesting that this decay mode corresponds to a beneficial entity/mechanism, such as the proposed formation of proline supramolecular aggregates at ultrahigh co-solute concentration. However, further investigation with spectroscopic and other orthogonal techniques would be warranted to validate this hypothesis in future studies.
Favorable Side-Chain Interactions: Relation to Viscosity and Stability
The larger viscosity reductions for Pro relative to the other two osmolytes may be explained in part by the strength of the interactions between the osmolytes and protein residue side chains. Although all three osmolytes are preferentially excluded from the protein backbone, they interact differently with the side chains (21). Trehalose is considered to have the weakest side chain interactions of the three, given the small overall free energy change for side chain transfer into a 1 M osmolyte solution. In contrast, Pro exhibits strongly favorable side chain interactions given the large negative transfer energies (21). More specifically, the transfer energies for apolar (hydrophobic) side chains into Tre solutions are positive (unfavorable), whereas they are significantly negative (favorable) in Pro solutions (21). The favorable side chain interactions of Pro are due to the pyrrolidine ring, which preferentially interacts with aromatic residues and may allow Pro to shield the hydrophobic residues (26,41). Glycine can also interact with aromatic residues via CH-π interactions (41), but to a much weaker extent than Pro given the fewer CH-groups available. The stronger side chain interactions may contribute to proline’s greater efficacy for weakening local attractive PPI and reducing the mAb viscosity compared to Tre and Gly.
The differences in Pro and Gly’s interactions with the protein side chain may also explain their different effects on the colloidal stability of mAb 1. An increase in osmolyte concentration is expected to improve mAb conformational stability due to osmotic depletion (20), which in turn maintains the mAb in the native folded state and minimizes self-association between hydrophobic sites. However, at high total solute (protein + osmolyte) concentrations, the increased proximity of the protein molecules to each other in the crowded system may enhance local electrostatic and hydrophobic attraction, leading to increased self-association despite the greater conformational stability. Proline appears to weaken these attractive interactions, evidenced by its efficacy for reducing the viscosity. In contrast, Gly does not appear to do so, as indicated by its unfavorable effect on the viscosity. This difference may explain why Pro progressively stabilizes mAb 1 with increasing osmolyte concentration (Fig. 5b, c; Table S8), while Gly has the opposite effect (Fig. 5b, c; Table S9).
Conclusions
Proline reduces the viscosity of a concentrated 225 mg/mL mAb 1 solution by up to 3-fold down to 25 cP at pH 6, but has no effect at pH 5. More interestingly, a 250:250 mM His(HCl):Pro binary system causes a greater viscosity reduction at pH 6 (from 84 cP to 31 cP at 225 mg/mL (calculated)) compared to a His system at the same total co-solute concentration (500 mM; 35 cP at 225 mg/mL). Notably, the osmolarity of the His-Pro binary system is 655 mOsm/L, which may be considered within the acceptable range, below that of the His-only system (810 mOsm/L), as Pro does not contribute any counterions to the solution. The lack of charge or counterions for Pro may also make it more advantageous than ionic viscosity-modifiers such as Arg and His in certain systems with strong electrostatic repulsion (ex. pH far from pI; strongly repulsive B 22 ’s etc.), where increasing the ionic strength through addition of ionic co-solutes may lead to stronger attraction and eventual phase instability (44). Pro similarly progressively increases the protein 4-week 40°C storage stability in terms of the soluble monomer content from 92.0% and 88.2% without added co-solute at pH 5 and 6 respectively, to ≥99% at 1300 mM Pro for both pH values. The reductions in viscosity and aggregation are not simply due to osmotic depletion, as two other neutral osmolytes, Gly and Tre, raise both properties. The viscosity of the mAb solution decreased from pH 6 to pH 5 without added co-solute, which may be attributed to the mAb’s larger positive net charge, reduced surface charge anisotropy, and weaker local anisotropic electrostatic attraction. It is likely that at pH 6, proline also weaken these interactions by modifying charged sites on the protein surface, given the large dipole moment of its zwitterionic functionality. Furthermore, the hydrophobic part of the pyrrolidine ring may bind to hydrophobic sites on the protein surface and weaken hydrophobic attraction. The faster normalized collective diffusion of mAb 1 at high proline concentration also suggests a weakening of attractive protein interactions. Although co-solute effects on protein morphology have rarely been studied at high protein concentrations, they would be warranted in the future to better understand how changes in morphology and PPI influence viscosity.
Abbreviations
- η :
-
Viscosity
- η inh :
-
Inherent viscosity
- CF:
-
Centrifugal filtration
- D c :
-
Collective diffusion coefficient
- D 0 :
-
Diffusion coefficient at infinite dilution
- Gly:
-
Glycine
- His:
-
Histidine
- Imid:
-
Imidazole
- LD:
-
Lyophilization dilution
- mAb:
-
Monoclonal antibody
- Phos:
-
Phosphate
- pI:
-
Isoelectric point
- PPI:
-
Protein-protein interactions
- Pro:
-
Proline
- SEC:
-
Size exclusion chromatography
- Tre:
-
Trehalose
References
Shire SJ, Shahrokh Z, Liu J. Challenges in the development of high protein concentration formulations. J Pharm Sci. 2004;93(6):1390–402.
Connolly Brian D, Petry C, Yadav S, Demeule B, Ciaccio N, Moore Jamie MR, et al. Weak interactions govern the viscosity of concentrated antibody solutions: high-throughput analysis using the diffusion interaction parameter. Biophys J. 2012;103(1):69–78.
Scherer TM, Liu J, Shire SJ, Minton AI. Intermolecular interactions of IgG1 monoclonal antibodies at high concentrations characterized by light scattering. J Phys Chem B. 2010;114(40):12948–57.
Lilyestrom WG, Yadav S, Shire SJ, Scherer TM. Monoclonal antibody self-association, cluster formation, and rheology at high concentrations. J Phys Chem B. 2013;117(21):6373–84.
Arora J, Hu Y, Esfandiary R, Sathish HA, Bishop SM, Joshi SB, et al. Charge-mediated Fab-Fc interactions in an IgG1 antibody induce reversible self-association, cluster formation, and elevated viscosity. mAbs. 2016;8(8):1561–74.
Quang LJ, Sandler SI, Lenhoff AM. Anisotropic contributions to protein–protein interactions. J Chem Theory Comput. 2014;10(2):835–45.
Chari R, Jerath K, Badkar AV, Kalonia DS. Long- and short-range electrostatic interactions affect the rheology of highly concentrated antibody solutions. Pharm Res. 2009;26(12):2607–18.
Yadav S, Laue TM, Kalonia DS, Singh SN, Shire SJ. The influence of charge distribution on self-association and viscosity behavior of monoclonal antibody solutions. Mol Pharm. 2012;9(4):791–802.
Buck PM, Chaudhri A, Kumar S, Singh SK. Highly viscous antibody solutions are a consequence of network formation caused by domain-domain electrostatic complementarities: insights from coarse-grained simulations. Mol Pharm. 2015;12(1):127–39.
Yadav S, Liu J, Shire SJ, Kalonia DS. Specific interactions in high concentration antibody solutions resulting in high viscosity. J Pharm Sci. 2010;99(3):1152–68.
Du W, Klibanov AM. Hydrophobic salts markedly diminish viscosity of concentrated protein solutions. Biotechnol Bioeng. 2011;108(3):632–6.
Kanai S, Liu J, Patapoff TW, Shire SJ. Reversible self-association of a concentrated monoclonal antibody solution mediated by Fab-Fab interaction that impacts solution viscosity. J Pharm Sci. 2008;97(10):4219–27.
Salvi G, De Los Rios P, Vendruscolo M. Effective interactions between chaotropic agents and proteins. Proteins: Struct, Funct, Bioinf. 2005;61(3):492–9.
Chang BS. Inventor; Integritybio Inc., assignee. Protein formulations containing amino acids patent WO2013063510 A1. 2013.
Inoue N, Takai E, Arakawa T, Shiraki K. Specific decrease in solution viscosity of antibodies by arginine for therapeutic formulations. Mol Pharm. 2014;11(6):1889–96.
Arakawa T, Ejima D, Tsumoto K, Obeyama N, Tanaka Y, Kita Y, et al. Suppression of protein interactions by arginine: a proposed mechanism of the arginine effects. Biophys Chem. 2007;127(1–2):1–8.
Chen B, Bautista R, Yu K, Zapata GA, Mulkerrin MG, Chamow SM. Influence of histidine on the stability and physical properties of a fully human antibody in aqueous and solid forms. Pharm Res. 2003;20(12):1952–60.
Liao SM, Du QS, Meng JZ, Pang ZW, Huang RB. The multiple roles of histidine in protein interactions. Chem Cent J. 2013;7
Heyda J, Mason PE, Jungwirth P. Attractive interactions between side chains of histidine-histidine and histidine-arginine-based cationic dipeptides in water. J Phys Chem B. 2010;114(26):8744–9.
Arakawa T, Timasheff SN. The stabilization of proteins by osmolytes. Biophys J. 1985;47(3):411–4.
Auton M, Rösgen J, Sinev M, Holthauzen LMF, Bolen DW. Osmolyte effects on protein stability and solubility: a balancing act between backbone and side-chains. Biophys Chem. 2011;159(1):90–9.
Baumann P, Schermeyer M-T, Burghardt H, Dürr C, Gärtner J, Hubbuch J. Prediction and characterization of the stability enhancing effect of the Cherry-Tag™ in highly concentrated protein solutions by complex rheological measurements and MD simulations. Int J Pharm. 2017;531(1):360–71.
Samuel D, Kumar TKS, Jayaraman G, Yang PW, Yu C. Proline is a protein solubilizing solute. Biochem Mol Biol Int. 1997;41(2):235–42.
Kumat TKS, Samuel D, Jayaraman G, Srimathi T, Yu C. The role of proline in the prevention of aggregation during protein folding in vitro. IUBMB Life. 1998;46(3):509–17.
Amend JP, Helgeson HC. Solubilities of the common L-α-amino acids as a function of temperature and solution pH. Pure Appl Chem. 1997;69(5):935–42.
Busch S, Lorenz CD, Taylor J, Pardo LC, McLain SE. Short-range interactions of concentrated proline in aqueous solution. J Phys Chem B. 2014;118(49):14267–77.
Ignatova Z, Gierasch LM. Inhibition of protein aggregation in vitro and in vivo by a natural osmoprotectant. Proc Natl Acad Sci U S A. 2006;103(36):13357–61.
Maeder W, Lieby P, Sebald A, Spycher M, Bolli R. Stability over 24 months and tolerability of a new 20% proline-stabilized polyclonal immunoglobulin for subcutaneous administration (SCIG). J Allergy Clin Immunol. 2010;125(2, Supplement 1):AB142.
Borwankar AU, Dinin AK, Laber JR, Twu A, Wilson BK, Maynard JA, et al. Tunable equilibrium nanocluster dispersions at high protein concentrations. Soft Matter. 2013;9(6):1766.
Johnston KP, Maynard JA, Truskett TM, Borwankar AU, Miller MA, Wilson BK, et al. Concentrated dispersions of equilibrium protein nanoclusters that reversibly dissociate into active monomers. ACS Nano. 2012;6(2):1357–69.
Hung JJ, Borwankar AU, Dear BJ, Truskett TM, Johnston KP. High concentration tangential flow ultrafiltration of stable monoclonal antibody solutions with low viscosities. J Membr Sci. 2016;508:113–26.
Borwankar AU, Dear BJ, Twu A, Hung JJ, Dinin AK, Wilson BK, et al. Viscosity reduction of a concentrated monoclonal antibody with arginine·HCl and arginine·glutamate. Ind Eng Chem Res. 2016;55(43):11225–34.
Dear BJ, Hung JJ, Truskett TM, Johnston KP. Contrasting the influence of cationic amino acids on the viscosity and stability of a highly concentrated monoclonal antibody. Pharm Res. 2017;34(1):193–207.
Zhao H, Brown Patrick H, Schuck P. On the distribution of protein refractive index increments. Biophys J. 2011;100(9):2309–17.
Liu J, Nguyen MDH, Andya JD, Shire SJ. Reversible self-association increases the viscosity of a concentrated monoclonal antibody in aqueous solution. J Pharm Sci. 2005;94(9):1928–40.
Heimenz PC, Rajagopalan R. Principles of colloid and surface chemistry. 3rd ed. New York: Marcel Dekker, Inc; 1997.
Dharmaraj VL, Godfrin PD, Liu Y, Hudson SD. Rheology of clustering protein solutions. Biomicrofluidics. 2016;10(4):043509.
Wang S, Zhang N, Hu T, Dai W, Feng X, Zhang X, et al. Viscosity-lowering effect of amino acids and salts on highly concentrated solutions of two IgG1 monoclonal antibodies. Mol Pharm. 2015;12(12):4478–87.
Whitaker N, Xiong J, Pace SE, Kumar V, Middaugh CR, Joshi SB, et al. A formulation development approach to identify and select stable ultra–high-concentration monoclonal antibody formulations with reduced viscosities. J Pharm Sci. 2017;106(11):3230–41.
Auton M, Bolen DW, Rosgen J. Structural thermodynamics of protein preferential solvation: osmolyte solvation of proteins, aminoacids, and peptides. Proteins: Struct, Funct, Bioinf. 2008;73(4):802–13.
Zondlo NJ. Aromatic-proline interactions: electronically tunable CH/pi interactions. Acc Chem Res. 2013;46(4):1039–49.
Nicoud L, Jagielski J, Pfister D, Lazzari S, Massant J, Lattuada M, et al. Kinetics of Monoclonal Antibody Aggregation from Dilute toward Concentrated Conditions. J Phys Chem B. 2016.
Svergun DI, Koch MH, Timmins PA, May RP. Small angle X-ray and neutron scattering from solutions of biological macromolecules. New York: Oxford University Press; 2013. p. 368.
Woldeyes MA, Calero-Rubio C, Furst EM, Roberts CJ. Predicting protein interactions of concentrated globular protein solutions using colloidal models. J Phys Chem B. 2017;121(18):4756–67.
Ghosh R, Calero-Rubio C, Saluja A, Roberts CJ. Relating protein-protein interactions and aggregation rates from low to high concentrations. J Pharm Sci. 2016;105(3):1086–96.
Calero-Rubio C, Ghosh R, Saluja A, Roberts CJ. Predicting protein-protein interactions of concentrated antibody solutions using dilute solution data and coarse-grained molecular models. J Pharm Sci. 2017;
Godfrin PD, Zarzar J, Zarraga IE, Porcar L, Falus P, Wagner NJ, et al. The effect of hierarchical cluster formation on the viscosity of concentrated monoclonal antibody formulations studied by neutron scattering. J Phys Chem B. 2015.
Lilyestrom WG, Shire SJ, Scherer TM. Influence of the cosolute environment on igg solution structure analyzed by small-angle X-ray scattering. J Phys Chem B. 2012;116(32):9611–8.
Scherer TM. Role of cosolute–protein interactions in the dissociation of monoclonal antibody clusters. J Phys Chem B. 2015;119(41):13027–38.
Shukla D, Trout BL. Interaction of arginine with proteins and the mechanism by which it inhibits aggregation. J Phys Chem B. 2010;114(42):13426–38.
Haque I, Singh R, Moosavi-Movahedi AA, Ahmad F. Effect of polyol osmolytes on ΔGD, the Gibbs energy of stabilisation of proteins at different pH values. Biophys Chem. 2005;117(1):1–12.
Kuhn LA, Swanson CA, Pique ME, Tainer JA, Getzoff ED. Atomic and residue hydrophilicity in the context of folded protein structures. Proteins: Struct, Funct, Bioinf. 1995;23(4):536–47.
Pemberton TA, Still BR, Christensen EM, Singh H, Srivastava D, Tanner JJ. Proline: mother nature’s cryoprotectant applied to protein crystallography. Acta Crystallogr Sect D: Biol Crystallogr. 2012;68(Pt 8):1010–8.
Jiang SY, Cao ZQ. Ultralow-fouling, functionalizable, and hydrolyzable zwitterionic materials and their derivatives for biological applications. Adv Mater. 2010;22(9):920–32.
Wang S, Zhang N, Hu T, Dai WG, Feng X, Zhang X, et al. Viscosity-Lowering Effect of Amino Acids and Salts on Highly Concentrated Solutions of two IgG1 Monoclonal Antibodies. Mol Pharmaceutics. 2015.
Saha D, Joshi YM, Bandyopadhyay R. Characteristics of the secondary relaxation process in soft colloidal suspensions. Epl. 2015;112(4)
Richter S, Boyko V, Schröter K. Gelation studies on a radical chain cross-linking copolymerization process: comparison of the critical exponents obtained by dynamic light scattering and rheology. Macromol Rapid Commun. 2004;25(4):542–6.
Acknowledgments and Disclosures
AbbVie provided financial support and the antibody used in this study. The University of Texas at Austin received research funds from AbbVie Inc. to conduct the study. This work was also supported by the Welch Foundation (F-1319, F-1696) and National Science Foundation (CBET-1247945, DGE-1110007). Any opinion, findings, and conclusions or recommendations expressed in this material are those of the authors(s) and do not necessarily reflect the views of the National Science Foundation.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
ESM 1
(DOCX 176 kb)
Rights and permissions
About this article

Cite this article
Hung, J.J., Dear, B.J., Dinin, A.K. et al. Improving Viscosity and Stability of a Highly Concentrated Monoclonal Antibody Solution with Concentrated Proline. Pharm Res 35, 133 (2018). https://doi.org/10.1007/s11095-018-2398-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11095-018-2398-1